
Exogenously Applied Cyclitols and Biosynthesized Silver Nanoparticles Affect the Soluble Carbohydrate Profiles of Wheat (Triticum aestivum L.) Seedling

 , , ,
, , , 
 ,
,  and
and 
Abstract
1. Introduction
2. Results
2.1. The Uptake of Cyclitols by Wheat Grains during Imbibition—Preliminary Experiment
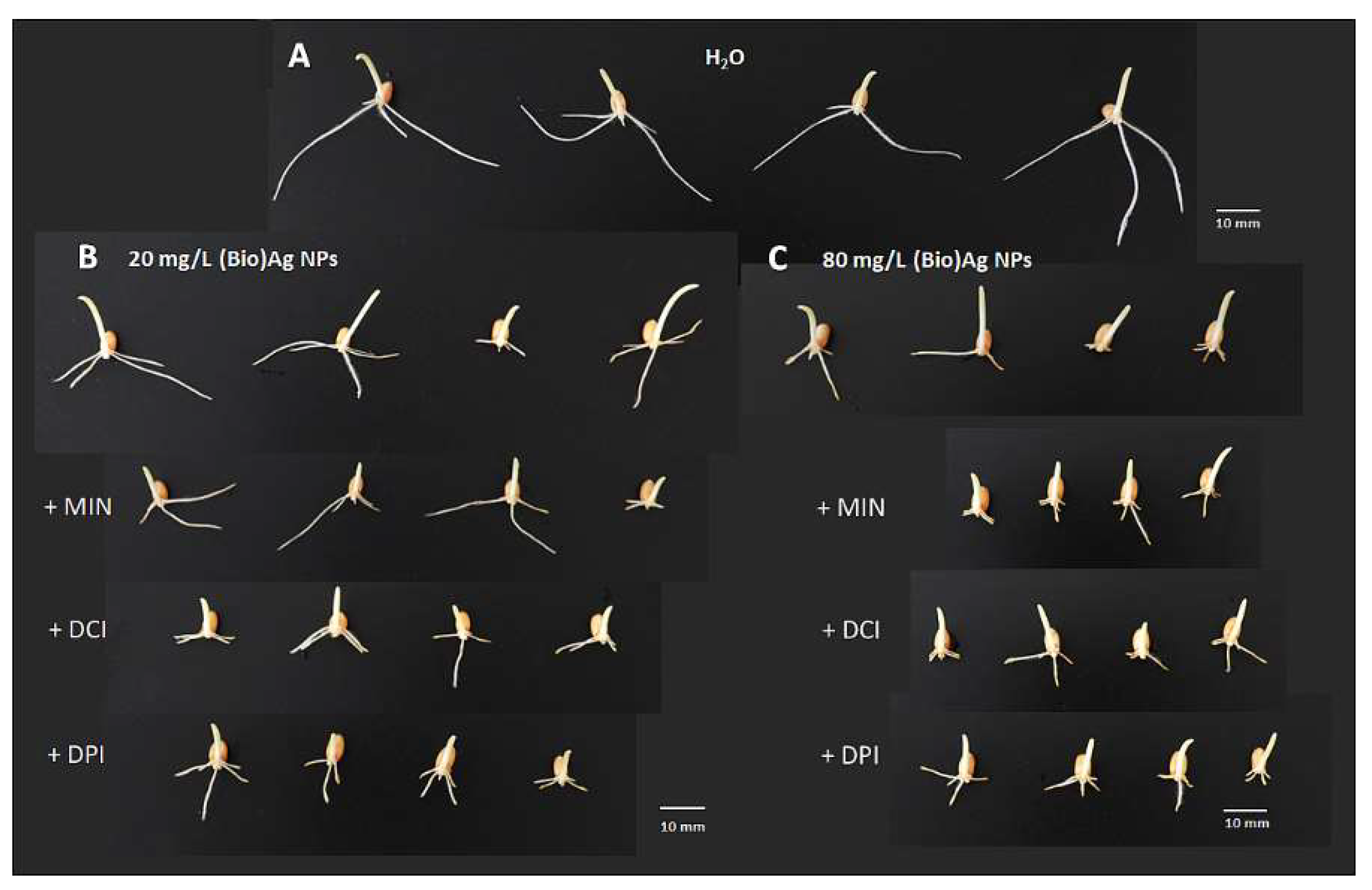
2.2. The Effect of Cyclitols on Phytotoxicity of Biosynthesized Silver Nanoparticles ((Bio)Ag NPs) to Wheat Seedlings
Soluble Carbohydrates
2.3. The Effect of Grain Priming with d-Pinitol, GSH and Their Mixtures on the Phytotoxicity of (Bio)Ag NPs against Seedlings
Soluble Carbohydrates
3. Discussion
3.1. The Role of Cyclitols in Reducing the Phytotoxic Effects of (Bio)Ag NPs
3.2. GSH and GSH/PIN in The Protection of Wheat Seedlings against (Bio)Ag NPs
3.3. The Effect of (Bio)Ag NPs and Cyclitols on Soluble Carbohydrates in Seedlings
4. Materials and Methods
4.1. Preparation of (Bio)Ag NP Suspensions
4.2. The Uptake and Transport of Cyclitols in Germinating Wheat Grains—A Preliminary Study
4.3. The Effect of Exogenous Cyclitols on the Phytotoxicity of (Bio)AgNPs
4.4. The Effects of Seed Priming with d-Pinitol, GSH and Their Mixtures on The Phytotoxicity of (Bio)AgNPs
4.5. Soluble Carbohydrate Analyses
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Shang, Y.; Hasan, M.K.; Ahammed, G.J.; Li, M.; Yin, H.; Zhou, J. Applications of nanotechnology in plant growth and crop protection: A review. Molecules 2019, 24, 2558. [Google Scholar] [CrossRef]
- Tripathi, D.K.; Tripathi, A.; Singh, S.; Singh, Y.; Vishwakarma, K.; Yadav, G.; Sharma, S.; Singh, V.K.; Mishra, R.K.; Upadhyay, R.G.; et al. Uptake, accumulation and toxicity of silver nanoparticle in autotrophic plants, and heterotrophic microbes: A concentric review. Front. Microbiol. 2017, 8, 7. [Google Scholar] [CrossRef] [PubMed]
- Juárez-Maldonado, A.; Ortega-Ortíz, H.; Morales-Díaz, A.B.; González-Morales, S.; Morelos-Moreno, A.; Cabrera-De la Fuente, M.; Sandoval-Rangel, A.; Cadenas-Pliego, G.; Benavides-Mendoza, A. Nanoparticles and nanomaterials as plant biostimulants. Int. J. Mol. Sci. 2019, 20, 162. [Google Scholar] [CrossRef] [PubMed]
- Salem Attia, T.M.; Elsheery, N.I. Nanomaterials: Scope, applications, and challenges in agriculture and soil reclamation. In Sustainable Agriculture Reviews 41, Sustainable Agriculture Reviews; Hayat, S., Ed.; Springer: Berlin/Heidelberg, Germany, 2020; Volume 41, pp. 1–39. [Google Scholar]
- Crisan, C.M.; Mocan, T.; Manolea, M.; Lasca, L.I.; Tăbăran, F.-A.; Mocan, L. Review on silver nanoparticles as a novel class of antibacterial solutions. Appl. Sci. 2021, 11, 1120. [Google Scholar] [CrossRef]
- Hamad, A.; Khashan, K.S.; Hadi, A. Silver nanoparticles and silver ions as potential antibacterial agents. J. Inorg. Organomet. Polym. Mater. 2020, 20, 4811–4828. [Google Scholar] [CrossRef]
- Syafiuddin, A.; Salmiati; Salim, M.R.; Kueh, A.B.H.; Hadibarata, T.; Nur, H. A review of silver nanoparticles: Research trends, global consumption, synthesis, properties, and future challenges. J. Chin. Chem. Soc. 2017, 64, 732–756. [Google Scholar] [CrossRef]
- Tariq, M.; Mohammad, K.N.; Ahmed, B.; Siddiqui, M.A.; Lee, J. Biological synthesis of silver nanoparticles and prospects in plant disease management. Molecules 2022, 27, 4754. [Google Scholar] [CrossRef]
- Kim, S.W.; Jung, J.H.; Lamsal, K.; Kim, Y.S.; Min, J.S.; Lee, Y.S. Antifungal effects of silver nanoparticles (AgNPs) against various plant pathogenic fungi. Mycobiology 2012, 40, 53–58. [Google Scholar] [CrossRef]
- Gulzar, S.; Zulfiqar, R.; Tahira, M.; Abbas, F.; Khadhim, M.; Pervaiz, S.; Noreen, S.; Hafeez, M.A.; Abbas, M.; Habib, J. Synthesis and characterization of silver nanoparticles against insect pests. Inter. J. Sci. Res. Publ. 2020, 10, 119–136. [Google Scholar] [CrossRef]
- Khan, M.; Khan, A.U.; Hasan, M.A.; Yadav, K.K.; Pinto, M.M.C.; Malik, N.; Yadav, V.K.; Khan, A.H.; Islam, S.; Sharma, G.K. Agro-nanotechnology as an emerging field: A novel sustainable approach for improving plant growth by reducing biotic stress. Appl. Sci. 2021, 11, 2282. [Google Scholar] [CrossRef]
- Ehsan, M.; Raja, N.I.; Mashwani, Z.U.R.; Zohra, E.; Abasi, F.; Ikram, M.; Mustafa, N.; Wattoo, F.H.; Proćków, J.; Pérez de la Lastra, J.M. Effects of phytogenically synthesized bimetallic Ag/ZnO nanomaterials and nitrogen-based fertilizers on biochemical and yield attributes of two wheat varieties. Nanomaterials 2022, 12, 2894. [Google Scholar] [CrossRef] [PubMed]
- Jhanzab, H.M.; Qayyum, A.; Bibi, Y.; Sher, A.; Hayat, M.T.; Iqbal, J.; Qamar, M.; Elesawy, B.H.; Ismail, K.A.; Gharib, A.F.; et al. Chemo-blended Ag & Fe nanoparticles effect on growth, physiochemical and yield traits of wheat (Triticum aestivum). Agronomy 2022, 12, 757. [Google Scholar] [CrossRef]
- Aguirre-Becerra, H.; Feregrino-Perez, A.A.; Esquivel, K.; Perez-Garcia, C.E.; Vazquez-Hernandez, M.C.; Mariana-Alvarado, A. Nanomaterials as an alternative to increase plant resistance to abiotic stresses. Front. Plant Sci. 2022, 13, 1023636. [Google Scholar] [CrossRef]
- Tripathi, D.; Singh, M.; Pandey-Rai, S. Crosstalk of nanoparticles and phytohormones regulate plant growth and metabolism under abiotic and biotic stress. Plant Stress 2022, 6, 100107. [Google Scholar] [CrossRef]
- Pociecha, E.; Gorczyca, A.; Dziurka, M.; Matras, E.; Oćwieja, M. Silver nanoparticles and silver ions differentially affect the phytohormone balance and yield in wheat. Agriculture 2021, 11, 729. [Google Scholar] [CrossRef]
- Dutta Gupta, S.; Agarwal, A.; Pradhan, S. Phytostimulatory effect of silver nanoparticles (AgNPs) on rice seedling growth: An insight from antioxidative enzyme activities and gene expression patterns. Ecotox. Environ. Saf. 2018, 161, 624–633. [Google Scholar] [CrossRef]
- Manaf, A.; Wang, X.; Tariq, F.; Jhanzab, H.M.; Bibi, Y.; Sher, A.; Razzaq, A.; Fiaz, S.; Tanveer, S.K.; Qayyum, A. Antioxidant enzyme activities correlated with growth parameters of wheat sprayed with silver and gold nanoparticle suspensions. Agronomy 2021, 11, 1494. [Google Scholar] [CrossRef]
- Nile, S.H.; Thiruvengadam, M.; Wang, Y.; Samynathan, R.; Shariati, M.A.; Rebezov, M.; Nile, A.; Sun, M.; Venkidasamy, B.; Xiao, J.; et al. Nano-priming as emerging seed priming technology for sustainable agriculture—Recent developments and future perspectives. J. Nanotechnol. 2022, 20, 254. [Google Scholar] [CrossRef]
- Khan, M.N.; Fu, C.; Li, J.; Tao, Y.; Li, Y.; Hu, J.; Chen, L.; Khan, Z.; Wu, H.; Li, Z. Seed nanopriming: How do nanomaterials improve seed tolerance to salinity and drought? Chemosphere 2023, 310, 136911. [Google Scholar] [CrossRef]
- Yan, A.; Chen, Z. Impacts of silver nanoparticles on plants: A focus on the phytotoxicity and underlying mechanism. Int. J. Mol. Sci. 2019, 20, 1003. [Google Scholar] [CrossRef]
- Szőllősi, R.; Molnár, A.; Kondak, S.; Kolbert, Z. Dual effect of nanomaterials on germination and seedling growth: Stimulation vs. phytotoxicity. Plants 2020, 9, 1745. [Google Scholar] [CrossRef] [PubMed]
- Siddiqi, K.S.; Husen, A. Plant response to silver nanoparticles: A critical review. Crit. Rev. Biotechnol. 2021, 42, 973–990. [Google Scholar] [CrossRef] [PubMed]
- McShan, D.; Ray, P.C.; Yu, H. Molecular toxicity mechanism of nanosilver. J. Food Drug Anal. 2014, 22, 116–127. [Google Scholar] [CrossRef] [PubMed]
- Lahuta, L.B.; Szablińska-Piernik, J.; Głowacka, K.; Stałanowska, K.; Railean-Plugaru, V.; Horbowicz, M.; Pomastowski, P.; Buszewski, B. The effect of biosynthesized silver nanoparticles on germination, early seedling development, and metabolome of wheat (Triticum aestivum L.). Molecules 2022, 27, 2303. [Google Scholar] [CrossRef]
- Lahuta, L.B.; Szablińska-Piernik, J.; Stałanowska, K.; Głowacka, K.; Horbowicz, M. The size-dependent effects of silver nanoparticles on germination, early seedling development and polar metabolite profile of wheat (Triticum aestivum L.). Int. J. Mol. Sci. 2022, 23, 13255. [Google Scholar] [CrossRef]
- Szablińska-Piernik, J.; Lahuta, L.B.; Stałanowska, K.; Horbowicz, M. The imbibition of pea (Pisum sativum L.) seeds in silver nitrate reduces seed germination, seedlings development and their metabolic profile. Plants 2022, 11, 1877. [Google Scholar] [CrossRef]
- Kannaujia, R.; Srivastava, C.M.; Prasad, V.; Singh, B.N.; Pandey, V. Phyllanthus emblica fruit extract stabilized biogenic silver nanoparticles as a growth promoter of wheat varieties by reducing ROS toxicity. Plant Physiol. Biochem. 2019, 149, 460–471. [Google Scholar] [CrossRef]
- Bayat, M.; Zargar, M.; Murtazova, K.M.-S.; Nakhaev, M.R.; Shkurkin, S.I. Ameliorating seed germination and seedling growth of nano-primed wheat and flax seeds using seven biogenic metal-based nanoparticles. Agronomy 2022, 12, 811. [Google Scholar] [CrossRef]
- Komatsu, S.; Yamaguchi, H.; Hitachi, K.; Tsuchida, K. Proteomic, biochemical, and morphological analyses of the effect of silver nanoparticles mixed with organic and inorganic chemicals on wheat growth. Cells 2022, 11, 1579. [Google Scholar] [CrossRef]
- Kranner, I.; Grill, D. Significance of thiol-disulfide exchange in resting stages of plant development. Bot. Acta 1996, 109, 8–141996. [Google Scholar] [CrossRef]
- Yang, F.; Basu, T.K.; Ooraikul, B. Studies on germination conditions and antioxidant contents of wheat grain. Int. J. Food Sci. Nutr. 2001, 52, 319–330. [Google Scholar] [CrossRef] [PubMed]
- Hasanuzzaman, M.; Borhannuddin Bhuyan, M.H.M.; Anee, T.I.; Parvin, K.; Nahar, K.; Al Mahmud, J.; Fujita, M. Regulation of ascorbate-glutathione pathway in mitigating oxidative damage in plants under abiotic stress. Antioxidants 2019, 8, 384. [Google Scholar] [CrossRef]
- Paradiso, A.; Bernardino, R.; de Pinto, M.C.; di Toppi, L.S.; Storelli, M.M.; Tommasi, F.; de Gara, L. Increase in ascorbate-glutathione metabolism as local precocious systemic responses induced by cadmium in durum wheat plants. Plant Cell Physiol. 2008, 49, 362–374. [Google Scholar] [CrossRef] [PubMed]
- Gerna, D.; Roach, T.; Stöggl, W.; Wagner, J.; Vaccino, P.; Limonta, M.; Kranner, I. Changes in low-molecular-weight thioldisulphide redox couples are part of bread wheat seed germination and early seedling growth. Free Radic. Res. 2017, 51, 568–581. [Google Scholar] [CrossRef]
- Farooq, M.A.; Zhang, X.; Zafar, M.M.; Ma, W.; Zhao, J. Roles of reactive oxygen species and mitochondria in seed germination. Front. Plant Sci. 2021, 12, 781734. [Google Scholar] [CrossRef]
- Karimi, J.; Mohsenzadeh, S.; Niazi, A.; Moghadam, A. Differential expression of mitochondrial manganese superoxide dismutase (SOD) in Triticum aestivum exposed to silver nitrate and silver nanoparticles. Iranian J. Biotechnol. 2017, 15, e1311. [Google Scholar] [CrossRef] [PubMed]
- Lourenço, S.C.; Moldão-Martins, M.; Alves, V.D. Antioxidants of natural plant origins: From sources to food industry applications. Molecules 2019, 24, 4132. [Google Scholar] [CrossRef]
- Signorelli, S.; Dans, P.D.; Coitiño, E.L.; Borsani, O.; Monza, J. Connecting proline and γ-aminobutyric acid in stressed plants through non-enzymatic reactions. PLoS ONE 2015, 10, e0115349. [Google Scholar] [CrossRef]
- Michell, R.H. Evolution of the diverse biological role of inositols. Biochem. Soc. Symp. 2007, 74, 223–246. [Google Scholar] [CrossRef]
- Riemer, E.; Pullagurla, N.J.; Yadav, R.; Rana, P.; Jessen, H.J.; Kamleitner, M.; Schaaf, G.; Laha, D. Regulation of plant biotic interactions and abiotic stress responses by inositol polyphosphates. Front. Plant Sci. 2022, 13, 944515. [Google Scholar] [CrossRef]
- Loewus, F.A.; Murthy, P.P.N. myo-Inositol metabolism in plants. Plant Sci. 2000, 150, 1–19. [Google Scholar] [CrossRef]
- Sparvoli, F.; Cominelli, E. Seed biofortification and phytic acid reduction: A conflict of interest for the plant? Plants 2015, 4, 728–755. [Google Scholar] [CrossRef]
- Kanter, U.; Usadel, B.; Guerineau, F.; Li, Y.; Pauly, M.; Tenhaken, R. The inositol oxygenase gene family of Arabidopsis is involved in the biosynthesis of nucleotide sugar precursors for cell-wall matrix polysaccharides. Planta 2005, 221, 243–254. [Google Scholar] [CrossRef] [PubMed]
- Valluru, R.; Van den Ende, W. Myo-inositol and beyond—Emerging networks under stress. Plant Sci. 2011, 181, 387–400. [Google Scholar] [CrossRef]
- Munir, S.; Mumtaz, M.A.; Ahiakpa, J.K.; Liu, G.; Chen, W.; Zhou, G.; Zheng, W.; Ye, Z.; Zhang, Y. Genome-wide analysis of Myo-inositol oxygenase gene family in tomato reveals their involvement in ascorbic acid accumulation. BMC Genomics 2020, 21, 284. [Google Scholar] [CrossRef] [PubMed]
- Couée, I.; Sulmon, C.; Gouesbet, G.; El Amrani, A. Involvement of soluble sugars in reactive oxygen species balance and responses to oxidative stress in plants. J. Exp. Bot. 2006, 57, 449–459. [Google Scholar] [CrossRef]
- Jiang, W.-D.; Hu, K.; Liu, Y.; Jiang, J.; Wu, P.; Zhao, J.; Zhang, Y.-A.; Zhou, X.-Q.; Feng, L. Dietary myo-inositol modulates immunity through antioxidant activity and the Nrf2 and E2F4/cyclin signalling factors in the head kidney and spleen following infection of juvenile fish with Aeromonas hydrophila. Fish Shellfish. Immunol. 2016, 49, 374–386. [Google Scholar] [CrossRef]
- Ali, F.; Manzoor, U.; Bhattacharya, R.; Bansal, A.K.; Chandrashekharaiah, K.S.; Singh, L.R.; Saraswati, S.M.; Uversky, V.; Dar, T.A. Brain metabolite, myo-inositol, inhibits catalase activity: A mechanism of the distortion of the antioxidant defense system in Alzheimer’s disease. ACS Omega 2022, 7, 12690–12700. [Google Scholar] [CrossRef]
- Orthen, B.; Popp, M.; Smirnoff, N. Hydroxyl radical scavenging properties of cyclitols. Proc. R. Soc. Edinb. Sect. B Biol. Sci. 1994, 102, 269–272. [Google Scholar] [CrossRef]
- Sánchez-Hidalgo, M.; León-González, A.J.; Gálvez-Peralta, M.; González-Mauraza, N.H.; Martin-Cordero, C. d-Pinitol: A cyclitol with versatile biological and pharmacological activities. Phytochem. Rev. 2021, 20, 211–224. [Google Scholar] [CrossRef]
- Płonka, J.; Szablińska-Piernik, J.; Buszewski, B.; Baranowska, I.; Lahuta, L.B. Analyses of antioxidative properties of selected cyclitols and their mixtures with flavanones and glutathione. Molecules 2022, 27, 158. [Google Scholar] [CrossRef]
- Ortbauer, M.; Popp, M. Functional role of polyhydroxy compounds on protein structure and thermal stability studied by circular dichroism spectroscopy. Plant Physiol. Biochem. 2008, 46, 428–434. [Google Scholar] [CrossRef] [PubMed]
- Merchant, A.; Richter, A.A. Polyols as biomarkers and bioindicators for 21st century plant breeding. Func. Plant Biol. 2011, 38, 934–940. [Google Scholar] [CrossRef] [PubMed]
- Pradas del Real, A.E.; Vidal, V.; Carrière, M.; Castillo-Michel, H.; Levard, C.; Chaurand, P.; Sarret, G. Silver nanoparticles and wheat roots: A complex interplay. Environ. Sci. Technol. 2017, 51, 5774–5782. [Google Scholar] [CrossRef] [PubMed]
- Yanik, F.; Vardar, F. Assessment of silver nanoparticle-induced morphological, biochemical and physiological alterations in wheat roots. Ann. Bot. 2019, 9, 83–94. [Google Scholar] [CrossRef]
- Barbasz, A.; Kreczmer, B.; Oćwieja, M. Effects of exposure of callus cells of two wheat varieties to silver nanoparticles and silver salt (AgNO3). Acta Physiol. Plant. 2016, 38, 76. [Google Scholar] [CrossRef]
- Vannini, C.; Domingo, G.; Onelli, E.; De Mattia, F.; Bruni, I.; Marsoni, M.; Bracale, M. Phytotoxic and genotoxic effects of silver nanoparticles exposure on germinating wheat seedlings. J. Plant Physiol. 2014, 171, 1142–1148. [Google Scholar] [CrossRef]
- Gorczyca, A.; Pociecha, E.; Maciejewska-Prończuk, J.; Kula-Maximenko, M.; Oćwieja, M. Phytotoxicity of silver nanoparticles and silver ions toward common wheat. Surf. Innov. 2021, 10, 48–58. [Google Scholar] [CrossRef]
- Matras, E.; Gorczyca, A.; Pociecha, E.; Przemieniecki, S.W.; Oćwieja, M. Phytotoxicity of silver nanoparticles with different surface properties on monocots and dicots model plants. J. Soil Sci. Plant Nutr. 2022, 22, 1647–1664. [Google Scholar] [CrossRef]
- Abdelsalam, N.R.; Abdel-Megeed, A.; Ali, H.M.; Salem, M.Z.M.; Al-Hayali, M.F.A.; Elshikh, M.S. Genotoxicity effects of silver nanoparticles on wheat (Triticum aestivum L.) root tip cells. Ecotoxicol. Environ. Saf. 2018, 155, 76–85. [Google Scholar] [CrossRef]
- Huang, D.; Dang, F.; Huang, Y.; Chen, N.; Zhou, D. Uptake, translocation, and transformation of silver nanoparticles in plants. Environ. Sci. Nano 2022, 9, 12–39. [Google Scholar] [CrossRef]
- Pryshchepa, O.; Pomastowski, P.; Buszewski, B. Silver nanoparticles: Synthesis, investigation techniques, and properties. Adv. Coll. Interface Sci. 2020, 284, 102246. [Google Scholar] [CrossRef]
- Gliga, A.R.; Skoglund, S.; Wallinder, I.O.; Fadeel, B.; Karlsson, H.L. Size-dependent cytotoxicity of silver nanoparticles in human lung cells: The role of cellular uptake, agglomeration and Ag release. Part. Fibre Toxicol. 2014, 11, 11. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Hu, C.; Shao, L. The antimicrobial activity of nanoparticles: Present situation and prospects for the future. Int. J. Nanomed. 2020, 15, 8023–8049. [Google Scholar] [CrossRef] [PubMed]
- Singh, P.; Kim, Y.J.; Wang, C.; Mathiyalagan, R.; Jin, C.G.; Yang, D.C. Biogenic silver and gold nanoparticles synthesized using red ginseng root extract, and their applications. Artif. Cells Nanomed. Biotechnol. 2016, 44, 811–816. [Google Scholar] [CrossRef] [PubMed]
- El-Batal, A.I.; Naeem, A.; Al-Abboud, M.A. Green synthesis of silver nanoparticles using Arthrospira platensis extract and its antibacterial activity against Escherichia coli. J. Inorg. Organometallic Polym. Mater. 2020, 30, 4206–4215. [Google Scholar]
- Lahuta, L.B.; Goszczyńska, J. Cyclitols in maturing grains of wheat, Triticale and barley. Acta Soc. Bot. Pol. 2010, 79, 181–187. [Google Scholar] [CrossRef]
- Lahuta, L.B.; Horbowicz, M.; Gojło, E.; Goszczyńska, J.; Górecki, R.J. Exogenously applied d-pinitol and d-chiro-inositol modifies the accumulation of alpha-d-galactosides in developing tiny vetch (Vicia hirsuta [L.] SF Gray) seeds. Acta Soc. Bot. Pol. 2005, 74, 287–296. [Google Scholar] [CrossRef]
- Lahuta, L.B.; Dzik, T. d-chiro-Inositol affects accumulation of raffinose family oligosaccharides in developing embryos of Pisum sativum. J. Plant Physiol. 2011, 168, 352–358. [Google Scholar] [CrossRef]
- Kawai, S.; Ohyama, T.; Kumazawa, K. Possibility of upward transport of d-pinitol in soybean plant: Investigation by petiole girdling treatment. Soil Sci. Plant Nutr. 1985, 31, 287–292. [Google Scholar] [CrossRef]
- Kordan, B.; Gabryś, B.; Dancewicz, K.; Lahuta, L.B.; Piotrowicz-Cieślak, A.; Rowińska, E. European yellow lupine Lupinus luteus and narrow-leaf lupine Lupinus angustifolius as hosts for pea aphid Acyrthosiphon pisum. Entomol. Exp. Appl. 2008, 128, 139–146. [Google Scholar] [CrossRef]
- Schneider, S. Inositol transport proteins. FEBS Lett. 2015, 589, 1049–1058. [Google Scholar] [CrossRef]
- Schneider, S.; Schneidereit, A.; Udvardi, P.; Hammes, U.; Gramann, M.; Dietrich, P.; Sauer, N. Arabidopsis INOSITOL TRANSPORTER2 mediates H+ symport of different inositol epimers and derivatives across the plasma membrane. Plant Physiol. 2007, 145, 1395–1407. [Google Scholar] [CrossRef] [PubMed]
- Obendorf, R.L.; Górecki, R.J. Soluble carbohydrates in legume seeds. Seed Sci. Res. 2012, 22, 219–242. [Google Scholar] [CrossRef]
- Pupel, P.; Szablińska-Piernik, J.; Lahuta, L.B. Two-step d-ononitol epimerization pathway in Medicago truncatula. Plant J. 2019, 100, 237–250. [Google Scholar] [CrossRef]
- Sengupta, S.; Mukherjee, S.; Goswami, L.; Sangma, S.; Mukherjee, A.; Roy, N.; Basak, P.; Majumder, A.L. Manipulation of inositol metabolism for improved plant survival under stress: A “network engineering approach”. J. Plant Biochem. Biotechnol. 2012, 21, 15–23. [Google Scholar] [CrossRef]
- Ahn, C.-H.; Hossain, M.A.; Lee, E.; Kanth, B.K.; Park, P.B. Increased salt and drought tolerance by d-pinitol production in transgenic Arabidopsis thaliana. Biochem. Bioph. Res. Com. 2018, 504, 315–320. [Google Scholar] [CrossRef] [PubMed]
- Li, C.-H.; Tien, H.-J.; Wen, M.-F.; Yen, H.E. Myo-inositol transport and metabolism participate in salt tolerance of halophyte ice plant seedlings. Physiol. Plant. 2021, 172, 1619–1629. [Google Scholar] [CrossRef] [PubMed]
- Zechmann, B.; Mauch, F.; Müller, M. Subcellular immunocytochemical analysis detects the highest concentrations of glutathione in mitochondria and not in plastids. J. Exp. Bot. 2008, 59, 4017–4027. [Google Scholar] [CrossRef]
- Nietzel, T.; Mostertz, J.; Ruberti, C.; Née, G.; Fuchs, P.; Wagner, S.; Moseler, A.; Müller-Schüssele, S.J.; Benamar, A.; Poschet, G.; et al. Redox-mediated kick-start of mitochondrial energy metabolism drives resource-efficient seed germination. Proc. Natl. Acad. Sci. USA 2020, 117, 741–751. [Google Scholar] [CrossRef]
- Van Aken, O. Mitochondrial redox systems as central hubs in plant metabolism and signaling. Plant Physiol. 2021, 186, 36–52. [Google Scholar] [CrossRef]
- Suzuki, N. Fine tuning of ROS, redox and energy regulatory systems associated with the functions of chloroplasts and mitochondria in plants under heat stress. Int. J. Mol. Sci. 2023, 24, 1356. [Google Scholar] [CrossRef]
- Cui, D.; Zhang, P.; Ma, Y.-H.; He, X.; Li, Y.-Y.; Zhano, Y.-C.; Zhang, Z.-Y. Phytotoxicity of silver nanoparticles to cucumber (Cucumis sativus) and wheat (Triticum aestivum). J. Zhejiang Univ. Sci. A 2014, 15, 662–670. [Google Scholar] [CrossRef]
- Guzmán-Ortiz, F.A.; Castro-Rosas, J.; Gómez-Aldapa, C.A.; Mora-Escobedo, R.; Rojas-León, A.; Rodríguez-Marín, M.L.; Falfán-Cortés, R.N.; Román-Gutiérrez, A.D. Enzyme activity during germination of different cereals: A review. Food Rev. Int. 2019, 35, 177–200. [Google Scholar] [CrossRef]
- Aoki, N.; Scofield, G.N.; Wang, X.-D.; Offler, C.E.; Patrick, J.W.; Furbank, R.T. Pathway of sugar transport in germinating wheat seeds. Plant Physiol. 2006, 141, 1255–1263. [Google Scholar] [CrossRef]
- Albrecht, G.; Mustroph, A. Localization of sucrose synthase in wheat roots: Increased in situ activity of sucrose synthase correlates with cell wall thickening by cellulose deposition under hypoxia. Planta 2003, 217, 252–260. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Pritchard, J.; Mieog, J.; Byrne, K.; Colgrave, M.N.; Wang, J.-R.; Ral, J.-P. Over-expression of a wheat late maturity alpha-amylase type 1 impact on starch properties during grain development and germination. Front. Plant Sci. 2022, 13, 811728. [Google Scholar] [CrossRef]
- Mahakham, W.; Sarmah, A.K.; Maensiri, S.; Theerakulpist, P. Nanopriming technology for enhancing germination and starch metabolism of aged rice seeds using photosynthesized silver nanoparticles. Sci. Rep. 2017, 7, 8263. [Google Scholar] [CrossRef]
- Wang, Q.; Li, R.; Li, N.; Ja, Y.; Wang, Y.; Chen, Y.; Panichayupakaranant, P.; Chen, H. The antioxidant activities, inhibitory effects, kinetics, and mechanisms of artocarpin and α-mangostin on α-glucosidase and α-amylase. Int. J. Biol. Macromol. 2022, 213, 880–891. [Google Scholar] [CrossRef]
- Čorković, I.; Gašo-Sokač, D.; Pichler, A.; Šimunović, J.; Kopjar, M. Dietary polyphenols as natural inhibitors of α-amylase and α-glucosidase. Life 2022, 12, 1692. [Google Scholar] [CrossRef]
- Mieog, J.C.; Janeček, Š.; Ral, J.-P. New insight in cereal starch degradation: Identification and structural characterization of four α-amylases in bread wheat. Amylase 2017, 1, 35–49. [Google Scholar] [CrossRef]
- Ciarkowska, A.; Wojtaczka, P.; Kęsy, J.; Ostrowski, M. Auxin homeostasis in maize (Zea mays) is regulated via 1-O-indole-3-acetyl-myo-inositol synthesis at early stages of seedling development and under abiotic stress. Planta 2023, 257, 23. [Google Scholar] [CrossRef] [PubMed]
- Sun, M.; Tuan, P.A.; Izydorczyk, M.S.; Ayele, B.T. Ethylene regulates post-germination seedling growth in wheat through spatial and temporal modulation of ABA/GA balance. J. Exp. Bot. 2020, 71, 1985–2004. [Google Scholar] [CrossRef]
- Strobl, S.M.; Kischka, D.; Heilmann, I.; Mouille, G.; Schneider, S. The tonoplastic inositol transporter INT1 from Arabidopsis thaliana impacts cell elongation in a sucrose-dependent way. Front. Plant Sci. 2018, 9, 1657. [Google Scholar] [CrossRef]
- Maiti, I.B.; Loewus, F.A. myo-Inositol metabolism in germinating wheat. Planta 1978, 142, 55–60. [Google Scholar] [CrossRef] [PubMed]
- Railean, V.; Pomastowski, P.; Płociński, T.; Gloc, M.; Dobrucka, R.; Kurzydłowski, K.J.; Buszewski, B. Consideration of a new approach to clarify the mechanism formation of AgNPs, AgNCl and AgNPs@AgNCl synthesized by biological method. Nanoscale Res. Lett. 2023, 18, 2. [Google Scholar] [CrossRef]
- Lahuta, L.B. Biosynthesis of raffinose family oligosaccharides and galactosyl pinitols in developing and maturing seeds of winter vetch (Vicia villosa Roth.). Acta Soc. Bot. Pol. 2006, 75, 219–227. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Solution | Carbohydrate | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Fructose | Glucose | Sucrose | Maltose | Maltotriose | Kestose | MIN | DCI | PIN | Total | ||
| Seedling | DDW | 33.63 a | 22.90 a | 39.54 a | 5.64 a | 1.37 a | 2.05 a | 0.97 b | - | - | 106.10 a |
| MIN | 33.21 a | 21.99 a | 32.21 b | 3.82 a | 1.37 a | 2.20 a | 1.31 a | - | - | 96.10 b | |
| DCI | 34.04 a | 21.09 a | 31.77 b | 5.76 a | 2.05 a | 2.08 a | 0.99 b | 2.29 | - | 97.17 b | |
| PIN | 32.56 a | 22.55 a | 29.97 b | 5.01 a | 1.49 a | 1.74 a | 0.92 b | - | 2.92 | 100.07 b | |
| Endosperm | DDW | 0.56 a | 18.38 a | 11.97 a | 88.44 a | 19.87 a | 1.21 a | 0.55 a | - | - | 140.97 a |
| MIN | 0.69 a | 20.96 a | 13.32 a | 85.57 a | 22.37 a | 1.04 ab | 0.61 a | - | - | 144.55 a | |
| DCI | 0.44 a | 18.15 ab | 10.39 a | 84.23 a | 21.81 a | 0.91 b | 0.49 a | 0.05 | - | 109.30 b | |
| PIN | 0.43 a | 14.82 b | 11.84 a | 64.56 b | 15.84 b | 0.95 b | 0.49 a | - | 0.27 | 136.47 a | |
| Germination Solution | ||||||
|---|---|---|---|---|---|---|
| H2O | Cyclitols * | (Bio)Ag NPs | ||||
| 20 mg/L | 80 mg/L | 20 mg/L + Cyclitols * | 80 mg/L + Cyclitols * | |||
| G, % | 95.83 b | 95.56 b | 95.00 b | 98.33 a | 92.22 b | 91.39 b |
| FW, mg | 56.10 a | 45.11 a | 37.09 a | 25.07 b | 29.19 b | 26.49 b |
| DW, mg | 5.18 a | 4.74 ab | 3.94 c | 3.58 c | 5.02 a | 4.51 bc |
| WC, % | 90.76 a | 89.43 a | 89.26 a | 85.59 b | 82.61 c | 82.62 c |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lahuta, L.B.; Szablińska-Piernik, J.; Stałanowska, K.; Horbowicz, M.; Górecki, R.J.; Railean, V.; Pomastowski, P.; Buszewski, B. Exogenously Applied Cyclitols and Biosynthesized Silver Nanoparticles Affect the Soluble Carbohydrate Profiles of Wheat (Triticum aestivum L.) Seedling. Plants 2023, 12, 1627. https://doi.org/10.3390/plants12081627
Lahuta LB, Szablińska-Piernik J, Stałanowska K, Horbowicz M, Górecki RJ, Railean V, Pomastowski P, Buszewski B. Exogenously Applied Cyclitols and Biosynthesized Silver Nanoparticles Affect the Soluble Carbohydrate Profiles of Wheat (Triticum aestivum L.) Seedling. Plants. 2023; 12(8):1627. https://doi.org/10.3390/plants12081627
Chicago/Turabian StyleLahuta, Lesław B., Joanna Szablińska-Piernik, Karolina Stałanowska, Marcin Horbowicz, Ryszard J. Górecki, Viorica Railean, Paweł Pomastowski, and Bogusław Buszewski. 2023. "Exogenously Applied Cyclitols and Biosynthesized Silver Nanoparticles Affect the Soluble Carbohydrate Profiles of Wheat (Triticum aestivum L.) Seedling" Plants 12, no. 8: 1627. https://doi.org/10.3390/plants12081627
APA StyleLahuta, L. B., Szablińska-Piernik, J., Stałanowska, K., Horbowicz, M., Górecki, R. J., Railean, V., Pomastowski, P., & Buszewski, B. (2023). Exogenously Applied Cyclitols and Biosynthesized Silver Nanoparticles Affect the Soluble Carbohydrate Profiles of Wheat (Triticum aestivum L.) Seedling. Plants, 12(8), 1627. https://doi.org/10.3390/plants12081627