

Bio-Intensive Tactics for the Management of Invasive Fall Armyworm for Organic Maize Production

 and
and
Abstract
1. Introduction
2. Results
2.1. Field Parasitism by Egg and Larval Parasitoids of FAW
2.2. Effect of Different Botanicals on Developmental Stages of FAW
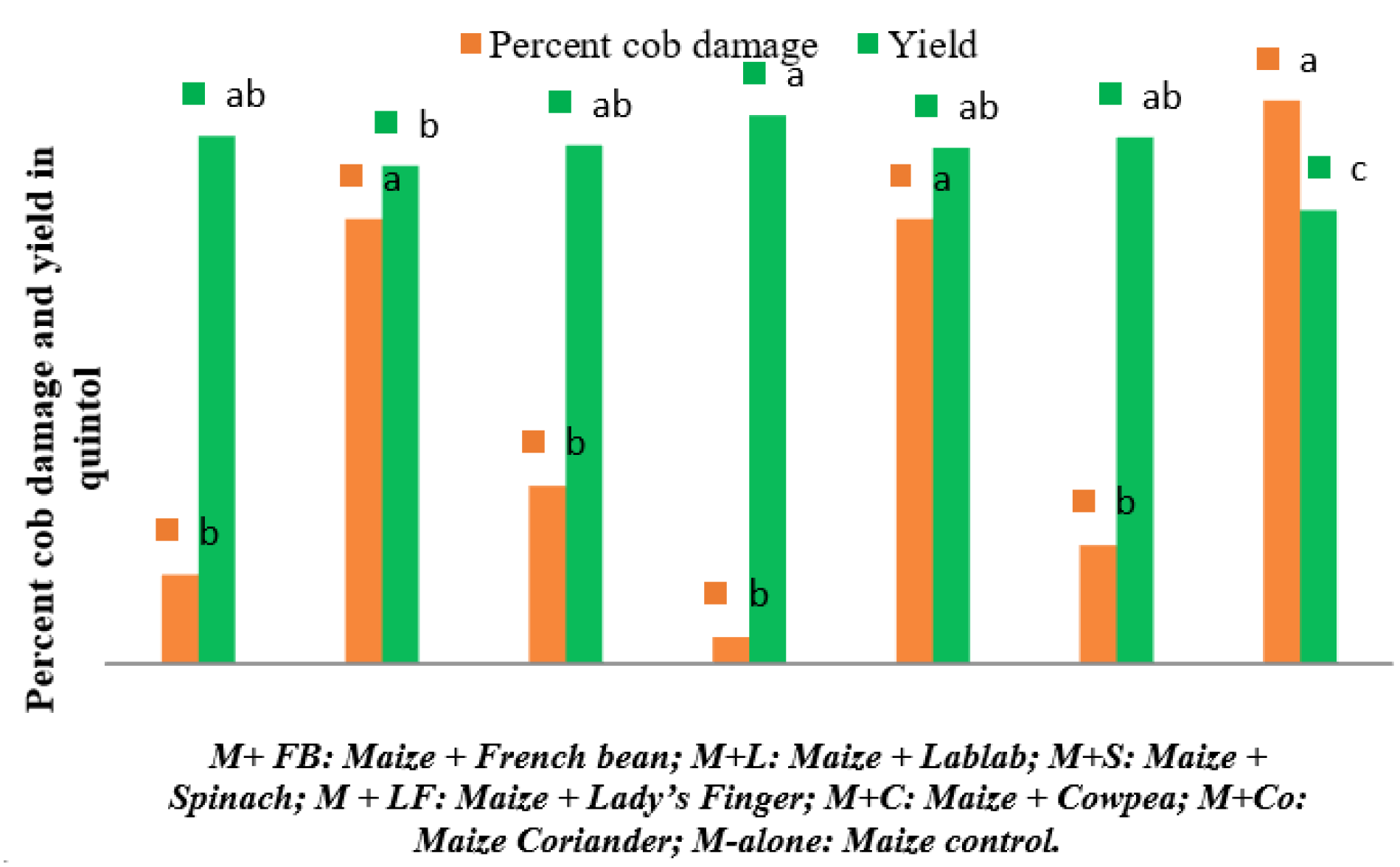
2.3. Effect of Intercropping on the Incidence of FAW
2.4. Efficacy of Biopesticides against Larvae of FAW
3. Discussion
4. Materials and Methods
4.1. Insect Rearing
4.2. Field Collection of FAW (Egg Masses and Larvae) and Laboratory Rearing
4.3. Evaluation of Botanicals against FAW
4.3.1. Preparation of Plant Extracts
4.3.2. Chemicals and Solvent
4.3.3. Preparation of Test Solution
4.3.4. Bioassay for Ovicidal Activity
4.3.5. Larvicidal Activity Bioassay by Leaf Dipping Method
4.3.6. Bioassay for Pupicidal Activity
4.4. Effect of Intercropping on the Incidence and Damage by FAW
4.5. Evaluation of Selected Bio-Pesticides against FAW
4.6. Data Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Goergen, G.; Kumar, P.L.; Sankung, S.B.; Togola, A.; Tamò, M. First report of outbreaks of the Fall Armyworm Spodoptera frugiperda (JE Smith) (Lepidoptera, Noctuidae), a new alien invasive pest in West and Central Africa. PLoS ONE 2016, 11, e0165632. [Google Scholar] [CrossRef]
- Ganiger, P.C.; Yeshwanth, H.M.; Muralimohan, K.; Vinay, N.; Kumar, A.R.V.; Chandrashekara, K. Occurrence of the new invasive pest, fall armyworm, Spodoptera frugiperda (JE Smith)(Lepidoptera: Noctuidae), in the maize fields of Karnataka, India. Curr. Sci. 2018, 115, 621–623. [Google Scholar] [CrossRef]
- Keerthi, M.C.; Mahesha, H.S.; Manjunatha, N.; Gupta, A.; Saini, R.P.; Shivakumara, K.T.; Bhargavi, H.A.; Gupta, G.; Kulkarni, N.S. Biology and oviposition preference of Fall Armyworm, Spodoptera frugiperda (JE Smith) (Lepidoptera: Noctuidae) on fodder crops and its natural enemies from Central India. Int. J. Pest Manag. 2021, 1–10. [Google Scholar] [CrossRef]
- Qi, G.J.; Ma, J.; Wan, J.; Ren, Y.L.; McKirdy, S.; Hu, G.; Zhang, Z.F. Source regions of the first immigration of Fall Armyworm, Spodoptera frugiperda (Lepidoptera: Noctuidae) invading Australia. Insects 2021, 12, 1104. [Google Scholar] [CrossRef] [PubMed]
- Devi, S. Fall Armyworm threatens food security in southern Africa. Lancet 2018, 391, 727. [Google Scholar] [CrossRef]
- Koffi, D.; Agboka, K.; Adenka, D.K.; Osae, M.; Tounou, A.K.; AnaniAdjevi, M.K.; Fening, K.O.; Meagher, R.L., Jr. Maize infestation offal Armyworm (Lepidoptera:Noctuidae) within agro-ecological zones of Togo and Ghana in West Africa 3 year after its invasion. Environ. Entomol. 2020, 49, 645–650. [Google Scholar] [CrossRef]
- Rwomushana, I.; Bateman, M.; Beale, T.; Beseh, P.; Cameron, K.; Chiluba, M.; Clottey, V.; Davis, T.; Day, R.; Early, R.; et al. Fall Armyworm: Impacts and implication for Africa; Evidence Note Update; CAB International: Wallingford, UK, 2018. [Google Scholar]
- DPPQS. 2019. Available online: http://ppqs.gov.in/sites/default/files/faw_do.pdf (accessed on 2 March 2022).
- Timilsena, B.P.; Niassy, S.; Kimathi, E.; Abdel-Rahman, E.M.; Seidl-Adams, I.; Wamalwa, M.; Tonnang, H.E.; Ekesi, S.; Hughes, D.P.; Rajotte, E.G.; et al. Potential distribution of Fall Armyworm in Africa and beyond, considering climate change and irrigation patterns. Sci. Rep. 2022, 12, 539. [Google Scholar] [CrossRef]
- Gurr, G.M.; Wratten, S.D.; Barbosa, P. Success in conservation biological control of arthropods. In Biological Control: Measures of Success; Springer: Dordrecht, The Netherlands; Kluwer Academic: Dordrecht, The Netherlands, 2000; pp. 105–132. [Google Scholar]
- Landis, D.A.; Wratten, S.D.; Gurr, G.M. Habitat management to conserve natural enemies of arthropod pests in agriculture. Annu. Rev. Entomol. 2000, 45, 175–201. [Google Scholar] [CrossRef]
- Pollard, K.A.; Holland, J.M. Arthropods within the woody element of hedge rows and their distribution pattern. Agric. For. Entomol. 2006, 8, 203–211. [Google Scholar] [CrossRef]
- Bone, N.J.; Thomson, L.J.; Ridland, P.M.; Cole, P.; Hoffmann, A.A. Cover crops in Victorian apple orchards: Effects on production, natural enemies and pests across a season. Crop Prot. 2009, 28, 675–683. [Google Scholar] [CrossRef]
- Ratnadass, A.; Fernandes, P.; Avelino, J.; Habib, R. Plant species diversity for sustainable management of crop pests and diseases in agroecosystems: A review. Agron. Sustain. Dev. 2012, 32, 273–303. [Google Scholar] [CrossRef]
- Keerthi, M.C.; Sravika, A.; Mahesha, H.S.; Gupta, A.; Bhargavi, H.A.; Ahmed, S. Performance of the native predatory bug, Eocanthecona furcellata (Wolff) (Hemiptera: Pentatomidae), on the Fall Armyworm, Spodoptera frugiperda (JE Smith) (Lepidoptera: Noctuidae), and its limitation under field condition. Egypt J. Biol. Pest Control. 2020, 30, 1–4. [Google Scholar] [CrossRef]
- Cook, S.M.; Khan, Z.R.; Pickett, J.A. The use of push-pull strategies in integrated pest management. Annu. Rev. Entomol. 2007, 52, 375–400. [Google Scholar] [CrossRef] [PubMed]
- Huss, C.P.; Holmes, K.D.; Blubaugh, C.K. Benefits and risks of intercropping for crop resilience and pest management. J. Econ. Entomol. 2022, 115, 1350–1362. [Google Scholar] [CrossRef]
- Sharanabasappa, S.; Kalleshwaraswamy, C.M.; Poorani, J.; Maruthi, M.S.; Pavithra, H.B.; Diraviam, J. Natural enemies of Spodoptera frugiperda (JE Smith) (Lepidoptera: Noctuidae),a recent invasive pest on maize in South India. Fla. Entomol. 2019, 102, 619–623. [Google Scholar]
- Myrick, S.; Norton, G.W.; Selvaraj, K.N.; Natarajan, K.; Muniappan, R. Economic impact of classical biological control of papaya mealybug in India. Crop Prot. 2014, 56, 82–86. [Google Scholar] [CrossRef]
- Sagar, D.; Sachin, S.S.; Keerthi, M.C.; Poorani, J.; Gupta, A.; Rahul, K.C. Native parasitoid complex of the invasive Fall Armyworm, Spodoptera frugiperda (J.E. Smith) from Northern India. Int. J. Trop. Insect Sci. 2021, 42, 2773–2778. [Google Scholar] [CrossRef]
- Udayakumar, A.; Shivalingaswamy, T.M.; Bakthavatsalam, N. Legume–based intercropping for the management of Fall Armyworm, Spodoptera frugiperda L. in maize. J. Plant Dis. Prot. 2021, 128, 775–779. [Google Scholar] [CrossRef]
- Navik, O.; Shylesha, A.N.; Patil, J.; Venkatesan, T.; Lalitha, Y.; Ashika, T.R. Damage, distribution and natural enemies of invasive Fall Armyworm, Spodoptera frugiperda (JE smith) under rainfed maize in Karnataka, India. Crop Prot. 2021, 143, 105536. [Google Scholar] [CrossRef]
- Sivakumar, G.; Mohan, M.; Kannan, M.; Elango, K.; RamKumar, P.; Venkatesan, T.; Rangeshwaran, R.; Mahesh, S.Y.; Dhanyakumar, O. Natural occurrence of entomopathogens on the invasive Fall Armyworm, Spodoptera frugiperda (J.E. Smith) in South India. Curr. Sci. 2020, 120, 619. [Google Scholar]
- Tiwari, S.; Youngman, R.R. Transgenic Bt corn hybrids and pest management in the USA. In Alternative Farming Systems, Biotechnology, Drought Stress and Ecological Fertilization; Springer: Dordrecht, The Netherlands, 2011; pp. 15–37. [Google Scholar]
- Tambo, J.A.; Day, R.K.; Lamontagne-Godwin, J.; Silvestri, S.; Beseh, P.K.; Oppong-Mensah, B.; Phiri, N.A.; Matimelo, M. Tackling fall armyworm (Spodoptera frugiperda) outbreak in Africa: An analysis of farmers’ control actions. Int. J. Pest Manag. 2020, 66, 298–310. [Google Scholar] [CrossRef]
- Matova, P.M.; Kamutando, C.N.; Magorokosho, C.; Kutywayo, D.; Gutsa, F.; Labuschagne, M. Fall-armyworm invasion, control practices and resistance breeding in Sub-Saharan Africa. Crop Sci. 2020, 60, 2951–2970. [Google Scholar] [CrossRef] [PubMed]
- Dudareva, N.; Negre, F.; Nagegowda, D.A.; Orlova, I. Plantvolatiles: Recent advances and future perspectives. Crit. Rev. Plant Sci. 2006, 25, 417–440. [Google Scholar] [CrossRef]
- Sombra, K.E.; deAguiar, C.V.; deOliveira, S.J.; Barbosa, M.G.; Zocolo, G.J.; Pastori, P.L. Potential pesticide of three botanicals against Spodoptera frugiperda (JE Smith) (Lepidoptera: Noctuidae). Chil. J. Agric. Res. 2020, 80, 617–628. [Google Scholar] [CrossRef]
- Negrini, M.; Fidelis, E.G.; Schurt, D.A.; Silva, F.D.S.; Pereira, R.S.; Bizzo, H.R. Insecticidal activity of botanicals in controlling Fall Armyworm, Spodoptera frugiperda. Arq. Inst. Biológico 2019, 86, 1–9. [Google Scholar]
- Mendoza-García, E.E.; Ortega-Arenas, L.D.; Serrato-Cruz, M.Á.; Villanueva-Jiménez, J.A.; López-Arroyo, J.I.; Pérez-Pacheco, R. Chemical composition, toxicity, and repellence of plant botanicals against Diaphorina citri (Hemiptera: Liviidae). Chil. J. Agric. Res. 2019, 79, 636–647. [Google Scholar] [CrossRef]
- Rani, A.T.; Kammar, V.; Keerthi, M.C.; Rani, V.; Majumder, S.; Pandey, K.K.; Singh, J. Biopesticides: An Alternative to Synthetic Insecticides. In Microbial Technology for Sustainable Environment; Springer: Singapore, 2021; pp. 439–466. [Google Scholar] [CrossRef]
- Yu, D.S.; Van Achterberg, C.; Horstmann, K. World Ichneumonoidea 2015: Taxonomy, Biology, Morphology and Distribution; CD/DVD; Taxapad: Nepean, ON, Canada, 2016. [Google Scholar]
- Achterberg, V.C. Order Hymenoptera, family Braconidae. The subfamily Agathidinae from the United Arab Emirates, with a review of the fauna of the Arabian Peninsula. Arthropod Fauna United Arab Emir. 2011, 4, 286–352. [Google Scholar]
- Sisay, B.; Simiyu, J.; Malusi, P.; Likhayo, P.; Mendesil, E.; Elibariki, N.; Wakgari, M.; Ayalew, G.; Tefera, T. First report of the Fall Armyworm, Spodoptera frugiperda (Lepidoptera: Noctuidae), natural enemies from Africa. J. Appl. Entomol. 2018, 142, 800–804. [Google Scholar] [CrossRef]
- Agboyi, L.K.; Goergen, G.; Beseh, P.; Mensah, S.A.; Clottey, V.A.; Glikpo, R.; Buddie, A.; Cafà, G.; Offord, L.; Day, R.; et al. Parasitoid complex of Fall Armyworm, Spodoptera frugiperda in Ghana and Benin. Insects 2020, 11, 68. [Google Scholar] [CrossRef]
- Otim, M.H.; AdumoAropet, S.; Opio, M.; Kanyesigye, D.; NakeletOpolot, H.; TekTay, W. Parasitoid distribution and parasitism of the Fall Armyworm, Spodoptera frugiperda (Lepidoptera: Noctuidae) in different maize producing regions of Uganda. Insects 2021, 12, 121. [Google Scholar] [CrossRef]
- Van Lenteren, J.C.; Bueno, V.H. Augmentative biological control of arthropods in Latin America. BioControl 2003, 48, 123–139. [Google Scholar] [CrossRef]
- Gause, G.F. The Struggle for Existence; Williams & Wilkins: Baltimore, MD, USA, 1934; 163p. [Google Scholar]
- Mitchell, P.; Arthur, W. Extinction due to evolution of a competitor. Evolution 1991, 45, 307–313. [Google Scholar] [CrossRef] [PubMed]
- Reitz, S.R.; Trumble, J.T. Competitive displacement among insects and arachnids. Annu. Rev. Entomol. 2002, 47, 435–465. [Google Scholar] [CrossRef]
- Ribeiro, L.P.; Dequech, S.T.; Camera, C.; Sturza, V.S.; Poncio, S.; Vendramim, J.D. Vertical and temporal distribution of Spodoptera frugiperda (JE Smith) (Lepidoptera: Noctuidae) egg masses, parasitized and non-parasitized, on maize plants. Maydica 2014, 59, 315–320. [Google Scholar]
- Regnault-Roger, C.; Vincent, C.; Arnason, J.T. Botanicals in insect control: Low risk products in a high-state world. Annu. Rev. Entomol. 2012, 57, 405–424. [Google Scholar] [CrossRef] [PubMed]
- Isman, M.B. A renaissance for botanical insecticides? Pest Manag. Sci. 2015, 71, 1587–1590. [Google Scholar] [CrossRef]
- Muturi, E.J.; Ramirez, J.L.; Zilkowski, B.; Flor-Weiler, L.B.; Rooney, A.P. Ovicidal and larvicidal effects of garlic and asafoetidabotanicals against West Nile virus vectors. J. Insect Sci. 2018, 18, 43. [Google Scholar] [CrossRef]
- Rice, P.J.; Coats, J.R. Insecticidal properties of several monoterpenoids to the house fly (Diptera: Muscidae), red flour beetle (Coleoptera: Tenebrionidae), and Southern corn root worm (Coleoptera: Chrysomelidae). J. Econ. Entomol. 1994, 87, 1172–1179. [Google Scholar] [CrossRef]
- Fujimoto, Y.; Murasaki, C.; Shimada, H.; Nishioka, S.; Kakinuma, K.; Singh, S.; Singh, M.; Gupta, Y.K.; Sahai, M. Annonaceous acetogenins from the seeds of Annona squamosa. Non-adjacent bis-tetrahydrofuran icacetogenins. Chem. Pharm. Bull. 1994, 42, 1175–1184. [Google Scholar] [CrossRef]
- Khalequzzaman, M.; Sultana, S. Insecticidal activity of Annona squamosa L. seed extracts against the red flour beetle, Tribolium castaneum (Herbst). J. Biosci. 2006, 14, 107–112. [Google Scholar] [CrossRef]
- El-Wakeil, N.E. Botanical pesticides and their mode of action. Gesunde Pflanz. 2013, 65, 125–149. [Google Scholar] [CrossRef]
- Campos, E.V.; Proença, P.L.; Oliveira, J.L.; Bakshi, M.; Abhilash, P.C.; Fraceto, L.F. Use of botanical insecticides for sustainable agriculture: Future perspectives. Ecol. Indic. 2018, 105, 1–13. [Google Scholar] [CrossRef]
- Hailu, G.; Niassy, S.; Khan, Z.R.; Ochatum, N.; Subramanian, S. Maize–legume intercropping and push–pull for management of Fall Armyworm, stemborers, and striga in Uganda. Agron. J. 2018, 110, 2513–2522. [Google Scholar] [CrossRef]
- Saminathan, V.R.; Murali Baskaran, R.K.; Mahadevan, N.R. Studies on the influence of intercropping on the conservation of Chrysoperla carnea (Stephens) for management of insect pests of cotton, Abstract. In Biotechnological Application for Integrated Pest Management; Ignacimuthu, S., Sen, A., Janarthanan, S., Eds.; Oxford & IBH Publishing Co., Pvt. Ltd.: New Delhi, India, 2002; pp. 107–110. [Google Scholar]
- Risch, S.J. A comparison by sweep sampling of the insect fauna from corn and sweet potato monoculture and dicultures inCosta Rica. Oecologia 1979, 42, 195–211. [Google Scholar] [CrossRef]
- Degri, M.M.; Mailafiya, D.M.; Mshelia, J.S. Effect of intercropping pattern on stem borer infestation in pearlmillet (Pennisetum glaucum L.) grown in the Nigerian Sudan Savannah. Adv. Entomol. 2014, 2, 81–86. [Google Scholar] [CrossRef][Green Version]
- Tanyi, C.B.; Nkongho, R.N.; Okolle, J.N.; Tening, A.S.; Ngosong, C. Effect of intercropping beans with maize and botanical extract on Fall Armyworm (Spodoptera frugiperda) infestation. Int. J. Agron. 2020, 26, 1–7. [Google Scholar] [CrossRef]
- Dhobi, C.B.; Zala, M.B.; Verma, H.S.; Sisodiya, D.B.; Thumar, R.K.; Patel, M.B.; Patel, J.K.; Borad, P.K. Evaluation of Bio-pesticides against Fall Armyworm, Spodoptera frugiperda (JE Smith) in Maize. Int. J. Curr. Microbiol. App. Sci. 2020, 9, 1150–1160. [Google Scholar] [CrossRef]
- Deshmukh, S.; Pavithra, H.B.; Kalleshwaraswamy, C.M.; Shivanna, B.K.; Maruthi, M.S.; Mota-Sanchez, D. Field efficacy of insecticides for management of invasive Fall Armyworm, Spodoptera frugiperda (JE Smith) (Lepidoptera: Noctuidae) on maize in India. Fla. Entomol. 2020, 103, 221–227. [Google Scholar] [CrossRef]
- Gujar, G.T.; Mittal, A.; Kumari, A.; Kalia, V. Host crop influence on the susceptibility of the American bollworm, Helicoverpa armigera, to Bacillus thuringiensis ssp. kurstaki HD-73. Entomol. Exp. Appl. 2004, 113, 165–172. [Google Scholar] [CrossRef]
- Sangha, J.S.; Astatkie, T.; Cutler, G.C. Ovicidal, larvicidal, and behavioral effects of some plant botanicals on diamond backmoth (Lepidoptera: Plutellidae). Can. Entomol. 2017, 149, 639–648. [Google Scholar] [CrossRef]
- Abbott, W.S. A method of computing the effectiveness of an insecticide. J. Econ. Entomol. 1925, 18, 265–267. [Google Scholar] [CrossRef]
- Baskar, K.; Kingsley, S.; Vendan, S.E.; Paulraj, M.G.; Duraipandiyan, V.; Ignacimuthu, S. Antifeedant, larvicidal and pupicidal activities of Atalantia monophylla (L) Correa against Helicoverpa armigera Hubner (Lepidoptera: Noctuidae). Chemosphere 2009, 75, 355–359. [Google Scholar] [CrossRef] [PubMed]
- Davis, F.M.; Williams, W.P. Visual Rating Scales for Screening Whorl-Stage Corn for Resistance to Fall Armyworm; Technical Bulletin186; Mississippi Agricultural and Forestry Research Experiment Station, Mississippi State University: Starkville, MS, USA, 1992. [Google Scholar]
- WASP-Web Agri Stat Package 2.0. Available online: https://ccari.icar.gov.in/wasp2.0/index.php (accessed on 20 April 2022).
- Legaspi, C.J.; French, J.V.; Garza Zuniga, A.; Legaspi, B.C. Population dynamics of the citrus leaf miner, Phyllocnistic citrella (Lep: Gracillariidae), and its natural enemies in Texas and México. Biol. Control. 2001, 21, 84–90. [Google Scholar] [CrossRef]
- Molina-Ochoa, J.; Hamm, J.J.; Lezama-Gutiérrez, R.; Lopez-Edwards, M.; Gonzalez-Ramirez, M.; Pescador-Rubio, A. A survey of Fall Armyworm (Lepidoptera: Noctuidae) parasitoids in the Mexican states of Michoacán, Colima, Jalisco, and Tamaulipas. Fla. Entomol. 2001, 84, 31–36. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parasitoid Species | Host Stage Attacked | July, 2021 (n = 926) | August, 2021 (n = 1036) | September, 2021 (n = 557) | October, 2021 (n = 222) | Relative Contribution to Total Parasitism (RP) | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| PE | RA (%) | PR (%) | PE | RA (%) | PR (%) | PE | RA (%) | PR (%) | PE | RA (%) | PR (%) | |||
| Chelonus nr. blackburni | Egg-larva | 175 | 43.77 | 18.57 | 125 | 32.30 | 12.07 | 4 | 2.53 | 0.72 | 0 | - | - | 10.98 |
| Chelonus formosanus | Egg-larva | 126 | 32.06 | 13.61 | 126 | 32.56 | 12.16 | 72 | 45.57 | 12.93 | 20 | 29.85 | 9.01 | 12.55 |
| Coccygidium sp. | Larva | 10 | 2.54 | 1.08 | 53 | 13.70 | 5.12 | 47 | 29.75 | 8.44 | 23 | 34.33 | 10.36 | 4.85 |
| Temelucha sp. | Larva | 12 | 3.05 | 1.30 | 42 | 10.85 | 4.05 | 5 | 3.16 | 0.90 | 0 | - | - | 2.15 |
| Cotesia ruficrus | Larva | 3 | 0.76 | 0.32 | 1 | 0.26 | 0.10 | 0 | - | - | 0 | - | - | 0.15 |
| Campoletis chlorideae | Larva | 3 | 0.76 | 0.32 | 3 | 0.78 | 0.29 | 1 | 0.63 | 0.18 | 0 | - | - | 0.26 |
| Dead parasitoids from field collected larvae | Egg-larva/Larva | 67 | 17.05 | 7.24 | 37 | 9.56 | 3.57 | 29 | 18.35 | 5.21 | 24 | 35.82 | 10.81 | 5.73 |
| Larva infected with entomopathogens | Larva | 11 | - | - | 32 | - | - | 9 | - | - | - | - | - | 1.90 |
| Total dead larvae | Larva | 59 | - | - | 122 | - | - | 108 | - | - | 42 | - | - | |
| Total PR (%) (n = 2741) | 42.44 | 37.36 | 28.37 | 30.18 | ||||||||||
| Parasitoids | July, 2021 (n = 64) | August, 2021 (n = 108) | September, 2021 (n = 115) | Total Egg Mass Collected (287) |
|---|---|---|---|---|
| % Egg MassParasitized | % Egg Mass Parasitized | % Egg Mass Parasitized | % of Total Egg Mass Parasitized | |
| T. chilonis | 56.25 | 24.05 | - | 21.60 |
| T. remus | - | - | 23.48 | 5.57 |
| T. chilonis + T. remus | - | 7.41 | 6.96 | 9.41 |
| Total percent parasitized | 56.25 | 31.46 | 30.44 | 36.58 |
| Sl. No | Botanicals (1%) | % Egg Hatch | % Inhibition of Egg Hatch |
|---|---|---|---|
| 1 | Citronellal | 65.41 ± 15.25 cd | 34.59 ± 15.25 cd |
| 2 | Citronella Oil | 64.03 ± 12.39 cd | 35.97 ± 12.39 cd |
| 3 | Annona | 88.61 ± 21.61 ab | 11.39 ± 21.61 ef |
| 4 | Citronellol | 87.60 ± 16.79 ab | 12.40 ± 16.79 ef |
| 5 | Geranium Oil | 95.31 ± 5.15 a | 4.69 ± 5.15 f |
| 6 | Picro | 92.69 ± 7.34 a | 7.31 ± 7.34 f |
| 7 | Geraniol | 9.24 ± 8.88 f | 90.76 ± 8.88 a |
| 8 | Clove, Syzygium aromaticum | 59.91 ± 19.53 cd | 40.09 ± 19.53 cd |
| 9 | Annona methanoic Extract | 39.37 ± 13.11 e | 60.63 ± 13.11 b |
| 10 | Sesame Oil | 87.13 ± 12.38 ab | 12.87 ± 12.38 ef |
| 11 | Chilli | 50 ± 23.59 de | 50 ± 23.59 bc |
| 12 | Karanjin | 91.65 ± 10.59 a | 8.35 ± 10.59 f |
| 13 | Garlic | 0.00 f | 100 ± 0.00 a |
| 14 | Annona Acetone Extract | 74.13 ± 8.80 bc | 25.87 ± 8.80 de |
| 15 | Control | 99.02 ± 1.53 a | 0.98 ± 1.53 f |
| F value | 26.183 | 26.183 | |
| p value | <0.0001 | <0.0001 | |
| Sl. No | Botanicals (1%) | % Mortality of Fall Armyworm Larvae | % Mortality of Pupae | Other Remarks | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| 24 h | 48 h | 72 h | 168 h | Larval Viability | Pupal Viability | Other Remarks | ||||
| 1 | Citronellal | 0 a | 30 abc | 30 abc | 30 abc | 100 | 85.7 | - | - | - |
| 2 | Citronella oil | 10 a | 60 c | 90 d | 100 e | 0 | 0 | - | 10 | - |
| 3 | Annona | 0 a | 10 ab | 40 bc | 100 e | 0 | 0 | AFA | - | - |
| 4 | Citronellol | 10 a | 10 ab | 20 abc | 90 de | 100 | 100 | AFA | - | - |
| 5 | Geranium oil | 40 b | 40 bc | 40 bc | 90 de | 100 | 100 | - | 10 | - |
| 6 | Picro | 10 a | 10 ab | 10 ab | 10 ab | 88.9 | 62.5 | - | - | AWD (20%) |
| 7 | Geraniol | 10 a | 20 ab | 40 bc | 40 bc | 16.7 | 100 | - | - | AWD (10%) |
| 8 | Clove | 0 a | 20 ab | 40 bc | 50 c | 60.0 | 66.7 | AFA | - | EM |
| 9 | Annona methane extract | 0 a | 10 ab | 10 ab | 10 ab | 77.8 | 71.4 | - | - | - |
| 10 | Sesame oil | 10 a | 10 ab | 40 c | 50 c | 100 | 60 | AWD | 10 | AWD (20%) |
| 11 | Chilli | 0 a | 0 a | 40 bc | 40 bc | 66.7 | 50 | PTE | - | EM |
| 12 | Karanjin | 0 a | 0 a | 10 ab | 10 ab | 100 | 88.9 | AWD | 10 | AWD (30%) |
| 13 | Garlic | 10 a | 20 ab | 40 bc | 60 cd | 75.0 | 66.7 | - | - | - |
| 14 | Annona acetone extract | 0 a | 0 a | 10 ab | 10 ab | 77.8 | 85.7 | - | - | AWD (40%) |
| 15 | Control | 0 a | 0 a | 0 a | 0 a | 100 | 100 | - | - | - |
| CV | 24.05 | 41.23 | 62.7 | 70.7 | ||||||
| CD @5% and @1% | 0.198 * NS@1% | 0.304 *; 0.399 ** | 0.378 *; 0.497 ** | 0.335 *; 0.440 ** | - | - | - | - | - | |
| Treatments | 5WAS | 6WAS | 7WAS | 8WAS | 9WAS | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| NOL | DS | NOL | DS | PWE | NOL | DS | PWE | NOL | DS | PWE | NOL | DS | PWE | |
| M + FB | 0.22 ± 0.38 | 0.78 ± 1.35 b | 0.83 ± 0.17 | 2.83 ± 2.52 | 0.67 ± 1.15 | 0.78 ± 0.05 a | 3.96 ± 1.03 b | 11.33 ± 1.53 b | 0.72 ± 0.04 b | 3.61 ± 0.53 b | 19.00 ± 9.64 ab | 0.54 ± 0.13 b | 3.65 ± 0.33 b | 8.33 ± 2.08 b |
| M + L | 0.00 | 0.00 b | 0.74 ± 0.22 | 2.55 ± 2.36 | 0.00 | 0.70 ± 0.17 a | 3.95 ± 0.55 b | 13.00 ± 2 b | 0.56 ± 0.09 c | 3.85 ± 0.69 b | 15.33 ± 4.04 bc | 0.64 ± 0.01 b | 3.46 ± 0.77 b | 12.33 ± 4.04 b |
| M + S | 0.22 ± 0.38 | 1.11 ± 1.92 b | 0.90 ± 0.09 | 4.22 ± 0.28 | 1.00 ± 1.73 | 0.84 ± 0.01 a | 3.44 ± 0.76 b | 12.00 ± 2.65 b | 0.69 ± 0.06 bc | 3.85 ± 0.79 b | 9.33 ± 6.66 bc | 0.68 ± 0.06 b | 3.82 ± 0.28 b | 8.00 ± 2.65 b |
| M + LF | 0.33 ± 0.58 | 0.00 b | 0.53 ± 0.47 | 2.05 ± 1.85 | 0.00 | 0.70 ± 0.17 a | 2.96 ± 0.14 b | 1.67 ± 0.58 c | 0.57 ± 0.14 c | 3.29 ± 1.21 b | 5.33 ± 5.13 c | 0.61 ± 0.1 b | 3.51 ± 0.08 b | 6.67 ± 1.15 b |
| M + C | 0.27 ± 0.46 | 1.00 ± 1.73 b | 0.82 ± 0.06 | 4.18 ± 0.55 | 1.67 ± 1.53 | 0.85 ± 0 a | 4.06 ± 0.55 b | 17.33 ± 7.57 ab | 0.70 ± 0.1 bc | 3.81 ± 1.29 b | 18.67 ± 7.23 ab | 0.70 ± 0.09 b | 4.49 ± 0.88 ab | 7.33 ± 4.04 b |
| M + Co | 0.00 | 0.00 b | 0.88 ± 0.1 | 3.95 ± 0.65 | 1.00 ± 1 | 0.67 ± 0.20 a | 3.51 ± 1.20 b | 11.00 ± 1 b | 0.66 ± 0.01 bc | 3.06 ± 0.06 b | 11.00 ± 5.29 bc | 0.68 ± 0.16 b | 3.37 ± 0.25 b | 8.67 ± 1.53 b |
| M-alone | 0.49 ± 0.43 | 4.07 ± 1.01 a | 0.94 ± 0.05 | 3.57 ± 1.22 | 2.33 ± 2.08 | 1.10 ± 0.19 b | 5.95 ± 0.47 a | 22.67 ± 10.79 a | 1.01 ± 0.08 a | 5.86 ± 0.39 a | 29.00 ± 9.85 a | 1.28 ± 0.26 a | 5.71 ± 1.12 a | 34.00 ± 16 a |
| p value | 0.768 (NS) | 0.019 | 0.420 (NS) | 0.401 (NS) | 0.356 (NS) | 0.028 | 0.016 | 0.007 | 0.0001 | 0.035 | 0.026 | 0.0001 | 0.012 | 0.003 |
| F value | 0.540 | 4.069 | 1.092 | 1.131 | 1.231 | 3.612 | 4.234 | 5.283 | 10.231 | 3.378 | 3.701 | 11.063 | 4.543 | 6.387 |
| 5WAS | 6WAS | 7WAS | 8WAS | 9WAS | |
|---|---|---|---|---|---|
| Maize + French bean | 2.63 | 12.29 bc | 18.57 bc | 27.07 b | 32.24 b |
| Maize + Lablab | 0.00 | 15.98 bc | 25.61 b | 32.47 b | 32.07 b |
| Maize + Spinach | 1.33 | 18.61 bc | 22.57 bc | 27.80 b | 30.62 b |
| Maize + Ladies Finger | 1.28 | 11.39 c | 12.25 c | 12.74 c | 13.81 c |
| Maize + Cowpea | 2.31 | 18.15 bc | 30.52 b | 35.04 b | 30.01 b |
| Maize + Coriander | 0.00 | 23.34 ab | 27.86 b | 30.33 b | 30.82 b |
| Monocrop of maize | 2.52 | 33.02 a | 48.86 a | 61.99 a | 67.10 a |
| F value | 0.781 (NS) | 3.830 | 7.307 | 11.872 | 10.711 |
| p value | 0.601 | p < 0.023 | p < 0.002 | p < 0.0001 | p < 0.0001 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Keerthi, M.C.; Suroshe, S.S.; Doddachowdappa, S.; Shivakumara, K.T.; Mahesha, H.S.; Rana, V.S.; Gupta, A.; Murukesan, A.; Casini, R.; Elansary, H.O.; et al. Bio-Intensive Tactics for the Management of Invasive Fall Armyworm for Organic Maize Production. Plants 2023, 12, 685. https://doi.org/10.3390/plants12030685
Keerthi MC, Suroshe SS, Doddachowdappa S, Shivakumara KT, Mahesha HS, Rana VS, Gupta A, Murukesan A, Casini R, Elansary HO, et al. Bio-Intensive Tactics for the Management of Invasive Fall Armyworm for Organic Maize Production. Plants. 2023; 12(3):685. https://doi.org/10.3390/plants12030685
Chicago/Turabian StyleKeerthi, Manikyanahalli Chandrashekara, Sachin Suresh Suroshe, Sagar Doddachowdappa, Kadanakuppe Thammayya Shivakumara, Hosapura Shekhararaju Mahesha, Virendra Singh Rana, Ankita Gupta, Ajith Murukesan, Ryan Casini, Hosam O. Elansary, and et al. 2023. "Bio-Intensive Tactics for the Management of Invasive Fall Armyworm for Organic Maize Production" Plants 12, no. 3: 685. https://doi.org/10.3390/plants12030685
APA StyleKeerthi, M. C., Suroshe, S. S., Doddachowdappa, S., Shivakumara, K. T., Mahesha, H. S., Rana, V. S., Gupta, A., Murukesan, A., Casini, R., Elansary, H. O., & Shakil, N. A. (2023). Bio-Intensive Tactics for the Management of Invasive Fall Armyworm for Organic Maize Production. Plants, 12(3), 685. https://doi.org/10.3390/plants12030685