
Marsupella lusitanica (Gymnomitriaceae, Marchantiophyta), a New Species of Sect. Ustulatae from Mountain Ranges of Portugal


Abstract
1. Introduction
2. Results
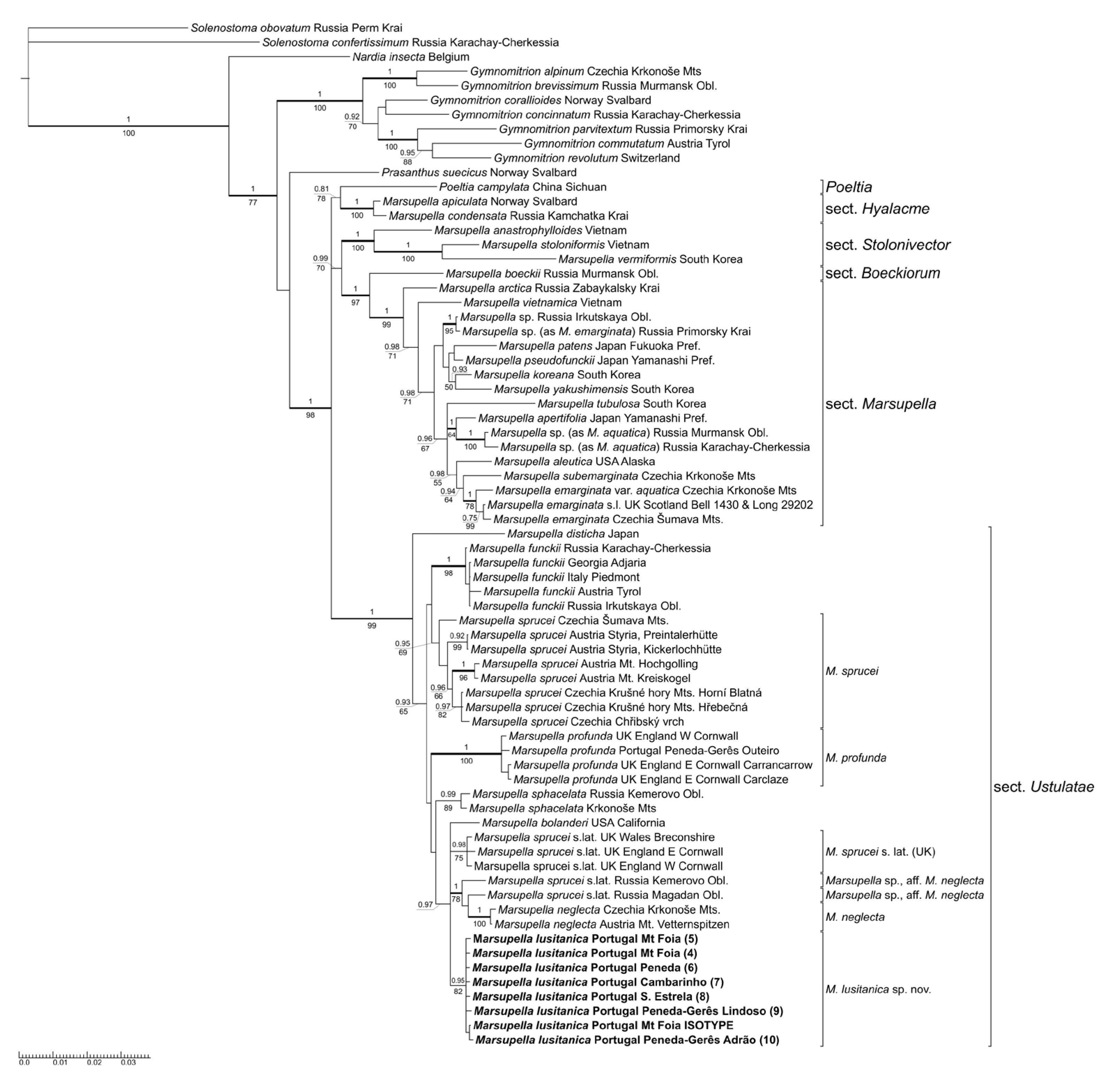
2.1. Molecular Data
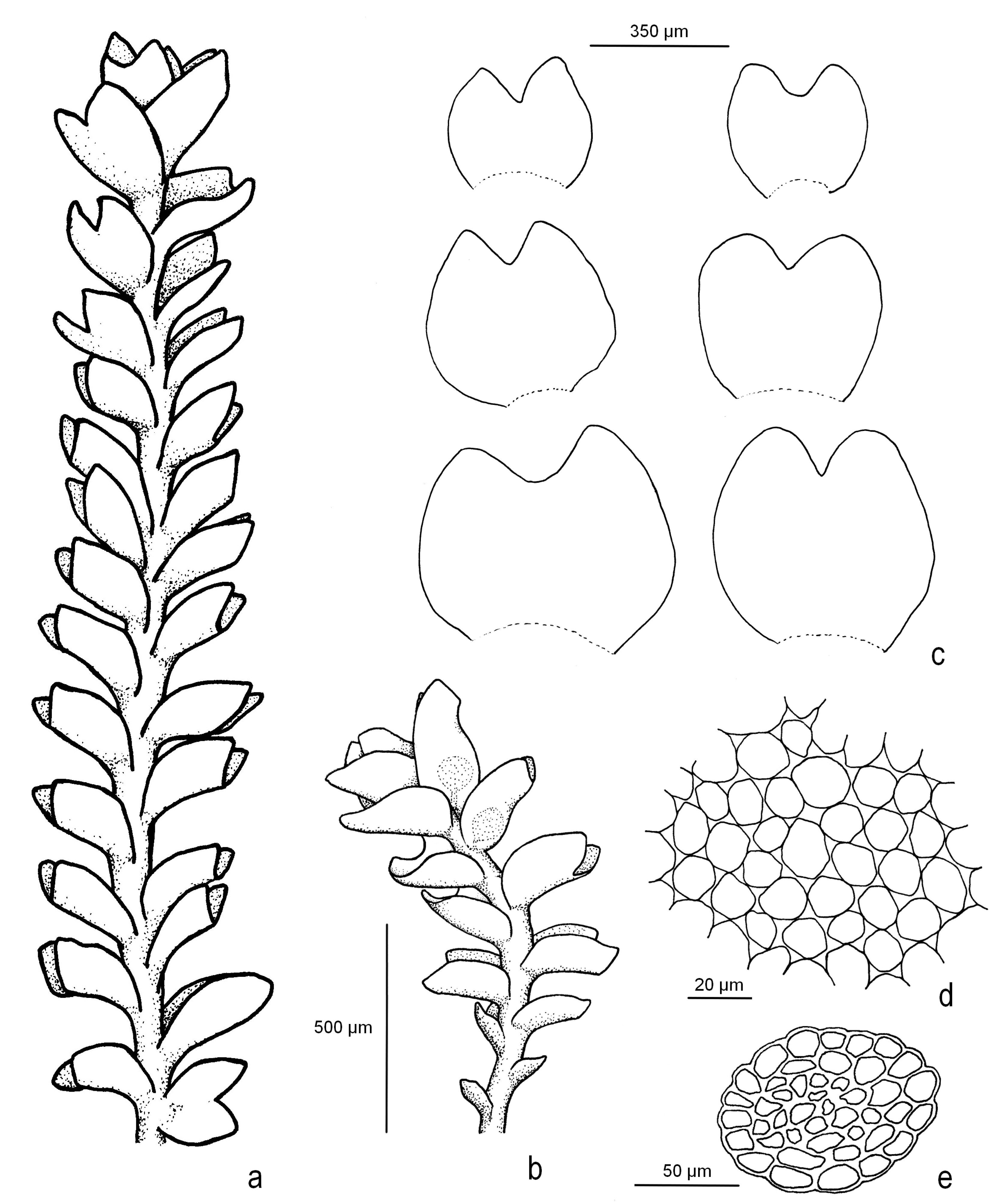
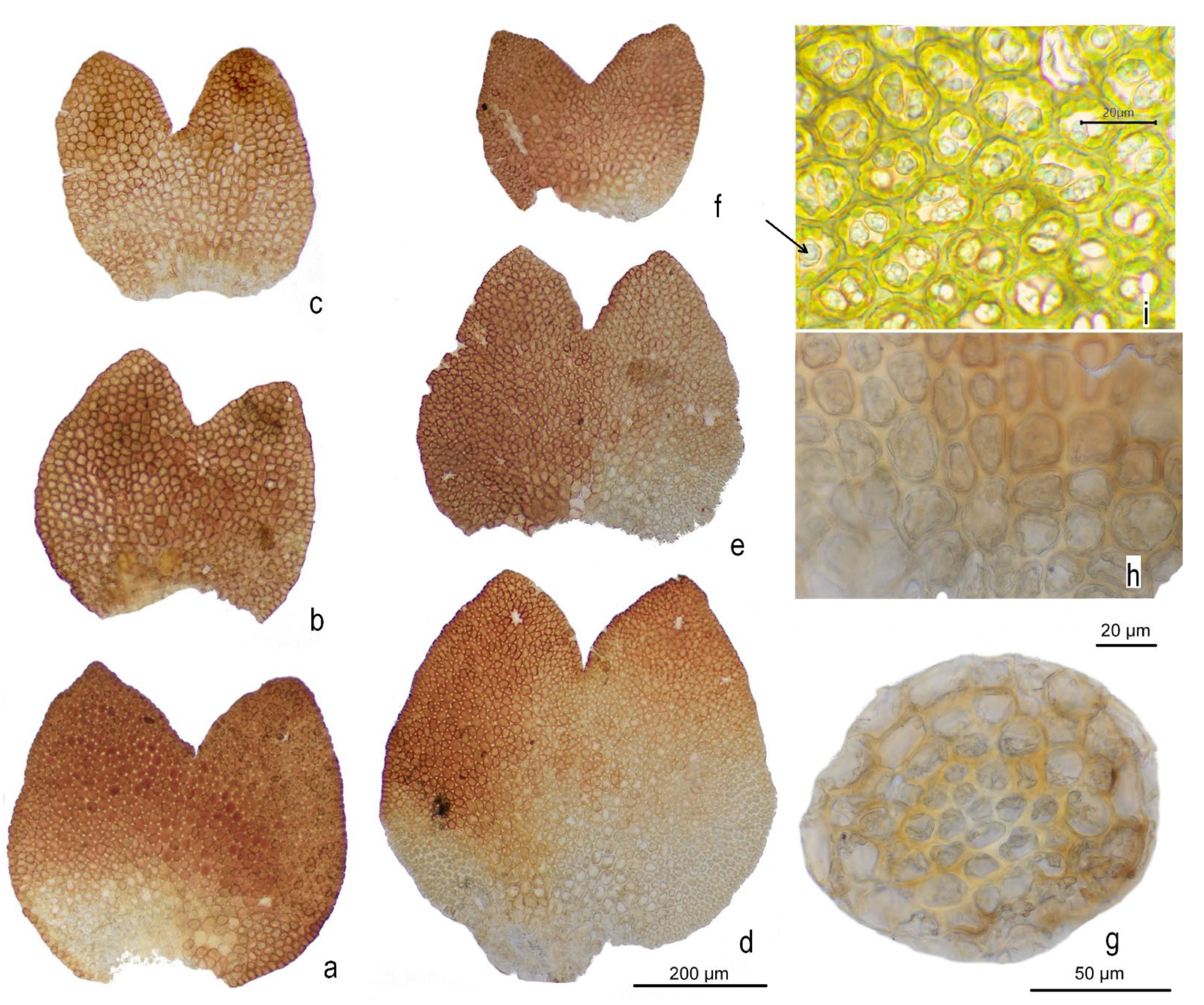
2.2. Description of the New Species
3. Discussion
3.1. Molecular Affinities
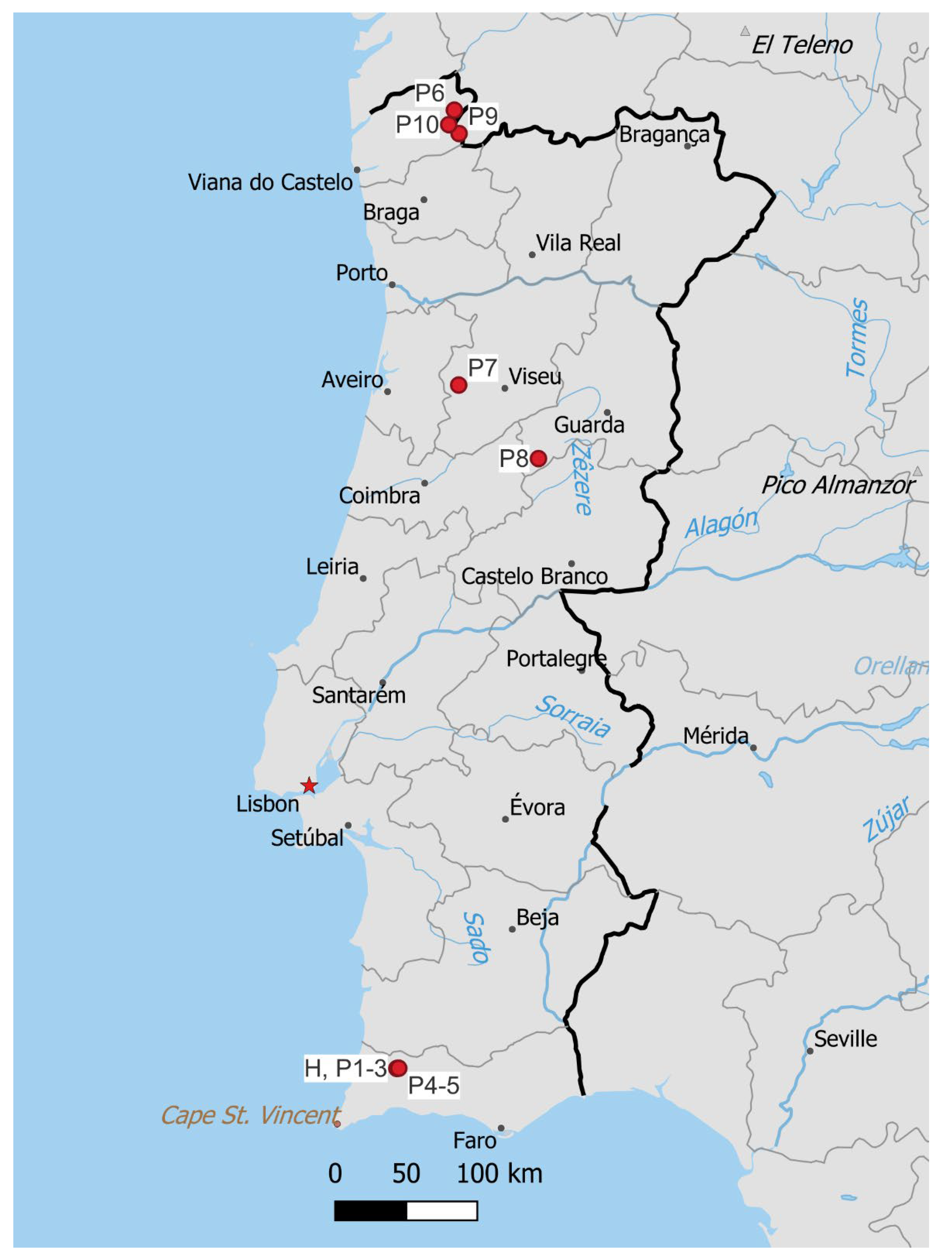
3.2. Habitat and Conservation
4. Materials and Methods
4.1. Molecular Investigation
4.2. Morphological Investigation
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Voucher Information | GenBank Accession Number | |
| ITS | trnF–trnL (trnT) | ||
| Gymnomitrion alpinum | Czech Republic: Krkonoše Mts., Úpská jáma, Kučera 21057 (CBFS) | OQ474588 | OQ507748 |
| Gymnomitrion brevissimum | Russia: Murmansk Obl., N. Konstantinova G 8171 (KPABG) | EU791833 | EU791711 |
| Gymnomitrion commutatum | Austria: Tyrol, Mt Kreuzjoch, Kučera 18862 (CBFS) | OQ474596 | OQ507758 |
| Gymnomitrion concinnatum | Russia: Caucasus, Karachay-Cherkessia, N. Konstantinova K 465a-05 (KPABG) | EU791831 | EU791710 |
| Gymnomitrion corallioides | Norway: Svalbard, N. Konstantinova 155-04 (KPABG) | EU791826 | EU791705 |
| Gymnomitrion parvitextum | Russia: Primorsky Krai, Mamontov 170-1-10 (KPABG) | MF521472 | MF521482 |
| Gymnomitrion revolutum | Switzerland, Kiebacher 2235 (LE) | OQ024235 | OQ029671 |
| Marsupella aleutica | USA: Alaska, Schofield 103958 (MO) | MH826408 | MH822632 |
| Marsupella anastrophylloides | Vietnam: Hà Giang Prov., Bakalin & Klimova, V-15-6-20 (VBGI) | OM480746 | OM489480 |
| Marsupella apertifolia | Japan: Yamanashi Pref., Bakalin J-87-25-15 (VBGI) | MH539835 | MH539892 |
| Marsupella apiculata | Norway: Svalbard, N. Konstantinova K 93-1-06 (KPABG) | EU791819 | EU791699 |
| Marsupella arctica | Russia: Zabaykalsky Krai, Kodar Range, Mamontov 2-July-2013 (CBFS) | OQ474591 | OQ507752 |
| Marsupella boeckii | Russia: Murmansk Obl., Revda, Kučera 11525 (CBFS) | OQ474590 | OQ507751 |
| Marsupella bolanderi | USA: California, Santa Barbara, Santa Yen Mts., 38802 (KPABG) | MF521463 | MF521475 |
| Marsupella condensata | Russia: Kamchatka Krai, Bakalin K-60-30-15 (VGBI) | MH539844 | MH539901 |
| Marsupella disticha | Japan: Deguchi & Yamaguchi, Bryophytes of Asia 170 (KPABG) | EU791824 | EU791703 |
| Marsupella emarginata | Czech Rep.: Šumava Mts., Mt. Plechý, Kučera 23914 (CBFS) | OQ474584 | OQ507743 |
| Marsupella emarginata | UK: Scotland, Bell 1430 | OL434465 | – |
| Marsupella emarginata var. aquatica | UK: Scotland, Long 29202 (E) | – | KF942959 |
| Marsupella emarginata var. aquatica | Czech Rep.: Krkonoše Mts., Úpská jáma, Kučera 20264 (CBFS) | OQ474583 | OQ507742 |
| Marsupella funckii | Russia: Caucasus, Karachay-Cherkessia, N. Konstantinova K 516-1-05 (KPABG) | EU791820 | EU791700 |
| Marsupella funckii | Georgia: Adjaria, Bakalin G-17-2-13 (VGBI) | MH539854 | MH539911 |
| Marsupella funckii | Italy: Piedmont, Ghiffa, Kučera 15227 (CBFS) | OQ474581 | OQ507740 |
| Marsupella funckii | Austria: Tyrol, Zillergrund, Kučera 18904 (CBFS) | OQ474587 | OQ507747 |
| Marsupella funckii | Russia: Irkutskaya Obl., Pik Taltsinskyi, Kučera 20642 (CBFS) | OQ474593 | OQ507754 |
| Marsupella koreana | South Korea: Gyeongsang Prov., Bakalin kor-23-18-15 (VGBI) | MH539850 | MH539907 |
| Marsupella lusitanica, isotype | Portugal: Algarve, Mt. Fóia, Porley s.n. 13-Sep-2018 (CBFS:20843) | OQ474573 | OQ507732 |
| Marsupella lusitanica | Portugal: Algarve, Mt. Fóia, Porley s.n. 21-Nov-2018 (CBFS:20844) | OQ474574 | OQ507733 |
| Marsupella lusitanica | Portugal: Algarve, Mt. Fóia, Porley s.n. 3-Dec-2015 (CBFS:20844) | – | OQ507746 |
| Marsupella lusitanica | Portugal: Minho, Peneda-Gerês NP, Peneda, Kučera 10535 (CBFS) | OQ474592 | OQ507753 |
| Marsupella lusitanica | Portugal: Beira Alta, Campia, Cambarinho, Kučera 10623 (CBFS) | – | OQ507756 |
| Marsupella lusitanica | Portugal: Beira Alta, Loriga, Kučera 10685 (CBFS) | OQ474595 | OQ507757 |
| Marsupella lusitanica | Portugal: Minho, Peneda-Gerês NP, Lindoso, Bell 257 (E) | LWT1248-22 | – |
| Marsupella lusitanica | Portugal: Minho, Peneda-Gerês NP, Adrão, Bell 241 (E) | LWT1269-22 | – |
| Marsupella neglecta | Czech Rep.: Krkonoše Mts., Mt. Sněžka, Kučera 18476 (CBFS) | OQ474575 | OQ507734 |
| Marsupella neglecta | Austria: Styria, Vetternspitzen, Kučera 14668 (CBFS) | OQ474594 | OQ507755 |
| Marsupella patens | Japan: Fukuoka Pref., Bakalin J-7-26a-14 (VBGI) | MH539846 | MH539903 |
| Marsupella profunda | Portugal: Minho, Peneda-Gerês, Outeiro, Bell 154 (E) | LWT1242-22 | – |
| Marsupella profunda | UK: England, vc2, Carrancarrow, Callaghan 1069 (E) | LWT1249-22 | – |
| Marsupella profunda | UK: England, vc2, Carclaze, Holyoak 09-47 | LWT1245-22 | – |
| Marsupella profunda | UK: England, vc1, Lower Bostraze, Holyoak 09-40 | LWT1246-22 | – |
| Marsupella pseudofunckii | Japan: Yamanashi Pref., Bakalin J-88-23-15 (VBGI) | MH539852 | MH539909 |
| Marsupella sp. (as M. aquatica) | Russia: Murmansk Obl., Konstantinova 152/5-87 (KPABG) | EU791813 | AF519201 |
| Marsupella sp. (as M. aquatica) | Russia: Caucasus, Karachay-Cherkessia, N. Konstantinova K 517-4-05 (KPABG) | EU791814 | EU791694 |
| Marsupella sp. (as M. emarginata) | Russia: Primorsky Krai, Bakalin & Klimova, Prim-16-14-16 (VBGI) | MH539848 | MH539905 |
| Marsupella sp. | Russia: Irkutskaya Obl., Snezhnaya River valley, Kučera 20641 (CBFS) | OQ474582 | OQ507741 |
| Marsupella sphacelata | Russia: Kemerovo Obl., N. Konstantinova 65/1-00 (KPABG) | EU791821 | AF519200 |
| Marsupella sphacelata | Czech Rep.: Krkonoše Mts., Úpská jáma, Kučera 21077 (CBFS) | OQ474597 | OQ507759 |
| Marsupella sprucei | Czech Rep.: Šumava Mts., Mt. Plechý, F. Müller s.n. 3-October-2007 (DR, dupl. CBFS:20918) | OQ474576 | OQ507735 |
| Marsupella sprucei | Czech Rep.: Krušné hory Mts., Horní Blatná, F. Müller s.n. 18-September-2018 (DR, dupl. CBFS:20919) | OQ474577 | OQ507736 |
| Marsupella sprucei | Czech Rep.: Krušné hory Mts., Hřebečná, S. Biedermann s.n. 18-September-2018 (DR, dupl. CBFS:20920) | OQ474578 | OQ507737 |
| Marsupella sprucei | Czech Rep.: Děčín distr., Chřibský vrch, F. Müller s.n. 17-July-2019 (DR, dupl. CBFS:20965) | OQ474579 | OQ507738 |
| Marsupella sprucei | Austria: Styria, Preintalerhütte, Köckinger 15430 (CBFS) | OQ474585 | OQ507744 |
| Marsupella sprucei | Austria: Styria, Kickerlochhütte, Köckinger 15429 (CBFS) | OQ474586 | OQ507745 |
| Marsupella sprucei | Austria: Salzburg, Mt. Hochgolling, Kučera 9363 (CBFS) | OQ474589 | OQ507749 |
| Marsupella sprucei | Austria: Styria, Mt. Kreiskogel, Kučera 6391 (CBFS) | – | OQ507750 |
| Marsupella sprucei s.lat. | Russia: Kemerovo Obl., N. Konstantinova 54-1-00 (KPABG) | EU791823 | HQ833031 |
| Marsupella sprucei s.lat. | Russia: Magadan Obl., Bakalin mag-38-39-11 (VGBI) | MH539856 | MH539913 |
| Marsupella sprucei s.lat. | UK: England, vc1, Bakers’ Pit, Bell 531 (E) | LWT1261-22 | – |
| Marsupella sprucei s.lat. | UK: England, vc2, St. Neot, Holyoak 99-465 | LWT1259-22 | – |
| Marsupella sprucei s.lat. | UK: Wales, vc42, Lower Neuadd Res., Bell 566 (E) | LWT1262-22 | – |
| Marsupella stoloniformis | Vietnam: Lao Cai Prov., Bakalin & Klimova, V-11-11-17 (VBGI) | MH539859 | MH539916 |
| Marsupella subemarginata | Czech Rep.: Krkonoše Mts., Mt. Kotel, Kučera 23513 (CBFS) | OQ474580 | OQ507739 |
| Marsupella vermiformis | South Korea: Jeju Prov., Choi 120911 (VBGI) | MH539857 | MH539914 |
| Marsupella vietnamica | Vietnam: Lao Cai Prov., Bakalin, V-2-101-16 (VBGI) | MH539862 | MH539919 |
| Marsupella tubulosa | South Korea, South Gyeongsang Prov., Bakalin Kor-23-15-15 (VBGI) | MH539861 | MH539918 |
| Marsupella yakushimensis | South Korea, Gangwon Prov., Choi 8347 (VBGI) | MH539864 | MH539921 |
| Nardia insecta | Belgium, N. Konstantinova 102077 (KPABG) | EU791836 | EU791714 |
| Poeltia campylata | China: Sichuan, Bakalin 48-2-17, 37210 (VGBI) | MH580596 | MH580593 |
| Prasanthus suecicus | Norway: Svalbard, N. Konstantinova K 121-5-06 (KPABG) | EU791825 | EU791704 |
| Solenostoma confertissimum | Russia: Caucasus, Karachay-Cherkessia, N. Konstantinova K 459-8a-05 (KPABG) | GQ220774 | GQ220758 |
| Solenostoma obovatum | Russia: Perm Krai, N. Konstantinova K 324-1-04 (KPABG) | GQ220771 | GQ220755 |
References
- Söderström, L.; Hagborg, A.; von Konrat, M.; Bartholomew-Began, S.; Bell, D.; Briscoe, L.; Brown, E.; Cargill, D.C.; da Costa, D.P.; Crandall-Stotler, B.J.; et al. World checklist of hornworts and liverworts. PhytoKeys 2016, 59, 1–828. [Google Scholar] [CrossRef] [PubMed]
- Mamontov, Y.S.; Vilnet, A.A.; Konstantinova, N.A.; Bakalin, V.A. Two new species of Gymnomitriaceae (Marchantiophyta) in the North Pacific. Bot. Pac. 2019, 8, 67–80. [Google Scholar] [CrossRef]
- Bakalin, V.A.; Fedosov, V.E.; Fedorova, A.V.; Nguyen, V.S. Integrative Taxonomic Revision of Marsupella (Gymnomitriaceae, Hepaticae) Reveals Neglected Diversity in Pacific Asia. Cryptogam. Bryol. 2019, 40, 59–85. [Google Scholar] [CrossRef]
- Bakalin, V.A.; Vilnet, A.A.; Mamontov, Y.S.; Schäfer-Verwimp, A.; Maltseva, Y.D.; Klimova, K.G.; Nguyen, V.S.; Choi, S.S. Stolonicaulon: A Section-Puzzle within Marsupella (Gymnomitriaceae, Marchantiophyta). Plants 2022, 11, 1596. [Google Scholar] [CrossRef] [PubMed]
- Hodgetts, N.G.; Söderström, L.; Blockeel, T.L.; Caspari, S.; Ignatov, M.S.; Konstantinova, N.A.; Lockhart, N.; Papp, B.; Schröck, C.; Sim-Sim, M.; et al. An annotated checklist of bryophytes of Europe, Macaronesia and Cyprus. J. Bryol. 2020, 42, 1–116. [Google Scholar] [CrossRef]
- Sérgio, C.; Garcia, C.; Sim-Sim, M.; Vieira, C.; Hespanhol, H.; Stow, S. Atlas e Livro Vermelho dos Briófitos Ameaçados de Portugal; MUHNAC, Documenta: Lisboa, Spain, 2013. [Google Scholar]
- Vilnet, A.A.; Konstantinova, N.A.; Troitsky, A.V. Molecular insight on phylogeny and systematics of the Lophoziaceae, Scapaniaceae, Gymnomitriaceae and Jungermanniaceae. Arctoa 2010, 19, 31–50. [Google Scholar] [CrossRef]
- Porley, R.; Papp, B.; Söderström, L.; Hallingbäck, T. European Bryophyte Conservation in the New Millennium; University of Malaya: Kuala Lumpur, Malaysia, 2008. [Google Scholar]
- Paton, J.A. The Liverwort Flora of the British Isles; Harley Books: Colchester, UK, 1999; ISBN 978-0-946589-60-9. [Google Scholar]
- Bell, D.; Long, D.; Hollingsworth, P. The Use of DNA Barcoding to Address Major Taxonomic Problems for Rare British Bryophytes; Final Revised Report; Royal Botanic Garden Edinburgh: Edinburgh, UK, 2013. [Google Scholar]
- Dayrat, B. Towards integrative taxonomy. Biol. J. Linn. Soc. 2005, 85, 407–415. [Google Scholar] [CrossRef]
- Ignatov, M.S.; Porley, R.D.; Ochyra, R.; Kuznetsova, O.I.; Ignatova, E.A. Coscinodon monchiquensis R.D. Porley, Ochyra & Ignatova (Grimmiaceae), a new species from the Algarve, southern Portugal. J. Bryol. 2018, 40, 125–136. [Google Scholar] [CrossRef]
- Porley, R.D.; Fedosov, V.; Plášek, V.; Fedorova, A. Undiscovered Biodiversity of the European Moss Flora: Neodicranella hamulosa (Aongstroemiaceae), a New Genus and Species from SW Portugal. Plants 2021, 10, 2289. [Google Scholar] [CrossRef]
- Garcia, C.; Sérgio, C.; Jansen, J. The bryophyte flora of the natural park of Serra da Estrela (Portugal): Conservation and Biogeographical approaches. Cryptogam. Bryol. 2008, 29, 49–73. [Google Scholar]
- Sérgio, C.; Garcia, C.; Hespanhol, H.; Vieira, C.; Stow, S.; Long, D. Diversidad de briófitos en el Parque Nacional Peneda-Gerês (Portugal): Seleccionando áreas importantes de plantas “Important Plant Areas (IPA)” basado en nuevos datos y anteriores referencias. Bot. Complut. 2012, 36, 39–50. [Google Scholar] [CrossRef]
- Sérgio, C.; Schumacker, R. Contribuiçao para o estudo da flora briológica do Parque Nacional da Peneda-Gerês. Port. Acta Biol. 1992, 16, 107–137. [Google Scholar]
- Hespanhol, H.; Vieira, C.; Lomba, Â.; Séneca, A. New data on bryophyte distribution in the Peneda-Gerês National Park (NW Portugal): The use of GIS for conservation remarks. Bol. Soc. Esp. Briol. 2005, 26–27, 59–74. [Google Scholar]
- Sérgio, C.; Silva, I. The bryophyte flora of the Vouga region (Aveiro, Beira Litoral). Port. Acta Biol. 2007, 22, 159–188. [Google Scholar]
- Simmons, M.P.; Ochoterena, H. Gaps as Characters in Sequence-Based Phylogenetic Analyses. Syst. Biol. 2000, 49, 369–381. [Google Scholar] [CrossRef]
- Grolle, R. Miscellanea Hepaticologica 151–160. Lindbergia 1975, 3, 47–56. [Google Scholar]
- Váňa, J.; Söderström, L.; Hagborg, A.; Von Konrat, M.; Engel, J.J. Early Land Plants Today: Taxonomy, systematics and nomenclature of Gymnomitriaceae. Phytotaxa 2010, 11, 1. [Google Scholar] [CrossRef]
- Müller, K. Die Lebermoose Deutschlands, Oesterreichs und der Schweiz, 2nd ed.; Dr. L. Rabenhorst’s Kryptogamen-Flora von Deutschland, Oesterreich und der Schweiz; E. Kummer: Leipzig, Germany, 1906–1911; Volume 6. [Google Scholar]
- Müller, K. Die Lebermoose Europas (Musci hepatici), 3rd ed.; Dr. L. Rabenhorst’s Kryptogamen-Flora von Deutschland, Oesterreich und der Schweiz; Geest & Portig: Leipzig, Germany, 1956; Volume 6. [Google Scholar]
- Schiffner, V.F. Kritische Bemerkungen über die europäische Lebermoose mit Bezug auf die Exemplare des Exsiccatenwerkes: Hepaticae europaeae exsiccatae. I. Serie. Sitzungsber. Deutsch. Naturwiss. Med. Ver. Böhmen “Lotos” Prag 1901, 49, 75–130. [Google Scholar]
- Limpricht, K.G. Über neue Arten und Formen der Gattung Sarcoscyphus Corda. Jahresber. Schles. Ges. Vaterländische Cult. 1880, 58, 179–184. [Google Scholar]
- Paton, J.A. Marsupella profunda Lindb in Cornwall, new to the British Isles. J. Bryol. 1990, 16, 1–4. [Google Scholar] [CrossRef]
- Konstantinova, N.A. Marsupella. In FNA Editors, Gymnomitriaceae, Bryophyte Flora of North America; Provisional Publication, 2020. Available online: http://www.mobot.org/plantscience/bfna/V3/Gymnomitriaceae.htm (accessed on 18 March 2023).
- Schiffner, V. Studien über kritische Arten der Gattungen Gymnomitrium und Marsupella. (Fortsetzung.). Oesterr. Bot. Z. 1903, 53, 166–172. [Google Scholar] [CrossRef]
- Damsholt, K. Illustrated Flora of Nordic Liverworts and Hornworts; Nordic Bryological Society: Lund, Sweden, 2002; ISBN 87-986781-2-4. [Google Scholar]
- Damsholt, K. The Liverworts of Greenland; Nordic Bryological Society: Lund, Sweden, 2013; ISBN 978-91-637-2514-2. [Google Scholar]
- Váňa, J. Notes on the genus Marsupella s. lat. (Gymnomitriaceae, Hepaticae) 1–10. Infrageneric taxa. Bryobrothera 1999, 5, 221–229. [Google Scholar]
- Grolle, R. Verzeichnis der Lebermoose Europas und benachbarter Gebiete. Feddes Repert. 1976, 76, 171–279. [Google Scholar] [CrossRef]
- Duda, J.; Váňa, J. Rozšíření játrovek v Československu—XXXVII [Die Verbreitung der Lebermoose in der Tschechoslowakei—XXXVII]. Časopis Slez. Muz., Sér. A, Vědy Přír. 1983, 32, 97–110. [Google Scholar]
- Malato Beliz, J. A Serra de Monchique: FLORA e Vegetação; Parques Naturais; Serviço Nacional de Parques Reservas e Património Paisagístico: Lisboa, Portugal, 1982. [Google Scholar]
- González-Clavijo, E.J.; Valadares, V. A Estrutura do Complexo de Monchique. In Proceedings of the Actas do VI Congresso Nacional de Geologia Ciências da Terra (UNL), Lisboa, Portugal, 4–6 June 2003; Volume No. Especial 5, pp. 37–40. [Google Scholar]
- Rock, N.M.S. Petrology and Petrogenesis of the Monchique Alkaline Complex, Southern Portugal. J. Petrol. 1978, 19, 171–214. [Google Scholar] [CrossRef]
- Bernard-Griffiths, J.; Gruau, G.; Cornen, G.; Azambre, B.; Mace, J. Continental Lithospheric Contribution to Alkaline Magmatism: Isotopic (Nd, Sr, Pb) and Geochemical (REE) Evidence from Serra de Monchique and Mount Ormonde Complexes. J. Petrol. 1997, 38, 115–132. [Google Scholar] [CrossRef]
- Lara, F.; Porley, R.D.; Draper, I.; Aleffi, M.; Garcia, C.; Garilleti, R. Ulota s.l. (Orthotrichaceae, Bryidae) at southernmost Mediterranean localities: Not a simple matter. Pl. Biosyst. 2022, 156, 1448–1455. [Google Scholar] [CrossRef]
- Hill, M.O.; Preston, C.D. The geographical relationships of British and Irish bryophytes. J. Bryol. 1998, 20, 127–226. [Google Scholar] [CrossRef]
- Patiño, J.; Bisang, I.; Goffinet, B.; Hedenäs, L.; McDaniel, S.; Pressel, S.; Stech, M.; Ah-Peng, C.; Bergamini, A.; Caners, R.T.; et al. Unveiling the nature of a miniature world: A horizon scan of fundamental questions in bryology. J. Bryol. 2022, 44, 1–34. [Google Scholar] [CrossRef]
- Pörtner, D.C.; Roberts, M.; Tignor, E.S.; Poloczanska, K.; Mintenbeck, A.; Alegría, M.; Craig, S.; Langsdorf, S.; Löschke, V.; Möller, A.; et al. IPCC Climate Change 2022: Impacts, Adaptation, and Vulnerability. Contribution of Working Group II to the Sixth Assessment Report of the Intergovernmental Panel on Climate Change; Cambridge University Press: Cambridge, UK; New York, NY, USA, 2022. [Google Scholar]
- Mitchell, R.J.; Simonson, W.; Flegg, L.A.; Santos, P.; Hall, J. A comparison of the resilience of four habitats to fire, and the implications of changes in community composition for conservation: A case study from the Serra de Monchique, Portugal. Plant Ecol. Divers. 2009, 2, 45–56. [Google Scholar] [CrossRef]
- Tedim, F.; Leone, V.; Amraoui, M.; Bouillon, C.; Coughlan, M.; Delogu, G.; Fernandes, P.; Ferreira, C.; McCaffrey, S.; McGee, T.; et al. Defining Extreme Wildfire Events: Difficulties, Challenges, and Impacts. Fire 2018, 1, 9. [Google Scholar] [CrossRef]
- Sérgio, C.; Bergamini, A.; Garcia, C.; Garilleti, R.; Infante, M.; Porley, R.D. Marsupella profunda. The IUCN Red List of Threatened Species 2019: E.T39195A87726839 2019. Available online: https://www.gbif.org/fr/species/176752393 (accessed on 18 March 2023). [CrossRef]
- Kučera, J.; Kuznetsova, O.I.; Manukjanová, A.; Ignatov, M.S. A phylogenetic revision of the genus Hypnum: Towards completion. Taxon 2019, 68, 628–660. [Google Scholar] [CrossRef]
- Taberlet, P.; Gielly, L.; Pautou, G.; Bouvet, J. Universal primers for amplification of three non-coding regions of chloroplast DNA. Plant. Mol. Biol. 1991, 17, 1105–1109. [Google Scholar] [CrossRef]
- Katoh, K.; Rozewicki, J.; Yamada, K.D. MAFFT online service: Multiple sequence alignment, interactive sequence choice and visualization. Brief. Bioinform. 2019, 20, 1160–1166. [Google Scholar] [CrossRef]
- Ronquist, F.; Teslenko, M.; van der Mark, P.; Ayres, D.L.; Darling, A.; Höhna, S.; Larget, B.; Liu, L.; Suchard, M.A.; Huelsenbeck, J.P. MrBayes 3.2: Efficient Bayesian Phylogenetic Inference and Model Choice Across a Large Model Space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef]
- Stamatakis, A. RAxML version 8: A tool for phylogenetic analysis and post-analysis of large phylogenies. Bioinformatics 2014, 30, 1312–1313. [Google Scholar] [CrossRef]
- Stöver, B.C.; Müller, K.F. TreeGraph 2: Combining and visualizing evidence from different phylogenetic analyses. BMC Bioinform. 2010, 11, 7. [Google Scholar] [CrossRef]
- Schuster, R.M. The Hepaticae and Anthocerotae of North America; Columbia University Press: New York, NY, USA; London, UK, 1974; Volume III. [Google Scholar]







| Character | M. sprucei s.lat. | M. funckii s.lat. | M. profunda | M. lusitanica |
|---|---|---|---|---|
| Leaf shape | Distinctly longer than wide to slightly wider than long | Slightly to distinctly wider than long, seldom longer than wide | Mostly longer than wide | Mostly longer than wide |
| Sinus angle | 50–100° | 50–70° | (20–)40–60(–80)° | 45–80° at base of sinus, 100–135°at lobe tips |
| Sinus depth | (1/5–)1/4–1/3(–1/2) | 1/3–1/2 (M. funckii s.str.,), 1/4–1/5 (M. badensis) | (1/5–)1/3–2/5 | 1/5–1/3(–2/5) of leaf length |
| Lobe apex | Acute to narrowly rounded, or seldom ovate and more broadly rounded | Acute or subacute | Obtuse or subobtuse, occ. subacute, often broadly ovate or lingulate | Subacute to occasionally obtuse |
| Leaf cell size | 12–18 μm (M. neglecta), 20–28 µm (M. sprucei) | 12–18 μm (to 25 µm in M. pygmaea, to 30 µm in M. ramosa) | 12–19 μm | (10)12–19(24) μm |
| Leafy shoot width | 0.3–0.6(0.8) mm | 0.3–0.6(0.8) mm (–1.1 mm in M. badensis) | 0.4–1.0 mm | 0.4–1.0 mm |
| Sexuality | Paroicous (rarely synoicous) | Dioicous | Paroicous | Dioicous |
| Color | Brownish green to dark brown, red or reddish black, seldom almost black | Brownish green to dark brown, reddish brown or almost black | Brownish green to dark brown, red or reddish black, seldom almost black and glossy when dry | Brown to brownish-ochre, new growth yellowish-green, to reddish-black and glossy when dry |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Porley, R.D.; Bell, D.; Kučera, J. Marsupella lusitanica (Gymnomitriaceae, Marchantiophyta), a New Species of Sect. Ustulatae from Mountain Ranges of Portugal. Plants 2023, 12, 1468. https://doi.org/10.3390/plants12071468
Porley RD, Bell D, Kučera J. Marsupella lusitanica (Gymnomitriaceae, Marchantiophyta), a New Species of Sect. Ustulatae from Mountain Ranges of Portugal. Plants. 2023; 12(7):1468. https://doi.org/10.3390/plants12071468
Chicago/Turabian StylePorley, Ronald D., David Bell, and Jan Kučera. 2023. "Marsupella lusitanica (Gymnomitriaceae, Marchantiophyta), a New Species of Sect. Ustulatae from Mountain Ranges of Portugal" Plants 12, no. 7: 1468. https://doi.org/10.3390/plants12071468
APA StylePorley, R. D., Bell, D., & Kučera, J. (2023). Marsupella lusitanica (Gymnomitriaceae, Marchantiophyta), a New Species of Sect. Ustulatae from Mountain Ranges of Portugal. Plants, 12(7), 1468. https://doi.org/10.3390/plants12071468