
Effects of Spirulina maxima on a Model of Sexual Dysfunction in Streptozotocin-Induced Diabetic Male Rats

 ,
,  ,
,  , , , and
, , , and
Abstract
1. Introduction
2. Results
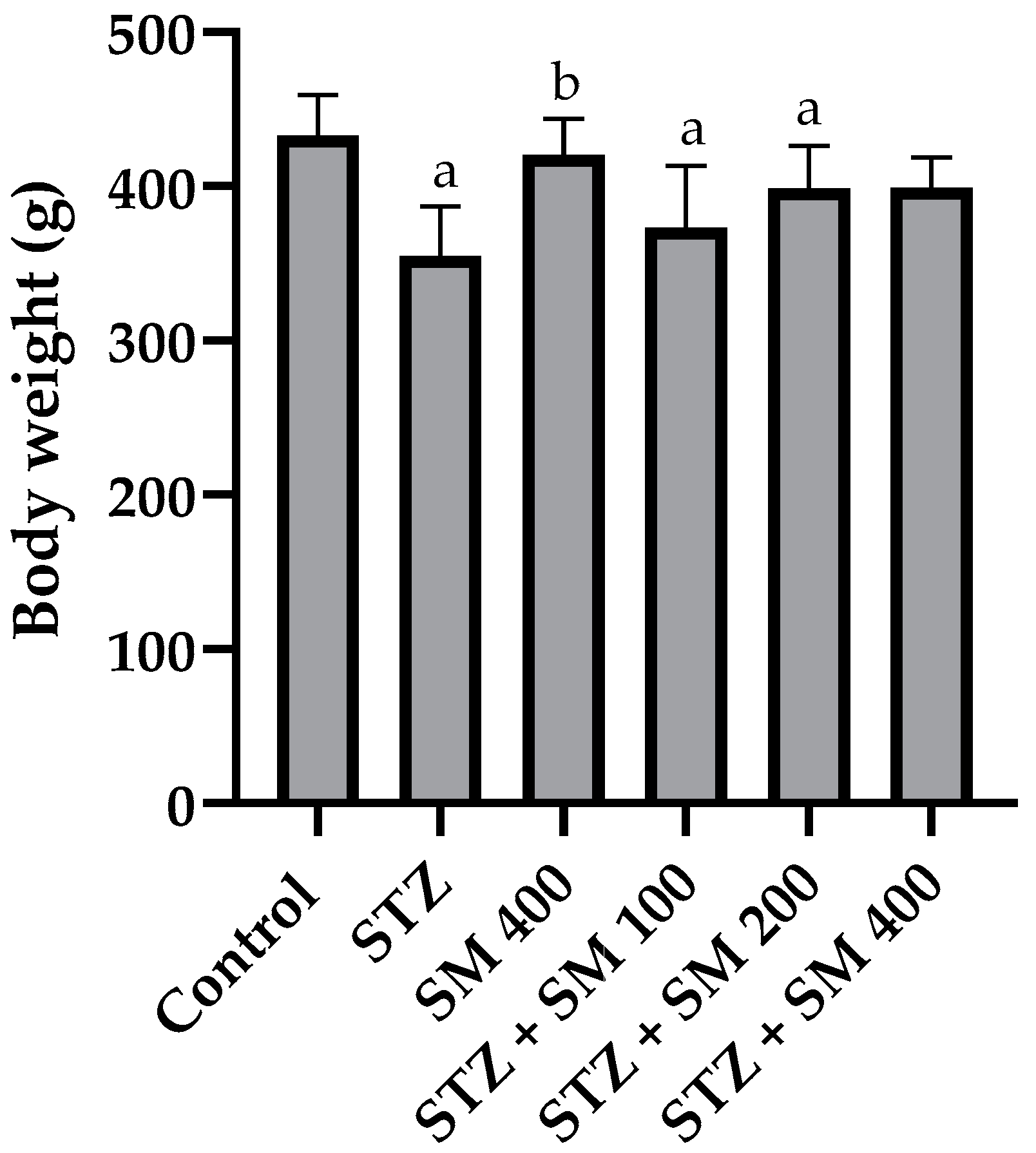
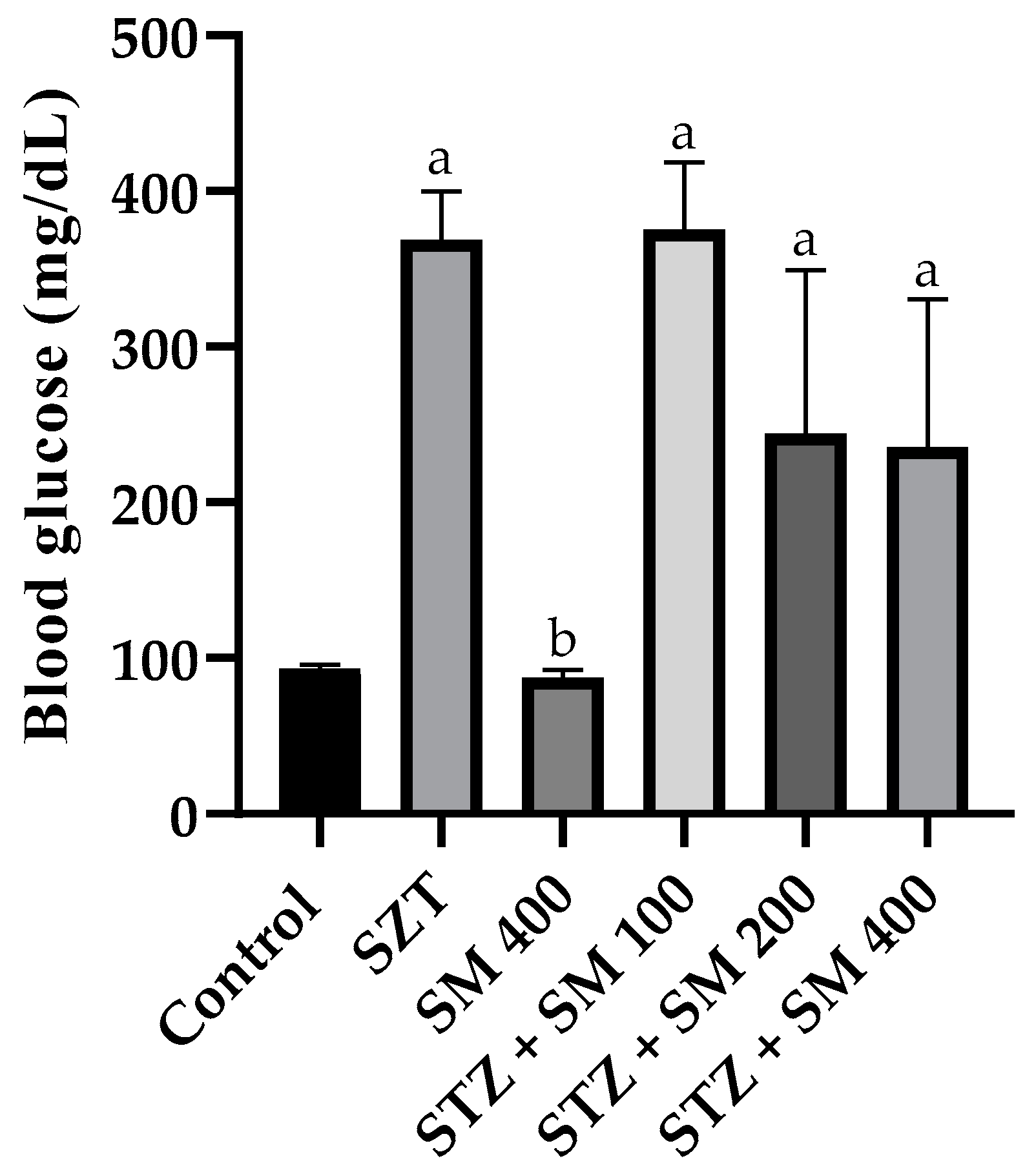
2.1. Body Weight and Glycemia
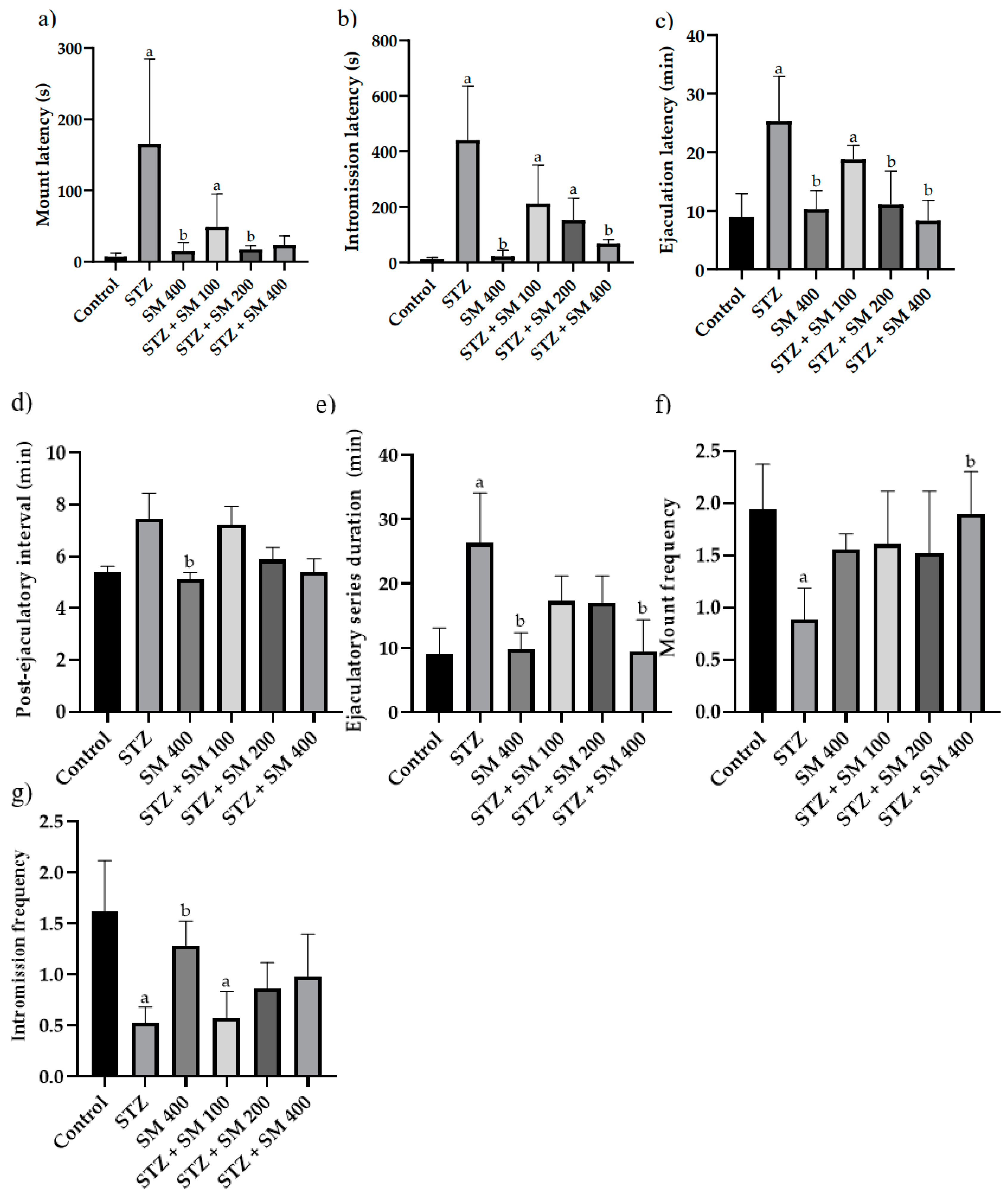
2.2. Copulatory Behavior
2.3. Sperm Quality
2.4. Sex Organ Weight
2.5. Biochemical Analyses
2.6. Testosterone Analysis
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Preparation of Females for Couplings, Animal Selection, and Preparation for Experiments
4.3. Induction of Diabetes Type II in Males
4.4. Mating Behavior Test
4.5. Sperm Collection and Analysis
4.6. Biochemical Analysis
4.7. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Kong, Z.L.; Johnson, A.; Ko, F.C.; He, J.L.; Cheng, S.C. Effect of Cistanche Tubulosa Extracts on Male Reproductive Function in Streptozotocin–Nicotinamide-Induced Diabetic Rats. Nutrients 2018, 10, 1562. [Google Scholar] [CrossRef] [PubMed]
- Ghaheri, M.; Miraghaee, S.; Babaei, A.; Mohammadi, B.; Kahrizi, D.; Saivosh Haghighi, Z.M.; Bahrami, G. Effect of Stevia rebaudiana Bertoni extract on sexual dysfunction in Streptozotocin-induced diabetic male rats. Cell. Mol. Biol. 2018, 64, 6–10. [Google Scholar] [CrossRef] [PubMed]
- Kalka, D. Depression symptoms, sexual satisfaction and satisfaction with a relationship in individuals with type 2 diabetes and sexual dysfunctions. Objawy depresji i satysfakcja seksualna a zadowolenie ze związku u osób z cukrzycą typu 2 z dysfunkcjami seksualnymi. Psychiatr. Pol. 2018, 52, 1087–1099. [Google Scholar] [CrossRef] [PubMed]
- Rai, A.; Das, S.; Chamallamudi, M.R.; Nandakumar, K.; Shetty, R.; Gill, M.; Sumalatha, S.; Devkar, R.; Gourishetti, K.; Kumar, N. Evaluation of the aphrodisiac potential of a chemically characterized aqueous extract of Tamarindus indica pulp. J. Ethnopharmacol. 2018, 210, 118–124. [Google Scholar] [CrossRef] [PubMed]
- Maiorino, M.I.; Bellastella, G.; Esposito, K. Diabetes and sexual dysfunction: Current perspectives. Diabetes. Metab. Syndr. Obes. 2014, 7, 95–105. [Google Scholar] [CrossRef]
- Wan, X.Z.; Li, T.T.; Zhong, R.T.; Chen, H.B.; Xia, X.; Gao, L.Y.; Gao, X.X.; Liu, B.; Zhang, H.Y.; Zhao, C. Anti-diabetic activity of PUFAs-rich extracts of Chlorella pyrenoidosa and Spirulina platensis in rats. Food Chem. Toxicol. 2019, 128, 233–239. [Google Scholar] [CrossRef]
- Lert-Amornpat, T.; Maketon, C.; Fungfuang, W. Effect of Kaempferia parviflora on sexual performance in streptozotocin-induced diabetic male rats. Andrologia 2017, 49, e12770. [Google Scholar] [CrossRef]
- Figueroa, M.C.; Pérez, I.; Mejía, R. Characterization of a type 2 diabetes model in female Wistar rats. Rev. MVZ Córdoba 2013, 18, 3699–3707. [Google Scholar] [CrossRef]
- World Health Organization. Informe Mundial Sobre la Diabetes; WHO: Geneva, Switzerland, 2016; pp. 1–84. Available online: https://apps.who.int/iris/bitstream/handle/10665/254649/9789243565255-spa.pdf (accessed on 6 November 2022).
- Yu, W.; Wan, Z.; Qiu, X.F.; Chen, Y.; Dai, Y.T. Resveratrol, an activator of SIRT1, restores erectile function in streptozotocin-induced diabetic rats. Asian J. Androl. 2013, 15, 646–651. [Google Scholar] [CrossRef]
- Elbatreek, M.H.; Pachado, M.P.; Cuadrado, A.; Jandeleit-Dahm, K.; Schmidt, H. Reactive Oxygen Comes of Age: Mechanism-Based Therapy of Diabetic End-Organ Damage. Trends Endocrinol. Metab. 2019, 30, 312–327. [Google Scholar] [CrossRef]
- Sahin, K.; Orhan, C.; Akdemir, F.; Tuzcu, M.; Gencoglu, H.; Sahin, N.; Turk, G.; Yilmaz, I.; Ozercan, I.H.; Juturu, V. Comparative evaluation of the sexual functions and NF-κB and Nrf2 pathways of some aphrodisiac herbal extracts in male rats. BMC Complement. Altern. Med. 2016, 16, 318. [Google Scholar] [CrossRef] [PubMed]
- Duangnin, N.; Phitak, T.; Pothacharoen, P.; Kongtawelert, P. In vitro and in vivo investigation of natural compounds from seed extract of Mucuna pruriens lacking l-DOPA for the treatment of erectile dysfunction. Asian Pac. J. Trop. Med. 2017, 10, 238–252. [Google Scholar] [CrossRef] [PubMed]
- Cristóbal-Luna, J.M.; Chamorro-Cevallos, G.; Monterubio-López, R.; Pérez-Ramos, J.; Pérez-González, C.; Pérez-Gutiérrez, S. Chapter 2: The effect of Arthrospira (Spirulina) maxima and its aqueous extract on fetal alcohol syndrome, induced in CD1 mice. In Spirulina and Its Health Benefits; Cochran, J., Ed.; NOVA: Hauppauge, NY, USA, 2021; pp. 45–92. [Google Scholar] [CrossRef]
- Miranda, M.S.; Cintra, R.G.; Bsrros, S.B.M. Antioxidant activity of the microalga Spirulina maxima. Braz. J. Med. Biol. Res. 1998, 31, 1075–1079. [Google Scholar] [CrossRef] [PubMed]
- Brito, A.F.; Silva, A.S.; de Souza, A.A.; Ferreira, P.B.; de Souza, I.L.L.; Araujo, L.C.D.C.; da Silva, B.A. Supplementation with Spirulina platensis Improves Tracheal Reactivity in Wistar Rats by Modulating Inflammation and Oxidative Stress. Front. Pharmacol. 2022, 13, 826649. [Google Scholar] [CrossRef]
- Ferreira, P.B.; Diniz, A.F.A.; Lacerda Júnior, F.F.; Silva, M.D.C.C.; Cardoso, G.A.; Silva, A.S.; da Silva, B.A. Supplementation with Spirulina platensis Prevents Uterine Diseases Related to Muscle Reactivity and Oxidative Stress in Rats Undergoing Strength Training. Nutrients 2021, 13, 3763. [Google Scholar] [CrossRef]
- Sadek, K.M.; Lebda, M.A.; Nasr, S.M.; Shoukry, M. Spirulina platensis prevents hyperglycemia in rats by modulating gluconeogenesis and apoptosis via modification of oxidative stress and MAPK-pathways. Biomed. Pharmacother. 2017, 92, 1085–1094. [Google Scholar] [CrossRef]
- Deng, R.; Chow, T.J. Hypolipidemic, antioxidant, and antiinflammatory activities of microalgae Spirulina. Cardiovasc. Ther. 2010, 28, e33–e45. [Google Scholar] [CrossRef]
- Shastri, D.; Kumar, M.; Kumar, A. Modulation of lead toxicity by Spirulina fusiformis. Phytother. Res. 1999, 13, 258–260. [Google Scholar] [CrossRef]
- Al-Dhabi, N.A. Heavy metal analysis in commercial Spirulina products for human consumption. Saudi J. Biol. Sci. 2013, 20, 383–388. [Google Scholar] [CrossRef]
- Gunes, S.; Tamburaci, S.; Dalay, M.C.; Deliloglu-Gurhan, I. In vitro evaluation of Spirulina platensis extract incorporated skin cream with its wound healing and antioxidant activities. Pharm. Biol. 2017, 55, 1824–1832. [Google Scholar] [CrossRef]
- Abdel-Daim, M.M.; Shaaban-Ali, M.; Madkour, F.F.; Elgendy, H. Oral Spirulina Platensis Attenuates Hyperglycemia and Exhibits Antinociceptive Effect in Streptozotocin-Induced Diabetic Neuropathy Rat Model. J. Pain Res. 2020, 13, 2289–2296. [Google Scholar] [CrossRef]
- Sinha, S.; Patro, N.; Patro, I.K. Amelioration of neurobehavioral and cognitive abilities of F1 progeny following dietary supplementation with Spirulina to protein malnourished mothers. Brain Behav. Immun. 2020, 85, 69–87. [Google Scholar] [CrossRef]
- Sinha, S.; Patro, N.; Patro, I.K. Maternal Protein Malnutrition: Current and Future Perspectives of Spirulina Supplementation in Neuroprotection. Front. Neurosci. 2018, 12, 966. [Google Scholar] [CrossRef]
- Shahzad, S.; Batool, Z.; Tabassum, S.; Ahmad, S.; Kamil, N.; Khaliq, S.; Nawaz, A.; Haider, S. Spirulina platensis (Blue-green algae): A miracle from sea combats the oxidative stress and improves behavioral deficits in an animal model of Schizophrenia. Pak. J. Pharm. Sci. 2020, 33 (Suppl. 4), 1847–1853. [Google Scholar]
- De, A.; Singh, M.F.; Singh, V.; Ram, V.; Bisht, S. Treatment effect of l-Norvaline on the sexual performance of male rats with streptozotocin induced diabetes. Eur. J. Pharmacol. 2016, 771, 247–254. [Google Scholar] [CrossRef] [PubMed]
- Farzadi, L.; Khaki, A.; Ghasemzadeh, A.; Ouladsahebmadarek, E.; Ghadamkheir, E.; Shadfar, S.; Khaki, A.A. Effect of rosmarnic acid on sexual behavior in diabetic male rats. Afr. J. Pharm. Pharmacol. 2011, 5, 1906–1910. [Google Scholar] [CrossRef]
- Al-Oanzi, Z.H. Erectile dysfunction attenuation by naringenin in streptozotocin-induced diabetic rats. J. Food Biochem. 2019, 43, e12885. [Google Scholar] [CrossRef] [PubMed]
- Minaz, N.; Razdan, R.; Hammock, B.D.; Mujwar, S.; Goswami, S.K. Impact of diabetes on male sexual function in streptozotocin-induced diabetic rats: Protective role of soluble epoxide hydrolase inhibitor. Biomed. Pharmacother. 2019, 115, 108897. [Google Scholar] [CrossRef] [PubMed]
- Chaturapanich, G.; Chaiyakul, S.; Verawatnapakul, V.; Pholpramool, C. Effects of Kaempferia parviflora extracts on reproductive parameters and spermatic blood flow in male rats. Reproduction 2008, 136, 515–522. [Google Scholar] [CrossRef] [PubMed]
- Sing, R.; Ali, A.; Jeyabalan, G.; Semwal, A.; Jaikishan. An overview of the current methodologies used for evaluation of aphrodisiac agents. J. Acute Dis. 2013, 2013, 85–91. [Google Scholar] [CrossRef]
- Erhabor, J.O.; Idu, M. Aphrodisiac potentials of the ethanol extract of Aloe barbadensis Mill. root in male Wistar rats. BMC Complement. Altern. Med. 2017, 17, 360. [Google Scholar] [CrossRef] [PubMed]
- Arya, A.; Cheah, S.C.; Looi, C.Y.; Taha, H.; Mustafa, M.R.; Mohd, M.A. The methanolic fraction of Centratherum antihelminticam seed down regulates pro-inflammatory cytokines, oxidative stress, and hyperglycemia in STZ-nicotinamide-induced type 2 diabetic rats. Food Chem. Toxicol. 2012, 50, 4209–4220. [Google Scholar] [CrossRef]
- Al-Roujeaie, A.S.; Abuohashih, H.M.; Ahmed, M.M.; Alkhamees, O.A. Effect of rutin on diabetic-induced erectile dysfunction: Possible involvement of testicular biomarkers in male rats. Andrologia 2017, 49, e12737. [Google Scholar] [CrossRef] [PubMed]
- Pontes, D.A.; Fernandes, G.S.A.; Piffer, R.C.; Gerardin, D.C.C.; Pereira, O.C.M.; Kempinas, W.G. Ejaculatory dysfunction in streptozotocin-induced diabetic rats: The role of testosterone. Pharmacol. Rep. 2011, 63, 130–138. [Google Scholar] [CrossRef] [PubMed]
- Abdrabou, M.I.; Elleithy, E.; Yasin, N.; Shaheen, Y.M.; Galal, M. Ameliorative effects of Spirulina maxima and Allium sativum on lead acetate-induced testicular injury in male albino rats with respect to caspase-3 gene expression. Acta Histochem. 2019, 121, 198–206. [Google Scholar] [CrossRef]
- Barkallah, M.; Slima, A.B.; Elleuch, F.; Fendri, I.; Pichon, C.; Abdelkafi, S.; Baril, P. Protective Role of Spirulina platensis against Bifenthrin-Induced Reprotoxicity in Adult Male Mice by Reversing Expression of Altered Histological, Biochemical, and Molecular Markers Including MicroRNAs. Biomolecules 2020, 10, 753. [Google Scholar] [CrossRef] [PubMed]
- Salama, A.F.; Kasem, S.M.; Tousson, E.; Elsisy, M.K. Protective role of L-carnitine and vitamin E on the testis of atherosclerotic rats. Toxicol. Ind. Health 2015, 31, 467–474. [Google Scholar] [CrossRef]
- Allouh, M.Z.; Daradka, H.M.; Abu-Ghaida, J.H. Influence of Cyperus esculentus tubers (tiger nut) on male rat copulatory behavior. BMC Complement. Altern. Med. 2015, 15, 331. [Google Scholar] [CrossRef]
- Cabrera, A.; Paredes, R.G. Effects of chronic estradiol or testosterone treatment upon sexual behavior in sexually sluggish male rats. Pharmacol. Biochem. Behav. 2012, 101, 336–341. [Google Scholar] [CrossRef]
- Noberasco, G.; Odetti, P.; Boeri, D.; Maiello, M.; Adezati, L. Malondialdehyde (MDA) level in diabetic subjects. Relationship with blood glucose and glycosylated hemoglobin. Biomed Pharmacother. 1991, 45, 193–196. [Google Scholar] [CrossRef]
- Younus, H. Therapeutic potentials of superoxide dismutase. Int. J. Health Sci. 2018, 12, 88–93. [Google Scholar]
- Hernández-Lepe, M.A.; Wall-Medrano, A.; Juárez-Oropeza, M.A.; Ramos-Jiménez, A.; Hernández-Torres, R.P. Spirulina y su efecto hipolipemiante y antioxidante en humanos: Una revisión sistemática [Spirulina and its hypolipidemic and antioxidant effects in humans: A systematic review]. Nutr. Hosp. 2015, 32, 494–500. [Google Scholar] [CrossRef] [PubMed]
- Lucio, R.A.; Tlachi, J.L.; López, A.A.; Zempoalteca, R.; Velázquez-Moctezuma, J. Análisis de los parámetros del eyaculado en la rata Wistar de laboratorio: Descripción de la técnica. Vet. Mex. 2008, 40, 405–415. [Google Scholar]
- Khajuria, D.K.; Razdan, R.; Mahapatra, D.R. Description of a new method of ovariectomy in female rats. Rev. Bras. Reumatol. 2012, 52, 462–470. [Google Scholar]
- Masiello, P.; Broca, C.; Gross, R.; Roye, M.; Manteghetti, M.; Hillaire-Buys, D.; Novelli, M.; Ribes, G. Experimental NIDDM: Development of a new model in adult rats administered streptozotocin and nicotinamide. Diabetes 1998, 47, 224–229. [Google Scholar] [CrossRef]
- Garrido-Acosta, O.; Limón, I.D.; García, E.; Anguiano-Robledo, L.; Barrientos-Alvarado, C.; Chamorro-Cevallos, G. Efecto Protector de Spirulina maxima en un modelo experimental de enfermedad de Parkinson inducido con MPP+ Protective effect of Spirulina maxima in an experimental Parkinson´s disease model induced by MPP+. Rev. Mex. Cienc. Farm. 2017, 48, 56–64. [Google Scholar]
- Fumero, B.; Fernandez-Vera, J.R.; Gonzalez-Mora, J.L.; Mas, M. Changes I, 5(6), n monoamine turnover in forebrain areas associated with masculine sexual behavior: A microdialysis study. Brain Res. 1994, 662, 233–239. [Google Scholar] [CrossRef]
- Andersen, H.R.; Nielsen, J.B.; Nielsen, F.; Grandjean, P. Antioxidative enzyme activities in human erythrocytes. Clin. Chem. 1997, 43, 562–568. [Google Scholar] [CrossRef]
- Paglia, D.E.; Valentine, W.N. Studies on the quantitative and qualitative characterization of erythrocyte glutathione peroxidase. J. Lab. Clin. Med. 1967, 70, 158–169. [Google Scholar]
- Matsuzawa, T.; Saitoh, H.; Sano, M.; Tomita, I.; Ohkawa, M.; Ikekawa, T. Studies on Antioxidant Effects of Hypsizigus marmoreus. II. Effects of Hypsizigus marmoreus for Antioxidant Activities of Tumor-Bearing Mice. Yakugaku Zasshi 1998, 118, 476–481. [Google Scholar] [CrossRef]
- Mitteer, D.R.; Greer, B.D. Using GraphPad Prism’s Heat Maps for Efficient, Fine-Grained Analyses of Single-Case Data. Behav. Anal. Pract. 2022, 15, 505–514. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment (mg/kg) | Sperm Count (X106/mL) | Sperm Motility | Sperm Viability | Sperm Abnormality (%) |
|---|---|---|---|---|
| Control | 69.42 ± 2.72 | 29.77 ± 7.36 | 61.74 ± 1.20 | 3.93 ± 0.98 |
| STZ | 39.44 ± 6.05 a | 9.48 ± 2.58 a | 27.56 ± 2.94 a | 6.08 ± 1.17 a |
| SM 400 | 75.29 ± 6.33 b | 20.52 ± 2.99 b | 61.49+ 2.20 b | 4.3 ± 1.13 b |
| STZ + SM 100 | 64.04 ± 6.22 b | 7.55 ± 1.89 a | 33.45 ± 4.00 | 4.1 ± 1.24 b |
| STZ + SM 200 | 72.50 ± 2.81 b | 16.38 ± 2.87 b | 44.95 ± 2.68 b | 3.9 ± 1.20 b |
| STZ + SM 400 | 74.20 ± 2.95 b | 8.52 ± 1.52 a | 43.82 ± 3.86 b | 3.2 ± 1.12 b |
| Treatments (mg/kg) | TBARS (nmol/mg Protein) | SOD (U/mg Protein) | GPX (mU/mL) |
|---|---|---|---|
| Control | 12.57 ± 0.42 | 4.11 ± 0.03 | 436.20 ± 24.63 |
| STZ | 17.65 ± 1.25 a | 1.59 ± 0.04 a | 209.83 ± 1.83 a |
| SM 400 | 13.91 ± 0.69 b | 4.24 ± 0.02 b | 430.10 ± 25.17 b |
| STZ + SM 100 | 13.79 ± 0.45 b | 2.81 ± 0.06 b | 332.65 ± 10.75 |
| STZ + SM 200 | 11.86 ± 0.64 b | 4.28 ± 0.03 b | 451.55 ± 11.55 b |
| STZ + SM 400 | 12.38 ± 0.79 b | 4.27 ± 0.02 b | 447.42 ± 11.72 b |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Olvera-Roldán, E.O.; Cristóbal-Luna, J.M.; García-Martínez, Y.; Mojica-Villegas, M.A.; Pérez-Pastén-Borja, R.; Gutiérrez-Salmeán, G.; Pérez-Gutiérrez, S.; García-Rodríguez, R.V.; Madrigal-Santillán, E.; Morales-González, J.A.; et al. Effects of Spirulina maxima on a Model of Sexual Dysfunction in Streptozotocin-Induced Diabetic Male Rats. Plants 2023, 12, 722. https://doi.org/10.3390/plants12040722
Olvera-Roldán EO, Cristóbal-Luna JM, García-Martínez Y, Mojica-Villegas MA, Pérez-Pastén-Borja R, Gutiérrez-Salmeán G, Pérez-Gutiérrez S, García-Rodríguez RV, Madrigal-Santillán E, Morales-González JA, et al. Effects of Spirulina maxima on a Model of Sexual Dysfunction in Streptozotocin-Induced Diabetic Male Rats. Plants. 2023; 12(4):722. https://doi.org/10.3390/plants12040722
Chicago/Turabian StyleOlvera-Roldán, Eduardo Osel, José Melesio Cristóbal-Luna, Yuliana García-Martínez, María Angélica Mojica-Villegas, Ricardo Pérez-Pastén-Borja, Gabriela Gutiérrez-Salmeán, Salud Pérez-Gutiérrez, Rosa Virginia García-Rodríguez, Eduardo Madrigal-Santillán, José A. Morales-González, and et al. 2023. "Effects of Spirulina maxima on a Model of Sexual Dysfunction in Streptozotocin-Induced Diabetic Male Rats" Plants 12, no. 4: 722. https://doi.org/10.3390/plants12040722
APA StyleOlvera-Roldán, E. O., Cristóbal-Luna, J. M., García-Martínez, Y., Mojica-Villegas, M. A., Pérez-Pastén-Borja, R., Gutiérrez-Salmeán, G., Pérez-Gutiérrez, S., García-Rodríguez, R. V., Madrigal-Santillán, E., Morales-González, J. A., & Chamorro-Cevallos, G. (2023). Effects of Spirulina maxima on a Model of Sexual Dysfunction in Streptozotocin-Induced Diabetic Male Rats. Plants, 12(4), 722. https://doi.org/10.3390/plants12040722