
In Vivo Study of Moringa oleifera Seed Extracts as Potential Sources of Neuroprotection against Rotenone-Induced Neurotoxicity

 ,
,  ,
,  ,
, 
Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Material
2.2. Chemicals
2.3. Preparation of M. oleifera Seed Extracts
2.4. Experimental Design and Extract Administration
2.5. Behavioral Tests for Motor Functions
2.6. Mice Brain Procurement
2.7. Antioxidant Capacity Determination
2.8. Histopathological Studies
2.9. Statistical Analysis
2.10. Gas Chromatography−Mass Spectrometry (GC-MS) Analysis of Ethanolic Extracts of M. oleifera
2.11. Identification of Bioactive Components from the Ethanolic Extract of M. oleifera
3. Results
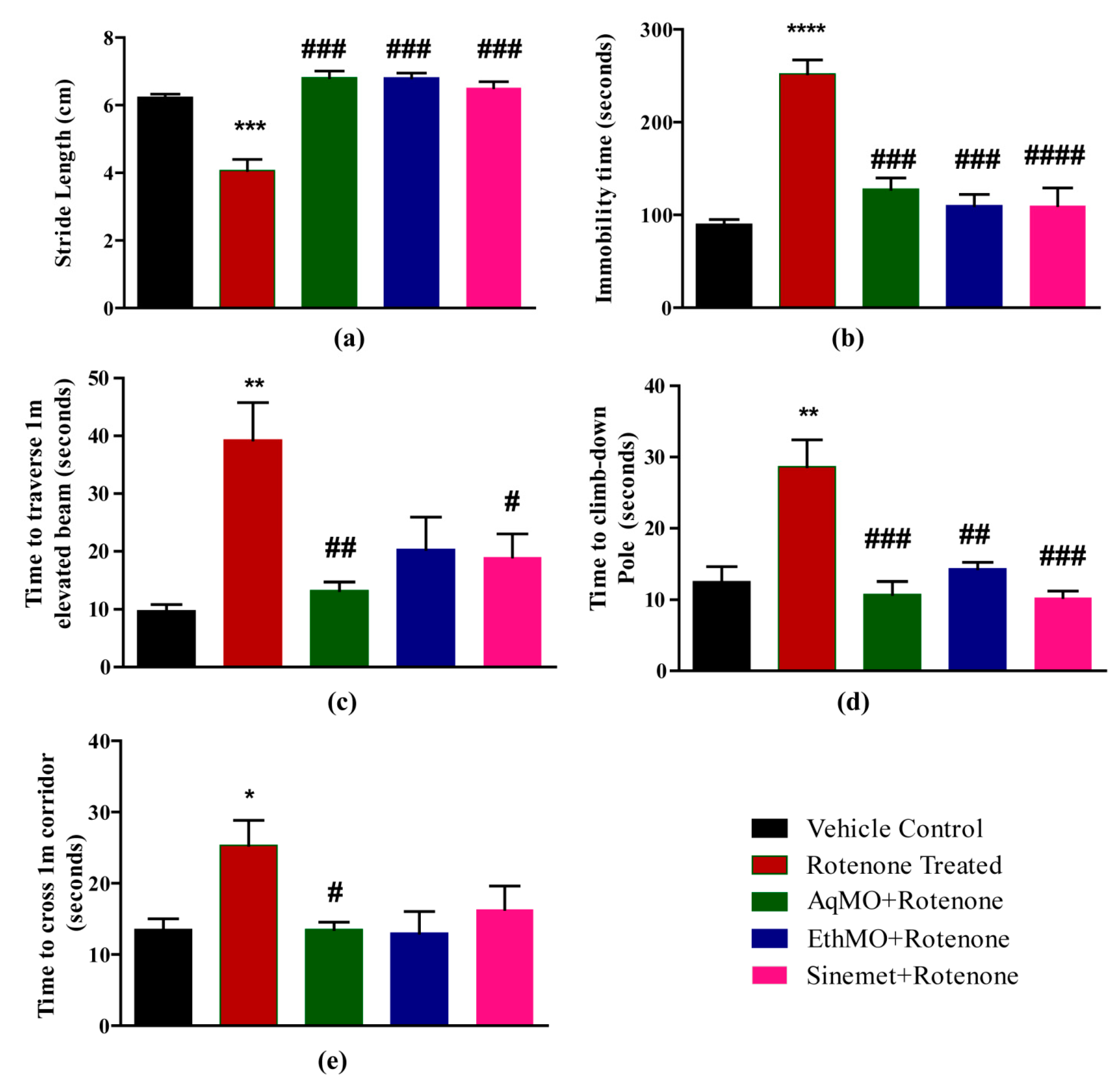
3.1. MO Maintains Locomotor Activities
3.2. MO Effects on Oxidative Stress
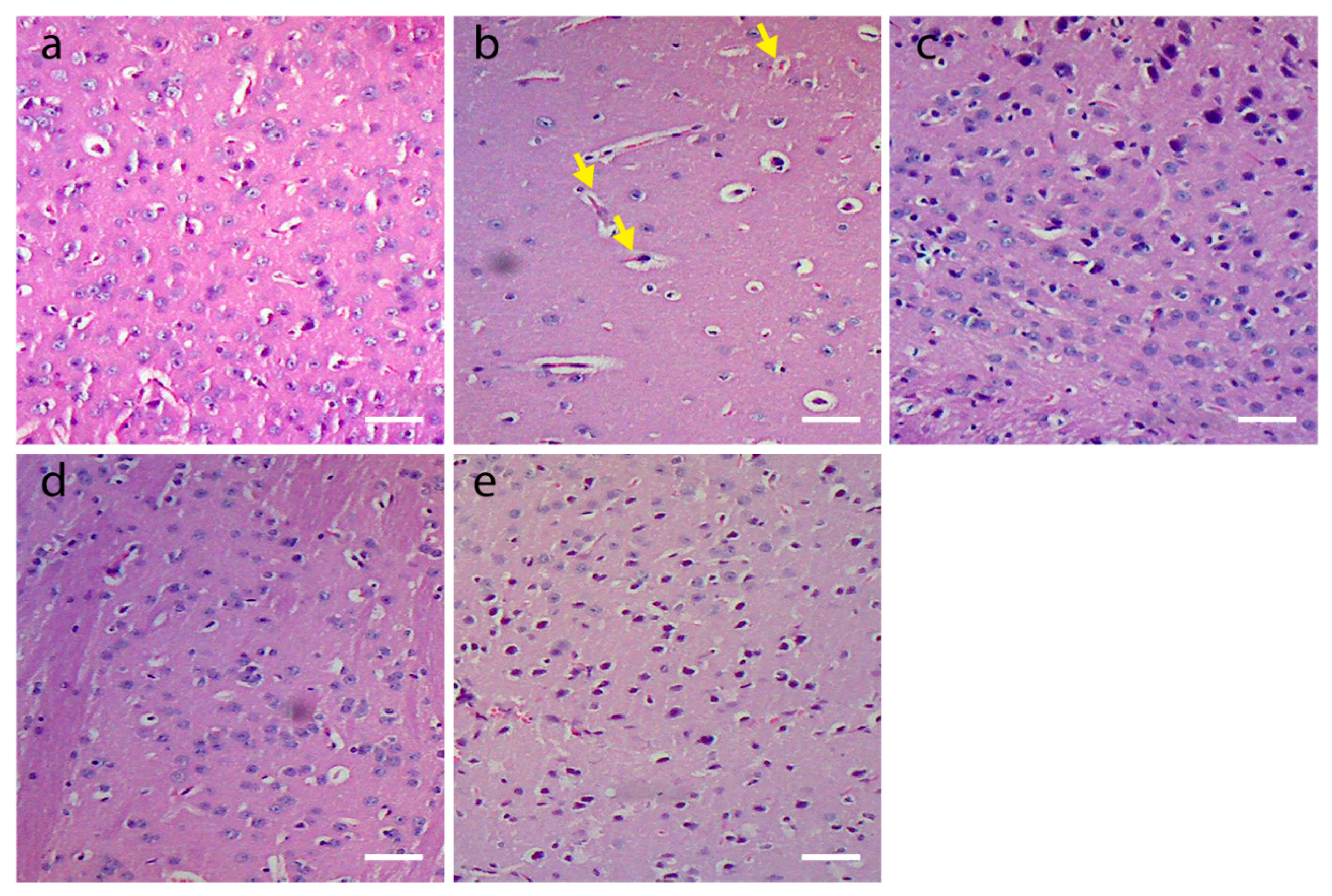
3.3. Histological Findings
3.4. Phytochemical Characterization of EthMO Extract Using GC-MS Analysis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Raza, C.; Anjum, R.; Shakeel, N.U.A. Parkinson’s disease: Mechanisms, translational models and management strategies. Life Sci. 2019, 226, 77–90. [Google Scholar] [CrossRef]
- Blesa, J.; Trigo-Damas, I.; Quiroga-Varela, A.; Jackson-Lewis, V.R. Oxidative stress and Parkinson’s disease. Front. Neuroanat. 2015, 9, 91. [Google Scholar] [CrossRef]
- Trist, B.G.; Hare, D.J.; Double, K.L. Oxidative stress in the aging substantia nigra and the etiology of Parkinson’s disease. Aging Cell 2019, 18, e13031. [Google Scholar] [CrossRef]
- Cheng, H.C.; Ulane, C.M.; Burke, R.E. Clinical progression in Parkinson disease and the neurobiology of axons. Ann. Neurol. 2010, 67, 715–725. [Google Scholar] [CrossRef]
- Li, J.L.; Lin, T.Y.; Chen, P.L.; Guo, T.N.; Huang, S.Y.; Chen, C.H.; Lin, C.H.; Chan, C.C. Mitochondrial Function and Parkinson’s Disease: From the Perspective of the Electron Transport Chain. Front. Mol. Neurosci. 2021, 14, 797833. [Google Scholar] [CrossRef]
- Gumber, A.; Ramaswamy, B.; Thongchundee, O. Effects of Parkinson’s on employment, cost of care, and quality of life of people with condition and family caregivers in the UK: A systematic literature review. Patient Relat. Outcome Meas. 2019, 10, 321–333. [Google Scholar] [CrossRef]
- Kim, T.W.; Koo, S.Y.; Studer, L. Pluripotent Stem Cell Therapies for Parkinson Disease: Present Challenges and Future Opportunities. Front. Cell Dev. Biol. 2020, 8, 729. [Google Scholar] [CrossRef]
- Limanaqi, F.; Biagioni, F.; Mastroiacovo, F.; Polzella, M.; Lazzeri, G.; Fornai, F. Merging the multi-target effects of phytochemicals in neurodegeneration: From oxidative stress to protein aggregation and inflammation. Antioxidants 2020, 9, 1022. [Google Scholar] [CrossRef]
- Velmurugan, B.K.; Rathinasamy, B.; Lohanathan, B.P.; Thiyagarajan, V.; Weng, C.-F. Neuroprotective role of phytochemicals. Molecules 2018, 23, 2485. [Google Scholar] [CrossRef]
- Renaud, J.; Martinoli, M.-G. Considerations for the use of polyphenols as therapies in neurodegenerative diseases. Int. J. Mol. Sci. 2019, 20, 1883. [Google Scholar] [CrossRef]
- Shahpiri, Z.; Bahramsoltani, R.; Farzaei, M.H.; Farzaei, F.; Rahimi, R. Phytochemicals as future drugs for Parkinson’s disease: A comprehensive review. Rev. Neurosci. 2016, 27, 651–668. [Google Scholar] [CrossRef]
- Uddin, M.S.; Al Mamun, A.; Kabir, M.T.; Ahmad, J.; Jeandet, P.; Sarwar, M.S.; Ashraf, G.M.; Aleya, L. Neuroprotective role of polyphenols against oxidative stress-mediated neurodegeneration. Eur. J. Pharmacol. 2020, 886, 173412. [Google Scholar] [CrossRef]
- Li, B.-Y.; Yuan, Y.-H.; Hu, J.-F.; Zhao, Q.; Zhang, D.-M.; Chen, N.-H. Protective effect of Bu-7, a flavonoid extracted from Clausena lansium, against rotenone injury in PC12 cells. Acta Pharmacol. Sin. 2011, 32, 1321–1326. [Google Scholar] [CrossRef]
- Bové, J.; Prou, D.; Perier, C.; Przedborski, S. Toxin-induced models of Parkinson’s disease. NeuroRx 2005, 2, 484–494. [Google Scholar] [CrossRef]
- Pan-Montojo, F.; Anichtchik, O.; Dening, Y.; Knels, L.; Pursche, S.; Jung, R.; Jackson, S.; Gille, G.; Spillantini, M.G.; Reichmann, H. Progression of Parkinson’s disease pathology is reproduced by intragastric administration of rotenone in mice. PLoS ONE 2010, 5, e8762. [Google Scholar] [CrossRef]
- El-Ghaiesh, S.H.; Bahr, H.I.; Ibrahiem, A.T.; Ghorab, D.; Alomar, S.Y.; Farag, N.E.; Zaitone, S.A. Metformin Protects from Rotenone-Induced Nigrostriatal Neuronal Death in Adult Mice by Activating AMPK-FOXO3 Signaling and Mitigation of Angiogenesis. Front. Mol. Neurosci. 2020, 13, 84. [Google Scholar] [CrossRef]
- Sherer, T.B.; Betarbet, R.; Testa, C.M.; Seo, B.B.; Richardson, J.R.; Kim, J.H.; Miller, G.W.; Yagi, T.; Matsuno-Yagi, A.; Greenamyre, J.T. Mechanism of toxicity in rotenone models of Parkinson’s disease. J. Neurosci. Off. J. Soc. Neurosci. 2003, 23, 10756–10764. [Google Scholar] [CrossRef]
- Jahan, I.A.; Hossain, M.H.; Ahmed, K.S.; Sultana, Z.; Biswas, P.K.; Nada, K. Antioxidant activity of Moringa oleifera seed extracts. Orient. Pharm. Exp. Med. 2018, 18, 299–307. [Google Scholar] [CrossRef]
- Chandrashekar, S.; Vijayakumar, R.; Chelliah, R.; Oh, D.H. Identification and Purification of Potential Bioactive Peptide of Moringa oleifera Seed Extracts. Plants 2020, 9, 1445. [Google Scholar] [CrossRef]
- Idris, M.A.; Jami, M.S.; Hammed, A.M.; Jamal, P. Moringa oleifera seed extract: A review on its environmental applications. Int. J. Appl. Environ. Sci. 2016, 11, 1469–1486. [Google Scholar]
- Bassey, K.; Mabowe, M.; Mothibe, M.; Witika, B.A. Chemical Characterization and Nutritional Markers of South African Moringa oleifera Seed Oils. Molecules 2022, 27, 5749. [Google Scholar] [CrossRef] [PubMed]
- Saini, R.K.; Sivanesan, I.; Keum, Y.S. Phytochemicals of Moringa oleifera: A review of their nutritional, therapeutic and industrial significance. 3 Biotech 2016, 6, 203. [Google Scholar] [CrossRef] [PubMed]
- Nafiu, A.O.; Akomolafe, R.O.; Alabi, Q.K.; Idowu, C.O.; Odujoko, O.O. Effect of fatty acids from ethanol extract of Moringa oleifera seeds on kidney function impairment and oxidative stress induced by gentamicin in rats. Biomed. Pharmacother. 2019, 117, 109154. [Google Scholar] [CrossRef]
- Makav, M.; Eroğlu, H.A. Recuperative effect of estrogen on rotenone-induced experimental model of Parkinson’s disease in rats. Environ. Sci. Pollut. Res. 2021, 28, 21266–21275. [Google Scholar] [CrossRef] [PubMed]
- Prasad, E.M.; Hung, S.Y. Behavioral Tests in Neurotoxin-Induced Animal Models of Parkinson’s Disease. Antioxidants 2020, 9, 1007. [Google Scholar] [CrossRef] [PubMed]
- Rudolf, J.; Dondorp, D.; Canon, L.; Tieo, S.; Chatzigeorgiou, M. Automated behavioural analysis reveals the basic behavioural repertoire of the urochordate Ciona intestinalis. Sci. Rep. 2019, 9, 2416. [Google Scholar] [CrossRef]
- Boix, J.; von Hieber, D.; Connor, B. Gait Analysis for Early Detection of Motor Symptoms in the 6-OHDA Rat Model of Parkinson’s Disease. Front. Behav. Neurosci. 2018, 12, 39. [Google Scholar] [CrossRef] [PubMed]
- Can, A.; Dao, D.T.; Terrillion, C.E.; Piantadosi, S.C.; Bhat, S.; Gould, T.D. The tail suspension test. J. Vis. Exp. JoVE 2012, 59, e3769. [Google Scholar] [CrossRef]
- Rajasankar, S.; Manivasagam, T.; Surendran, S. Ashwagandha leaf extract: A potential agent in treating oxidative damage and physiological abnormalities seen in a mouse model of Parkinson’s disease. Neurosci. Lett. 2009, 454, 11–15. [Google Scholar] [CrossRef]
- Glajch, K.E.; Fleming, S.M.; Surmeier, D.J.; Osten, P. Sensorimotor assessment of the unilateral 6-hydroxydopamine mouse model of Parkinson’s disease. Behav. Brain Res. 2012, 230, 309–316. [Google Scholar] [CrossRef]
- Blume, S.R.; Cass, D.K.; Tseng, K.Y. Stepping test in mice: A reliable approach in determining forelimb akinesia in MPTP-induced Parkinsonism. Exp. Neurol. 2009, 219, 208–211. [Google Scholar] [CrossRef] [PubMed]
- Wright, J.R.; Colby, H.D.; Miles, P.R. Cytosolic factors which affect microsomal lipid peroxidation in lung and liver. Arch Biochem. Biophys. 1981, 206, 296–304. [Google Scholar] [CrossRef]
- Faheem, M.; Adeel, M.; Khaliq, S.; Lone, K.P.; El-Din-H-Sayed, A. Bisphenol-A induced antioxidants imbalance and cytokines alteration leading to immune suppression during larval development of Labeo rohita. Environ. Sci. Pollut. Res. 2020, 27, 26800–26809. [Google Scholar] [CrossRef] [PubMed]
- Baiguera, C.; Alghisi, M.; Pinna, A.; Bellucci, A.; De Luca, M.A.; Frau, L.; Morelli, M.; Ingrassia, R.; Benarese, M.; Porrini, V.; et al. Late-onset Parkinsonism in NFkappaB/c-Rel-deficient mice. Brain 2012, 135, 2750–2765. [Google Scholar] [CrossRef]
- Stutz, B.; Nasrallah, C.; Nigro, M.; Curry, D.; Liu, Z.-W.; Gao, X.-B.; Elsworth, J.D.; Mintz, L.; Horvath, T.L. Dopamine neuronal protection in the mouse Substantia nigra by GHSR is independent of electric activity. Mol. Metab. 2019, 24, 120–138. [Google Scholar] [CrossRef]
- Cardiff, R.D.; Miller, C.H.; Munn, R.J. Manual hematoxylin and eosin staining of mouse tissue sections. Cold Spring Harb. Protoc. 2014, 2014, 655–658. [Google Scholar] [CrossRef] [PubMed]
- Mohammadzadeh, L.; Hosseinzadeh, H.; Abnous, K.; Razavi, B.M. Neuroprotective potential of crocin against malathion-induced motor deficit and neurochemical alterations in rats. Environ. Sci. Pollut. Res. Int. 2018, 25, 4904–4914. [Google Scholar] [CrossRef]
- Bonito-Oliva, A.; Masini, D.; Fisone, G. A mouse model of non-motor symptoms in Parkinson’s disease: Focus on pharmacological interventions targeting affective dysfunctions. Front. Behav. Neurosci. 2014, 8, 290. [Google Scholar] [CrossRef] [PubMed]
- Wegrzynowicz, M.; Bar-On, D.; Calo’, L.; Anichtchik, O.; Iovino, M.; Xia, J.; Ryazanov, S.; Leonov, A.; Giese, A.; Dalley, J.W.; et al. Depopulation of dense α-synuclein aggregates is associated with rescue of dopamine neuron dysfunction and death in a new Parkinson’s disease model. Acta Neuropathol. 2019, 138, 575–595. [Google Scholar] [CrossRef]
- Ebrahimi-Ghiri, M.; Shahini, F.; Zarrindast, M.R. The effect of URB597, exercise or their combination on the performance of 6-OHDA mouse model of Parkinson disease in the elevated plus maze, tail suspension test and step-down task. Metab. Brain Dis. 2021, 36, 2579–2588. [Google Scholar] [CrossRef]
- Asakawa, T.; Fang, H.; Sugiyama, K.; Nozaki, T.; Hong, Z.; Yang, Y.; Hua, F.; Ding, G.; Chao, D.; Fenoy, A.J.; et al. Animal behavioral assessments in current research of Parkinson’s disease. Neurosci. Biobehav. Rev. 2016, 65, 63–94. [Google Scholar] [CrossRef] [PubMed]
- Bresgen, N.; Eckl, P.M. Oxidative stress and the homeodynamics of iron metabolism. Biomolecules 2015, 5, 808–847. [Google Scholar] [CrossRef] [PubMed]
- Faucheux, B.A.; Martin, M.E.; Beaumont, C.; Hauw, J.J.; Agid, Y.; Hirsch, E.C. Neuromelanin associated redox-active iron is increased in the substantia nigra of patients with Parkinson’s disease. J. Neurochem. 2003, 86, 1142–1148. [Google Scholar] [CrossRef] [PubMed]
- Testa, C.M.; Sherer, T.B.; Greenamyre, J.T. Rotenone induces oxidative stress and dopaminergic neuron damage in organotypic substantia nigra cultures. Brain Research. Mol. Brain Res. 2005, 134, 109–118. [Google Scholar] [CrossRef]
- Pearce, R.K.B.; Owen, A.; Daniel, S.; Jenner, P.; Marsden, C.D. Alterations in the distribution of glutathione in the substantia nigra in Parkinson’s disease. J. Neural Transm. 1997, 104, 661–677. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.L.; Zhang, J.; Li, Y.P.; Dong, L.; Chen, Y.Z. Potential use of glutathione as a treatment for Parkinson’s disease. Exp. Ther. Med. 2021, 21, 125. [Google Scholar] [CrossRef] [PubMed]
- Fitzmaurice, P.S.; Ang, L.; Guttman, M.; Rajput, A.H.; Furukawa, Y.; Kish, S.J. Nigral glutathione deficiency is not specific for idiopathic Parkinson’s disease. Mov. Disord. Off. J. Mov. Disord. Soc. 2003, 18, 969–976. [Google Scholar] [CrossRef] [PubMed]
- Jewett, M.; Dickson, E.; Brolin, K.; Negrini, M.; Jimenez-Ferrer, I.; Swanberg, M. Glutathione S-Transferase Alpha 4 Prevents Dopamine Neurodegeneration in a Rat Alpha-Synuclein Model of Parkinson’s Disease. Front. Neurol. 2018, 9, 222. [Google Scholar] [CrossRef] [PubMed]
- Whitworth, A.J.; Theodore, D.A.; Greene, J.C.; Beneš, H.; Wes, P.D.; Pallanck, L.J. Increased glutathione S-transferase activity rescues dopaminergic neuron loss in a Drosophila model of Parkinson’s disease. Proc. Natl. Acad. Sci. USA 2005, 102, 8024–8029. [Google Scholar] [CrossRef]
- Cannon, J.R.; Tapias, V.; Na, H.M.; Honick, A.S.; Drolet, R.E.; Greenamyre, J.T. A highly reproducible rotenone model of Parkinson’s disease. Neurobiol. Dis. 2009, 34, 279–290. [Google Scholar] [CrossRef]
- Kumar, R.; Khurana, N.; Singh, S.K.; Khurana, S.; Verma, S.; Sharma, N.; Vyas, M.; Dua, K.; Khursheed, R.; Awasthi, A.; et al. Improved neuroprotective activity of Fisetin through SNEDDS in ameliorating the behavioral alterations produced in rotenone-induced Parkinson’s model. Environ. Sci. Pollut. Res. Int. 2022, 29, 50488–50499. [Google Scholar] [CrossRef] [PubMed]
- Jayaraj, R.L.; Beiram, R.; Azimullah, S.; M, F.N.; Ojha, S.K.; Adem, A.; Jalal, F.Y. Noscapine Prevents Rotenone-Induced Neurotoxicity: Involvement of Oxidative Stress, Neuroinflammation and Autophagy Pathways. Molecules 2021, 26, 4627. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Animal Group Title | Treatment | Duration |
|---|---|---|
| 1. Vehicle Control | DMSO containing sunflower oil treated (1 mL/kg) + distilled water, oral administration (10 mL/kg) | 21 days (once a day) |
| 2. Rotenone Control | Rotenone (2.5 mg/kg) in 2.5% DMSO, subcutaneous administration (1 mL/kg) + distilled water, oral administration (10 mL/kg) | 21 days (once a day) |
| 3. EthMO Seed Extract Treated | Rotenone (2.5 mg/kg) in 2.5% DMSO containing sunflower oil, subcutaneous administration (1 mL/kg) + 200 mg/kg EthMO extract, oral administration (10 mL/kg) | 21 days (once a day) |
| 4. AqMO Seed Extract Treated | Rotenone (2.5 mg/kg) in 2.5% DMSO, subcutaneous administration (1 mL/kg) + 200 mg/kg AqMO extract, oral administration (10 mL/kg) | 21 days (once a day) |
| 5. Sinemet Control | Rotenone (2.5 mg/kg) in 2.5% DMSO, subcutaneous administration (1 mL/kg) + Sinemet (20 mg/kg) in distilled water, oral administration (10 mL/kg) | 21 days (once a day) |
| Peak# | R. Time | Compounds | M.F. | M.W. | CAS No. | EthMO (%) |
|---|---|---|---|---|---|---|
| 1 | 6.582 | 3-Pyrrolidinol | C4H9NO | 87 | 2799-21-5 | 14.5 |
| 2 | 8.301 | 3-Ethoxycarbonyl-5-hydroxytetrahydropyran-2-one | C8H12O5 | 188 | - | 15.9 |
| 3 | 11.794 | Diethyl Phthalate | C12H14O4 | 222 | 84-66-2 | 8.8 |
| 4 | 14.190 | Hexadecanoic acid, methyl ester | C17H34O2 | 270 | 112-39-0 | 2.5 |
| 5 | 14.268 | (9Z)-8-Methyl-9-tetradecenoic acid | C15H28O2 | 240 | 457653-87-1 | 3.8 |
| 6 | 16.465 | Hexadecyl 2,2,3,3,3-pentafluoropropanoate | C19H33F5O2 | 388 | 6222-7-7 | 8.3 |
| 7 | 17.437 | Hexadecanoic acid, 2,3-bis[(trimethylsilyl)oxy]propyl ester | C25H54O4Si2 | 474 | 1188-74-5 | 11.4 |
| 8 | 17.823 | Hexadecanoic acid | C16H32O2 | 256 | 57-10-3 | 7.0 |
| 9 | 18.203 | Octadecanoic acid | C18H36O2 | 284 | 57-11-4 | 3.0 |
| 10 | 18.771 | Di-n-octyl phthalate | C24H38O4 | 390 | 117-84-0 | 14.9 |
| Total Identified | 90.1 | |||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Raza, C.; Mohsin, S.; Faheem, M.; Hanif, U.; Alkhathlan, H.Z.; Shaik, M.R.; Riaz, H.A.; Anjum, R.; Jurrat, H.; Khan, M. In Vivo Study of Moringa oleifera Seed Extracts as Potential Sources of Neuroprotection against Rotenone-Induced Neurotoxicity. Plants 2024, 13, 1479. https://doi.org/10.3390/plants13111479
Raza C, Mohsin S, Faheem M, Hanif U, Alkhathlan HZ, Shaik MR, Riaz HA, Anjum R, Jurrat H, Khan M. In Vivo Study of Moringa oleifera Seed Extracts as Potential Sources of Neuroprotection against Rotenone-Induced Neurotoxicity. Plants. 2024; 13(11):1479. https://doi.org/10.3390/plants13111479
Chicago/Turabian StyleRaza, Chand, Sehrish Mohsin, Mehwish Faheem, Uzma Hanif, Hamad Z. Alkhathlan, Mohammed Rafi Shaik, Hasib Aamir Riaz, Rabia Anjum, Husna Jurrat, and Merajuddin Khan. 2024. "In Vivo Study of Moringa oleifera Seed Extracts as Potential Sources of Neuroprotection against Rotenone-Induced Neurotoxicity" Plants 13, no. 11: 1479. https://doi.org/10.3390/plants13111479
APA StyleRaza, C., Mohsin, S., Faheem, M., Hanif, U., Alkhathlan, H. Z., Shaik, M. R., Riaz, H. A., Anjum, R., Jurrat, H., & Khan, M. (2024). In Vivo Study of Moringa oleifera Seed Extracts as Potential Sources of Neuroprotection against Rotenone-Induced Neurotoxicity. Plants, 13(11), 1479. https://doi.org/10.3390/plants13111479