
Physiological Analysis of Source–Sink Relationship in Rice Genotypes with Contrasting Grain Yields

 ,
,
Abstract
:1. Introduction
2. Results
2.1. Yield and Yield Components
2.2. Source Traits
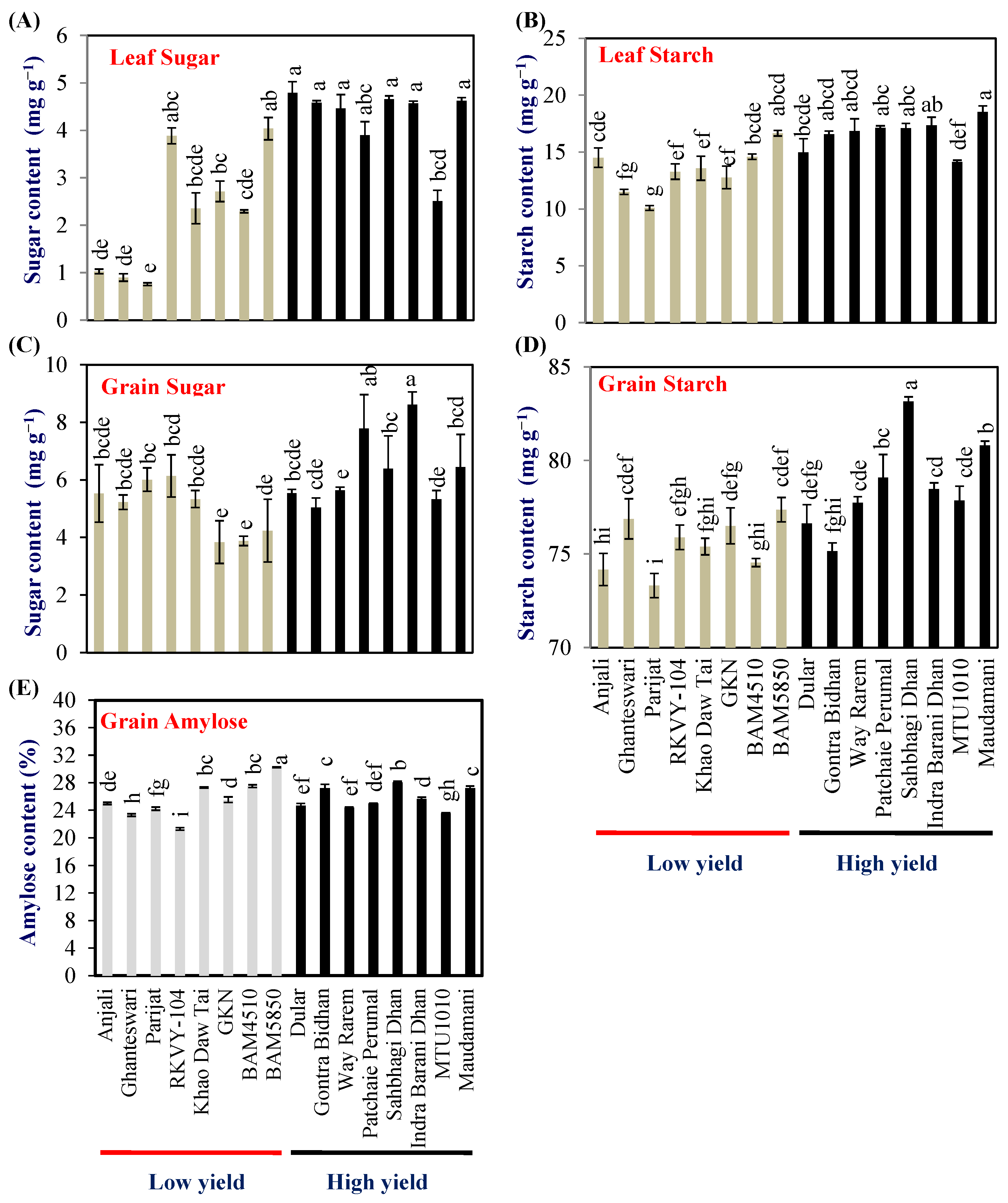
2.3. Leaf Sugar and Starch Content
2.4. Grain Sugar and Starch Content
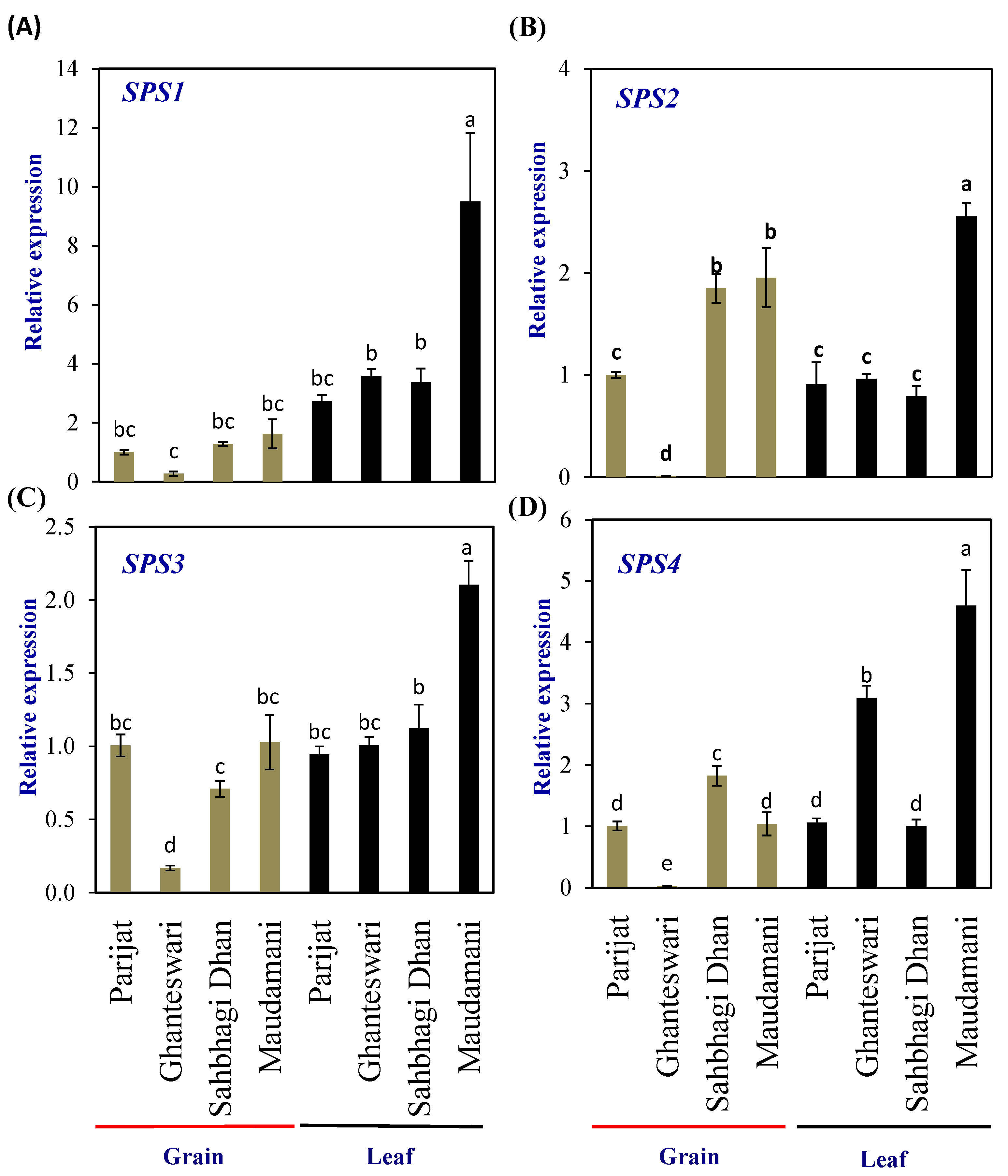
2.5. Expression Profiling of Photosynthesis Regulatory Genes
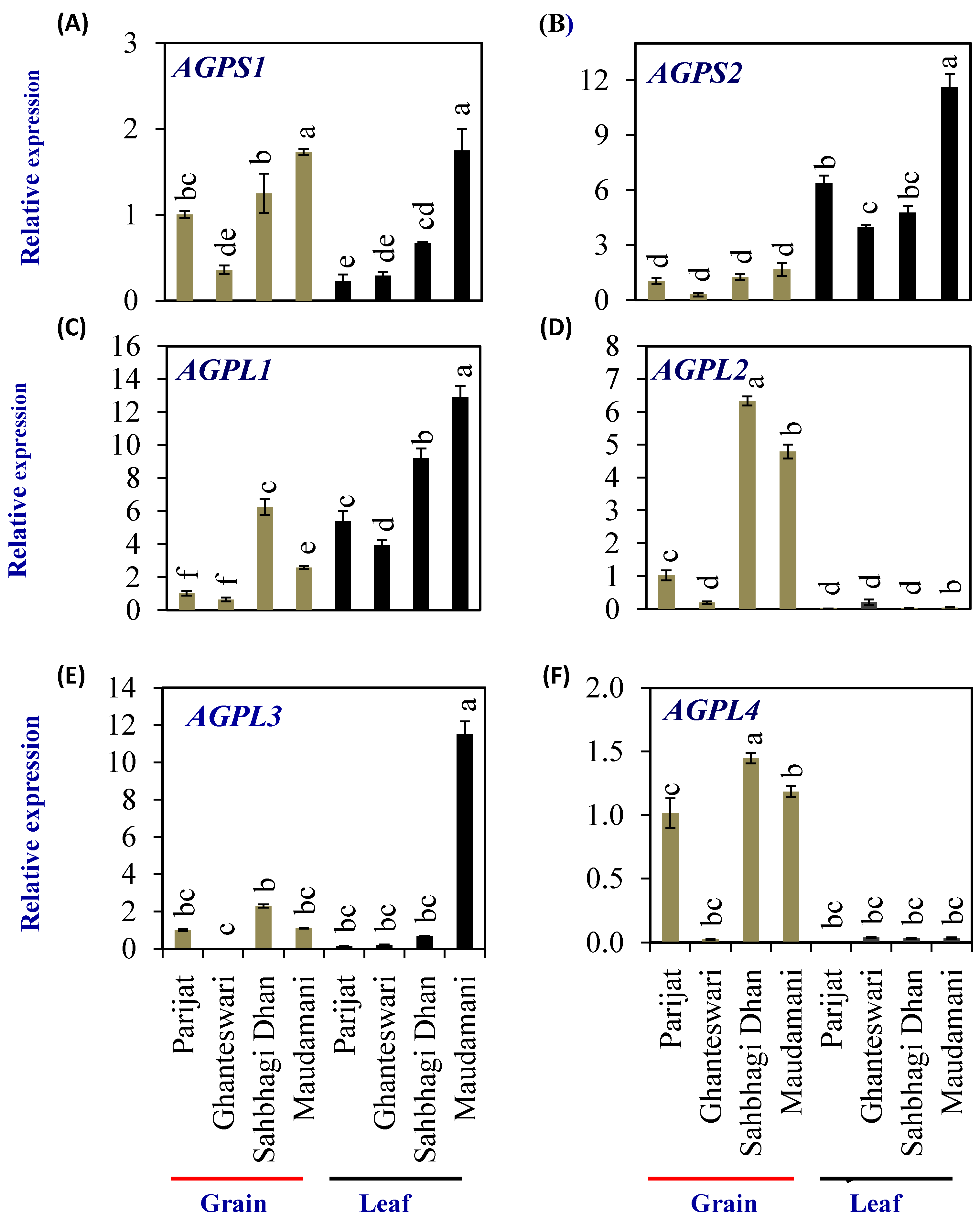
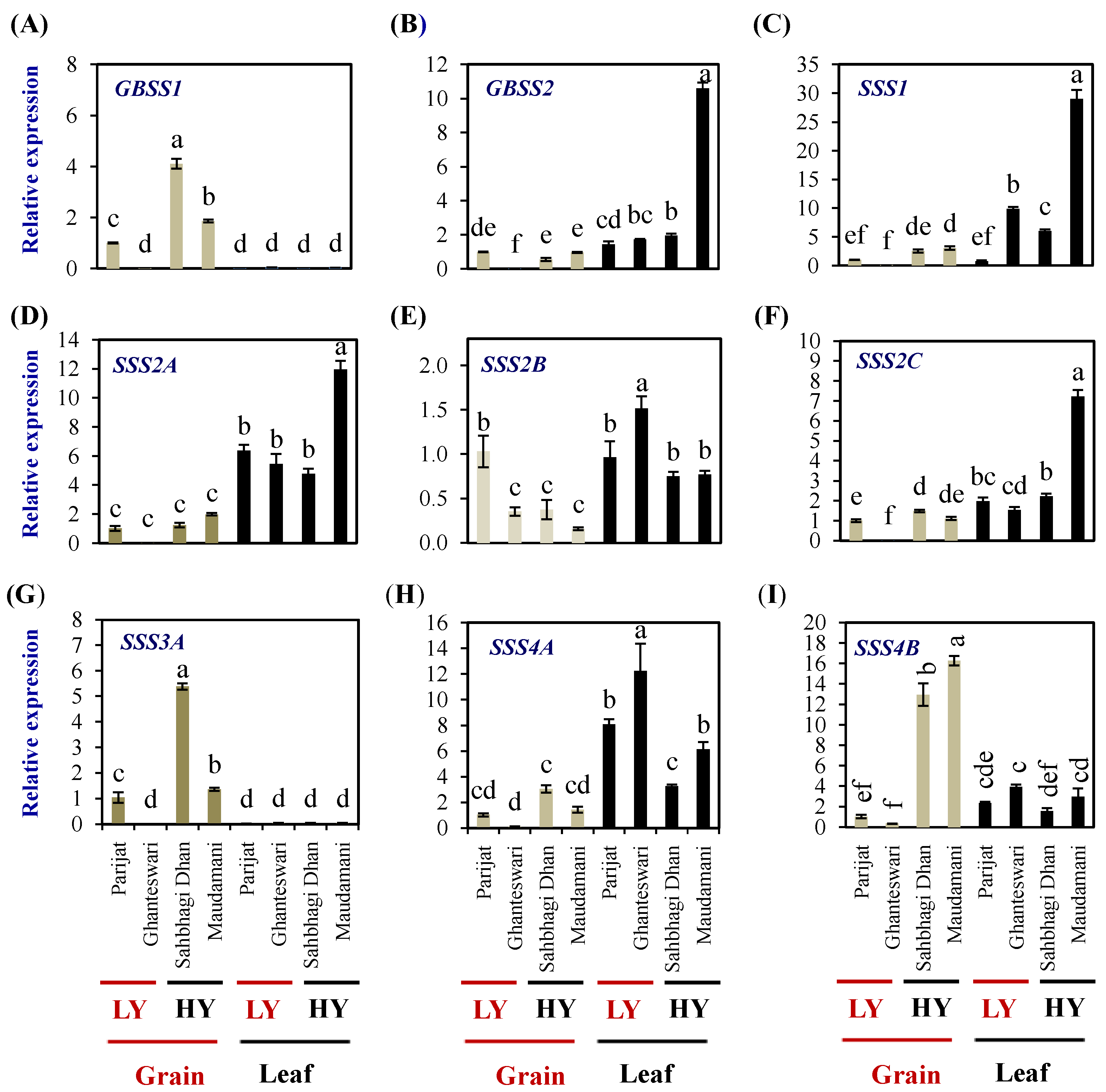
2.6. Expression Profiling of Starch-Synthesis Genes in Selected Genotypes
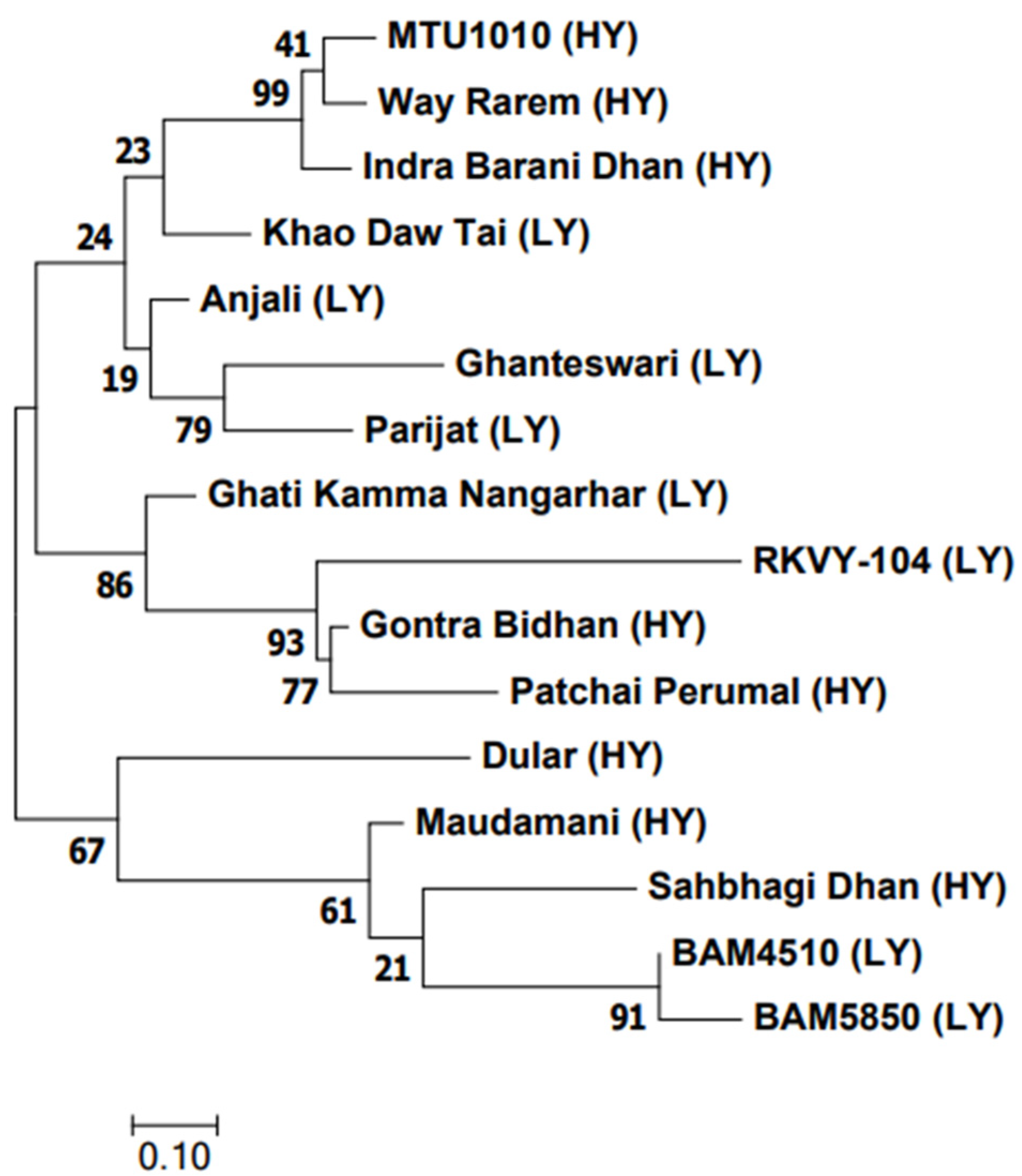
2.7. SNP-Based Allele Mining and Generation of Dendrogram
3. Discussion
4. Materials and Methods
4.1. Plant Material, Experimental Design, and Crop Management
4.2. Physiological Parameters
4.3. Biochemical Analysis
Estimation of Starch, Sugar, and Amylose Content
4.4. RNA Extraction and Gene-Expression Analysis
4.5. Allele Mining of Candidate Genes
4.6. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kurata, N.; Moore, G.; Nagamura, Y.; Foote, T.; Yano, M.; Minobe, Y.; Gale, M. Conservation of genome structure between rice and wheat. Bio/Technology 1994, 12, 276–278. [Google Scholar] [CrossRef]
- Garris, A.J.; Tai, T.H.; Coburn, J.; Kresovich, S.; McCouch, S. Genetic structure and diversity in Oryza sativa L. Genetics 2005, 169, 1631–1638. [Google Scholar] [CrossRef] [PubMed]
- Mogga, M.; Sibiya, J.; Shimelis, H.; Mbogo, D.; Muzhingi, T.; Lamo, J.; Yao, N. Correction Diversity analysis and genome-wide association studies of grain shape and eating quality traits in rice (Oryza sativa L.) using DArT markers. PLoS ONE 2019, 14, e0212078. [Google Scholar] [CrossRef] [PubMed]
- Murchie, E.H.; Pinto, M.; Horton, P. Agriculture and the new challenges for photosynthesis research. New Phytol. 2009, 181, 532–552. [Google Scholar] [CrossRef] [PubMed]
- Mohapatra, T.; Robin, S.; Sarla, N.; Sheshashayee, M.; Singh, A.K.; Singh, K.; Singh, N.K.; Sevanthi, A.M.; Sharma, R.P. EMS induced mutants of upland rice variety Nagina22: Generation and characterization. Proc. Indian Natl. Sci. Acad. 2014, 80, 163–172. [Google Scholar] [CrossRef]
- Huang, L.Y.; Yang, D.S.; Li, X.X.; Peng, S.B.; Wang, F. Coordination of high grain yield and high nitrogen use efficiency through large sink size and high post-heading source capacity in rice. Field Crops Res. 2019, 233, 49–58. [Google Scholar] [CrossRef]
- Vishwakarma, C.; Krishna, G.K.; Kapoor, R.T.; Mathur, K.; Lal, S.L.; Saini, R.P.; Yadava, P.; Chinnusamy, V. Bioengineering of canopy photosynthesis in rice for securing global food security: A critical review. Agronomy 2023, 13, 489. [Google Scholar] [CrossRef]
- Peng, S.; Khush, G.S.; Virk, P.; Tang, Q.; Zou, Y. Progress in ideotype breeding to increase rice yield potential. Field Crops Res. 2008, 108, 32–38. [Google Scholar] [CrossRef]
- Zhang, Z.; Chu, G.; Liu, L.; Wang, Z.; Wang, X.; Zhang, H.; Yang, J.; Zhang, J. Mid-season nitrogen application strategies for rice varieties differing in panicle size. Field Crops Res. 2013, 150, 9–18. [Google Scholar] [CrossRef]
- Jiang, Y.H.; Zhang, H.C.; Zhao, K.; Xu, J.W.; Wei, H.H.; Long, H.Y.; Wang, W.T.; Dai, Q.G.; Huo, Z.Y.; Xu, K.; et al. Difference in yield and its components characteristics of different type rice cultivars in the lower reaches of the Yangtze River. Chinese J. Rice Sci. 2014, 28, 621–631. [Google Scholar]
- Yang, J.; Peng, S.; Zhang, Z.; Wang, Z.; Visperas, R.M.; Zhu, Q. Grain and dry matter yields and partitioning of assimilates in japonica/indica hybrid rice. Crop Sci. 2002, 42, 766–772. [Google Scholar] [CrossRef]
- Ambavaram, M.M.R.; Basu, S.; Krishnan, A.; Ramegowda, V.; Batlang, U.; Rahman, L.; Baisakh, N.; Pereira, A. Coordinated regulation of photosynthesis in rice increases yield and tolerance to environmental stress. Nat. Commun. 2014, 5, 5302. [Google Scholar] [CrossRef] [PubMed]
- Richards, R.A. Selectable traits to increase crop photosynthesis and yield of grain crops. J. Exp. Bot. 2000, 51, 447–458. [Google Scholar] [CrossRef] [PubMed]
- Cheng, S.; Zhuang, J.; Fan, Y.; Du, J.; Cao, L. Progress in research and development on hybrid rice: A super-domesticate in China. Ann. Bot. 2007, 100, 959–966. [Google Scholar] [CrossRef] [PubMed]
- Pan, Y.; Gao, S.; Xie, K.; Lu, Z.; Meng, X.; Wang, S.; Lu, J.; Guo, S. Higher radiation use efficiency produces greater biomass before heading and grain yield in super hybrid rice. Agronomy 2020, 10, 209. [Google Scholar] [CrossRef]
- Matsubara, K.; Yamamoto, E.; Kobayashi, N.; Ishii, T.; Tanaka, J.; Tsunematsu, H.; Yoshinaga, S.; Matsumura, O.; Yonemaru, J.-I.; Mizobuchi, R.; et al. Improvement of rice biomass yield through QTL-based selection. PLoS ONE 2016, 11, e0151830. [Google Scholar] [CrossRef]
- Long, S.P. Photosynthesis engineered to increase rice yield. Nat. Food 2020, 1, 105. [Google Scholar] [CrossRef]
- Yoon, D.K.; Ishiyama, K.; Suganami, M.; Tazoe, Y.; Watanabe, M.; Imaruoka, S.; Ogura, M.; Ishida, H.; Suzuki, Y.; Obara, M.; et al. Transgenic rice overproducing Rubisco exhibits increased yields with improved nitrogen-use efficiency in an experimental paddy field. Nat. Food 2020, 1, 134–139. [Google Scholar] [CrossRef]
- Okamura, M.; Aoki, N.; Hirose, T.; Yonekura, M.; Ohto, C.; Ohsugi, R. Tissue specificity and diurnal change in gene expression of the sucrose phosphate synthase gene family in rice. Plant Sci. 2011, 181, 159–166. [Google Scholar] [CrossRef]
- Fischer, R.A.; Gregory, O.E. Breeding and cereal yield progress. Crop Sci. 2010, 50, S-85–S-98. [Google Scholar] [CrossRef]
- Huang, M.; Zou, Y.B.; Jiang, P.; Xia, B.; Ibrahim, M.; Ao, H.J. Relationship between grain yield and yield components in super hybrid rice. Agric. Sci. China 2011, 10, 1537–1544. [Google Scholar] [CrossRef]
- Wei, H.-H.; Yang, Y.-I.; Sao, X.-Y.; Shi, T.-Y.; Meng, T.-Y.; Lu, Y.; Tao, Y.; Li, I.; Ding, E.; Chen, Y.-L.; et al. Higher leaf area through leaf width and lower leaf angle were the primary morphological traits for yield advantage of japonica/indica hybrids. J. Integr. Agric. 2020, 19, 483–494. [Google Scholar] [CrossRef]
- Sheehy, J.E.; Mitchell, P.L. Designing Rice for the 21st Century: The Three Laws of Maximum Yield; Discussion Paper Series 48; International Rice Research Institute: Los Baños, Philippines, 2013; 19p. [Google Scholar]
- Jahn, C.E.; Mckay, J.K.; Mauleon, R.; Stephens, J.; McNally, K.L.; Bush, D.R.; Leung, H.; Leach, J.E. Genetic variation in biomass traits among 20 diverse rice varieties. Plant Physiol. 2011, 155, 157–168. [Google Scholar] [CrossRef]
- Honda, S.; Ohkubo, S.; San, N.S.; Nakkasame, A.; Tomisawa, K.; Katsura, K.; Ookawa, T.; Nagano, A.J.; Adachi, S. Maintaining higher leaf photosynthesis after heading stage could promote biomass accumulation in rice. Sci. Rep. 2021, 11, 7579. [Google Scholar] [CrossRef]
- Feng, Y.; Wu, H.; Liu, H.; He, Y.; Yin, Z. Effects of OsRCA Overexpression on rubisco activation state and photosynthesis in maize. Plants 2023, 11, 1614. [Google Scholar] [CrossRef]
- Smith, J.; Wijewardene, I.; Cai, Y.; Esmaeili, N.; Shen, G.; Hequet, E.; Ritchie, G.; Payton, P.; Zhang, H. Co-overexpression of RCA and AVP1 in cotton substantially improves fiber yield for cotton under drought, moderate heat, and salt stress conditions. Curr. Res. Biotechnol. 2023, 5, 100123. [Google Scholar] [CrossRef]
- Tamoi, M.; Nagaoka, M.; Miyagawa, Y.; Shigeoka, S. Contribution of fructose-1,6-bisphosphatase and sedoheptulose-1,7-bisphosphatase to the photosynthetic rate and carbon flow in the Calvin cycle in transgenic plants. Plant Cell Physiol. 2006, 47, 380–390. [Google Scholar] [CrossRef]
- Li, Y.Y.; Guo, L.-N.; Liang, C.-Z.; Meng, Z.-G.; Tahira, S.; Guo, S.-D.; Zhang, R. Overexpression of Brassica napus cytosolic fructose-1,6-bisphosphatase and sedoheptulose-1,7-bisphosphatase genes significantly enhanced tobacco growth and biomass. J. Integr. Agric. 2022, 21, 49–59. [Google Scholar] [CrossRef]
- Cao, Y.; Zhang, Z.; Zhang, T.; You, Z.; Geng, J.; Wang, Y.; Hu, T.; Yang, P. Overexpression of zeaxanthin epoxidase gene from Medicago sativa enhances the tolerance to low light in transgenic tobacco. Acta. Biochim. Pol. 2018, 65, 431–435. [Google Scholar] [CrossRef] [PubMed]
- Li, R.; Li, M.; Ashraf, U.; Liu, S.; Zhang, J. Exploring the relationships between yield and yield-related traits for rice varieties released in China From 1978 to 2017. Front. Plant. Sci. 2019, 10, 543. [Google Scholar] [CrossRef] [PubMed]
- Tiwari, K.K.; Singh, A.; Pattnaik, S.; Sandhu, M.; Kaur, S.; Jain, S.; Tiwari, S.; Mehrotra, S.; Anumalla, M.; Samal, R.; et al. Identification of a diverse mini-core panel of Indian rice germplasm based on genotyping using microsatellite markers. Plant Breed. 2015, 134, 164–171. [Google Scholar] [CrossRef]
- Arnon, D.I.; Whatley, F.R. Is chloride a coenzyme of photosynthesis? Science 1949, 110, 554–556. [Google Scholar] [CrossRef] [PubMed]
- Yemm, E.W.; Willis, A. The estimation of carbohydrates in plant extracts by anthrone. Biochem. J. 1954, 57, 508. [Google Scholar] [CrossRef] [PubMed]
- Hovenkamp-Hermelink, J.H.M.; Jacobsen, E.; Ponstein, A.S.; Visser, R.G.F.; Vos-Scheperkeuter, G.H.; Bijmolt, E.W.; de Vries, J.N.; Witholt, B.; Feenstra, W.J. Isolation of an amylose-free starch mutant of the potato (Solanum tuberosum L.). Theor. Appl. Genet. 1987, 75, 217–221. [Google Scholar] [CrossRef]
- Chomczynski, P.; Sacchi, N. Single-step method of RNA isolation by acid guanidinium thiocyanate-phenol-chloroform extraction. Anal. Biochem. 1987, 162, 156–159. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–428. [Google Scholar] [CrossRef]
- Singh, N.; Jayaswal, P.K.; Panda, K.; Mandal, P.; Kumar, V.; Singh, B.; Mishra, S.; Singh, Y.; Singh, R.; Rai, V.; et al. Single-copy gene based 50 K SNP chip for genetic studies and molecular breeding in rice. Sci. Rep. 2015, 5, 11600. [Google Scholar] [CrossRef]
- Droc, G.; Ruiz, M.; Larmande, P.; Pereira, A.; Piffanelli, P.; Morel, J.B.; Dievart, A.; Courtois, B.; Guiderdoni, E.; Perin, C. Ory Genes DB: A database for rice reverse genetics. Nucleic Acids Res. 2006, 34, 736–740. [Google Scholar] [CrossRef]
- Artimo, P.; Jonnalagedda, M.; Arnold, K.; Baratin, D.; Csardi, G.; De Castro, E.; Duvaud, S.; Flegel, V.; Fortier, A.; Gasteiger, E.; et al. ExPASy: SIB bioinformatics resource portal. Nucleic Acids Res. 2012, 40, 597–603. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef]
- Tamura, K.; Nei, M. Estimation of the number of nucleotide substitutions in the control region of mitochondrial DNA in humans and chimpanzees. Mol. Biol. Evol. 1993, 10, 512–526. [Google Scholar] [PubMed]
- Felsenstein, J. Confidence limits on phylogenies: An approach using the bootstrap. Evolution 1985, 39, 783–791. [Google Scholar] [CrossRef] [PubMed]
- R Core Team. R: A Language and Environment for Statistical Computing. R Foundation for Statistical Computing; R Core Team: Vienna, Austria, 2013. [Google Scholar]
- De Mendiburu, F. Agricolae: Statistical Procedures for Agricultural Research. R Package Version 2019, 1.3-1. Available online: https://CRAN.R-project.org/package=agricolae (accessed on 15 April 2022).




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sl No. | Genotypes | Details of the Genotypes/Parentage etc. |
|---|---|---|
| Low Yielding (LY) | ||
| 1 | Anjali | Parentage: PR-19-2 X RR-149-1129, released in 2002 by the National Rice Research Institute (NRRI), Cuttack, early maturing (90–95 days), semi-tall (85–100 cm), suitable for rainfed, direct-seeded conditions, grains: short bold, white, moderately tolerant to drought, resistant to brown spot, moderately resistant to leaf blast, yield: 3 t/ha under direct seeded, and 4 t/ha under transplanted conditions. |
| 2 | Ghanteswari | Parentage—IR-2061-628- 1-6-4-3 x N-2-2, released in 1994, early maturing (90–95 days), dwarf (85 cm), resistant to blast, neck blast, yield: 35 Q/ha. High silicon uptake ability. |
| 3 | Parijat | Parentage—TKM-6xT(N)1, released in 1985, duration 95–100 days, dwarf (85 cm), medium slender grains, moderately resistant to sheath blight, blast, leaf blight and helminthosporium leaf spot, yield: 30–40 q/ha. |
| 4 | RKVY-104 | Duration (105 days), medium plant height, |
| 5 | Khao Daw Tai | Duration (105 days) |
| 6 | Ghati Kamma Nangarhar | Duration (110 days), medium plant height. |
| 7 | BAM4510 | IC-6294, germplasm line from Northeast India |
| 8 | BAM5850 | IC38224, germplasm line from Northeast India |
| High Yielding (HY) | ||
| 9 | Dular | It is an upland-adapted, drought-tolerant traditional genotype from India and has shown consistent performance in drought screening at IRRI. It is early maturing. |
| 10 | Gontra Bidhan 3 | Duration—110–115 days, plant height—95–100 cm, short medium bold grain type, average yield—6t/ha, suitable for medium to medium upland. |
| 11 | Way Rarem | It is an Indica variety from Indonesia. Medium plant height and higher chlorophyll contents. |
| 12 | Patchai Perumal | It is a water-deficit stress-tolerant variety.Tall plant height, mostly cultivated in Kerala, India and Sri Lanka. |
| 13 | Sahbhagi Dhan | Dwarf statured (90–100 cm), drought-tolerant variety suitable for upland, rainfed direct seeded as well as transplanted conditions. It bears golden-husked long bold grains and yields 3.8–4.5 t/ha. It is resistant to leaf blast and moderately resistant to brown spot, sheath rot, stem borer, leaf folder, etc. It is cultivated in Jharkhand and Odisha. Released by NRRI, Cuttack in 2009. |
| 14 | Indira Barani Dhan-1 | Parentage—Swarna x IR 42253, maturity—111–115 days, medium slender grain, suitable for rainfed shallow lowland ecosystem of Chhattisgarh plains and adapted to aerobic conditions, average yield 40–45 q/ha. |
| 15 | MTU 1010 | An elite, high-yielding, short-duration, widely cultivated megavariety, possessing long slender grain type, was developed and released by Acharya NG Ranga Agricultural University (ANGRAU), Guntur, Andhra P. |
| 16 | Maudamani | Parentage/Pedigree: Dandi/Naveen/Dandi. It is also known as CR DHAN 307. Released by ICAR-National Rice Research Institute, Cuttack in 2014. |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vishwakarma, C.; Krishna, G.K.; Kapoor, R.T.; Mathur, K.; Dalal, M.; Singh, N.K.; Mohapatra, T.; Chinnusamy, V. Physiological Analysis of Source–Sink Relationship in Rice Genotypes with Contrasting Grain Yields. Plants 2024, 13, 62. https://doi.org/10.3390/plants13010062
Vishwakarma C, Krishna GK, Kapoor RT, Mathur K, Dalal M, Singh NK, Mohapatra T, Chinnusamy V. Physiological Analysis of Source–Sink Relationship in Rice Genotypes with Contrasting Grain Yields. Plants. 2024; 13(1):62. https://doi.org/10.3390/plants13010062
Chicago/Turabian StyleVishwakarma, Chandrapal, Gopinathan Kumar Krishna, Riti Thapar Kapoor, Komal Mathur, Monika Dalal, Nagendra Kumar Singh, Trilochan Mohapatra, and Viswanathan Chinnusamy. 2024. "Physiological Analysis of Source–Sink Relationship in Rice Genotypes with Contrasting Grain Yields" Plants 13, no. 1: 62. https://doi.org/10.3390/plants13010062
APA StyleVishwakarma, C., Krishna, G. K., Kapoor, R. T., Mathur, K., Dalal, M., Singh, N. K., Mohapatra, T., & Chinnusamy, V. (2024). Physiological Analysis of Source–Sink Relationship in Rice Genotypes with Contrasting Grain Yields. Plants, 13(1), 62. https://doi.org/10.3390/plants13010062