
The Plant Volatile-Sensing Mechanism of Insects and Its Utilization


Abstract
1. Introduction
2. Plant Volatiles
3. Molecular Perception of PVOCs by Insects
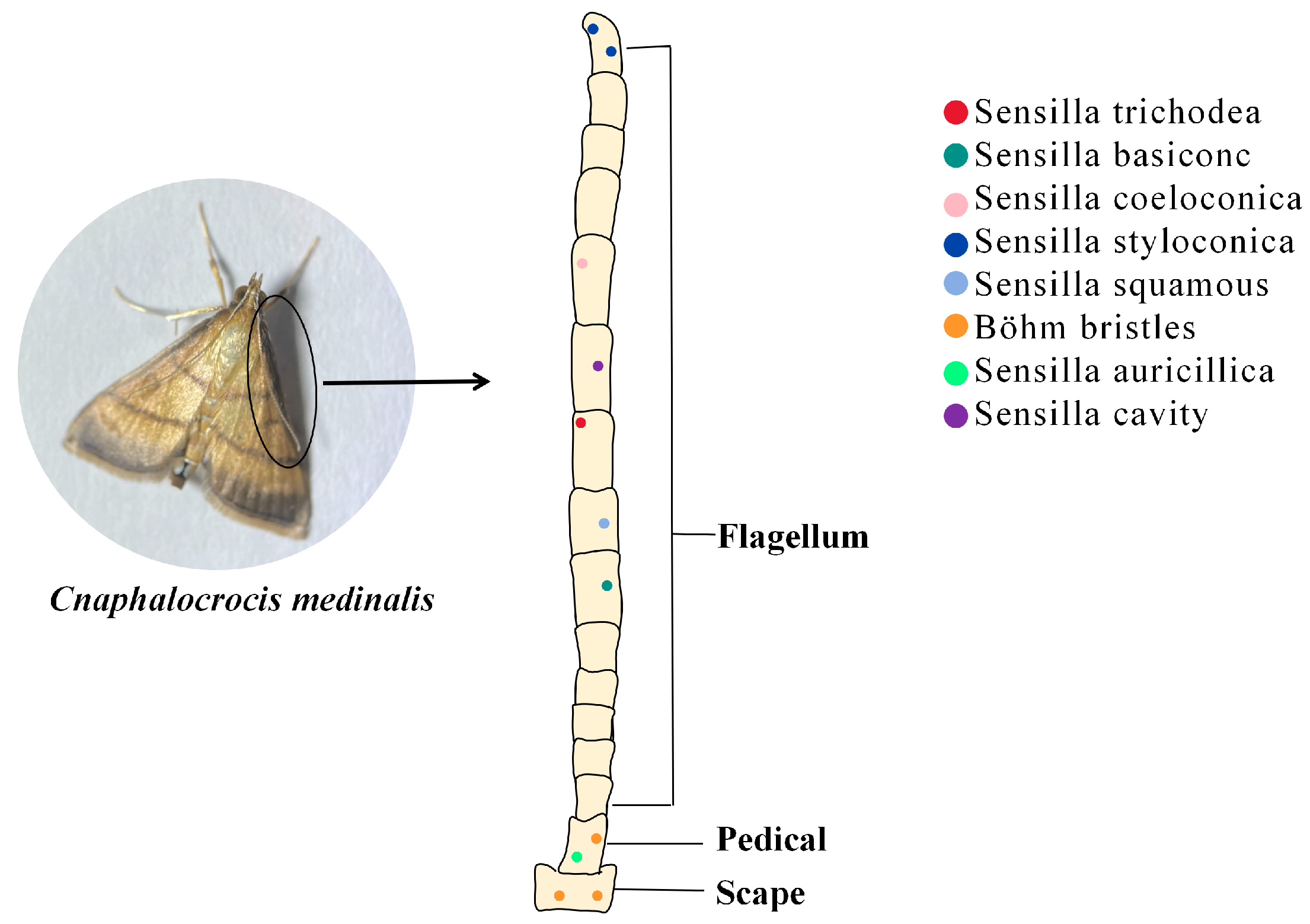
3.1. The Role of Insect Antennal Olfactory Sensors in the Recognition of PVOCs
3.2. The Role of Insect Antennal Olfaction-Related Proteins in the Recognition of PVOCs
4. The Attraction Mechanism of Plant Volatiles to Insects: A Case Study of C. medinalis
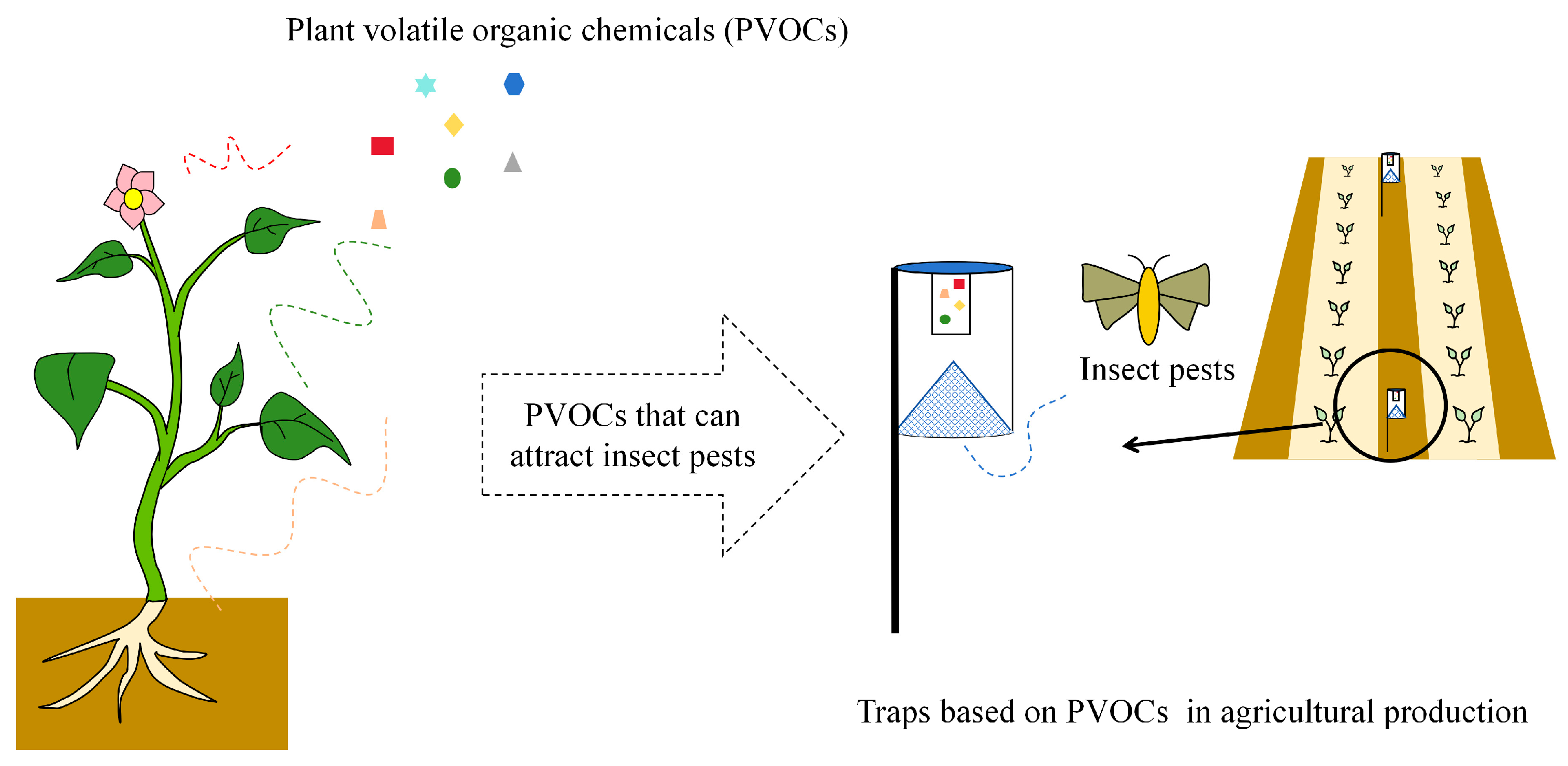
5. Development and Utilization of Pest Attractants Based on PVOCs
6. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Wu, J.; Baldwin, I.T. New insights into plant responses to the attack from insect herbivores. Annu. Rev. Genet. 2010, 44, 1–24. [Google Scholar] [CrossRef] [PubMed]
- Plettner, E. Insect pheromone olfaction: New targets for the design of species-selective pest control agents. Curr. Med. Chem. 2002, 9, 1075–1085. [Google Scholar] [CrossRef] [PubMed]
- Zu, P.; Zhang, D.Y.; Luo, Y.B. Chemical communication between plants and insects. J. Syst. Evol. 2023, 61, 441–444. [Google Scholar] [CrossRef]
- Fang, J.X.; Chen, D.F.; Shi, X.; Zhang, S.F.; Liu, F.; Shen, W.X.; Jia, C.Y.; Ma, S.C.; Zhang, Z.; Kong, X.B. Solid-phase microextraction and cuticular hydrocarbon differences related to reproductive activity in juniper bark borer Semanotus bifasciatus Motschulsky. J. Syst. Evol. 2023, 61, 498–505. [Google Scholar] [CrossRef]
- Rizvi, S.A.H.; George, J.; Reddy, G.V.P.; Zeng, X.; Guerrero, A. Latest developments in insect sex pheromone research and its application in agricultural pest management. Insects 2021, 12, 484. [Google Scholar] [CrossRef]
- Wyckhuys, K.A.G.; Lu, Y.; Zhou, W.; Cock, M.J.W.; Naranjo, S.E.; Fereti, A.; Williams, F.E.; Furlong, M.J. Ecological pest control fortifies agricultural growth in Asia–Pacific economies. Nat. Ecol. Evol. 2020, 4, 1522–1530. [Google Scholar] [CrossRef]
- Zhao, J.; Cai, W.L.; Shen, Y.Y.; Zhu, H.Y.; Pu, L.; Xie, M.Q.; Zou, Y.L.; Hua, H.X. Current situation and prospect of green rice pest control technology. J. Huazhong Agric. Univ. 2022, 41, 92–104. (In Chinese) [Google Scholar]
- Beck, J.J.; Torto, B.; Vannette, R.L. Eavesdropping on plant-insect-microbe chemical communications in agricultural ecology: A virtual issue on semiochemicals. J. Agric. Food Chem. 2017, 65, 5101–5103. [Google Scholar] [CrossRef]
- Liu, H.; Sun, X.; Shi, Z.; An, X.; Khashaveh, A.; Li, Y.; Gu, S.; Zhang, Y. Identification and functional analysis of odorant-binding proteins provide new control strategies for Apolygus lucorum. Int. J. Biol. Macromol. 2023, 224, 1129–1141. [Google Scholar] [CrossRef]
- Pelosi, P.; Iovinella, I.; Zhu, J.; Wang, G.; Dani, F.R. Beyond chemoreception: Diverse tasks of soluble olfactory proteins in insects. Biol. Rev. Camb. Philos. Soc. 2018, 93, 184–200. [Google Scholar] [CrossRef]
- Zeng, L.T.; Zhou, X.C.; Fu, X.M.; Hu, Y.L.; Gu, D.C.; Hou, X.L.; Dong, F.; Yang, Z.Y. Effect of the biosynthesis of the volatile compound phenylacetaldehyde on chloroplast modifications in tea (Camellia sinensis) plants. Hortic. Res. 2023, 10, uhad003. [Google Scholar] [CrossRef] [PubMed]
- Dudareva, N.; Klempien, A.; Muhlemann, J.K.; Kaplan, I. Biosynthesis, function and metabolic engineering of plant volatile organic compounds. New Phytol. 2013, 198, 16–32. [Google Scholar] [CrossRef] [PubMed]
- Laothawornkitkul, J.; Taylor, J.E.; Paul, N.D.; Hewitt, C.N. Biogenic volatile organic compounds in the Earth system. New Phytol. 2009, 183, 27–51. [Google Scholar] [CrossRef]
- Schiestl, F.; Ayasse, M. Post-pollination emission of a repellent compound in a sexually deceptive orchid: A new mechanism for maximising reproductive success? Oecologia 2001, 126, 531–534. [Google Scholar] [CrossRef] [PubMed]
- Majetic, C.J.; Raguso, R.A.; Ashman, T.L. Sources of floral scent variation: Can environment define floral scent phenotype? Plant Signal. Behav. 2009, 4, 129–131. [Google Scholar] [CrossRef] [PubMed]
- Martel, C.; Rakosy, D.; Romero, P.E.; Jersáková, J.; Ayasse, M. The evolution of tachinid pollination in Neotinea ustulata is related to floral cuticular composition and the combined high relative production of (Z)-11-C23/C25enes. J. Syst. Evol. 2021, 61, 487–497. [Google Scholar] [CrossRef]
- Guo, M.; Du, L.; Chen, Q.; Feng, Y.; Zhang, J.; Zhang, X.; Tian, K.; Cao, S.; Huang, T.; Jacquin-Joly, E.; et al. Odorant receptors for detecting flowering plant cues are functionally conserved across moths and butterflies. Mol. Biol. Evol. 2021, 38, 1413–1427. [Google Scholar] [CrossRef]
- Feng, H.H.; Wang, X.Y.; Luo, Y.B.; Huang, S.Q. Floral scent emission is the highest at the second night of anthesis in Lonicera japonica (Caprifoliaceae). J. Syst. Evol. 2023, 61, 530–537. [Google Scholar] [CrossRef]
- Brevik, K.; Schoville, S.D.; Mota-Sanchez, D.; Chen, Y.H. Pesticide durability and the evolution of resistance: A novel application of survival analysis. Pest Manag. Sci. 2018, 74, 1953–1963. [Google Scholar] [CrossRef]
- Yan, X.Z.; Ma, L.; Li, X.F.; Chang, L.; Liu, Q.Z.; Song, C.F.; Zhao, J.Y.; Qie, X.T.; Deng, C.P.; Wang, C.Z.; et al. Identification and evaluation of cruciferous plant volatiles attractive to Plutella xylostella L. (Lepidoptera: Plutellidae). Pest Manag. Sci. 2023, 79, 5270–5282, Accepted Author Manuscript. [Google Scholar] [CrossRef]
- Meagher, R.L.; Landolt, P.J. Binary floral lure attractive to velvetbean caterpillar adults (Lepidoptera: Noctuidae). Fla. Entomol. 2010, 93, 73–79. [Google Scholar] [CrossRef]
- Desurmont, G.A.; Arx, M.V.; Turlings, T.C.J.; Schiestl, F.P. Floral odors can interfere with the foraging behavior of parasitoids searching for hosts. Front. Ecol. Evol. 2020, 8, 148. [Google Scholar] [CrossRef]
- Wu, S.; Liu, F.; Zeng, W.; Xiao, Z.; Li, J.; Teng, K.; Guo, Q.; Zhao, J.; Du, Y. Evaluation of floral-derived volatile blend for attracting aphid parasitoids and lady beetles in the tobacco fields. Biol. Control 2022, 172, 104979. [Google Scholar] [CrossRef]
- Schneider, D. Insect antennae. Annu. Rev. Entomol. 1964, 9, 103–122. [Google Scholar] [CrossRef]
- Mclver, S.B. Structure of cuticular mechanoreceptors of arthropods. Annu. Rev. Entomol. 1975, 20, 381–397. [Google Scholar] [CrossRef]
- Roh, H.S.; Park, K.C.; Oh, H.W.; Park, C.G. Morphology and distribution of antennal sensilla of two tortricid moths, Cydia pomonella and C. succedana (Lepidoptera). Microsc. Res. Tech. 2016, 79, 1069–1081. [Google Scholar] [CrossRef]
- Zacharuk, R.Y. Antennae and sensilla. Compr. Insect Physiol. Biochem. Pharmacol. 1985, 17, 1283–1284. [Google Scholar]
- Pophof, B.; Stange, G.; Abrell, L. Volatile organic compounds as signals in a plant-herbivore system: Electrophysiological responses in olfactory sensilla of the moth Cactoblastis cactorum. Chem. Senses 2005, 30, 51–68. [Google Scholar] [CrossRef]
- Khallaf, A.; Knaden, M. Evolutionary neuroecology of olfactory-mediated sexual communication and host specialization in Drosophila—A review. Entomol. Exp. Appl. 2022, 170, 289–302. [Google Scholar] [CrossRef]
- Brilli, F.; Loreto, F.; Baccelli, I. Exploiting plant volatile organic compounds (VOCs) in agriculture to improve sustainable defense strategies and productivity of crops. Front. Plant Sci. 2019, 10, 264. [Google Scholar] [CrossRef]
- Kwon, H.W.; Lu, T.; Rützler, M.; Zwiebel, L.J. Olfactory responses in a gustatory organ of the malaria vector mosquito Anopheles gambiae. Proc. Natl. Acad. Sci. USA 2006, 103, 13526–13531. [Google Scholar] [CrossRef] [PubMed]
- Sun, X.; Wang, M.Q.; Zhang, G.A. Ultrastructural observations on antennal sensilla of Cnaphalocrocis medinalis (Lepidoptera: Pyralidae). Microsc. Res. Tech. 2011, 74, 113–121. [Google Scholar] [CrossRef] [PubMed]
- Piersanti, S.; Rebora, M. The antennae of damselfly larvae. Arthropod Struct. Dev. 2018, 47, 36–44. [Google Scholar] [CrossRef] [PubMed]
- Ma, R.Y.; Du, J.W. Antennal sensilla of insect. Chin. J. Appl. Entomol. 2000, 37, 179–183. (In Chinese) [Google Scholar]
- Li, C.D.; Yang, D.R.; Shen, F.R.; Yang, Y.X. Scanning electron microscopic observation on antennal sensilla of Hepialus yulongcnsis. Zool. Res. 1994, 11, 83–86. (In Chinese) [Google Scholar]
- Liébanas, G.; Sáez, Á.; Luna, Á.; Romero-Vidal, P.; Palma, A.; Pérez, J.M. The morphology of Colpocephalum pectinatum (Phthiraptera: Amblycera: Menoponidae) under scanning electron microscopy. Arthropod Struct. Dev. 2021, 64, 101085. [Google Scholar] [CrossRef]
- Dong, W.Y.; Wang, B.; Wang, G.R. Morphological and ultrastructural characterization of antennal sensilla and the detection of floral scent volatiles in Eupeodes corollae (Diptera: Syrphidae). Front. Neuroanat. 2021, 15, 791900. [Google Scholar] [CrossRef]
- Chen, Q.; Liu, X.; Cao, S.; Ma, B.; Guo, M.; Shen, J.; Wang, G. Fine structure and olfactory reception of the labial palps of Spodoptera frugiperda. Front. Physiol. 2021, 12, 680697. [Google Scholar] [CrossRef]
- Liu, Y.Q.; Li, J.; Ban, L.P. Morphology and distribution of antennal sensilla in three species of Thripidae (Thysanoptera) infesting alfalfa Medicago sativa. Insects 2021, 12, 81. [Google Scholar] [CrossRef]
- Shi, X.; Zhang, S.F.; Liu, F.; Xu, F.Y.; Zhang, F.B.; Guo, X.B.; Zhang, Z.; Kong, X.B. SEM analysis of sensilla on the mouthparts and antennae of Asian larch bark beetle Ips subelongatus. Micron 2021, 140, 102976. [Google Scholar] [CrossRef]
- Du, Y.J.; Yan, F.S.; Tang, J. Structure and function of olfactory receptors in Aphis glycines antennae. Acta Entomol. Sin. 1995, 38, 1–7. (In Chinese) [Google Scholar]
- Brito, N.F.; Moreira, M.F.; Melo, A.C. A look inside odorant-binding proteins in insect chemoreception. J. Insect Physiol. 2016, 95, 51–65. [Google Scholar] [CrossRef]
- Krieger, J.; Breer, H. Olfactory reception in invertebrates. Science 1999, 286, 720–723. [Google Scholar] [CrossRef] [PubMed]
- Fan, J.; Francis, F.; Liu, Y.; Chen, J.L.; Cheng, D.F. An overview of odorant-binding protein functions in insect peripheral olfactory reception. Genet. Mol. Res. 2011, 10, 3056–3069. [Google Scholar] [CrossRef] [PubMed]
- Breer, H.; Krieger, J.; Raming, K. A novel class of binding proteins in the antennae of the silk moth Antheraea pernyi. Insect Biochem. 1990, 20, 735–740. [Google Scholar] [CrossRef]
- Rihani, K.; Ferveur, J.F.; Briand, L. The 40-year mystery of insect odorant-binding proteins. Biomolecules 2021, 11, 509. [Google Scholar] [CrossRef]
- Zhang, Y.; Yang, B.; Wang, G.R. Research progress of soluble proteins on chemosensationin insects. J. Environ. Entomol. 2019, 41, 229–240. (In Chinese) [Google Scholar]
- Wu, F.; Zhang, L.; Qiu, Y.L.; Li, H.L. Research progress of olfactory binding proteins in insects. Acta Entomol. Sin. 2021, 64, 523–535. (In Chinese) [Google Scholar]
- Sun, Y.L.; Dong, J.F.; Song, Y.Q.; Wang, S.L. GOBP1 from the variegated cutworm Peridroma saucia (Hübner) (Lepidoptera: Noctuidae) displays high binding affinities to the behavioral attractant (Z)-3-Hexenyl acetate. Insects 2021, 12, 939. [Google Scholar] [CrossRef]
- Zhou, J.J. Odorant-binding proteins in insects. Vitam. Horm. 2010, 83, 241–272. [Google Scholar]
- Leal, W.S. Odorant reception in insects: Roles of receptors, binding proteins, and degrading enzymes. Annu. Rev. Entomol. 2013, 58, 373–391. [Google Scholar] [CrossRef] [PubMed]
- Venthur, H.; Zhou, J.J. Odorant receptors and odorant-binding proteins as insect pest control targets: A comparative analysis. Front. Physiol. 2018, 9, 1163. [Google Scholar] [CrossRef] [PubMed]
- Li, T.T.; Liu, W.C.; Zhu, J.; Yang, Y.H.; Ma, C.; Lu, C.; Zhang, K.X. Crystal structure and ligand identification of odorant binding protein 4 in the natural predator Chrysopa pallens. Int. J. Biol. Macromol. 2019, 141, 1004–1012. [Google Scholar] [CrossRef] [PubMed]
- Northey, T.; Venthur, H.; De, B.F.; Chauviac, F.X.; Cole, A.; Ribeiro, K.A.J.; Grossi, G.; Falabella, P.; Field, L.M.; Keep, N.H.; et al. Crystal structures and binding dynamics of odorant-binging protein 3 form two aphid species Megoura viciae and Nasonovia ribisniari. Sci. Rep. 2016, 6, 24739. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez, D.; Rihani, K.; Neiers, F.; Poirier, N.; Fraichard, S.; Gotthard, G.; Chertemps, T.; Maïbèche, M.; Ferveur, J.F.; Briand, L. The Drosophila odorant-binding protein 28a is involved in the detection of the floral odour ß-ionone. Cell. Mol. Life Sci. 2020, 77, 2565–2577. [Google Scholar] [CrossRef] [PubMed]
- Sandler, B.H.; Nikonova, L.; Leal, W.S.; Clardy, J. Sexual attraction in the silkworm moth: Structure of the pheromone-binding-protein-bombykol complex. Chem. Biol. 2000, 7, 143–151. [Google Scholar] [CrossRef]
- Horst, R.; Damberger, F.; Luginbühl, P.; Güntert, P.; Peng, G.; Nikonova, L.; Leal, W.S.; Wüthrich, K. NMR structure reveals intramolecular regulation mechanism for pheromone binding and release. Proc. Natl. Acad. Sci. USA 2001, 98, 14374–14379. [Google Scholar] [CrossRef]
- Li, G.; Chen, X.; Li, B.; Zhang, G.; Li, Y.; Wu, J. Binding properties of general odorant binding proteins from the oriental fruit moth, Grapholita molesta (Busck) (Lepidoptera: Tortricidae). PLoS ONE 2016, 11, e0155096. [Google Scholar] [CrossRef]
- Nagnan-Le, M.P.; Huet, J.C.; Maibeche, M.; Pernollet, J.C.; Descoins, C. Purification and characterization of multiple forms of odorant/pheromone binding proteins in the antennae of Mamestra brassicae (Noctuidae). Insect Biochem. Mol. Biol. 1996, 26, 59–67. [Google Scholar] [CrossRef]
- Ahmed, T.; Zhang, T.T.; Wang, Z.Y.; He, K.L.; Bai, S.X. C-terminus methionene specifically involved in binding corn odorants to odorant binding protein 4 in Macrocentrus cingulum. Front. Physiol. 2017, 8, 62. [Google Scholar] [CrossRef]
- Dong, K.; Duan, H.X.; Liu, J.T.; Sun, L.; Gu, S.H.; Yang, R.N.; Dhiloo, K.H.; Gao, X.W.; Zhang, Y.J.; Guo, Y.Y. Key site residues of pheromone-binding protein 1 involved in interacting with sex pheromone components of Helicoverpa armigera. Sci. Rep. 2017, 7, 16859. [Google Scholar] [CrossRef] [PubMed]
- del Mármol, J.; Yedlin, M.A.; Ruta, V. The structural basis of odorant recognition in insect olfactory receptors. Nature 2021, 597, 126–131. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Wang, W.L.; Zhang, Y.X.; Zhang, B.X.; Rao, X.J.; Liu, X.M.; Wang, D.M.; Li, S.G. Transcriptome sequencing reveals abundant olfactory genes in the antennae of the rice leaffolder, Cnaphalocrocis medinalis (Lepidoptera: Pyralidae). Entomol. Sci. 2017, 20, 177–188. [Google Scholar] [CrossRef]
- Zhang, Y.; Yang, B.; Yu, J.; Pang, B.P.; Wang, G.R. Expression profiles and functional prediction of ionotropic receptors in Asian corn borer, Ostrinia furnacalis (Lepidoptera: Crambidae). J. Integr. Agric. 2022, 21, 474–485. [Google Scholar]
- Jiang, X.; Jiang, J.; Yu, M.; Zhang, S.; Qin, Y.; Xu, Y.; Francis, F.; Fan, J.; Chen, J. Functional analysis of odorant-binding proteins for the parasitic host location to implicate convergent evolution between the grain aphid and its parasitoid Aphidius gifuensis. Int. J. Biol. Macromol. 2023, 226, 510–524. [Google Scholar] [CrossRef] [PubMed]
- Zeng, F.F.; Sun, X.; Dong, H.B.; Wang, M.Q. Analysis of a cDNA library from the antenna of Cnaphalocrocis medinalis and the expression pattern of olfactory genes. Biochem. Biophys. Res. Commun. 2013, 433, 463–469. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Shi, X.X.; Zhu, Q.Z.; Jiao, W.J.; Zhu, Z.J.; Yu, H.; Wang, G.Y.; Zhu, Z.R. Identification and expression profiles of putative chemosensory protein genes in Cnaphalocrocis medinalis (Lepidoptera: Pyralidae). J. Asia-Pac. Entomol. 2015, 18, 99–105. [Google Scholar] [CrossRef]
- Zeng, F.F.; Zhao, Z.F.; Yan, M.J.; Zhou, W.; Zhang, Z.; Zhang, A.; Lu, Z.X.; Wang, M.Q. Identification and comparative expression profiles of chemoreception genes revealed from major chemoreception organs of the rice leaf folder, Cnaphalocrocis medinalis (Lepidoptera: Pyralidae). PLoS ONE 2015, 10, e0144267. [Google Scholar] [CrossRef]
- Sun, X.; Zhao, Z.; Zeng, F.; Zhang, A.; Lu, Z.; Wang, M. Functional characterization of a pheromone-binding protein from rice leaffolder Cnaphalocrocis medinalis in detecting pheromones and host plant volatiles. Bull. Entomol. Res. 2016, 106, 781–789. [Google Scholar] [CrossRef]
- Sun, S.F.; Zeng, F.F.; Yi, S.C.; Wang, M.Q. Molecular screening of behaviorally active compounds with CmedOBP14 from the rice leaf folder Cnaphalocrocis medinalis. J. Chem. Ecol. 2019, 45, 849–857. [Google Scholar] [CrossRef]
- Duan, S.G.; Li, D.Z.; Wang, M.Q. Chemosensory proteins used as target for screening behaviourally active compounds in the rice pest Cnaphalocrocis medinalis (Lepidoptera: Pyralidae). Insect Mol. Biol. 2019, 28, 123–135. [Google Scholar] [CrossRef] [PubMed]
- Zeng, F.F.; Liu, H.; Zhang, A.; Lu, Z.X.; Leal, W.S.; Abdelnabby, H.; Wang, M.Q. Three chemosensory proteins from the rice leaf folder Cnaphalocrocis medinalis involved in host volatile and sex pheromone reception. Insect Mol. Biol. 2018, 27, 710–723. [Google Scholar] [CrossRef] [PubMed]
- Kevan, P.G.; Baker, H.G. Insects as flower visitors and pollinators. Annu. Rev. Entomol. 1983, 28, 407–453. [Google Scholar] [CrossRef]
- Schiestl, F.P. The evolution of floral scent and insect chemical communication. Ecol. Lett. 2010, 13, 643–656. [Google Scholar] [CrossRef] [PubMed]
- von Arx, M.; Schmidt-Büsser, D.; Guerin, P.M. Plant volatiles enhance behavioral responses of grapevine moth males, Lobesia botrana to sex pheromone. J. Chem. Ecol. 2012, 38, 222–225. [Google Scholar] [CrossRef]
- Ochieng, S.A.; Park, K.C.; Baker, T.C. Host plant volatiles synergize responses of sex pheromone-specific olfactory receptor neurons in male Helicoverpa zea. J. Comp. Physiology. A Neuroethol. Sens. Neural Behav. Physiol. 2002, 188, 325–333. [Google Scholar] [CrossRef]
- Chaffiol, A.; Dupuy, F.; Barrozo, R.B.; Kropf, J.; Renou, M.; Rospars, J.P.; Anton, S. Pheromone modulates plant odor responses in the antennal lobe of a moth. Chem. Senses 2014, 39, 451–463. [Google Scholar] [CrossRef]
- Molnár, B.P.; Kárpáti, Z.; Nagy, A.; Szarukán, I.; Csabai, J.; Koczor, S.; Tóth, M. Development of a female-targeted lure for the box tree moth Cydalima perspectalis (Lepidoptera: Crambidae): A preliminary report. J. Chem. Ecol. 2019, 45, 657–666. [Google Scholar] [CrossRef]
- Meagher, R.L. Trapping noctuid moths with synthetic floral volatile lures. Entomol. Exp. Appl. 2002, 103, 3. [Google Scholar] [CrossRef]
- Guedot, C.; Landolt, P.J.; Smithhisler, C.L. Odorants of the flowers of butterfly bush, Buddleja davidii, as possible attractants of pest species of moths. Fla. Entomol. 2008, 91, 576–582. [Google Scholar]
- Li, P.; Zhu, J.; Qin, Y. Enhanced attraction of Plutella xylostella (Lepidoptera: Plutellidae) to pheromone-baited traps with the addition of green leaf volatiles. J. Econ. Entomol. 2012, 105, 1149–1156. [Google Scholar] [CrossRef] [PubMed]
- Ju, Q.; Guo, X.Q.; Li, X.; Jiang, X.J.; Jiang, X.G.; Ni, W.L.; Qu, M.J. Plant volatiles increase sex pheromone attraction of Holotrichia parallela (Coleoptera: Scarabaeoidea). J. Chem. Ecol. 2017, 43, 236–242. [Google Scholar] [CrossRef] [PubMed]
- Pregitzer, P.; Schubert, M.; Breer, H.; Hansson, B.S.; Sachse, S.; Krieger, J. Plant odorants interfere with detection of sex pheromone signals by male Heliothis virescens. Front. Cell. Neurosci. 2012, 6, 42. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Su, Q.F.; Wang, L.S.; Lv, M.W.; Hou, Y.X.; Li, S.S. Linalool: A ubiquitous floral volatile mediating the communication between plants and insects. J. Syst. Evol. 2023, 61, 538–549. [Google Scholar] [CrossRef]
- Foote, G.G.; Runyon, J.B.; Fettig, C.J.; Foote, N.E.; Ross, D.W. Wild bee response to application of the douglas-fir beetle anti-aggregation pheromone, 3-methylcyclohex-2-en-1-one. J. Econ. Entomol. 2021, 114, 2121–2126. [Google Scholar] [CrossRef] [PubMed]
- Wang, A.J.; Zhang, K.X.; Gao, Y.L.; Weng, A.Z.; Wang, L.Y.; Zhang, Y.H.; Zhang, Z.; She, D.M.; Ning, J.; Mei, X.D. Synthesis and bioactivity studies of sex pheromone analogs of the diamond back moth, Plutella xylostella. Pest Manag. Sci. 2019, 75, 1045–1055. [Google Scholar] [CrossRef]
- Guo, X.; Yu, Q.; Chen, D.; Wei, J.; Yang, P.; Yu, J.; Wang, X.; Kang, L. 4-Vinylanisole is an aggregation pheromone in locusts. Nature 2020, 584, 584–588. [Google Scholar] [CrossRef]
- Khallaf, M.A.; Cui, R.; Weißflog, J.; Erdogmus, M.; Svatoš, A.; Dweck, H.K.M.; Valenzano, D.R.; Hansson, B.S.; Knaden, M. Large-scale characterization of sex pheromone communication systems in Drosophila. Nat. Commun. 2021, 12, 4165. [Google Scholar] [CrossRef]
- Field, L.M.; Pickett, J.A.; Wadhams, L.J. Molecular studies in insect olfaction. Insect Mol. Biol. 2000, 9, 545–551. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
| Plant Organs | Types of Volatiles |
|---|---|
| Green leaf | C6/C9 aldehydes, alcohols, and esters |
| Flower | Terpenoids, phenylpropanoids/benzenoids, and fatty acid derivatives |
| Root | Alcohols, ketones, aldehydes, esters, terpenes, furans, organic acids, aromatic compounds, and sulfur compounds |
| Gene Name | GenBank a | Functions of Olfactory-Related Proteins |
|---|---|---|
| CmedPBP4 | KC507185.1 | (Z)-13-octadecenyl acetate, (Z)-11-hexadecenal, (Z)-13-octadecenol, cyclohexanol, nerolidol, cedrol, dodecanal, ionone, (−)-α-cedrene, (Z)-farnesene, β-myrcene, R-(+)-limonene, (−)-limonene, (+)-3-carene [69] |
| CmedOBP14 | KP975125.1 | cedrol, β-ionone, nerolidol, 3-carene, 1-octen-3-ol, tetradecane, D-limonene, 3-pentanol, L-limonene, P-cymene, (E)-2-hexen-1-ol, 2-heptanone, (Z)-hex-3-en-1-ol, hexadecane, cyclohexanol, linalool, 2-tridecanone, henicosane, heptan-1-ol, α-cedrene, octadecane, sabinene, dodecanal, eicosane, nonadecane, trans-2-hexenal [70] |
| CmedCSP1 | KC507178.1 | (+/−)-linalool, nerolidol, tetradecane, nerolidol, cedrol, cis-β-farnesene, terpinene-4-ol, α-terpineol, α-terpinene, β-myrcene, sabinene, P-cymene, methyl benzoate, γ-terpinene, heptano, R-(+)-limonene, 2-heptanone, (1R)-(+)-α-pinene [72] |
| CmedCSP2 | KC507180.1 | cyclohexanol, heptadecane, octadecane, nonadecane, eicosane, (−)-α-cedrene, nerolidol, cedrol, cis-β-farnesene, α-terpineol, α-terpinene, β-myrcene, sabinene, P-cymene, methyl benzoate, (−)-(E)-caryophyllene, γ-terpinene, octane, ionone, 2-heptanone [72] |
| CmedCSP3 | KC507182.1 | octadecane, (−)-α-cedrene, cis-β-farnesene, (−)-(E)-caryophyllene, (Z)-11-hexadecenyl acetate, (Z)-11-hexadecenal [72] |
| CmedCSP33 | KP975096.1 | 2-heptanone, β-ionone, R-(+)-limonene, cyclohexanol, (E)-2-hexen-1-ol, 3-pentanol, nerolidol [71] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Qian, Q.; Cui, J.; Miao, Y.; Xu, X.; Gao, H.; Xu, H.; Lu, Z.; Zhu, P. The Plant Volatile-Sensing Mechanism of Insects and Its Utilization. Plants 2024, 13, 185. https://doi.org/10.3390/plants13020185
Qian Q, Cui J, Miao Y, Xu X, Gao H, Xu H, Lu Z, Zhu P. The Plant Volatile-Sensing Mechanism of Insects and Its Utilization. Plants. 2024; 13(2):185. https://doi.org/10.3390/plants13020185
Chicago/Turabian StyleQian, Qi, Jiarong Cui, Yuanyuan Miao, Xiaofang Xu, Huiying Gao, Hongxing Xu, Zhongxian Lu, and Pingyang Zhu. 2024. "The Plant Volatile-Sensing Mechanism of Insects and Its Utilization" Plants 13, no. 2: 185. https://doi.org/10.3390/plants13020185
APA StyleQian, Q., Cui, J., Miao, Y., Xu, X., Gao, H., Xu, H., Lu, Z., & Zhu, P. (2024). The Plant Volatile-Sensing Mechanism of Insects and Its Utilization. Plants, 13(2), 185. https://doi.org/10.3390/plants13020185