
Effect of Ethanolic Extract of Vernonia amygdalina on the Proliferation, Viability and Function of Mouse Induced Pluripotent Stem Cells and Cardiomyocytes


Abstract
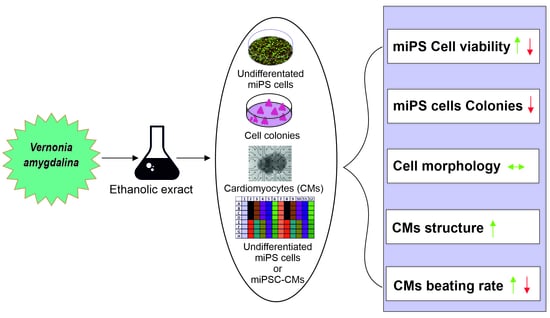
{kind=link}
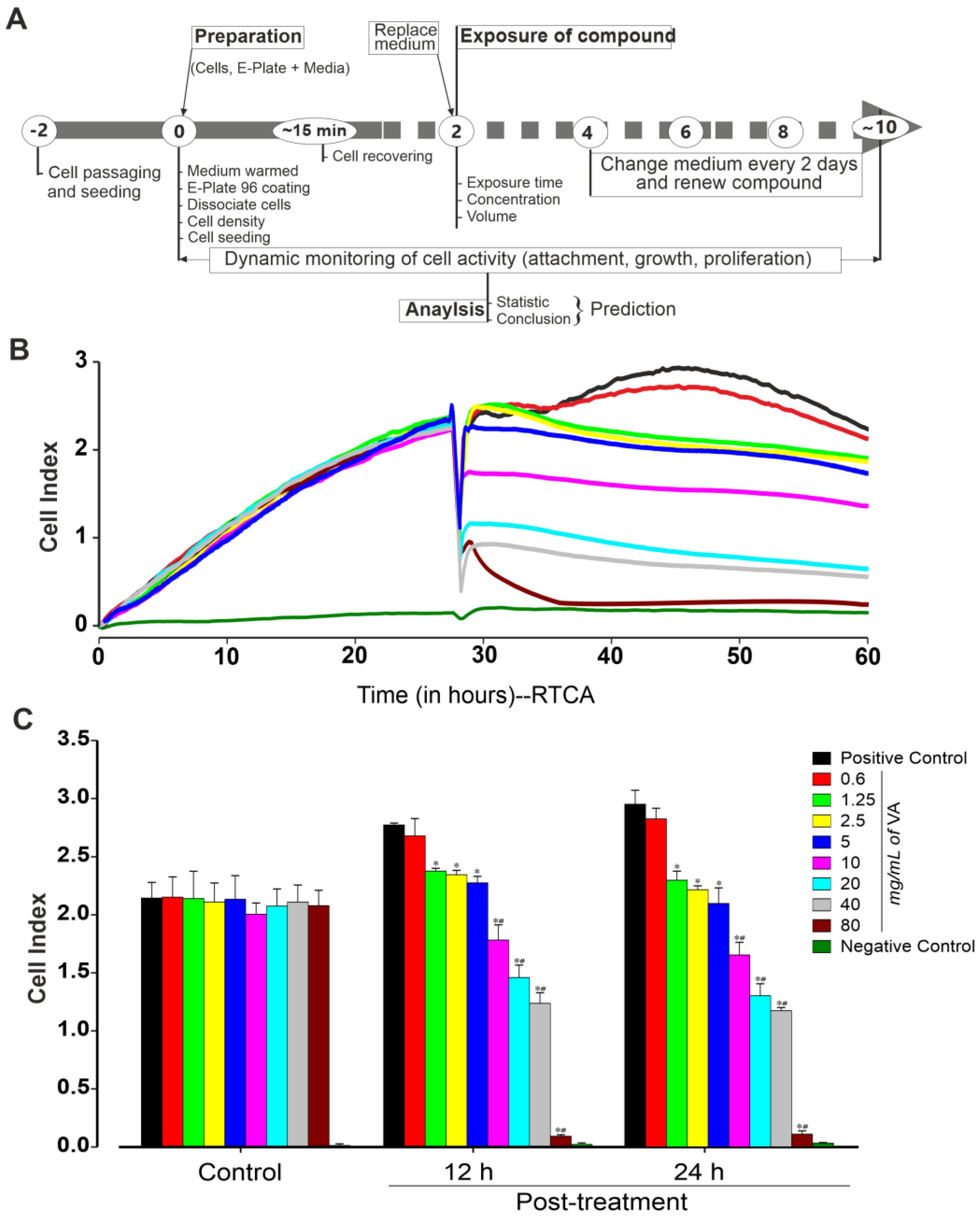{kind=link}
{kind=link}
{kind=link}
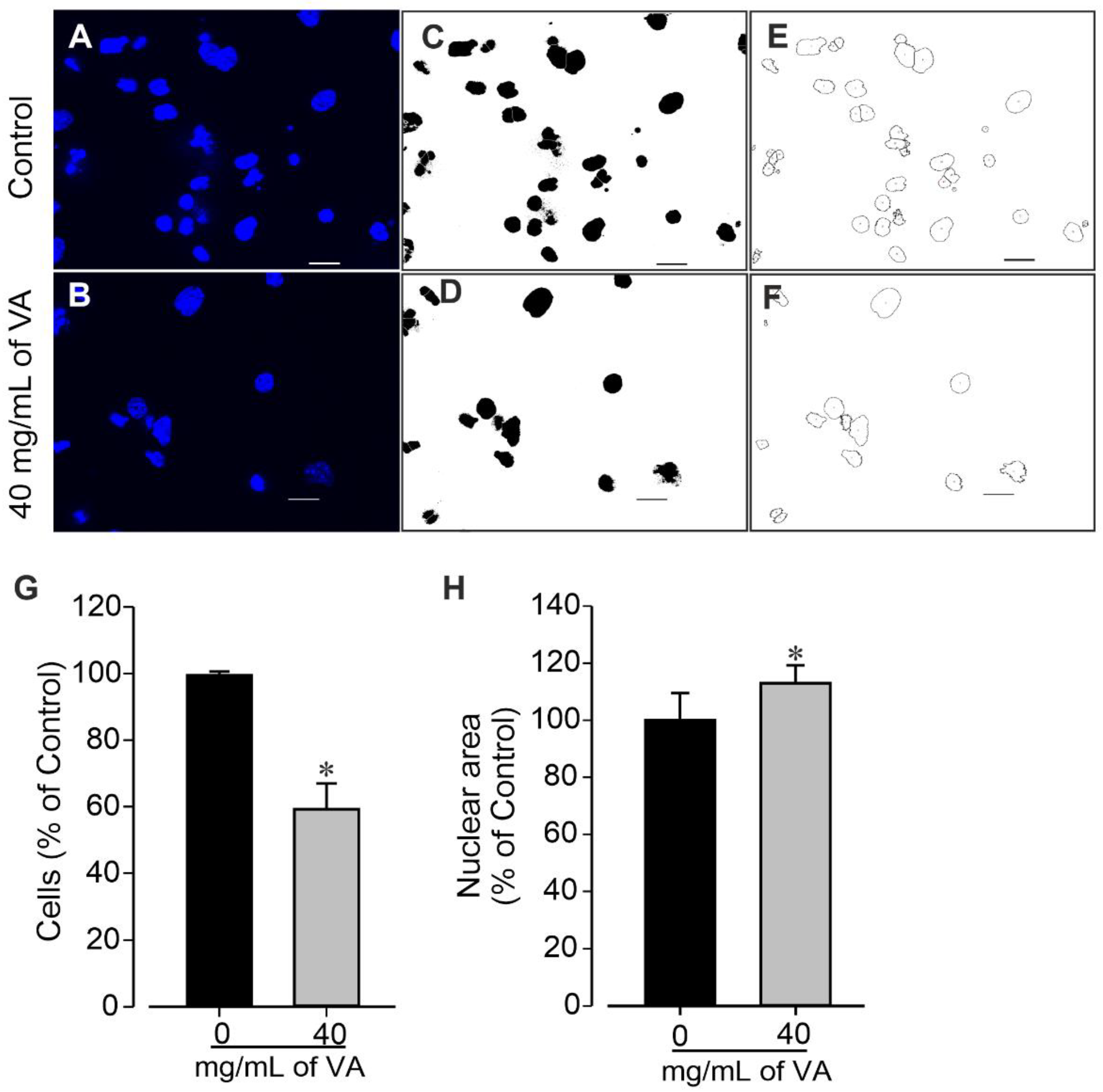{kind=link}
{kind=link}
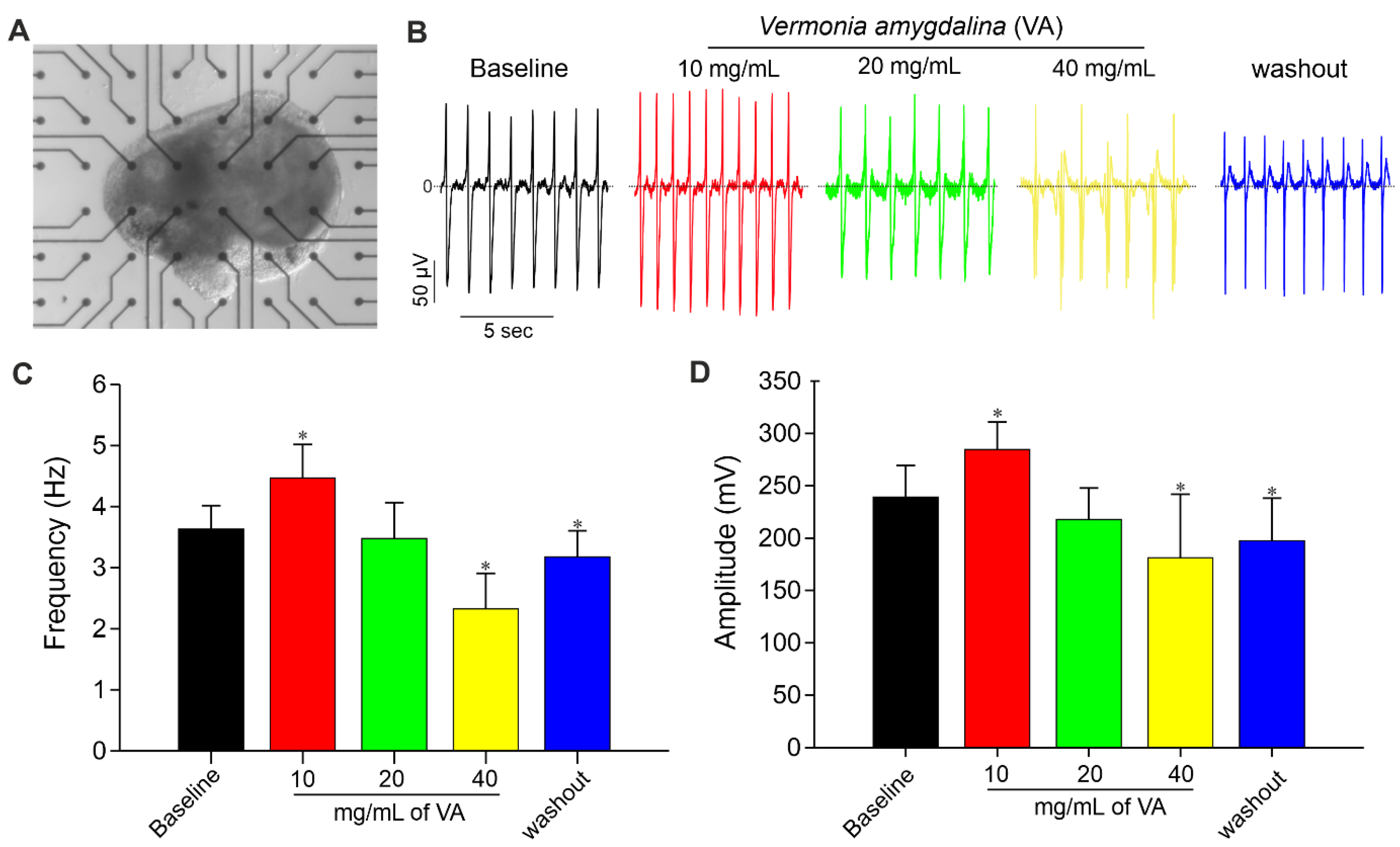{kind=link}
1. Introduction
2. Materials and Methods
2.1. Plant Extract Preparation
2.2. Mouse iPS Cells Culture and Treatment
2.3. xCELLigence Real-Time Cell Analyzers Platform
2.4. Microelectrode Array and Extracellular Field Potential Recordings
2.5. Cell Viability, Cytotoxicity and Apoptosis
2.6. Quantification of Cell Nuclei with ImageJ
2.7. Statistics
3. Results
3.1. Effect of Ethanolic Extract of V. amygdalina on Pluripotent Stem Cells
3.2. Effect of Ethanolic Extract of V. amygdalina on miPSCs’ Viability
3.3. Effect of Ethanolic Extract of V. amygdalina on miPSCs’ Colony Formation and Nuclei
3.4. Impact of Ethanolic Extract of V. amygdalina on Cardiac Differentiation of Pluripotent Stem Cells
4. Discussion
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Nembo, E.N.; Hescheler, J.; Nguemo, F. Stem cells in natural product and medicinal plant drug discovery-An overview of new screening approaches. Biomed. Pharmacother. 2020, 131, 110730. [Google Scholar] [CrossRef] [PubMed]
- Alara, O.R.; Abdurahman, N.H.; Ukaegbu, C.I.; Hassan, Z.; Kabbashi, N.A. Dataset on LC-Q-TOF/MS tentative identification of phytochemicals in the extract of Vernonia amygdalina leaf through positive ionization. Data Brief 2018, 21, 1686–1689. [Google Scholar] [CrossRef] [PubMed]
- Clement, E.; Erharuyi, O.; Vincent, I.; Joy, A.; Christopher, A.; Anthony, A.; Onyekaba, O.J.; Iftikhar, A.; Abiodun, F. Significance of bitter leaf (Vernonia amagdalina) in tropical diseases and beyond: A review. Malar Chemoth Cont. 2014, 3, 1–10. [Google Scholar]
- Ugbogu, O.C.; Emmanuel, O.; Agi, G.O.; Ibe, C.; Ekweogu, C.N.; Ude, V.C.; Uche, M.E.; Nnanna, R.O.; Ugbogu, E.A. A review on the traditional uses, phytochemistry, and pharmacological activities of clove basil (Ocimum gratissimum L.). Heliyon 2021, 7, e08404. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.T.; Liao, S.F.; Wu, Z.L.; Chang, C.W.; Wu, J.Y. Simultaneous study of antioxidant activity, DNA protection and anti-inflammatory effect of Vernonia amygdalina leaves extracts. PLoS ONE 2020, 15, e0235717. [Google Scholar] [CrossRef]
- Guo, L.; Abrams, R.M.; Babiarz, J.E.; Cohen, J.D.; Kameoka, S.; Sanders, M.J.; Chiao, E.; Kolaja, K.L. Estimating the risk of drug-induced proarrhythmia using human induced pluripotent stem cell-derived cardiomyocytes. Toxicol. Sci. 2011, 123, 281–289. [Google Scholar] [CrossRef]
- Abassi, Y.A.; Xi, B.; Li, N.; Ouyang, W.; Seiler, A.; Watzele, M.; Kettonhofen, R.; Bohlen, H.; Ehlich, A.; Kolossov, E.; et al. Dynamic Monitoring of Beating Periodicity of Stem Cell Derived Cardiomyocytes as a Predictive Tool for Preclinical Safety Assessment. Br. J. Pharmacol. 2012, 165, 1424–1441. [Google Scholar] [CrossRef] [PubMed]
- Nguemo, F.; Saric, T.; Pfannkuche, K.; Watzele, M.; Reppel, M.; Hescheler, J. In vitro model for assessing arrhythmogenic properties of drugs based on high-resolution impedance measurements. Cell. Physiol. Biochem. 2012, 29, 819–832. [Google Scholar] [CrossRef] [PubMed]
- Wabo Pone, J.; Fossi Tankoua, O.; Yondo, J.; Komtangi, M.C.; Mbida, M.; Bilong Bilong, C.F. The In Vitro Effects of Aqueous and Ethanolic Extracts of the Leaves of Ageratum conyzoides (Asteraceae) on Three Life Cycle Stages of the Parasitic Nematode Heligmosomoides bakeri (Nematoda: Heligmosomatidae). Vet. Med. Int. 2011, 2011, 140293. [Google Scholar] [CrossRef]
- Alabi, Q.K.; Adeyemi, W.J. Vernonia amygdalina (Del) as an antioxidant, aspirin toxicity, and oxidative stress. In Toxicology; Elsevier: Amsterdam, The Netherlands, 2021; pp. 491–504. [Google Scholar]
- Fatima, A.; Xu, G.X.; Nguemo, F.; Kuzmenkin, A.; Burkert, K.; Hescheler, J.; Šarić, T. Murine transgenic iPS cell line for monitoring and selection of cardiomyocytes. Stem Cell Res. 2016, 17, 266–272. [Google Scholar] [CrossRef]
- Nguemo, F.; Nembo, E.N.; Kamga Kapchoup, M.V.; Enzmann, F.; Hescheler, J. QuinoMit Q10-Fluid attenuates hydrogen peroxide-induced irregular beating in mouse pluripotent stem cell-derived cardiomyocytes. Biomed. Pharmacother. 2021, 142, 112089. [Google Scholar] [CrossRef]
- Nembo, E.N.; Atsamo, A.D.; Nguelefack, T.B.; Kamanyi, A.; Hescheler, J.; Nguemo, F. In vitro chronotropic effects of Erythrina senegalensis DC (Fabaceae) aqueous extract on mouse heart slice and pluripotent stem cell-derived cardiomyocytes. J. Ethnopharmacol. 2015, 165, 163–172. [Google Scholar] [CrossRef]
- Pahlavan, S.; Tousi, M.S.; Ayyari, M.; Alirezalu, A.; Ansari, H.; Saric, T.; Baharvand, H. Effects of hawthorn (Crataegus pentagyna) leaf extract on electrophysiologic properties of cardiomyocytes derived from human cardiac arrhythmia-specific induced pluripotent stem cells. FASEB J. 2018, 32, 1440–1451. [Google Scholar] [CrossRef]
- Strober, W. Trypan blue exclusion test of cell viability. Curr. Protoc. Immunol. 2001, 21, A-3B. [Google Scholar] [CrossRef]
- Antonio, A.S.; Wiedemann, L.S.M.; Galante, E.B.F.; Guimaraes, A.C.; Matharu, A.S.; Veiga-Junior, V.F. Efficacy and sustainability of natural products in COVID-19 treatment development: Opportunities and challenges in using agro-industrial waste from Citrus and apple. Heliyon 2021, 7, e07816. [Google Scholar] [CrossRef]
- Liu, C.; Feng, X.; Li, G.; Gokulnath, P.; Xiao, J. Generating 3D human cardiac constructs from pluripotent stem cells. EBioMedicine 2022, 76, 103813. [Google Scholar] [CrossRef] [PubMed]
- Ndjenda Ii, M.K.; Nguelefack-Mbuyo, E.P.; Hescheler, J.; Nguelefack, T.B.; Nguemo, F. Assessment of the In Vitro Cytotoxicity Effects of the Leaf Methanol Extract of Crinum zeylanicum on Mouse Induced Pluripotent Stem Cells and Their Cardiomyocytes Derivatives. Pharmaceuticals 2021, 14, 1208. [Google Scholar] [CrossRef] [PubMed]
- Luo, X.; Jiang, Y.; Fronczek, F.R.; Lin, C.; Izevbigie, E.B.; Lee, K.S. Isolation and structure determination of a sesquiterpene lactone (vernodalinol) from Vernonia amygdalina extracts. Pharm. Biol. 2011, 49, 464–470. [Google Scholar] [CrossRef]
- Yusoff, S.F.; Haron, F.F.; Tengku Muda Mohamed, M.; Asib, N.; Sakimin, S.Z.; Abu Kassim, F.; Ismail, S.I. Antifungal Activity and Phytochemical Screening of Vernonia amygdalina Extract against Botrytis cinerea Causing Gray Mold Disease on Tomato Fruits. Biology 2020, 9, 286. [Google Scholar] [CrossRef]
- Howard, C.B.; McDowell, R.; Feleke, K.; Deer, E.; Stamps, S.; Thames, E.; Singh, V.; Pervin, S. Chemotherapeutic Vulnerability of Triple-negative Breast Cancer Cell-derived Tumors to Pretreatment with Vernonia amygdalina Aqueous Extracts. Anticancer Res. 2016, 36, 3933–3943. [Google Scholar] [PubMed]
- Syahputra, R.A.; Harahap, U.; Dalimunthe, A.; Pandapotan, M.; Satria, D. Protective effect of Vernonia amygdalina Delile against doxorubicin-induced cardiotoxicity. Heliyon 2021, 7, e07434. [Google Scholar] [CrossRef] [PubMed]
- Olshansky, B.; Sabbah, H.N.; Hauptman, P.J.; Colucci, W.S. Parasympathetic nervous system and heart failure: Pathophysiology and potential implications for therapy. Circulation 2008, 118, 863–871. [Google Scholar] [CrossRef] [PubMed]
- Ch’ng, Y.S.; Loh, Y.C.; Tan, C.S.; Ahmad, M.; Asmawi, M.Z.; Wan Omar, W.M.; Yam, M.F. Vasorelaxant properties of Vernonia amygdalina ethanol extract and its possible mechanism. Pharm. Biol. 2017, 55, 2083–2094. [Google Scholar] [CrossRef]
- Zhang, Y.; Dong, C. Regulatory mechanisms of mitogen-activated kinase signaling. Cell. Mol. Life Sci. 2007, 64, 2771–2789. [Google Scholar] [CrossRef] [PubMed]
- Streicher, J.M.; Ren, S.; Herschman, H.; Wang, Y. MAPK-activated protein kinase-2 in cardiac hypertrophy and cyclooxygenase-2 regulation in heart. Circ. Res. 2010, 106, 1434–1443. [Google Scholar] [CrossRef] [PubMed]
- Rose, B.A.; Force, T.; Wang, Y. Mitogen-activated protein kinase signaling in the heart: Angels versus demons in a heart-breaking tale. Physiol. Rev. 2010, 90, 1507–1546. [Google Scholar] [CrossRef] [PubMed]
- Guo, Y.J.; Pan, W.W.; Liu, S.B.; Shen, Z.F.; Xu, Y.; Hu, L.L. ERK/MAPK signalling pathway and tumorigenesis. Exp. Ther. Med. 2020, 19, 1997–2007. [Google Scholar] [CrossRef]
- Zheng, M.; Kang, L.; Uchino, T.; Liu, G.; Wang, Y.; Ono, K. Mitogen-activated protein kinase p38 modulates pacemaker ion channels differentiation in P19-derived pluripotent cells. J. Physiol. Sci. 2020, 70, 39. [Google Scholar] [CrossRef]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nguemfo Tchankugni, A.; Mbida, M.; Hescheler, J.; Nguemo, F. Effect of Ethanolic Extract of Vernonia amygdalina on the Proliferation, Viability and Function of Mouse Induced Pluripotent Stem Cells and Cardiomyocytes. Plants 2023, 12, 1108. https://doi.org/10.3390/plants12051108
Nguemfo Tchankugni A, Mbida M, Hescheler J, Nguemo F. Effect of Ethanolic Extract of Vernonia amygdalina on the Proliferation, Viability and Function of Mouse Induced Pluripotent Stem Cells and Cardiomyocytes. Plants. 2023; 12(5):1108. https://doi.org/10.3390/plants12051108
Chicago/Turabian StyleNguemfo Tchankugni, Arlette, Mpoame Mbida, Jürgen Hescheler, and Filomain Nguemo. 2023. "Effect of Ethanolic Extract of Vernonia amygdalina on the Proliferation, Viability and Function of Mouse Induced Pluripotent Stem Cells and Cardiomyocytes" Plants 12, no. 5: 1108. https://doi.org/10.3390/plants12051108
APA StyleNguemfo Tchankugni, A., Mbida, M., Hescheler, J., & Nguemo, F. (2023). Effect of Ethanolic Extract of Vernonia amygdalina on the Proliferation, Viability and Function of Mouse Induced Pluripotent Stem Cells and Cardiomyocytes. Plants, 12(5), 1108. https://doi.org/10.3390/plants12051108