
Conjoining Trees for the Provision of Living Architecture in Future Cities: A Long-Term Inosculation Study

 ,
,  , ,
, ,  , ,
, ,  and
and 
Abstract
1. Introduction
1.1. Conjoined Trees as Architectural Structures
1.2. State of the Art on Inosculation Studies
1.3. Inosculations Form Functional Units
1.4. Aim of the Study
2. Material and Methods
2.1. Plant Material
2.2. Morphometric Studies
2.3. Macroscopic Anatomical Studies and 3D Reconstruction
2.4. Micro-CT Scans
2.5. Staining of Water-Conducting Tissues
2.6. Statistics
3. Results
3.1. Morphometric Analyses
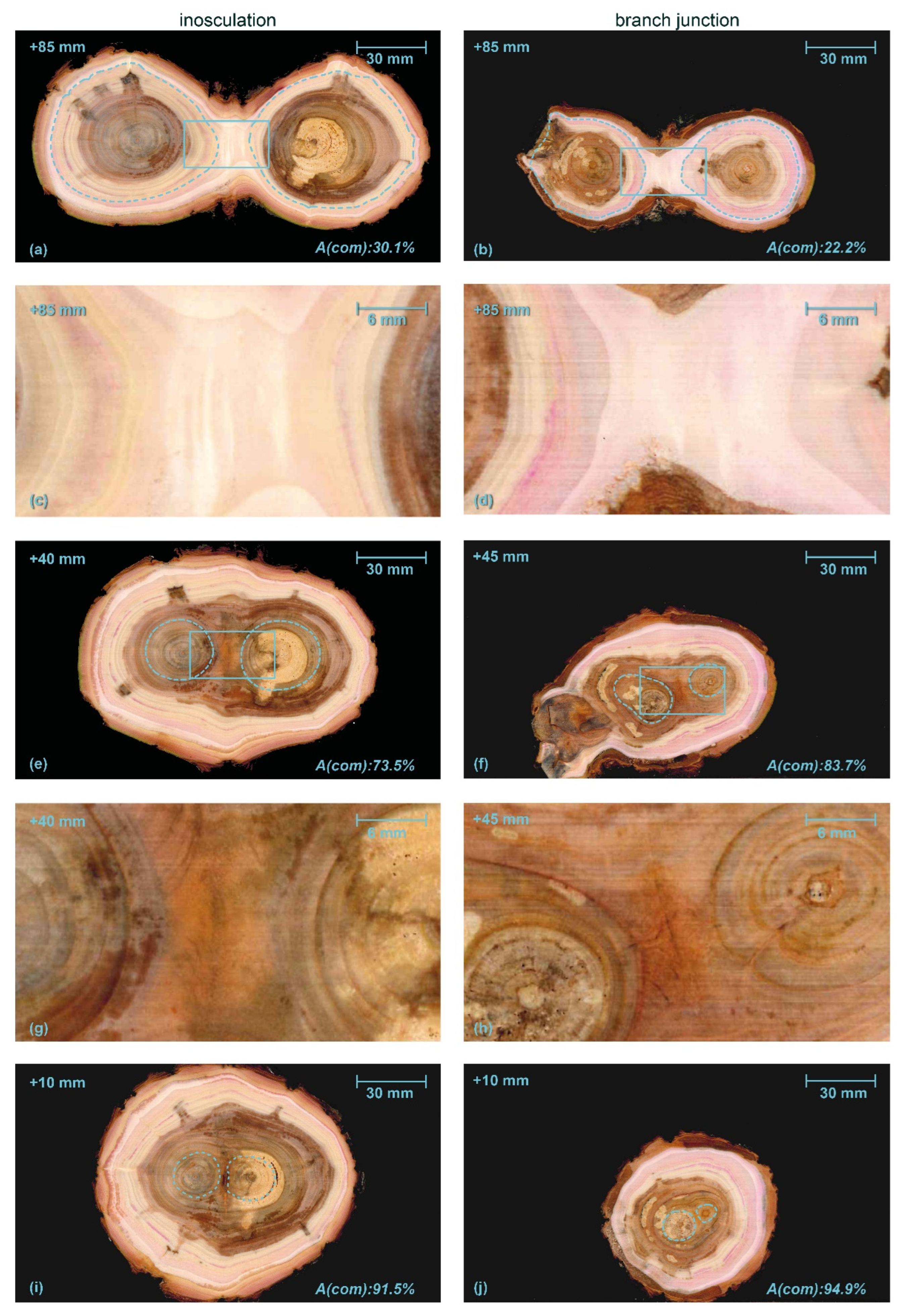
3.2. Macroscopic Anatomical Analyses
3.3. Volumetric 3D Reconstruction
3.4. Micro-CT Analyses
4. Discussion
4.1. Morphometric Measurements as a Tool for Inosculation Assessment
4.1.1. Theoretical Scenarios of Inosculation Morphology
4.1.2. Morphology of Salix alba and Platanus × hispanica Inosculations
4.2. Comparison of Artificial Inosculations and Natural Branch Junctions
4.2.1. Similarities between Inosculations and Branch Junctions
4.2.2. Dissimilarities between Inosculations and Branch Junctions
4.3. Limitations and Opportunities of Conjoined Trees
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- United Nations. Transforming Our World: The 2030 Agenda for Sustainable Development: A/RES/70/1. 2015. Available online: https://www.un.org/en/development/desa/population/migration/generalassembly/docs/globalcompact/A_RES_70_1_E.pdf (accessed on 17 October 2022).
- Ludwig, F. Baubotanik: Designing with Living Material. In Materiality and Architecture; Löschke, S., Ed.; Routledge: Milton Park, UK, 2016; pp. 206–216. [Google Scholar]
- Kirsch, K. Naturbauten aus Lebenden Gehölzen; Organischer Landbau-Verlag Kurt Walter Lau: Xanten, Germany, 1996. [Google Scholar]
- Arbona, J.; Greden, L.; Joachim, M. Nature’s Technology: The Fab Tree Hab House. Thresholds 2003, 26, 48–53. [Google Scholar] [CrossRef]
- Reams, R. Arborsculpture. Solutions for a Small Planet; Arborsmith Studios: Williams, OR, USA, 2005. [Google Scholar]
- Wang, X.; Gard, W.; Borska, H.; Ursem, B.; van de Kuilen, J. Vertical greenery systems: From plants to trees with self-growing interconnections. Eur. J. Wood Wood Prod. 2020, 78, 1031–1043. [Google Scholar] [CrossRef]
- Bowler, D.E.; Buyung-Ali, L.; Knight, T.M.; Pullin, A.S. Urban greening to cool towns and cities: A systematic review of the empirical evidence. Landsc. Urban Plan. 2010, 97, 147–155. [Google Scholar] [CrossRef]
- Nowak, D.J.; Crane, D.E. Carbon storage and sequestration by urban trees in the USA. Environ. Pollut. 2002, 116, 381–389. [Google Scholar] [CrossRef] [PubMed]
- Kuehler, E.; Hathaway, J.; Tirpak, A. Quantifying the benefits of urban forest systems as a component of the green infrastructure stormwater treatment network. Ecohydrology 2017, 10, e1813. [Google Scholar] [CrossRef]
- Rötzer, T.; Rahman, M.A.; Moser-Reischl, A.; Pauleit, S.; Pretzsch, H. Process based simulation of tree growth and ecosystem services of urban trees under present and future climate conditions. Sci. Total Environ. 2019, 676, 651–664. [Google Scholar] [CrossRef]
- Rahman, M.A.; Stratopoulos, L.M.F.; Moser-Reischl, A.; Zölch, T.; Häberle, K.-H.; Rötzer, T.; Pretzsch, H.; Pauleit, S. Traits of trees for cooling urban heat islands: A meta-analysis. Build. Environ. 2020, 170, 106606. [Google Scholar] [CrossRef]
- Ludwig, F.; Schönle, D.; Bellers, M. Klimaaktive baubotanische Stadtquartiere, Bautypologien und Infrastrukturen: Modellprojekte und Planungswerkzeuge. Klimopass-Berichte. LUBW Landesanstalt für Umwelt, Messungen und Naturschutz Baden-Württemberg. 2015. Available online: https://pd.lubw.de/91679 (accessed on 18 October 2022).
- Ludwig, F. Baubotanik—Lebendarchitektur. In Lebende Bauten−Trainierbare Tragwerke; De Bruyn, G., Ludwig, F., Schwertfeger, H., Eds.; Lit: Berlin, Germany, 2009; pp. 165–196. [Google Scholar]
- Ludwig, F. Baubotanik—Möglichkeiten und Grenzen des Konstruierens lebender Tragwerke. In Konstruktion und Gestalt—Leichte Konstruktionen; Baier, B., Koenen, R., Müller, J., Schmerbach, S., Eds.; Druck & Verlagshaus Mainz: Aachen, Germany, 2008; pp. 59–85. [Google Scholar]
- Ludwig, F. Botanische Grundlagen der Baubotanik und deren Anwendung im Entwurf. Ph.D. Thesis, University of Stuttgart, Stuttgart, Germany, 2012. [Google Scholar]
- Ludwig, F.; Middleton, W.; Vees, U. Baubotanik: Living Wood and Organic Joints. In Rethinking Wood—Future Dimensions of Timber Assembly; Birkhäuser: Basel, Switzerland, 2019; pp. 262–275. [Google Scholar]
- Ludwig, F.; Middleton, W.; Gallenmüller, F.; Rogers, P.; Speck, T. Living bridges using aerial roots of Ficus elastica—An interdisciplinary perspective. Sci. Rep. 2019, 9, 12226. [Google Scholar] [CrossRef]
- Middleton, W.; Habibi, A.; Shankar, S.; Ludwig, F. Characterizing Regenerative Aspects of Living Root Bridges. Sustainability 2020, 12, 3267. [Google Scholar] [CrossRef]
- Graefe, R. Geleitete Linden. Daidalos Z. Für Archit. Kunst Kult. 1987, 23, 17–29. [Google Scholar]
- Graefe, R. Bauten aus Lebenden Bäumen; Geymüller: Aachen, Germany, 2014. [Google Scholar]
- Ludwig, F. Lebende Konstruktionen—Eine historische Einführung in die Baubotanik. nodium 2018, 10, 46–49. [Google Scholar]
- Cattle, C. Grown furniture: A move towards design for sustainability. Ph.D. Thesis, Buckinghamshire New University, High Wycombe, UK, 2010. [Google Scholar]
- Harris, R.W. Arboriculture: Integrated Management of Landscape Trees, Shrubs, and Vines, 2nd ed.; Prentice-Hall International: Upper Sadle River, NY, USA, 1992. [Google Scholar]
- Roloff, A. Baumpflege; Ulmer: Stuttgart, Germany, 2013. [Google Scholar]
- Wang, X.; Gard, W.; de Vries, N.; van de Kuilen, J.-W. Anatomical and Mechanical Features of Self-Growing Connections in Plants. preprint 2022. [Google Scholar] [CrossRef]
- Graham, B.; Bormann, F. Natural root grafts. Bot. Rev. 1966, 32, 255–292. [Google Scholar] [CrossRef]
- Bormann, F. The structure, function, and ecological significance of root grafts in Pinus strobus L. Ecol. Monogr. 1966, 36, 2–26. [Google Scholar] [CrossRef]
- Tarroux, E.; DesRochers, A.; Tremblay, F. Molecular analysis of natural root grafting in jack pine (Pinus banksiana) trees: How does genetic proximity influence anastomosis occurrence? Tree Genet. Genomes 2014, 10, 667–677. [Google Scholar] [CrossRef]
- Tarroux, E.; DesRochers, A. Effect of natural root grafting on growth response of jack pine (Pinus banksiana; Pinaceae). Am. J. Bot. 2011, 98, 967–974. [Google Scholar] [CrossRef]
- Basnet, K.; Scatena, F.; Likens, G.E.; Lugo, A.E. Ecological consequences of root grafting in tabonuco (Dacryodes excelsa) trees in the Luquillo Experimental Forest, Puerto Rico. Biotropica 1993, 25, 28–35. [Google Scholar] [CrossRef]
- Bormann, F.; Graham, B.F. The occurrence of natural root grafting in eastern white pine, Pinus strobus L., and its ecological implications. Ecology 1959, 40, 677–691. [Google Scholar] [CrossRef]
- Millner, E.M. Natural grafting in Hedera helix. New Phytol. 1932, 31, 2–25. [Google Scholar] [CrossRef]
- Harrington, M.J.; Speck, O.; Speck, T.; Wagner, S.; Weinkamer, R. Biological archetypes for self-healing materials. In Self-Healing Materials. Advances in Polymer Science; Hager, M., van der Zwaag, S., Schubert, U., Eds.; Springer: Cham, Switzerland, 2015; Volume 273, pp. 307–344. [Google Scholar]
- Shinozaki, K.; Yoda, K.; Hozumi, K.; Kira, T. A quantitative Analysis of Plant Form—The Pipe Model Theory: I. Basic Analyses. Jpn. J. Ecol. 1964, 14, 97–105. [Google Scholar]
- Shinozaki, K.; Yoda, K.; Hozumi, K.; Kira, T. A quantitative Analysis of Plant Form—The Pipe Model Theory: II. Further Evidence of the Theory and its Application in Forest Ecology. Jpn. J. Ecol. 1964, 14, 133–139. [Google Scholar]
- Lehnebach, R.; Beyer, R.; Letort, V.; Heuret, P. The pipe model theory half a century on: A review. Ann. Bot. 2018, 121, 773–795. [Google Scholar] [CrossRef] [PubMed]
- Godin, C. Representing and encoding plant architecture: A review. Ann. For. Sci. 2000, 57, 413–438. [Google Scholar] [CrossRef]
- Perttunen, J.; Änen, R.S.; Nikinmaa, E.; Salminen, H.; Saarenmaa, H. LIGNUM: A tree model based on simple structural units. Ann. Bot. 1996, 77, 87–98. [Google Scholar] [CrossRef]
- Mattheck, C.; Kubler, H. Wood—The internal Optimization of Trees; Springer: Heidelberg, Germany, 1997. [Google Scholar]
- Burkhart, H.E.; Tomé, M. Tree form and stem taper. In Modeling Forest Trees and Stands; Springer: Dordrecht, The Netherlands, 2012; pp. 9–41. [Google Scholar]
- Veit, N. Wie Verhalten sich Verwachsungen bei Unterschiedlichen Verbindungstechniken. Tech. Staatsschule Für Gart. Hohenh. 2012, unpublished. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2019. [Google Scholar]
- Kingsford, C.; Salzberg, S.L. What are decision trees? Nat. Biotechnol. 2008, 26, 1011–1013. [Google Scholar] [CrossRef]
- Langer, M.; Kelbel, M.C.; Speck, T.; Müller, C.; Speck, O. Twist-to-bend ratios and safety factors of petioles having various geometries, sizes and shapes. Front. Plant Sci. 2021, 12, 765605. [Google Scholar] [CrossRef]
- Wolff-Vorbeck, S.; Speck, O.; Langer, M.; Speck, T.; Dondl, P.W. Charting the twist-to-bend ratio of plant axes. J. R. Soc. Interface 2022, 19, 20220131. [Google Scholar] [CrossRef]
- Speck, T.; Spatz, H.-C.; Vogellehner, D. Contributions to the Biomechanics of Plants. I. Stabilities of Plant Stems with Strengthening Elements of Different Cross-Sections against Weight and Wind Forces. Bot. Acta 1990, 103, 111–122. [Google Scholar] [CrossRef]
- Middleton, W.; Erdal, H.I.; Detter, A.; D’Acunto, P.; Ludwig, F. Comparing structural models of linear elastic responses to bending in inosculated joints. Trees 2023, 1–13. [Google Scholar] [CrossRef]
- Isebrands, J.G.; Larson, P.R. Vascular anatomy of the nodal region in Populus deltoides Bartr. Am. J. Bot. 1977, 64, 1066–1077. [Google Scholar] [CrossRef]
- Larson, P.R.; Isebrands, J.G. Functional significance of the nodal constricted zone in Populus deltoides. Can. J. Bot. 1978, 56, 801–804. [Google Scholar] [CrossRef]
- Li, H.; Zhang, X.; Li, Z.; Wen, J.; Tan, X. A review of research on tree risk assessment methods. Forests 2022, 13, 1556. [Google Scholar] [CrossRef]
- Linhares, C.S.; Gonçalves, R.; Martins, L.M.; Knapic, S. Structural stability of urban trees using visual and instrumental techniques: A review. Forests 2021, 12, 1752. [Google Scholar] [CrossRef]













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variable | Platanus × hispanica | Salix alba |
|---|---|---|
| At [cm] | 5.7 (1.5) | 7.4 (1.1) |
| Ab [cm] | 7.5 (2.5) | 9.8 (0.6) |
| Bt [cm] | 6.0 (1.9) | 8.2 (1.8) |
| Bb [cm] | 6.6 (2.9) | 9.2 (1.8) |
| At/Ab [-] | 0.80 (0.07) | 0.78 (0.06) |
| Bt/Bb [-] | 0.85 (0.13) | 0.88 (0.06) |
| (At/Ab)/(Bt/Bb) [-] | 0.92 (0.06) | 0.87 (0.09) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mylo, M.D.; Ludwig, F.; Rahman, M.A.; Shu, Q.; Fleckenstein, C.; Speck, T.; Speck, O. Conjoining Trees for the Provision of Living Architecture in Future Cities: A Long-Term Inosculation Study. Plants 2023, 12, 1385. https://doi.org/10.3390/plants12061385
Mylo MD, Ludwig F, Rahman MA, Shu Q, Fleckenstein C, Speck T, Speck O. Conjoining Trees for the Provision of Living Architecture in Future Cities: A Long-Term Inosculation Study. Plants. 2023; 12(6):1385. https://doi.org/10.3390/plants12061385
Chicago/Turabian StyleMylo, Max D., Ferdinand Ludwig, Mohammad A. Rahman, Qiguan Shu, Christoph Fleckenstein, Thomas Speck, and Olga Speck. 2023. "Conjoining Trees for the Provision of Living Architecture in Future Cities: A Long-Term Inosculation Study" Plants 12, no. 6: 1385. https://doi.org/10.3390/plants12061385
APA StyleMylo, M. D., Ludwig, F., Rahman, M. A., Shu, Q., Fleckenstein, C., Speck, T., & Speck, O. (2023). Conjoining Trees for the Provision of Living Architecture in Future Cities: A Long-Term Inosculation Study. Plants, 12(6), 1385. https://doi.org/10.3390/plants12061385