

‘140R’ Rootstock Regulates Resveratrol Content in ‘Cabernet Sauvignon’ Grapevine Leaves Through miRNA

Abstract
1. Introduction
2. Results
2.1. Aboveground Morphology of Grapevine Plants
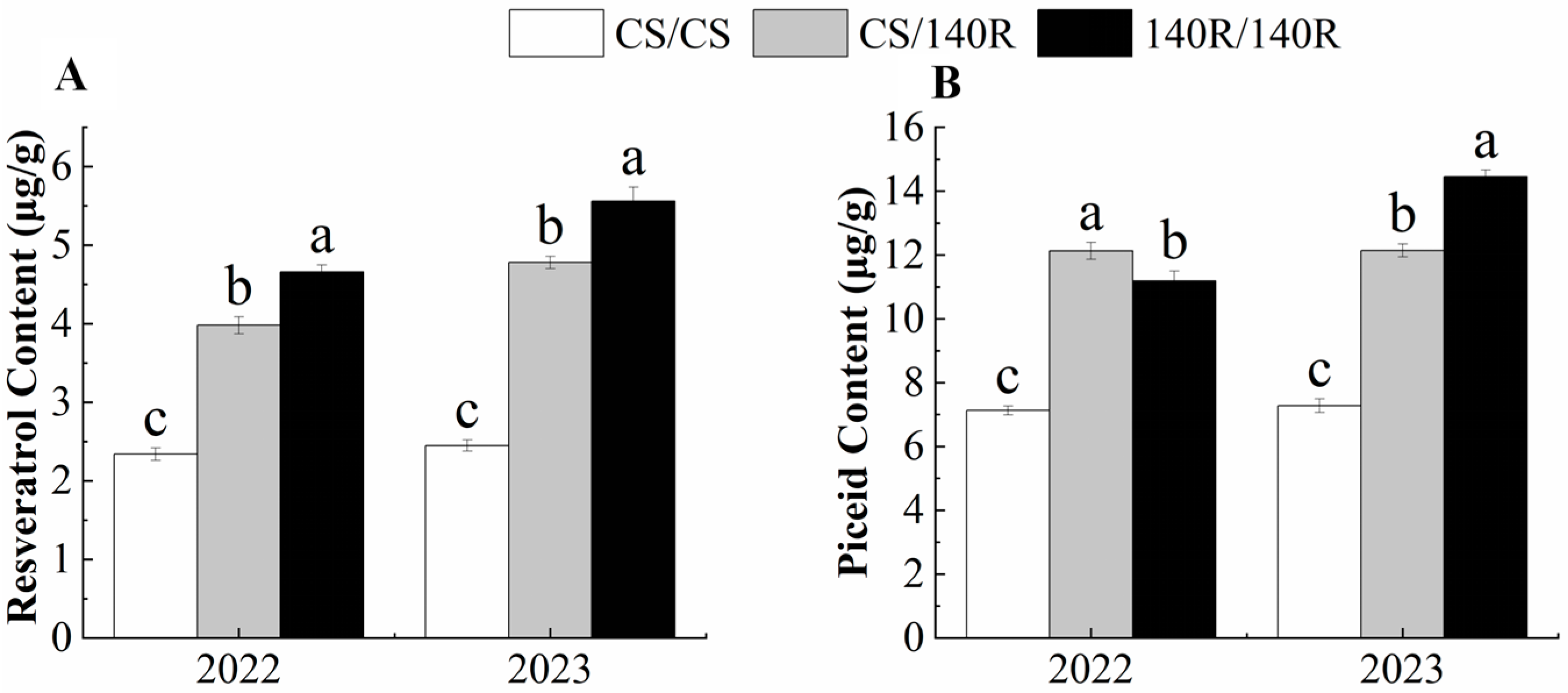
2.2. Resveratrol and Piceid Content of Grape Leaves
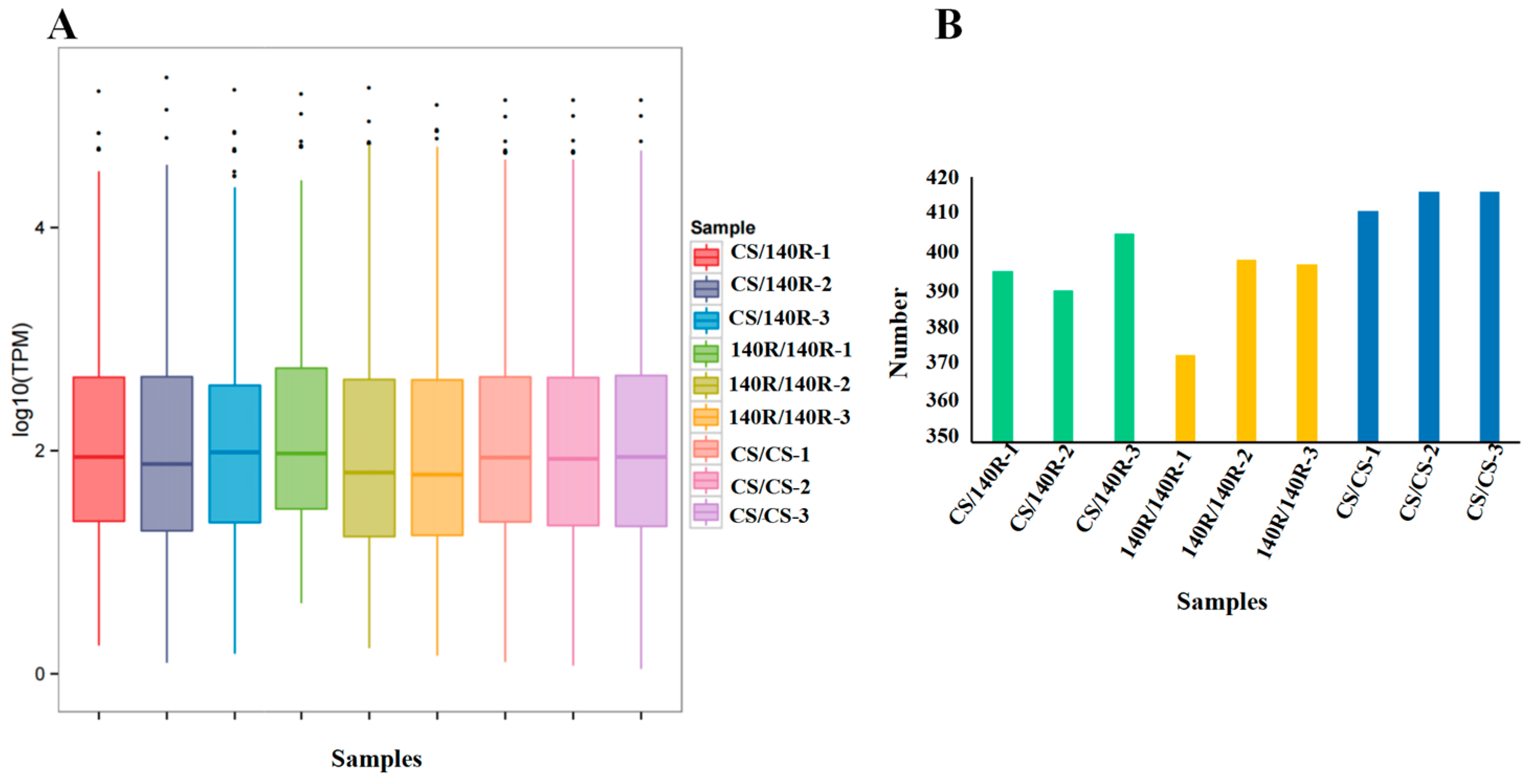
2.3. Sequencing and Analysis of sRNA Libraries
2.4. Identification of Grapevine miRNAs
2.5. Quantitative Analysis of Grapevine miRNAs
2.6. Differential miRNA Analysis
2.7. miRNA Target Gene Analysis and Functional Annotations of the Differentially Expressed miRNAs
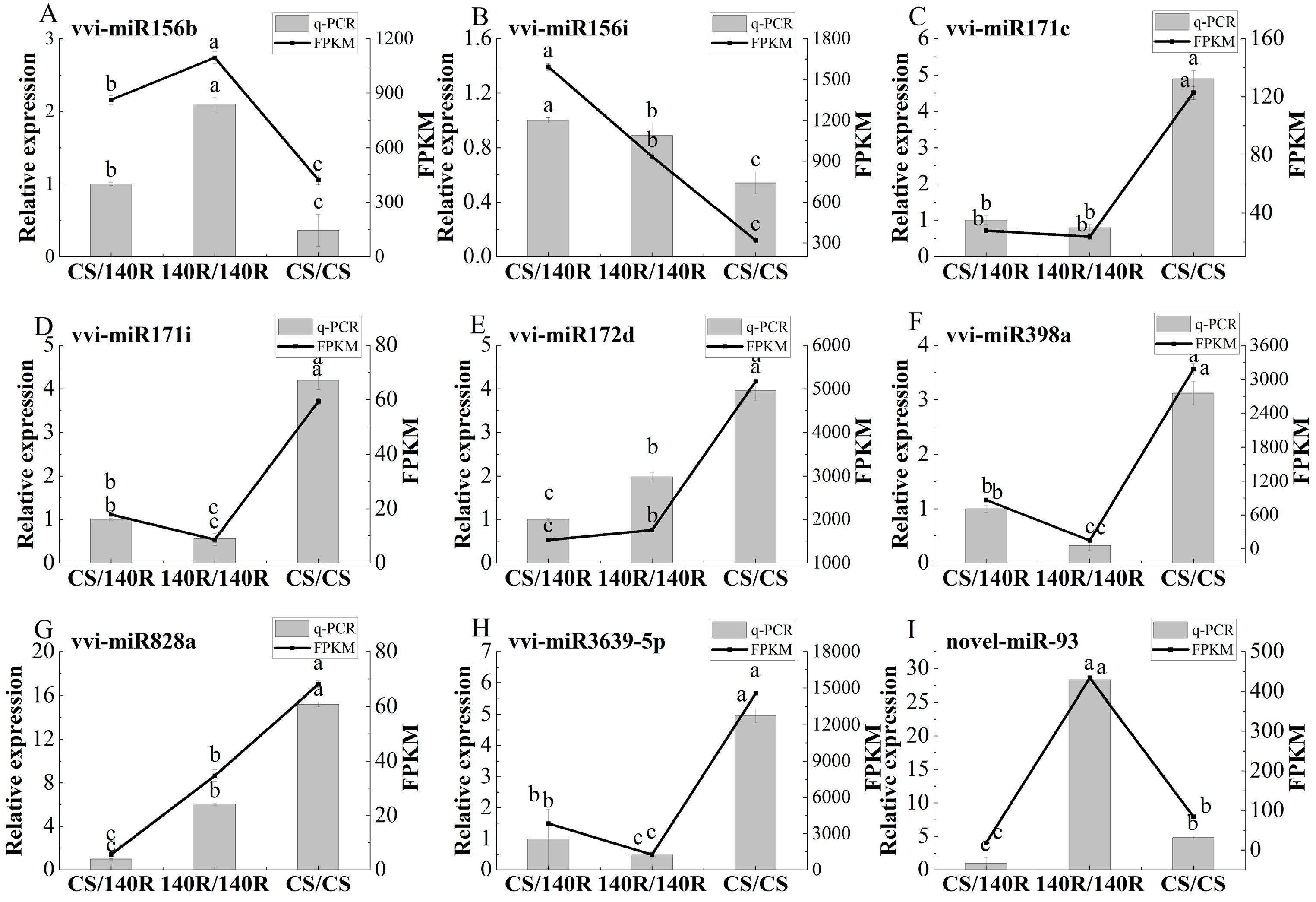
2.8. RT-qPCR Verification of Differentially Expressed miRNAs
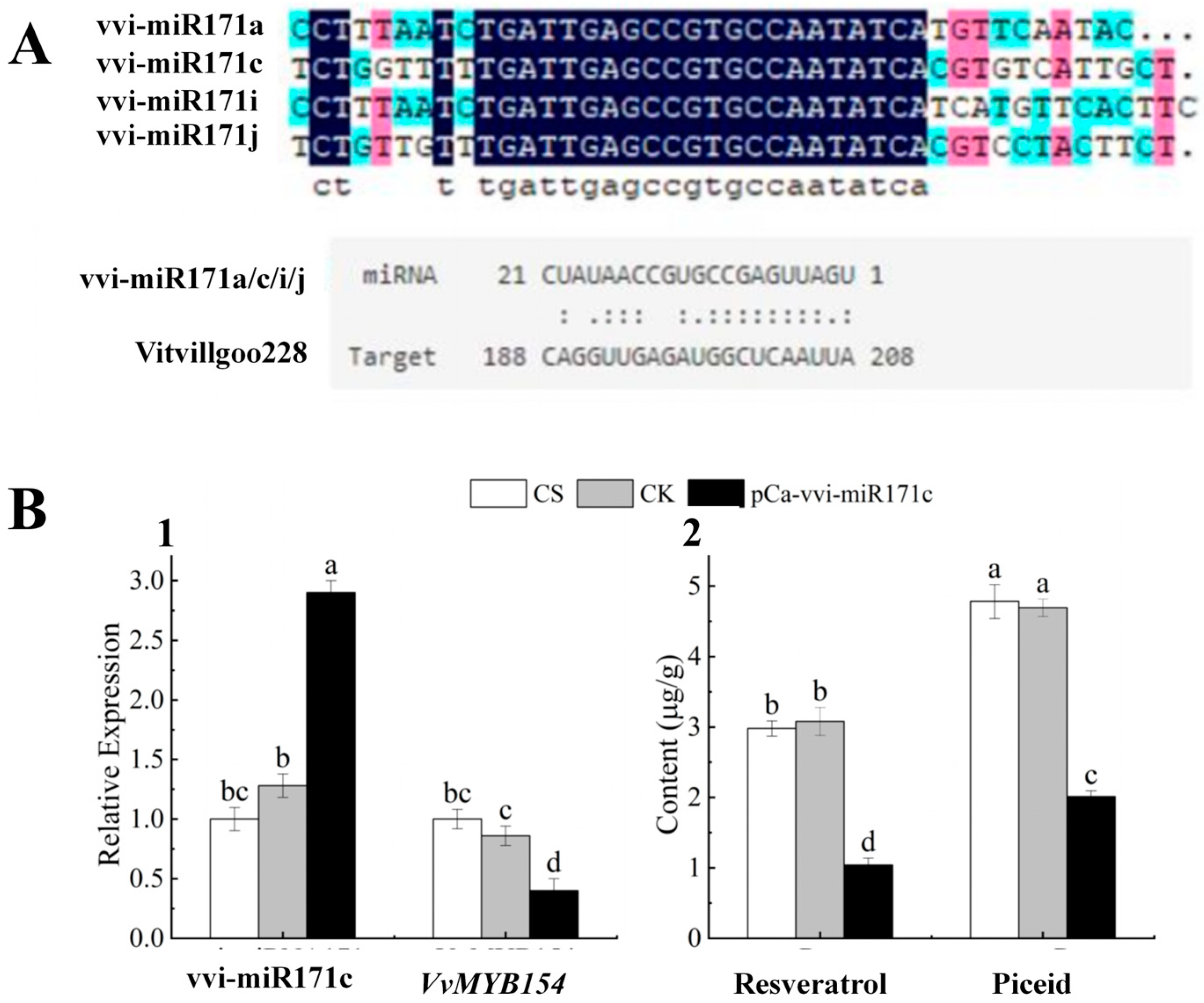
2.9. Functional Characterisation of vvi-miR171c
3. Discussion
4. Materials and Methods
4.1. Experimental Site and Material Culture
4.2. Determination of the Res and Piceid Content
4.3. Small RNA Library Construction and Sequencing Analysis
4.3.1. RNA Extraction and sRNA Library Construction
4.3.2. sRNA Annotation and miRNA Identification
4.3.3. Analysis of Differentially Expressed miRNAs
4.3.4. miRNA Target Gene Prediction and Functional Annotation
4.3.5. RT-qPCR Verification of Differentially Expressed miRNAs and Their Target Genes
4.3.6. Plasmid Construction and Generation of Transgenic Plants
4.3.7. Transient Transformation of vvi-miR171c in Grapevine
4.3.8. Statistical Analyses
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Myles, S.; Boylo, A.R.; Owens, C.L.; Brown, P.J.; Grassi, F.; Aradhya, M.K.; Prins, B.; Reynolds, A.; Chia, J.M.; Ware, D.; et al. Genetic structure and domestication history of the grape. Proc. Natl. Acad. Sci. USA 2011, 108, 3530–3535. [Google Scholar] [CrossRef]
- Guo, J.; Liu, C.; Pan, X.; Wang, J. Progress in the study of resveratrol in Vitis vinifera. J. Fruit Sci. 2002, 19, 199–204. (In Chinese) [Google Scholar]
- Li, A.; Wang, X.; Liu, D.; Zhao, M.; Qian, Y.; Fang, J. Changes in resveratrol content in various organs during the growth and development of four table grape varieties. J. Fruit Sci. 2014, 31, 1079–1085. [Google Scholar] [CrossRef]
- Li, T.; Li, S.; Zhang, Q.; Zhang, Z.; Liu, Y.; Wu, Y. Comparison of resveratrol content in different tissue parts of grapes. J. Gansu Agric. Univ. 2009, 44, 64–67. (In Chinese) [Google Scholar]
- Chen, L.; Han, Y. Differences in resveratrol content in different grape varieties and tissues. Acta Hortic. Sin. 1999, 26, 118–119. (In Chinese) [Google Scholar] [CrossRef]
- Dercks, W.; Creasy, L.L. The significance of stilbene phytoalexins in the Plasmopara viticola-grapevine interaction. Physiol. Mol. Plant Pathol. 1989, 34, 189–202. [Google Scholar] [CrossRef]
- Pezet, R.; Gindork Viret, O.; Spring, J.L. Glycosylation and oxidative dimerization of resveratrol are respectively associated to sensitivity and resistance of grapevine cultivars to downy mildew. Physiol. Mol. Plant Phathol. 2004, 65, 297–303. [Google Scholar] [CrossRef]
- Lepak, A.; Gutmann, A.; Kulmer, S.T.; Nidetzky, B. Creating a water-soluble Resveratrol-based antioxidant by site-selective enzymatic glucosylation. Chembiochem 2015, 16, 1870–1874. [Google Scholar] [CrossRef]
- Regev-Shoshani, G.; Shoseyov, O.; Bilkis, I.; Kerem, Z. Glycosylation of resveratrol protects it from enzymic oxidation. Biochem. J. 2003, 374, 157–163. [Google Scholar] [CrossRef]
- Yan, C.; Yang, N.; Wang, X.; Wang, Y. VqBGH40a isolated from Chinese wild Vitis quinquangularis degrades trans-piceid and enhances trans-resveratrol. Plant Sci. 2021, 310, 110989. [Google Scholar] [CrossRef]
- Zhang, F.; Song, X.; Zhong, H.; Zhou, X.; Han, S.; Zhang, W.; Pan, M. Study on the effects of Rootstocks on leaf quality and photosynthetic efficiency of Cabernet Sauvignon 9 grape. Xinjiang Agric. Sci. 2017, 54, 1223–1231. (In Chinese) [Google Scholar]
- Li, C.; Bai, S.; Geng, G.; Chen, G.; Zhao, R.; Cai, J. Effects of different rootstocks on the growth and development of ‘Cabernet Pearl’ grapes. J. Fruit Sci. 2016, 33, 1241–1250. [Google Scholar]
- Zhai, C.; Zhao, B.; Sun, J.; Yu, S.; Zhang, E.; Li, G.; Dong, X.; Zhang, Z. Effect of resistant rootstock on the process and content of resveratrol in ‘Cabernet Sauvignon’ grape leaves. Northwest Bot. J. 2017, 37, 1773–1780. (In Chinese) [Google Scholar]
- He, W.; Sun, J.; Zhao, B.; Zhang, B.; Liu, L.; Pan, L.; Chang, X. Effects of different rootstocks on resveratrol content and its related enzyme activities in ‘Cabernet Sauvignon’ grape berries. J. Fruit Sci. 2019, 36, 738–747. (In Chinese) [Google Scholar] [CrossRef]
- Tsaballa, A.; Athanasiadis, C.; Pasentsis, K.; Ganopoulos, I.; Nianiou-Obeidat, I.; Tsaftaris, A. Molecular studies of inheritable grafting induced changes in pepper (Capsicum annuum) fruit shape. Sci. Hortic. 2013, 149, 2–8. [Google Scholar] [CrossRef]
- Deng, Z.; Wu, H.; Li, D.; Li, L.; Wang, Z.; Yuan, W.; Xing, Y.; Li, C.; Liang, D. Root-to-shoot long-distance mobile. miRNAs identified from Nicotiana rootstocks. Int. J. Mol. Sci. 2022, 22, 12821. [Google Scholar] [CrossRef]
- Baron, D.; Amaro, A.C.E.; Pina, A.; Ferreira, G. An overview of grafting re-establishment in woody fruit species. Sci. Hortic. Amst. 2019, 243, 84–91. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Reinhart, B.J.; Weinstein, E.G.; Rhoades, M.W.; Bartel, B.; Bartel, D.P. MicroRNAs in plants. Genes Dev. 2002, 16, 1616–1626. [Google Scholar] [CrossRef]
- Zhang, B. MicroRNA: A new target for improving plant tolerance to abiotic stress. J. Exp. Bot. 2015, 66, 1749–1761. [Google Scholar] [CrossRef]
- Jiang, X. Molecular Mechanism of the Effect of Different Rootstocks on the Development of “Yanfu 3” Apple Scion. Master’s Thesis, Yantai University, Yantai, China, 2019. [Google Scholar]
- Xu, Y.; Zhu, S.; Liu, N.; Li, Q.; Zhao, X. Effect of grafting on microRNA expression in citrus. Acta Hortic. Sinica. 2017, 44, 1263–1274. [Google Scholar] [CrossRef]
- Wang, C. Identification, and Mode of Action of Grape microRNAs and Their Target Genes. Master’s Thesis, Nanjing Agricultural University, Nanjing, China, 2012. [Google Scholar]
- Sun, X. Identification and Interaction of Grapevine microRNA164 and microRNA171 and Their Targets. Master’s Thesis, Nanjing Agricultural University, Nanjing, China, 2015. [Google Scholar]
- Jiang, C. Mechanism of VqMYB154 Transcription Factor Regulation of Powdery Mildew Resistance in Chinese Wild Grape. Master’s Thesis, Northwest A&F University, Yangling, China, 2021. [Google Scholar]
- Wang, D. Expression of Transcription Factors WRKY and bZIP in Wild Grape in China and Their Regulation of Disease Resistance. Master’s Thesis, Northwest A&F University, Yangling, China, 2019. [Google Scholar]
- Zuo, J.; Wang, Q.; Han, C.; Ju, Z.; Cao, D.; Zhu, B.; Luo, Y.; Gao, L. SRNAome and degradome sequencing analysis reveals specific regulation of sRNA in response to chilling injury in tomato fruit. Physiol. Plant. 2017, 160, 142–154. [Google Scholar] [CrossRef] [PubMed]
- Dey, S.; Sarkar, A.; Chowdhury, S.; Singh, R.; Mukherjee, A.; Ghosh, Z.; Kundu, P. Heightened miR6024-NLR interactions facilitate necrotrophic pathogenesis in tomato. Plant. Mol. Biol. 2022, 109, 717–739. [Google Scholar] [CrossRef] [PubMed]
- Mu, H.; Li, Y.; Yuan, L.; Jiang, J.; Wei, Y.; Duan, W.; Fan, P.; Li, S.; Liang, Z.; Wang, L. MYB30 and MYB14 form a repressor-activator module with WRKY8 that controls stilbene biosynthesis in grapevine. Plant Cell 2023, 35, 552–573. [Google Scholar] [CrossRef]
- Chitwood, D.H.; Nogueira, F.; Howell, M.; Montgomery, T.A.; Carrington, J.C.; Timmermans, M.C. Pattern formation via small RNA mobility. Genes Dev. 2009, 23, 549–554. [Google Scholar] [CrossRef]
- Yoo, B.C.; Kragler, F.; Varkonyi-Gasic, E.; Haywood, V.; Archer-Evans, S.; Lee, Y.M.; Lough, T.J.; Lucas, W.J. A systemic small RNA signaling system in plants. Plant Cell 2004, 16, 1979–2000. [Google Scholar] [CrossRef]
- Varkonyi-Gasic, E.; Gould, N.; Sandanayaka, M.; Sutherland, P.; MacDiarmid, R.M. Characterisation of microRNAs from apple (Malus domestica ‘Royal Gala’) vascular tissue and phloem sap. BMC Plant Biol. 2010, 10, 159. [Google Scholar] [CrossRef]
- Jones-Rhoades, M.W.; Bartel, D.P.; Bartel, B. MicroRNAS and their regulatory roles in plants. Annu. Rev. Plant. Biol. Biol. 2006, 57, 19–53. [Google Scholar] [CrossRef]
- Khraiwesh, B.; Zhu, J.K.; Zhu, J. Role of miRNAs and siRNAs in biotic and abiotic stress responses of plants. Biochim. Biophys. Acta 2012, 1819, 137–148. [Google Scholar] [CrossRef]
- Ferdous, J.; Sanchez-Ferrero, J.C.; Langridge, P. Differential expression of microRNAs and potential targets under drought stress in barley. Plant Cell Environ. 2017, 40, 11–24. [Google Scholar] [CrossRef]
- An, N.; Fan, S.; Yang, Y.; Chen, X.; Dong, F.; Wang, Y.; Xing, L.; Zhao, C.; Han, M. Identification and Characterization of miRNAs in Self-Rooted and Grafted Malus Reveals Critical Networks Associated with Flowering. Int. J. Mol. Sci. 2018, 19, 2384. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Wang, L.; Ma, L.; Xi, H.; Duan, W.; Wang, J.; Li, S. Individual and combined effects of CaCl2 and UV–C on the biosynthesis of resveratrols in Grape Leaves and Berry Skins. J. Agric. Food Chem. 2013, 61, 7135–7141. [Google Scholar] [CrossRef] [PubMed]
- Jiu, S.; Zhu, X.; Wang, J. Genome-Wide mapping and analysis of grapevine microRNAs and their potential target genes. Plant Genome 2015, 8, plantgenome2014.12.0091. [Google Scholar] [CrossRef] [PubMed]
- Hu, Z.; Liu, A.; Gitau, M.; Huang, X.; Chen, L.; Fu, J. Insights into the MicroRNA-regulated response of bermudagrass to cold and salt stress. Environ. Exp. Bot. 2018, 145, 64–74. [Google Scholar] [CrossRef]
- Jiang, J.; Lv, M.; Liang, Y.; Ma, Z.; Cao, J. Identification of novel and conserved miRNAs involved in pollen development in Brassica campestris ssp. chinensis by high-throughput sequencing and degradome analysis. BMC Genom. 2014, 15, 146. [Google Scholar] [CrossRef]
- Ma, C.; Lu, Y.; Bai, S.; Zhang, W.; Duan, X.; Meng, D.; Wang, Z.; Wang, A.; Zhou, Z.; Li, T. Cloning and characterization of miRNAs and their targets, including a novel miRNA targeted NBS–LRR protein class gene in apple (Golden Delicious). Mol. Plant 2014, 7, 218–230. [Google Scholar] [CrossRef]
- Saminathan, T.; Bodunrin, A.; Singh, N.V.; Devarajan, R.; Nimmakayala, P.; Jeff, M. Reddy Genome-wide identification of microRNAs in pomegranate (Punica granatum L.) by high-throughput sequencing. BMC Plant Biol. 2016, 16, 100289. [Google Scholar] [CrossRef]
- Zhang, L.; Fan, D.; Li, H.; Chen, Q.; Zhang, Z.; Liu, M.; Liu, J.; Song, Y.; He, J.; Xu, W.; et al. Characterization and identification of grapevine heat stress-responsive microRNAs revealed the positive regulated function of vvi-miR167 in thermostability. Plant Sci. 2023, 329, 111623. [Google Scholar] [CrossRef]
- Zhang, B.; Pan, X.; Wang, Q.; Cob, G.P.; Anderson, T.A. Identification and characterization of new plant microRNAs using EST analysis. Cell. Res. 2005, 15, 1074–1075. [Google Scholar] [CrossRef]
- Rajagopalan, R.; Vaucheret, H.; Trejo, J. A diverse and evolutionarily fluid set of microRNAs in Arabidopsis thaliana. Genes Dev. 2006, 20, 3407–3425. [Google Scholar] [CrossRef]
- Lin, S.; Santi, C.; Jobet, E.; Lacut, E.; El, K.N.; Karlowski, W.M.; Verdeil, J.L.; Breitler, J.C.; Périn, C.; Ko, S.S.; et al. Complex regulation of two target genes encoding SPX-MFS proteins by rice miR827 in response to phosphate starvation. Plant Cell Physiol. 2010, 51, 2119–2131. [Google Scholar] [CrossRef] [PubMed]
- Wei, L.; Du, Y.; Xiang, J.; Zheng, T.; Cheng, J.; Wu, J. Integrated mRNA and miRNA transcriptome analysis of grape in responses to salt stress. Front Plant Sci. 2023, 14, 1173857. [Google Scholar] [CrossRef] [PubMed]
- Gandikota, M.; Birkenbihl, R.P.; Höhmann, S.; Cardon, G.H.; Saedler, H.; Huijser, P. The mirna156/157 recognition element in the 3′ utr of the Arabidopsis sbp box gene spl3 prevents early flowering by translational inhibition in seedlings. Plant J. 2007, 49, 683–693. [Google Scholar] [CrossRef] [PubMed]
- Ma, Z.; Hu, X.; Cai, W.; Huang, W.; Zhou, X.; Luo, Q.; Yang, H.; Wang, J.; Huang, J. Arabidopsis miR171-targeted scarecrow-like proteins bind to GT cis-elements and mediate gibberellin-regulated chlorophyll biosynthesis under light conditions. PLoS Genet. 2014, 10, e1004519. [Google Scholar] [CrossRef]
- Wang, L.; Mai, Y.; Zhang, Y.; Luo, Q.; Yang, H. Microrna171c-targeted scl6-ii, scl6-iii, and scl6-iv genes regulate shoot branching in Arabidopsis. Mol. Plant. 2010, 3, 794–806. [Google Scholar] [CrossRef]
- He, W. Effects of Different Rootstocks on Resveratrol Synthesis and Related Gene Expression Levels in “Cabernet Sauvignon” Grapes. Master’s Thesis, Shihezi University, Shihezi, China, 2020. [Google Scholar]
- Zhu, F.; Chen, M.; Ye, N.; Qiao, W.; Gao, B.; Liu, Y. Comparative performance of the BGISEQ-500 and Illumina HiSeq4000 sequencing platforms for transcriptome analysis in plants. Plant Methods 2018, 14, 69. [Google Scholar] [CrossRef]
- Langmead, B.; Trapnell, C.; Pop, M.; Salzberg, S.L. Ultrafast and memory-efficient alignment of short DNA sequences to the human genome. Genome Biol. 2009, 10, R25. [Google Scholar] [CrossRef]
- Shi, X.; Cao, S.; Wang, X.; Huang, S.; Wang, Y.; Liu, Z.; Liu, W.; Leng, X.; Peng, Y.; Wang, N.; et al. The complete reference genome for grapevine (Vitis vinifera L.) genetics and breeding. Hortic. Res. 2023, 10, uhad061. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Friedlander, M.; Mackowiak, S.; Li, N.; Chen, W.; Rajewsky, N. miRDeep2 accurately identifies known and hundreds of novel microRNA genes in seven animal clades. Nucleic Acids Res. 2012, 40, 37–52. [Google Scholar] [CrossRef]
- Zhang, Z.; Jiang, L.; Wang, J. MTide: An integrated tool for the identification of miRNA-target interaction in plants. Bioinformatics 2014, 31, 290–291. [Google Scholar] [CrossRef]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef] [PubMed]
- Robinson, M.D.; Mccarthy, D.J.; Smyth, G.K. edgeR: A Bioconductor package for differential expression analysis of digital gene expression data. Bioinformatics 2009, 26, 139–140. [Google Scholar] [CrossRef] [PubMed]
- Allen, E.; Xie, Z.; Gustafson, A.M.; Carrington, J.C. microRNA-directed phasing during trans-acting siRNA biogenesis in plants. Cell 2005, 121, 207–221. [Google Scholar] [CrossRef] [PubMed]
- Deng, Y.; Li, J.; Wu, S.; Zhu, Y.; Chen, Y.; He, F. Integrated nr database in protein annotation system and its localization. Comput. Eng. 2006, 32, 71–74. [Google Scholar] [CrossRef]
- Ashburner, M.; Ball, C.A.; Blake, J.A.; Botstein, D.; Butler, H.; Cherry, J.M.; Davis, A.P.; Dolinski, K.; Dwight, S.S.; Eppig, J.T.; et al. Gene ontology: Tool for the unification of biology. The Gene Ontology Consortium. Nat. Genet. 2000, 25, 25–29. [Google Scholar] [CrossRef]
- Roman, L.T.; Michael, Y.G.; Darren, A.N. The COG database: A tool for genome scale analysis of protein functions and evolution. Nucleic Acids Res. 2007, 28, 33–36. [Google Scholar] [CrossRef]
- Minoru, K.; Susumu, G.; Shuichi, K. The KEGG resource for deciphering the genome. Nucleic Acids Res. 2004, 32, D277–D280. [Google Scholar] [CrossRef]
- Koonin, E.V.; Fedorova, N.D.; Jackson, J.D. A comprehensive evolutionary classification of proteins encoded in complete eukaryotic genomes. Genome Biol. 2004, 5, R7. [Google Scholar] [CrossRef]
- Liave, C.; Kasschau, K.D.; Rector, M.A.; Carrington, J.C. Endogenous and silencing-associated small RNAs in plants. Plant Cell 2002, 14, 1605–1619. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| miRNA Name | log2(CS/CS vs. CS/140R) | miRNA Name | log2FC(CS/CS vs. 140R/140R) |
|---|---|---|---|
| vvi-miR171c | −2.055713777 | novel_miR_110 | −3.638579789 |
| vvi-miR169c | −3.99381221 | novel_miR_175 | −2.462009742 |
| novel_miR_166 | −3.908111265 | vvi-miR828a | −1.703482327 |
| vvi-miR828a | −3.383976384 | novel_miR_177 | −1.702342234 |
| novel_miR_128 | −3.043316763 | vvi-miR169l | −1.555720976 |
| novel_miR_2 | −3.043316763 | novel_miR_1 | 9.725505306 |
| novel_miR_263 | 2.734070564 | vvi-miR398a | 4.280026387 |
| novel_miR_172 | 2.86081601 | novel_miR_183 | 3.475394554 |
| novel_miR_181 | 2.86081601 | vvi-miR479 | 2.744971769 |
| vvi-miR3627-5p | 3.168791739 | vvi-miR319c | 2.024953876 |
| vvi-miR3627-3p | 3.412211104 | novel_miR_52 | 2.03448677 |
| vvi-miR3624-5p | 3.465130458 | vvi-miR319f | 2.036424038 |
| vvi-miR3624-3p | 3.545753663 | vvi-miR399e | 2.199961988 |
| Compound | Retention Time | Standard Curves | Coefficient of Determination |
|---|---|---|---|
| trans-Res | 2.931 | y = 132,447x − 120,557 | 0.9993 |
| Trans-Pd | 4.328 | y = 103,081x − 74,534 | 0.9996 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhu, C.; Zhang, Z.; Liu, Z.; Shi, W.; Zhang, D.; Zhao, B.; Sun, J. ‘140R’ Rootstock Regulates Resveratrol Content in ‘Cabernet Sauvignon’ Grapevine Leaves Through miRNA. Plants 2024, 13, 3057. https://doi.org/10.3390/plants13213057
Zhu C, Zhang Z, Liu Z, Shi W, Zhang D, Zhao B, Sun J. ‘140R’ Rootstock Regulates Resveratrol Content in ‘Cabernet Sauvignon’ Grapevine Leaves Through miRNA. Plants. 2024; 13(21):3057. https://doi.org/10.3390/plants13213057
Chicago/Turabian StyleZhu, Chunmei, Zhijun Zhang, Zhiyu Liu, Wenchao Shi, Dongliang Zhang, Baolong Zhao, and Junli Sun. 2024. "‘140R’ Rootstock Regulates Resveratrol Content in ‘Cabernet Sauvignon’ Grapevine Leaves Through miRNA" Plants 13, no. 21: 3057. https://doi.org/10.3390/plants13213057
APA StyleZhu, C., Zhang, Z., Liu, Z., Shi, W., Zhang, D., Zhao, B., & Sun, J. (2024). ‘140R’ Rootstock Regulates Resveratrol Content in ‘Cabernet Sauvignon’ Grapevine Leaves Through miRNA. Plants, 13(21), 3057. https://doi.org/10.3390/plants13213057