


The Effect of Combined Elicitation with Light and Temperature on the Chlorogenic Acid Content, Total Phenolic Content and Antioxidant Activity of Berula erecta in Tissue Culture

 ,
, 

Abstract

1. Introduction
2. Results
2.1. Influence of Light Quality and Temperature on Plant Development
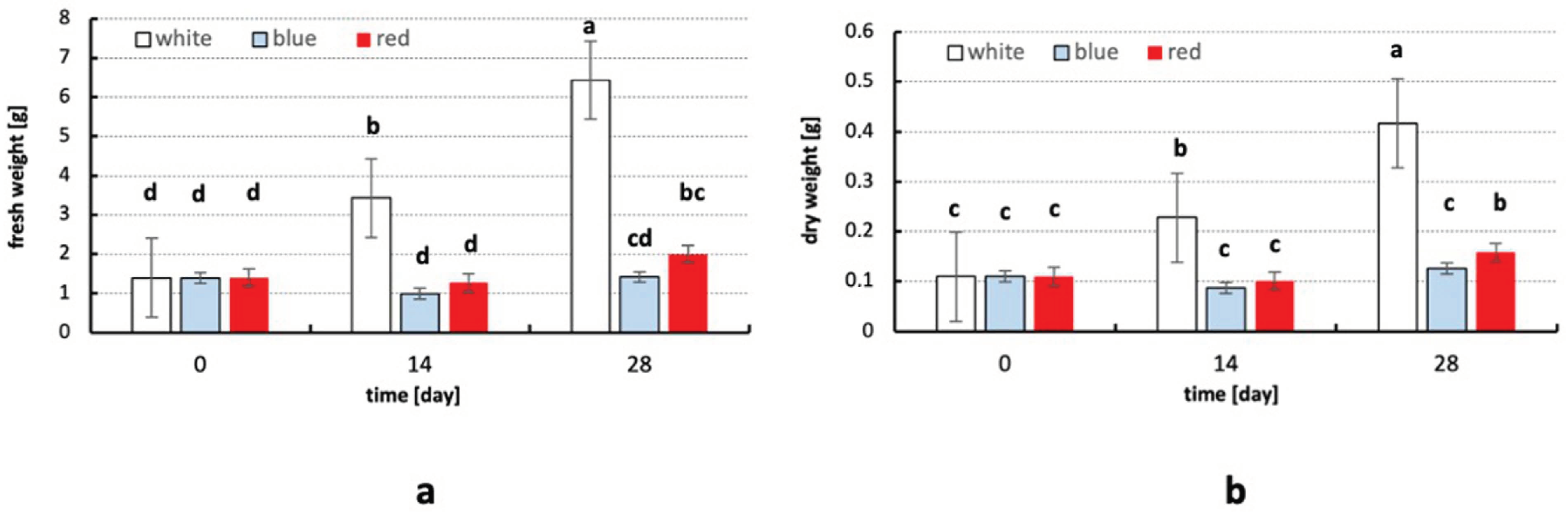
2.2. Influence of Light Quality and Temperature on Fresh and Dry Weight
2.3. Influence of Light Quality and Temperature on Chlorogenic Acid Contents
2.4. Influence of Light Wavelength and Temperature on Total Phenolic Content
2.5. Influence of Light Wavelength and Temperature on the Antioxidant Activity of Ethanolic Extracts of Berula erecta
2.6. Correlations between TPC and Antioxidant Activity in Relation to Chlorogenic Acid Content
2.7. Influence of Light Wavelength and Temperature on the Yield of Chlorogenic Acid in Tissue Culture of Berula erecta Grown In Vitro
3. Discussion
3.1. The Influence of Light Quality and Temperature on Plant Development
3.2. The Influence of Light Quality and Temperature on Fresh and Dry Weight of Plants
3.3. The Influence of Light Wavelength and Temperature on Chlorogenic Acid and Total Phenolics’ Content in Berula erecta
3.4. The Influence of Light Wavelength and Temperature on the Antioxidant Activity of Plant Ethanolic Extracts of Berula erecta
3.5. Correlations between TPC and Antioxidant Activity in Relation to Chlorogenic Acid Content
3.6. The Influence of Light Wavelength and Temperature on the Yield of Chlorogenic Acid in a Tissue Culture of Berula erecta Grown In Vitro
4. Materials and Methods
4.1. Chemicals
4.2. Plant Material
4.3. Plant Development
4.4. Determination of Fresh and Dry Weights
4.5. Ultrasonic Extraction (UE) of Bioactive Compounds from Berula erecta
4.6. Determination of the Total Phenolic Content (TPC)
4.7. DPPH-Radical-Scavenging Activity of Antioxidants
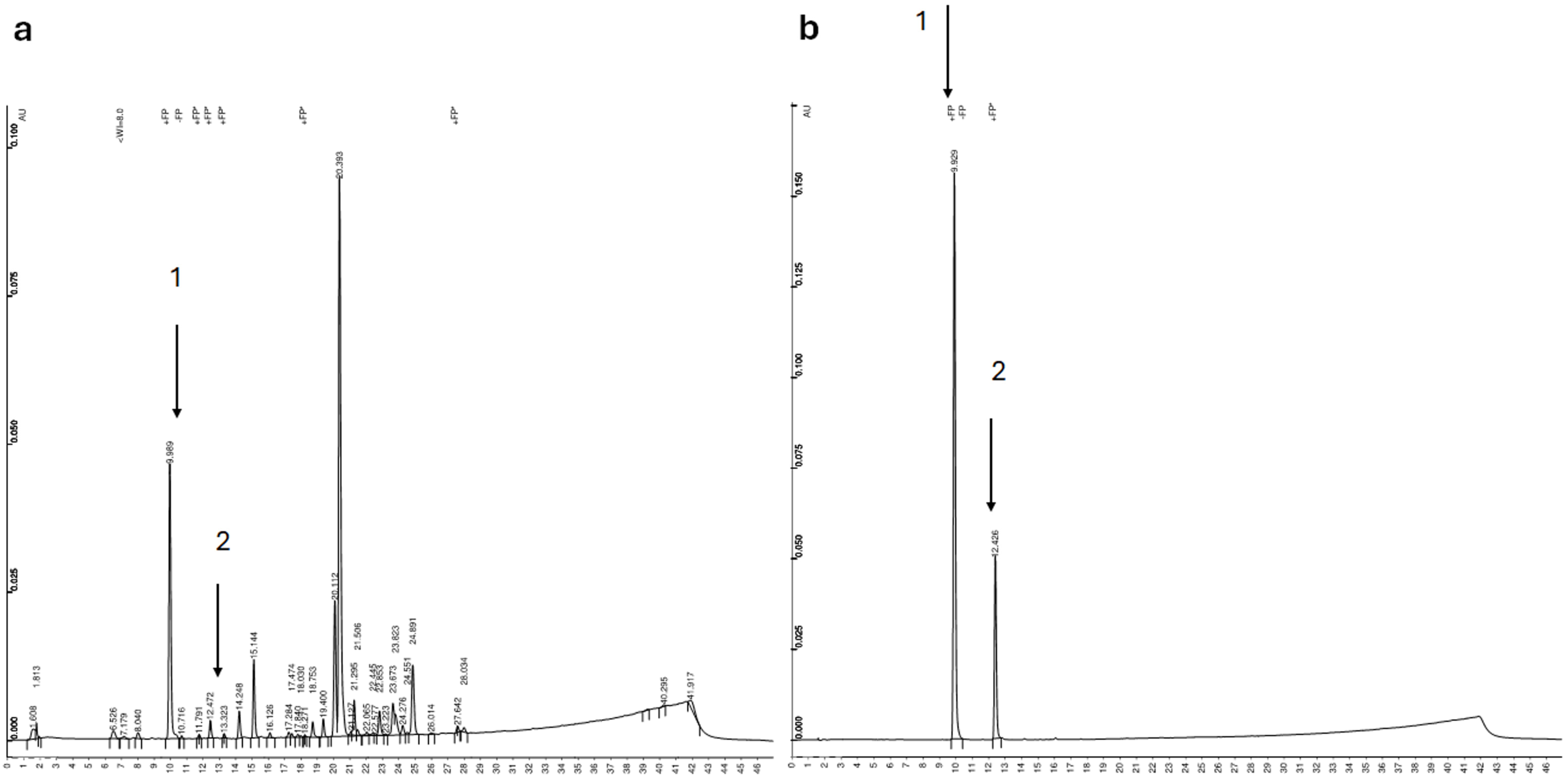
4.8. The Identification and Quantification of Chlorogenic Acid Using the HPLC-UV Technique (Chromatographic System, Conditions and Validation of the Method)
4.9. Statistics
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Rashmi, H.B.; Negi, P.S. Phenolic acids from vegetables: A review on processing stability and health benefits. Food Res. Int. 2020, 136, 109298. [Google Scholar] [CrossRef] [PubMed]
- Heleno, S.A.; Martins, A.; Queiroz, M.J.R.; Ferreira, I.C. Bioactivity of phenolic acids: Metabolites versus parent compounds: A review. Food Chem. 2015, 173, 501–513. [Google Scholar] [CrossRef] [PubMed]
- Kremr, D.; Bajer, T.; Bajerová, P.; Surmová, S.; Ventura, K. Unremitting problems with chlorogenic acid nomenclature: A review. Química Nova. 2016, 39, 530–533. [Google Scholar] [CrossRef]
- e Silva, N.V.; Mazzafera, P.; Cesarino, I. Should I stay or should I go: Are chlorogenic acids mobilized towards lignin biosynthesis? Phytochemistry 2019, 166, 112063. [Google Scholar] [CrossRef] [PubMed]
- Cha, M.N.; Kim, H.J.; Kim, B.G.; Ahn, J.H. Synthesis of chlorogenic acid and p-coumaroyl shikimates from glucose using engineered Escherichia coli. J. Microbiol. Biotechnol. 2014, 24, 1109–1117. [Google Scholar] [CrossRef] [PubMed]
- Naveed, M.; Hejazi, V.; Abbas, M.; Kamboh, A.A.; Khan, G.J.; Shumzaid, M.; Ahmad, F.; Babazadeh, D.; FangFang, X.; Modarresi-Ghazani, F.; et al. Chlorogenic acid (CGA): A pharmacological review and call for further research. Biomed. Pharmacother. 2018, 97, 67–74. [Google Scholar] [CrossRef] [PubMed]
- Shafaghatlonbar, M.; Bagherzade, G. Dual role of chlorogenic acid as an influential precursor in synthesizing nano-sized Cu (II) complexes and investigating its catalytic role in the oxidation of alcohols and its antibacterial activity. J. Organomet. Chem. 2023, 996, 122758. [Google Scholar] [CrossRef]
- Zeng, L.; Xiang, R.; Fu, C.; Qu, Z.; Liu, C. The Regulatory effect of chlorogenic acid on gut-brain function and its mechanism: A systematic review. Biomed. Pharmacother. 2022, 149, 112831. [Google Scholar] [CrossRef]
- Shang, H.; Liu, D.; Ji, H.J. The antioxidation activity in vivo and in vitro of flavonoids and chlorogenic acid isolated from Eucommia ulmoides leaves. Adv. Mater. Res. 2012, 550, 1270–1273. [Google Scholar] [CrossRef]
- Kabir, F.; Katayama, S.; Tanji, N.; Nakamura, S. Antimicrobial effects of chlorogenic acid and related compounds. J. Korean Soc. Appl. Biol. Chem. 2014, 57, 359–365. [Google Scholar] [CrossRef]
- Gonçalves, B.; Moeenfard, M.; Rocha, F.; Alves, A.; Estevinho, B.N.; Santos, L. Microencapsulation of a natural antioxidant from coffee—Chlorogenic acid (3-caffeoylquinic acid). Food Bioprocess Technol. 2017, 10, 1521–1530. [Google Scholar] [CrossRef]
- Rostami, F.; Radjabian, T.; Abrishamchi, P. Enhancement of phenolic acids accumulation in Salvia abrotanoides (Kar.) Sytsma shoot cultures under elicitation with nitric oxide. Plant Cell Tissue Organ Cult. (PCTOC) 2022, 149, 441–453. [Google Scholar] [CrossRef]
- Halder, M.; Sarkar, S.; Jha, S. Elicitation: A biotechnological tool for enhanced production of secondary metabolites in hairy root cultures. Eng. Life Sci. 2019, 19, 880–895. [Google Scholar] [CrossRef] [PubMed]
- Khurshid, R.; Ullah, M.A.; Tungmunnithum, D.; Drouet, S.; Shah, M.; Zaeem, A.; Hameed, S.; Hano, C.; Abbasi, B.H. Lights triggered differential accumulation of antioxidant and antidiabetic secondary metabolites in callus culture of Eclipta alba L. PLoS ONE 2020, 15, e0233963. [Google Scholar] [CrossRef] [PubMed]
- Castro, A.H.F.; Coimbra, M.C.; Crema, C.R.N.; de Oliveira Thomasi, R.M.; Cardoso, V.R. Elicitors as a biotechnological tool for in vitro production of bioactive phenolic compounds. In Exploring Plant Cells for the Production of Compounds of Interest; Springer: Berlin/Heidelberg, Germany, 2021; pp. 195–226. [Google Scholar] [CrossRef]
- Mohammad, S.; Khan, M.A.; Ali, A.; Khan, L.; Khan, M.S. Feasible production of biomass and natural antioxidants through callus cultures in response to varying light intensities in olive (Olea europaea. L.) cult. Arbosana. J. Photochem. Photobiol. B Biol. 2019, 193, 140–147. [Google Scholar] [CrossRef] [PubMed]
- Le, A.T.; Yu, J.K.; Han, G.D.; Do, T.K.; Chung, Y.S. Potential Use of Colored LED Lights to Increase the Production of Bioactive Metabolites Hedyotis corymbosa (L.) Lam. Plants 2022, 11, 225. [Google Scholar] [CrossRef] [PubMed]
- Rai, N.; Kumari, S.; Singh, S.; Saha, P.; Pandey, A.K.; Pandey-Rai, S. Modulation of morpho-physiological attributes and in situ analysis of secondary metabolites using Raman spectroscopy in response to red and blue light exposure in Artemisia annua. Environ. Exp. Bot. 2024, 217, 105563. [Google Scholar] [CrossRef]
- Jan, R.; Asaf, S.; Numan, M.; Lubna Kim, K.M. Plant secondary metabolite biosynthesis and transcriptional regulation in response to biotic and abiotic stress conditions. Agronomy 2021, 11, 968. [Google Scholar] [CrossRef]
- Fürtauer, L.; Weiszmann, J.; Weckwerth, W.; Nägele, T. Dynamics of plant metabolism during cold acclimation. Int. J. Mol. Sci. 2019, 20, 5411. [Google Scholar] [CrossRef]
- Ambrožič-Dolinšek, J.; Podgrajšek, A.; Šabeder, N.; Grudnik, Z.M.; Urbanek Krajnc, A.; Todorović, B.; Ciringer, T. The potential of berula erecta in vitro for As bioaccumulation and phytoremediation of water environments. Environ. Pollut. Bioavailab. 2023, 35, 2205010. [Google Scholar] [CrossRef]
- Mechora, Š.; Rižnik, T.; Urbanek Krajnc, A.; Ambrožič-Dolinšek, J. Response of Berula erecta to Lead in Combination with Selenium. Bull. Environ. Contam. Toxicol. 2020, 105, 51–61. [Google Scholar] [CrossRef] [PubMed]
- Thiviya, P.; Gamage, A.; Piumali, D.; Merah, O.; Madhujith, T. Apiaceae as an important source of antioxidants and their applications. Cosmetics 2021, 8, 111. [Google Scholar] [CrossRef]
- Javidnia, K.; Miri, R.; Assadollahi, M. Constituents of the essential oil of Berula angustifolia from Iran. Chem. Nat. Compd. 2011, 46, 990–991. [Google Scholar] [CrossRef]
- Tabaraki, R.; Nateghi, A.; Ahmady-Asbchin, S. In vitro assessment of antioxidant and antibacterial activities of six edible plants from Iran. J. Acupunct. Meridian Stud. 2013, 6, 159–162. [Google Scholar] [CrossRef]
- Murashige, T.; Skoog, F. A revised medium for rapid growth and bio assays with tobacco tissue cultures. Physiol. Plant. 1962, 15, 473–497. [Google Scholar] [CrossRef]
- Park, J.S.; Seong, Z.K.; Kim, M.S.; Ha, J.H.; Moon, K.B.; Lee, H.J.; Lee, H.K.; Jeon, J.H.; Park, S.U.; Kim, H.S. Production of flavonoids in callus cultures of Sophora flavescens Aiton. Plants 2020, 9, 688. [Google Scholar] [CrossRef]
- Dias, M.I.; Sousa, M.J.; Alves, R.C.; Ferreira, I.C. Exploring plant tissue culture to improve the production of phenolic compounds: A review. Ind. Crops Prod. 2016, 82, 9–22. [Google Scholar] [CrossRef]
- Palta, J.P. Leaf chlorophyll content. Remote Sens. Rev. 1990, 5, 207–213. [Google Scholar] [CrossRef]
- Kancheva, R.; Borisova, D.; Georgiev, G. Chlorophyll assessment and stress detection from vegetation optical properties. Ecol. Eng. Environ. Prot. 2014, 1, 34–43. [Google Scholar]
- Heaton, J.W.; Marangoni, A.G. Chlorophyll degradation in processed foods and senescent plant tissues. Trends Food Sci. Technol. 1996, 7, 8–15. [Google Scholar] [CrossRef]
- Ambrožič-Dolinšek, J.; Oblonšek, S.; Ciringer, T. Slow Growth Storage of Berula erecta in vitro–Effect of Sucrose, Sorbitol and Temperature. Agric. Sci. 2022, 19, 33–41. [Google Scholar] [CrossRef]
- Ma, Y.; Li, X.; Gu, Z.; Li, J.A. Leaf color and growth change of Sedum rubrotinctum caused by two commercial chemical products. HortScience 2019, 54, 434–444. [Google Scholar] [CrossRef]
- Su, X.; Griffin, J.; Xu, J.; Ouyang, P.; Zhao, Z.; Wang, W. Identification and quantification of anthocyanins in purple-fleshed sweet potato leaves. Heliyon 2019, 5, e01964. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Zhai, J.; Shao, L.; Lin, W.; Peng, C. Accumulation of anthocyanins: An adaptation strategy of Mikania micrantha to low temperature in winter. Front. Plant Sci. 2019, 10, 1049. [Google Scholar] [CrossRef]
- Samkumar, A.; Jones, D.; Karppinen, K.; Dare, A.P.; Sipari, N.; Espley, R.V.; Jaakola, L. Red and blue light treatments of ripening bilberry fruits reveal differences in signalling through abscisic acid-regulated anthocyanin biosynthesis. Plant Cell Environ. 2021, 44, 3227–3245. [Google Scholar] [CrossRef]
- Tao, R.; Bai, S.; Ni, J.; Yang, Q.; Zhao, Y.; Teng, Y. The blue light signal transduction pathway is involved in anthocyanin accumulation in ‘Red Zaosu’ pear. Planta 2018, 248, 37–48. [Google Scholar] [CrossRef] [PubMed]
- Mattioli, R.; Francioso, A.; Mosca, L.; Silva, P. Anthocyanins: A comprehensive review of their chemical properties and health effects on cardiovascular and neurodegenerative diseases. Molecules 2020, 25, 3809. [Google Scholar] [CrossRef]
- Izzo, L.G.; Mele, B.H.; Vitale, L.; Vitale, E.; Arena, C. The role of monochromatic red and blue light in tomato early photomorphogenesis and photosynthetic traits. Environ. Exp. Bot. 2020, 179, 104195. [Google Scholar] [CrossRef]
- Kim, H.M.; Hwang, S.J. The growth and development of ‘mini chal’ tomato plug seedlings grown under various wavelengths using light emitting diodes. Agronomy 2019, 9, 157. [Google Scholar] [CrossRef]
- Wang, J.; Lu, W.; Tong, Y.; Yang, Q. Leaf morphology, photosynthetic performance, chlorophyll fluorescence, stomatal development of lettuce (Lactuca sativa L.) exposed to different ratios of red light to blue light. Front. Plant Sci. 2016, 7, 250. [Google Scholar] [CrossRef]
- Mengxi, L.; Zhigang, X.; Yang, Y.; Yijie, F. Effects of different spectral lights on Oncidium PLBs induction, proliferation, and plant regeneration. Plant Cell Tissue Organ Cult (PCTOC) 2011, 106, 1–10. [Google Scholar] [CrossRef]
- Al-Rukabi, M.N.; Leunov, V.I.; Tarakanov, I.G.; Tereshonkova, T.A. The Effect of LED Lighting on The Growth of Seedlings of Hybrid Tomato. In IOP Conference Series: Earth and Environmental Science; IOP Publishing: Bristol, UK, 2021; Volume 910, p. 012127. [Google Scholar] [CrossRef]
- Afridi, M.; Fayyaz, M.; Rehman, M.; Akhter, N.; Masood, S.; Khan, S.; Ahmad, K. Effect of light emitting diode (LED) on the growth and photosynthetic characteristics of Brassica Juncea. J. Bio-Mol. Sci. (JBMS) 2020, 7, 1–9. [Google Scholar]
- Janas, K.M.; Cvikrová, M.; Pałagiewicz, A.; Szafranska, K.; Posmyk, M.M. Constitutive elevated accumulation of phenylpropanoids in soybean roots at low temperature. Plant Sci. 2002, 163, 369–373. [Google Scholar] [CrossRef]
- Heitman, E.; Ingram, D.K. Cognitive and neuroprotective effects of chlorogenic acid. Nutr. Neurosci. 2017, 20, 32–39. [Google Scholar] [CrossRef]
- Wu, C.H.; Murthy, H.N.; Hahn, E.J.; Paek, K.Y. Large-scale cultivation of adventitious roots of Echinacea purpurea in airlift bioreactors for the production of chichoric acid, chlorogenic acid and caftaric acid. Biotechnol. Lett. 2007, 29, 1179–1182. [Google Scholar] [CrossRef] [PubMed]
- Siahpoush, A.; Ghasemi, N.; Ardakani, M.S.; Asghari, G. Production of chlorogenic acid in Varthemia persica DC (var. persica) callus cultures. Res. Pharm. Sci. 2011, 6, 63. [Google Scholar] [PubMed]
- Thiem, B.; Kikowska, M.; Krawczyk, A.; Więckowska, B.; Sliwinska, E. Phenolic acid and DNA contents of micropropagated Eryngium planum L. Plant Cell Tissue Organ Cult. (PCTOC) 2013, 114, 197–206. [Google Scholar] [CrossRef]
- Szopa, A.; Ekiert, H. In vitro cultures of Schisandra chinensis (Turcz.) Baill. (Chinese magnolia vine)—A potential biotechnological rich source of therapeutically important phenolic acids. Appl. Biochem. Biotechnol. 2012, 166, 1941–1948. [Google Scholar] [CrossRef] [PubMed]
- Trajtemberg, S.P.; Apóstolo, N.M.; Fernández, G. Calluses of Cynara cardunculus var. cardunculus cardoon (Asteraceae): Determination of cynarine and chlorogenic acid by automated high-performance capillary electrophoresis. Vitr. Cell. Dev. Biol.-Plant 2006, 42, 534–537. [Google Scholar] [CrossRef]
- Chen, C.C.; Agrawal, D.C.; Lee, M.R.; Lee, R.J.; Kuo, C.L.; Wu, C.R.; Chang, H.C. Influence of LED light spectra on in vitro somatic embryogenesis and LC–MS analysis of chlorogenic acid and rutin in Peucedanum japonicum Thunb.: A medicinal herb. Bot. Stud. 2016, 57, 9. [Google Scholar] [CrossRef]
- Matkowski, A. Plant in vitro culture for the production of antioxidants—A review. Biotechnol. Adv. 2008, 26, 548–560. [Google Scholar] [CrossRef] [PubMed]
- Murthy, H.N.; Praveen, N. Influence of macro elements and nitrogen source on adventitious root growth and withanolide-A production in Withania somnifera (L.) Dunal. Nat. Prod. Res. 2012, 26, 466–473. [Google Scholar] [CrossRef] [PubMed]
- Pałka, P.; Muszyńska, B.; Szewczyk, A.; Pawłowska, B. Elicitation and Enhancement of Phenolics Synthesis with Zinc Oxide Nanoparticles and LED Light in Lilium candidum L. Cultures In Vitro. Agronomy 2023, 13, 1437. [Google Scholar] [CrossRef]
- Kikowska, M.; Budzianowski, J.; Krawczyk, A.; Thiem, B. Accumulation of rosmarinic, chlorogenic and caffeic acids in in vitro cultures of Eryngium planum L. Acta Physiol. Plant. 2012, 34, 2425–2433. [Google Scholar] [CrossRef]
- Liu, Z.B.; Chen, J.G.; Yin, Z.P.; Shangguan, X.C.; Peng, D.Y.; Lu, T.; Lin, P. Methyl jasmonate and salicylic acid elicitation increase content and yield of chlorogenic acid and its derivatives in Gardenia jasminoides cell suspension cultures. Plant Cell Tissue Organ Cult. (PCTOC) 2018, 134, 79–93. [Google Scholar] [CrossRef]
- Kuzel, S.; Vydra, J.A.N.; Triska, J.A.N.; Vrchotova, N.; Hruby, M.; Cigler, P. Elicitation of pharmacologically active substances in an intact medical plant. J. Agric. Food Chem. 2009, 57, 7907–7911. [Google Scholar] [CrossRef]
- Koch, U.; Kühnl, T.; Conradt, W.; Wellmann, E. Differential effects of light and fungal elicitor on chlorogenic acid and caffeoylshikimic acid metabolism. Plant Sci. 1990, 70, 167–174. [Google Scholar] [CrossRef]
- Das, S.; Sultana, K.W.; Mondal, M.; Chandra, I.; Ndhlala, A.R. Unveiling the Dual Nature of Heavy Metals: Stressors and Promoters of Phenolic Compound Biosynthesis in Basilicum polystachyon (L.) Moench In Vitro. Plants 2023, 13, 98. [Google Scholar] [CrossRef] [PubMed]
- Mechora, Š.; Sotler, M.; Urbanek Krajnc, A.; Ambrožič-Dolinšek, J. How selenium affects Berula erecta. Water Air Soil Pollut. 2016, 227, 1–12. [Google Scholar] [CrossRef]
- Wang, X.; Qin, L.; Zhou, J.; Li, Y.; Fan, X. A novel design to screen chlorogenic acid-producing microbial strains from the environment. Sci. Rep. 2018, 8, 14756. [Google Scholar] [CrossRef]
- Saleh, N.A.; El-Negoumy, S.I.; El-Hadidi, M.N.; Hosni, H.A. Comparative study of the flavonoids of some local members of the Umbelliferae. Phytochemistry 1983, 22, 1417–1420. [Google Scholar] [CrossRef]
- Park, Y.H.; Han, S.I.; Oh, B.; Kim, H.S.; Jeon, M.S.; Kim, S.; Choi, Y.E. Microalgal secondary metabolite productions as a component of biorefinery: A review. Bioresour. Technol. 2022, 344, 126206. [Google Scholar] [CrossRef] [PubMed]
- Wianowska, D.; Gil, M. Recent advances in extraction and analysis procedures of natural chlorogenic acids. Phytochem. Rev. 2019, 18, 273–302. [Google Scholar] [CrossRef]
- Dirar, A.I.; Alsaadi, D.H.M.; Wada, M.; Mohamed, M.A.; Watanabe, T.; Devkota, H.P. Effects of extraction solvents on total phenolic and flavonoid contents and biological activities of extracts from Sudanese medicinal plants. S. Afr. J. Bot. 2019, 120, 261–267. [Google Scholar] [CrossRef]
- Todorović, B.; Grujić, V.J.; Urbanek Krajnc, A.; Kranvogl, R.; Ambrožič-Dolinšek, J. Identification and content of astaxanthin and its esters from microalgae Haematococcus pluvialis by HPLC-DAD and LC-QTOF-MS after extraction with various solvents. Plants 2021, 10, 2413. [Google Scholar] [CrossRef] [PubMed]
- Ivanović, M.; Islamčević Razboršek, M.; Kolar, M. Innovative extraction techniques using deep eutectic solvents and analytical methods for the isolation and characterization of natural bioactive compounds from plant material. Plants 2020, 9, 1428. [Google Scholar] [CrossRef]
- Kubica, P.; Szopa, A.; Prokopiuk, B.; Komsta, Ł.; Pawłowska, B.; Ekiert, H. The influence of light quality on the production of bioactive metabolites–verbascoside, isoverbascoside and phenolic acids and the content of photosynthetic pigments in biomass of Verbena officinalis L. cultured in vitro. J. Photochem. Photobiol. B Biol. 2020, 203, 111768. [Google Scholar] [CrossRef]
- Kim, E.Y.; Park, S.A.; Park, B.J.; Lee, Y.; Oh, M.M. Growth and antioxidant phenolic compounds in cherry tomato seedlings grown under monochromatic light-emitting diodes. Hortic. Environ. Biotechnol. 2014, 55, 506–513. [Google Scholar] [CrossRef]
- Biswal, B.; Jena, B.; Giri, A.K.; Acharya, L. Monochromatic light elicited biomass accumulation, antioxidant activity, and secondary metabolite production in callus culture of Operculina turpethum (L.). Plant Cell Tissue Organ Cult. (PCTOC) 2022, 149, 123–134. [Google Scholar] [CrossRef]
- Szopa, A.; Ekiert, H. The importance of applied light quality on the production of lignans and phenolic acids in Schisandra chinensis (Turcz.) Baill. cultures in vitro. Plant Cell Tissue Organ Cult. (PCTOC) 2016, 127, 115–121. [Google Scholar] [CrossRef]
- Lefsrud, M.G.; Kopsell, D.A.; Kopsell, D.E.; Curran-Celentano, J. Air temperature affects biomass and carotenoid pigment accumulation in kale and spinach grown in a controlled environment. HortScience 2005, 40, 2026–2030. [Google Scholar] [CrossRef]
- Naikoo, M.I.; Dar, M.I.; Raghib, F.; Jaleel, H.; Ahmad, B.; Raina, A.; Naushin, F. Role and regulation of plants phenolics in abiotic stress tolerance: An overview. In Plant Signaling Molecules: Role and Regulation under Stressful Environments; Woodhead Publishing: Sawston, UK, 2019; pp. 157–168. [Google Scholar] [CrossRef]
- Lattanzio, V.; Cardinali, A.; Ruta, C.; Fortunato, I.M.; Lattanzio, V.M.; Linsalata, V.; Cicco, N. Relationship of secondary metabolism to growth in oregano (Origanum vulgare L.) shoot cultures under nutritional stress. Environ. Exp. Bot. 2009, 65, 54–62. [Google Scholar] [CrossRef]
- Caretto, S.; Linsalata, V.; Colella, G.; Mita, G.; Lattanzio, V. Carbon fluxes be-tween primary metabolism and phenolic pathway in plant tissues under stress. Int. J. Mol. Sci. 2015, 16, 26378–26394. [Google Scholar] [CrossRef] [PubMed]
- García-Pérez, P.; Lozano-Milo, E.; Landín, M.; Gallego, P.P. Combining medicinal plant in vitro culture with machine learning technologies for maximizing the production of phenolic compounds. Antioxidants 2020, 9, 210. [Google Scholar] [CrossRef] [PubMed]
- Jaafar, H.Z.; Ibrahim, M.H.; Karimi, E. Phenolics and flavonoids compounds phenylalanine ammonia lyase and antioxidant activity responses to elevated CO2 in Labisia pumila (Myrisinaceae). Molecules 2012, 17, 6331–6347. [Google Scholar] [CrossRef]
- Zhou, Z.; Chen, X.; Zhang, M.; Blanchard, C. Phenolics, flavonoids, proanthocyanidin and antioxidant activity of brown rice with different pericarp colors following storage. J. Stored Prod. Res. 2014, 59, 120–125. [Google Scholar] [CrossRef]
- Sei, G.; Ella, F.M.A.; Emad, M.H.; Shalaby, E.; Doha, H. Antioxidant activity of phenolic compounds from different grape wastes. J. Food Process. Technol. 2014, 5, 2. [Google Scholar] [CrossRef]
- Liu, Y.; Liu, X.; Zhong, F.; Tian, R.; Zhang, K.; Zhang, X.; Li, T. Comparative study of phenolic compounds and antioxidant activity in different species of cherries. J. Food Sci. 2011, 76, C633–C638. [Google Scholar] [CrossRef] [PubMed]
- Swigonska, S.; Amarowicz, R.; Król, A.; Mostek, A.; Badowiec, A.; Weidner, S. Influence of abiotic stress during soybean germination followed by recovery on the phenolic compounds of radicles and their antioxidant capacity. Acta Soc. Bot. Pol. 2014, 83, 209–218. [Google Scholar] [CrossRef]
- Merah, O.; Sayed-Ahmad, B.; Talou, T.; Saad, Z.; Cerny, M.; Grivot, S.; Hijazi, A. Biochemical composition of cumin seeds, and biorefining study. Biomolecules 2020, 10, 1054. [Google Scholar] [CrossRef]
- Lazarević, J.; Radulović, N.; Palić, R.; Zlatković, B. Chemical analysis of volatile constituents of Berula erecta (Hudson) Coville subsp. erecta (Apiaceae) from Serbia. J. Essent. Oil Res. 2010, 22, 153–156. [Google Scholar] [CrossRef]
- Ngo, T.C.; Dao, D.Q.; Nguyen, M.T.; Nam, P.C. A DFT analysis on the radical scavenging activity of oxygenated terpenoids present in the extract of the buds of Cleistocalyx operculatus. RSC Adv. 2017, 7, 39686–39698. [Google Scholar] [CrossRef]
- Ayla, K.; Demirci, B.; Muhittin, D.; Süleyman, D. A monotypic species from Turkey: Characterization of the essential oil of Berula erecta (Apiaceae). Nat. Volatiles Essent. Oils 2018, 5, 7–10. [Google Scholar]
- Abd-ElGawad, A.; El Gendy, A.E.N.; El-Amier, Y.; Gaara, A.; Omer, E.; Al-Rowaily, S.; Elshamy, A. Essential oil of Bassia muricata: Chemical characterization, antioxidant activity, and allelopathic effect on the weed Chenopodium murale. Saudi J. Biol. Sci. 2020, 27, 1900–1906. [Google Scholar] [CrossRef]
- Muanda, F.; Koné, D.; Dicko, A.; Soulimani, R.; Younos, C. Phytochemical composition and antioxidant capacity of three malalianian medicinal plant parts. Evid.-Based Complement. Altern. Med. 2011, 2011, 674320. [Google Scholar] [CrossRef]
- Everette, J.D.; Bryant, Q.M.; Green, A.M.; Abbey, Y.A.; Wangila, G.W.; Walker, R.B. Thorough study of reactivity of various compound classes toward the Folin−Ciocalteu reagent. J. Agric. Food Chem. 2010, 58, 8139–8144. [Google Scholar] [CrossRef] [PubMed]
- Yoo, K.S.; Lee, E.J.; Leskovar, D.; Patil, B.S. Development of an automated method for Folin-Ciocalteu total phenolic assay in artichoke extracts. J. Food Sci. 2012, 77, C1279–C1284. [Google Scholar] [CrossRef]
- Cilliers, J.J.L.; Singleton, V.L.; Lamuela-Raventos, R.M. Total polyphenols in apples and ciders; correlation with chlorogenic acid. J. Food Sci. 1990, 55, 1458–1459. [Google Scholar] [CrossRef]
- Jeszka-Skowron, M.; Stanisz, E.; De Peña, M.P. Relationship between antioxidant capacity, chlorogenic acids and elemental composition of green coffee. Lwt 2016, 73, 243–250. [Google Scholar] [CrossRef]
- Anh-Dao, L.T.; Minh-Huy, D.; Bui-Phuc, T.; Nhut-Truong, N.; Tu-Chi, T.; Thi-Hoa, L.; Cong-Hau, N. Determination of Caffeine, Chlorogenic Acid, Total Phenolic Contents, and Antioxidant Capacities for Arabica and Robusta Coffee from Vietnam. Anal. Lett. 2024, 1–15. [Google Scholar] [CrossRef]
- Joly, N.; Souidi, K.; Depraetere, D.; Wils, D.; Martin, P. Potato by-products as a source of natural chlorogenic acids and phenolic compounds: Extraction, characterization, and antioxidant capacity. Molecules 2020, 26, 177. [Google Scholar] [CrossRef]
- Šamec, D.; Karalija, E.; Šola, I.; Vujčić Bok, V.; Salopek-Sondi, B. The role of polyphenols in abiotic stress response: The influence of molecular structure. Plants 2021, 10, 118. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.; Bertin, R.; Froldi, G. EC50 estimation of antioxidant activity in DPPH assay using several statistical programs. Food Chem. 2013, 138, 414–420. [Google Scholar] [CrossRef] [PubMed]
- Pitakpawasutthi, Y.; Thitikornpong, W.; Palanuvej, C.; Ruangrungsi, N. Chlorogenic acid content, essential oil compositions, and in vitro antioxidant activities of Chromolaena odorata leaves. J. Adv. Pharm. Technol. Res. 2016, 7, 37–42. [Google Scholar] [CrossRef]
- Karuppusamy, S. A review on trends in production of secondary metabolites from higher plants by in vitro tissue, organ and cell cultures. J. Med. Plants Res. 2009, 3, 1222–1239. [Google Scholar]
- Gonçalves, S.; Romano, A. Production of plant secondary metabolites by using biotechnological tools. Second. Metab.-Sources Appl. 2018, 5, 81–99. [Google Scholar] [CrossRef]
- Nicasio-Torres, M.D.P.; Meckes-Fischer, M.; Aguilar-Santamaría, L.; Garduño-Ramírez, M.L.; Chávez-Ávila, V.M.; Cruz-Sosa, F. Production of chlorogenic acid and isoorientin hypoglycemic compounds in Cecropia obtusifolia calli and in cell suspension cultures with nitrate deficiency. Acta Physiol. Plant. 2012, 34, 307–316. [Google Scholar] [CrossRef]
- Fu, X.; Yin, Z.P.; Chen, J.G.; Shangguan, X.C.; Wang, X.; Zhang, Q.F.; Peng, D.Y. Production of chlorogenic acid and its derivatives in hairy root cultures of Stevia rebaudiana. J. Agric. Food Chem. 2015, 63, 262–268. [Google Scholar] [CrossRef] [PubMed]
- Sitarek, P.; Kowalczyk, T.; Picot, L.; Michalska-Hejduk, D.; Bijak, M.; Białas, A.J.; Skała, E. Growth of Leonurus sibiricus L. roots with over-expression of AtPAP1 transcriptional factor in closed bioreactor, production of bioactive phenolic compounds and evaluation of their biological activity. Ind. Crops Prod. 2018, 122, 732–739. [Google Scholar] [CrossRef]
- Nartop, P. Engineering of biomass accumulation and secondary metabolite production in plant cell and tissue cultures. In Plant Metabolites and Regulation under Environmental Stress; Academic Press: Cambridge, MA, USA, 2018; pp. 169–194. [Google Scholar] [CrossRef]
- Ozyigit, I.I.; Dogan, I.; Hocaoglu-Ozyigit, A.; Yalcin, B.; Erdogan, A.; Yalcin, I.E.; Kaya, Y. Production of secondary metabolites using tissue culture-based biotechnological applications. Front. Plant Sci. 2023, 14, 1132555. [Google Scholar] [CrossRef]
- Cardoso, J.C.; Oliveira, M.E.B.D.; Cardoso, F.D.C. Advances and challenges on the in vitro production of secondary metabolites from medicinal plants. Hortic. Bras. 2019, 37, 124–132. [Google Scholar] [CrossRef]
- Xiao, F.; Lian, J.; Tu, S.; Xie, L.; Li, J.; Zhang, F.; Zhong, W. Metabolic engineering of Saccharomyces cerevisiae for high-level production of chlorogenic acid from glucose. ACS Synth. Biol. 2022, 11, 800–811. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Wang, H.; Chen, J.; Hu, M.; Shan, X.; Zhou, J. Efficient Production of Chlorogenic Acid in Escherichia coli Via Modular Pathway and Cofactor Engineering. J. Agric. Food Chem. 2023, 71, 15204–15212. [Google Scholar] [CrossRef] [PubMed]
- da Silveira, J.S.; Durand, N.; Lacour, S.; Belleville, M.P.; Perez, A.; Loiseau, G.; Dornier, M. Solid-state fermentation as a sustainable method for coffee pulp treatment and production of an extract rich in chlorogenic acids. Food Bioprod. Process. 2019, 115, 175–184. [Google Scholar] [CrossRef]
- Myo, H.; Nantarat, N.; Khat-Udomkiri, N. Changes in bioactive compounds of coffee pulp through fermentation-based biotransformation using Lactobacillus plantarum TISTR 543 and its antioxidant activities. Fermentation 2021, 7, 292. [Google Scholar] [CrossRef]
- Awwad, S.; Issa, R.; Alnsour, L.; Albals, D.; Al-Momani, I. Quantification of caffeine and chlorogenic acid in green and roasted coffee samples using HPLC-DAD and evaluation of the effect of degree of roasting on their levels. Molecules 2021, 26, 7502. [Google Scholar] [CrossRef] [PubMed]
- Gallardo-Ignacio, J.; Santibáñez, A.; Oropeza-Mariano, O.; Salazar, R.; Montiel-Ruiz, R.M.; Cabrera-Hilerio, S.; Nicasio-Torres, P. Chemical and Biological Characterization of Green and Processed Coffee Beans from Coffea arabica Varieties. Molecules 2023, 28, 4685. [Google Scholar] [CrossRef] [PubMed]
- Budryn, G.; Nebesny, E.; Podsędek, A.; Żyżelewicz, D.; Materska, M.; Jankowski, S.; Janda, B. Effect of different extraction methods on the recovery of chlorogenic acids, caffeine and Maillard reaction products in coffee beans. Eur. Food Res. Technol. 2009, 228, 913–922. [Google Scholar] [CrossRef]
- Mohammad Salamatullah, A.; Hayat, K.; Mabood Husain, F.; Asif Ahmed, M.; Arzoo, S.; Mohammed Alghunaymi, A.; Bourhia, M. Effect of microwave roasting and extraction solvents on the bioactive properties of coffee beans. Evid.-Based Complement. Altern. Med. 2021, 2021, 4908033. [Google Scholar] [CrossRef]
- Masek, A.; Latos-Brozio, M.; Kałużna-Czaplińska, J.; Rosiak, A.; Chrzescijanska, E. Antioxidant properties of green coffee extract. Forests 2020, 11, 557. [Google Scholar] [CrossRef]
- Ivanović, M.; Grujić, D.; Cerar, J.; Islamčević Razboršek, M.; Topalić-Trivunović, L.; Savić, A.; Kolar, M. Extraction of Bioactive Metabolites from Achillea millefolium L. with Choline Chloride Based Natural Deep Eutectic Solvents: A Study of the Antioxidant and Antimicrobial Activity. Antioxidants 2022, 11, 724. [Google Scholar] [CrossRef] [PubMed]
- Islamčević Razboršek, M. Stability studies on trans-rosmarinic acid and GC–MS analysis of its degradation product. J. Pharm. Biomed. Anal. 2011, 55, 1010–1016. [Google Scholar] [CrossRef] [PubMed]
- Stopinšek, M.; Islamčević Razboršek, M. Fenolne Spojine v Ekstraktih Slovenskih Čajev. 2019, pp. 1–6. Available online: https://www.dlib.si/details/URN:NBN:SI:doc-NN71DJLP (accessed on 17 January 2024).
- Islamčević Razboršek, M.; Ivanović, M.; Kolar, M. Validated stability-indicating GC-MS method for characterization of forced degradation products of trans-caffeic acid and trans-ferulic acid. Molecules 2021, 26, 2475. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment | Content of Chlorogenic Acid [mg/1 g DW] | Total Phenolic Content [mg GA/1 g DW] |
|---|---|---|
| The beginning of the experiment | 1.893 ± 0.020 e | 17.29 ± 0.11 e |
| 14 days, white light, 23 °C | 1.309 ± 0.002 f | 16.41 ± 0.35 f |
| 28 days, white light, 23 °C | 3.042 ± 0.019 b | 23.77 ± 0.66 b |
| 14 days, blue light, 12 °C | 2.604 ± 0.022 c | 21.30 ± 0.24 c |
| 28 days, blue light, 12 °C | 4.049 ± 0.021 a | 28.12 ± 0.38 a |
| 14 days, red light, 12 °C | 0.927 ± 0.014 g | 14.76 ± 0.28 g |
| 28 days, red light, 12 °C | 2.050 ± 0.012 d | 20.30 ± 0.13 d |
| Treatment | Antioxidant Activity [mmol Trolox/1 g DW] |
|---|---|
| The beginning of the experiment | 0.0485 ± 0.0002 d |
| 14 days, white light, 23 °C | 0.0469 ± 0.0012 d |
| 28 days, white light, 23 °C | 0.0687 ± 0.0006 b |
| 14 days, blue light, 12 °C | 0.0649 ± 0.0016 bc |
| 28 days, blue light, 12 °C | 0.1008 ± 0.0003 a |
| 14 days, red light, 12 °C | 0.0296 ± 0.0007 e |
| 28 days, red light, 12 °C | 0.0638 ± 0.0013 c |
| Spectrophotometric Assay | Chlorogenic Acid Content |
|---|---|
| TPC | 0.980 ** |
| Antioxidant activity (DPPH assay) | 0.963 ** |
| Treatment | Yield of Chlorogenic Acid Obtained in Tissue Culture of Berula erecta per Litre of Prepared MS Medium [mg/1 L MS Medium] |
|---|---|
| The beginning of the experiment | 20.846 ± 0.220 f |
| 14 days, white light, 23 °C | 29.787 ± 0.046 d |
| 28 days, white light, 23 °C | 126.874 ± 0.792 b |
| 14 days, blue light, 12 °C | 22.683 ± 0.192 e |
| 28 days, blue light, 12 °C | 168.873 ± 0.265 a |
| 14 days, red light, 12 °C | 9.402 ± 0.142 g |
| 28 days, red light, 12 °C | 32.520 ± 0.190 c |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Senekovič, J.; Ciringer, T.; Ambrožič-Dolinšek, J.; Islamčević Razboršek, M. The Effect of Combined Elicitation with Light and Temperature on the Chlorogenic Acid Content, Total Phenolic Content and Antioxidant Activity of Berula erecta in Tissue Culture. Plants 2024, 13, 1463. https://doi.org/10.3390/plants13111463
Senekovič J, Ciringer T, Ambrožič-Dolinšek J, Islamčević Razboršek M. The Effect of Combined Elicitation with Light and Temperature on the Chlorogenic Acid Content, Total Phenolic Content and Antioxidant Activity of Berula erecta in Tissue Culture. Plants. 2024; 13(11):1463. https://doi.org/10.3390/plants13111463
Chicago/Turabian StyleSenekovič, Jan, Terezija Ciringer, Jana Ambrožič-Dolinšek, and Maša Islamčević Razboršek. 2024. "The Effect of Combined Elicitation with Light and Temperature on the Chlorogenic Acid Content, Total Phenolic Content and Antioxidant Activity of Berula erecta in Tissue Culture" Plants 13, no. 11: 1463. https://doi.org/10.3390/plants13111463
APA StyleSenekovič, J., Ciringer, T., Ambrožič-Dolinšek, J., & Islamčević Razboršek, M. (2024). The Effect of Combined Elicitation with Light and Temperature on the Chlorogenic Acid Content, Total Phenolic Content and Antioxidant Activity of Berula erecta in Tissue Culture. Plants, 13(11), 1463. https://doi.org/10.3390/plants13111463