
Construction of an Efficient Genetic Transformation System for Watercress (Nasturtium officinale W. T. Aiton)

 ,
, 

Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Chemical Reagents
2.3. Construction of DR5::EGFP Vector
2.4. PCR Positive Detection
2.5. Observation of GFP Fluorescence
2.6. Efficient Regeneration of Watercress
2.7. Mannitol Concentration and Treatment of Negative Pressure Time
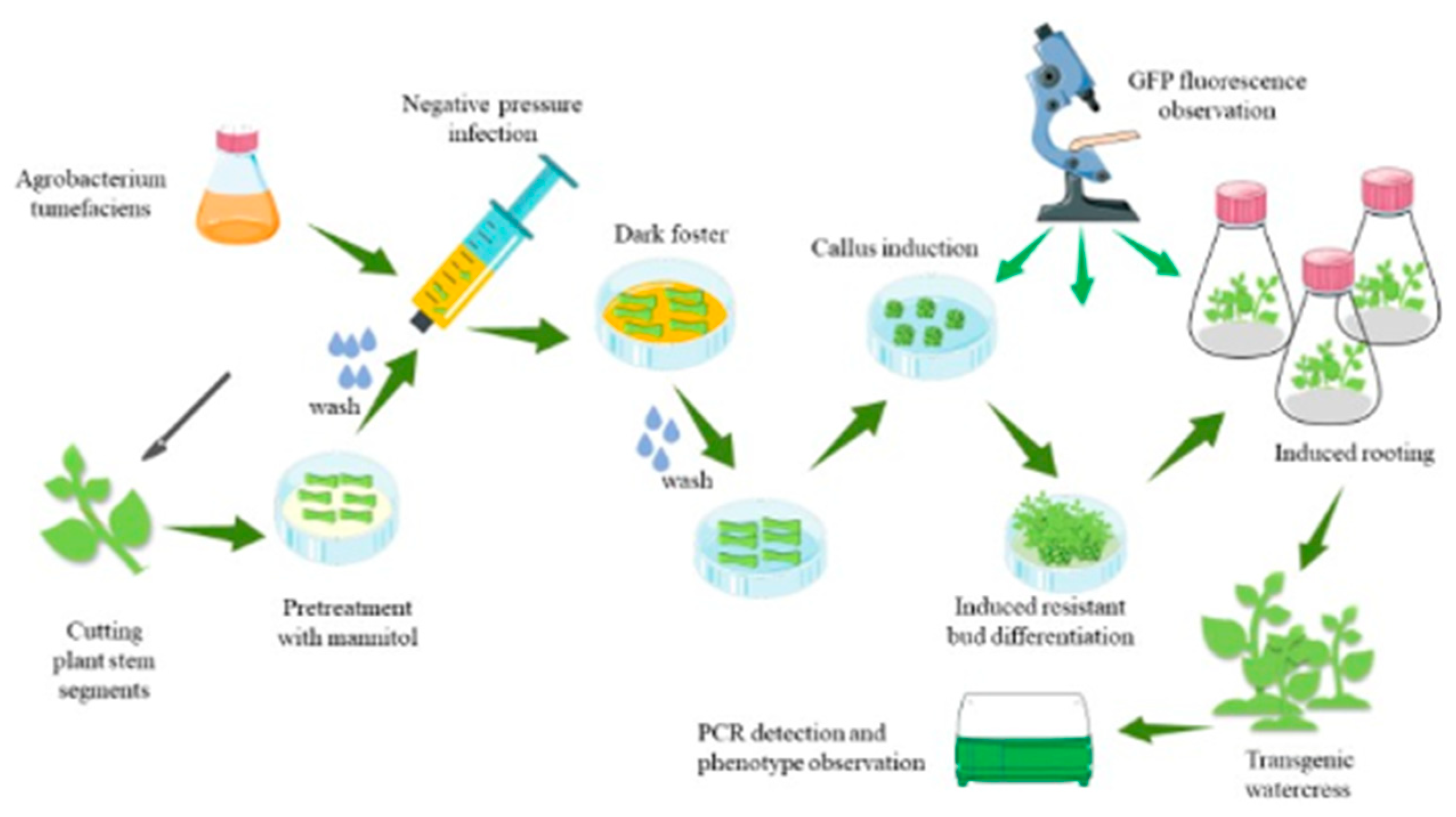
2.8. Transformation Steps
- (1)
- Select healthy watercress with consistent growth, remove the leaves, and cut the stem into sections of about 0.5 cm.
- (2)
- Place the stem in the Murashige and Skoog (MS) medium (pH 5.2, supplemented with 0.6 M mannitol + 4 mg/L 6-Benzylaminopurine (6-BA)) for 24 h of pretreatment.
- (3)
- Prepare the bacterial solution in lysogeny broth (LB) medium with antibiotics and activate it 24 h in advance. Two hours before infection, resuspend the bacteria in MS liquid medium (pH 5.2, containing 50 mg/L 3′,5′-Dimethoxy-4′-hydroxyacetophenone (AS), and remove phosphates). Then, activate the bacterial solution at 24 °C and adjust the optical density (OD) to 0.2~0.3 during infection.
- (4)
- Wash the pretreated stem with sterile water, place it in a syringe, add the bacterial solution, and seal it for 3 min of vacuum infiltration.
- (5)
- Inoculate the stem into the MS medium (pH 5.2, containing 4 mg/L 6-BA + 1.5 mg/L Thidiazuron (TDZ) + 1.5 mg/L 2,4-Dichlorophenoxyacetic acid (2,4-D) + 50 mg/L inositol + 10 mg/L AS), and cultivate in the dark at 24 °C for 3 days.
- (6)
- Wash the stem with sterile water (containing 200 mg/L cefotaxime sodium (Cef) and 100 mg/L Timentin (TMT)), dry it, and place it in the MS medium (pH 5.2, containing 4 mg/L 6-BA + 1.5 mg/L TDZ + 1.5 mg/L 2,4-D + 50 mg/L inositol + 200 mg/L Cef + 100 mg/L TMT). Induce callus formation and observe GFP fluorescence by cultivating under light for 16 h a day and at 24 °C for 15 days.
- (7)
- Place the callus on MS medium (pH 5.2, supplemented with 3 mg/L 6-BA + 3 mg/L TDZ + 200 mg/L Kanamycin (Kan)), cultivate under light for 16 h a day at 24 °C for 15 days, and induce the differentiation of large amounts of resistant buds.
- (8)
- Inoculate the resistant buds into the MS medium (pH 5.7, containing 200 mg/L Kan), cultivate under light for 16 h a day at 24 °C for 7 days, induce root formation, and observe. Before transplanting the seedlings, the cap of the tissue culture bottle was half-opened and half-closed 3 days in advance, and the regenerated plants of watercress were domesticated in the light culture box, and then, the seedlings were removed from the tissue culture bottle.
- (9)
- The regenerated plants were adapted to tissue culture in vitro (it was cultured in water with Hogan’s solution and cultured at 24 °C for 16 h), and PCR detection was performed.
2.9. Statistical Analysis
3. Results
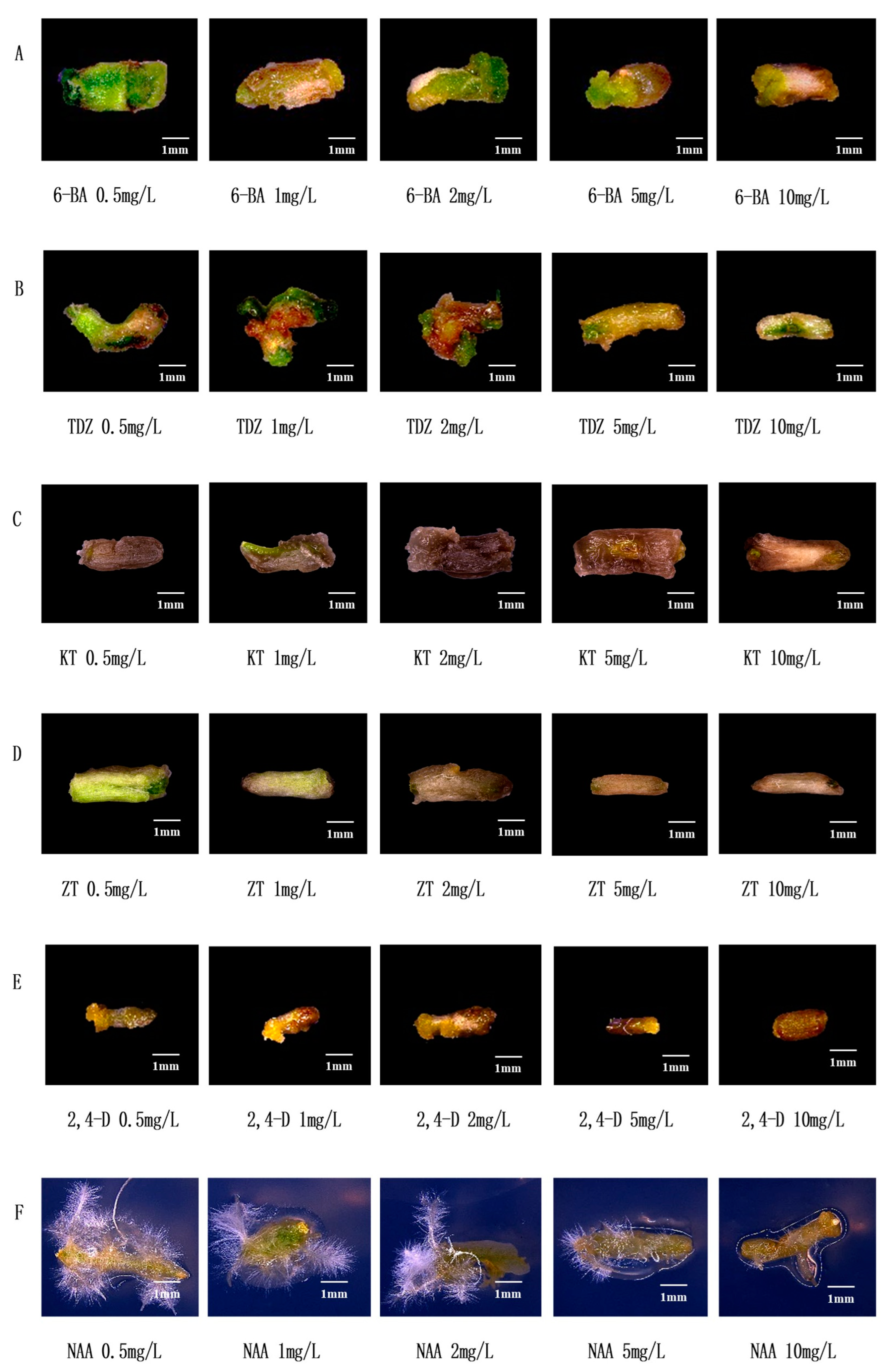
3.1. Effects of Common Hormones on Stem Segments of Watercress
3.2. Establishment and Optimization of Regeneration System
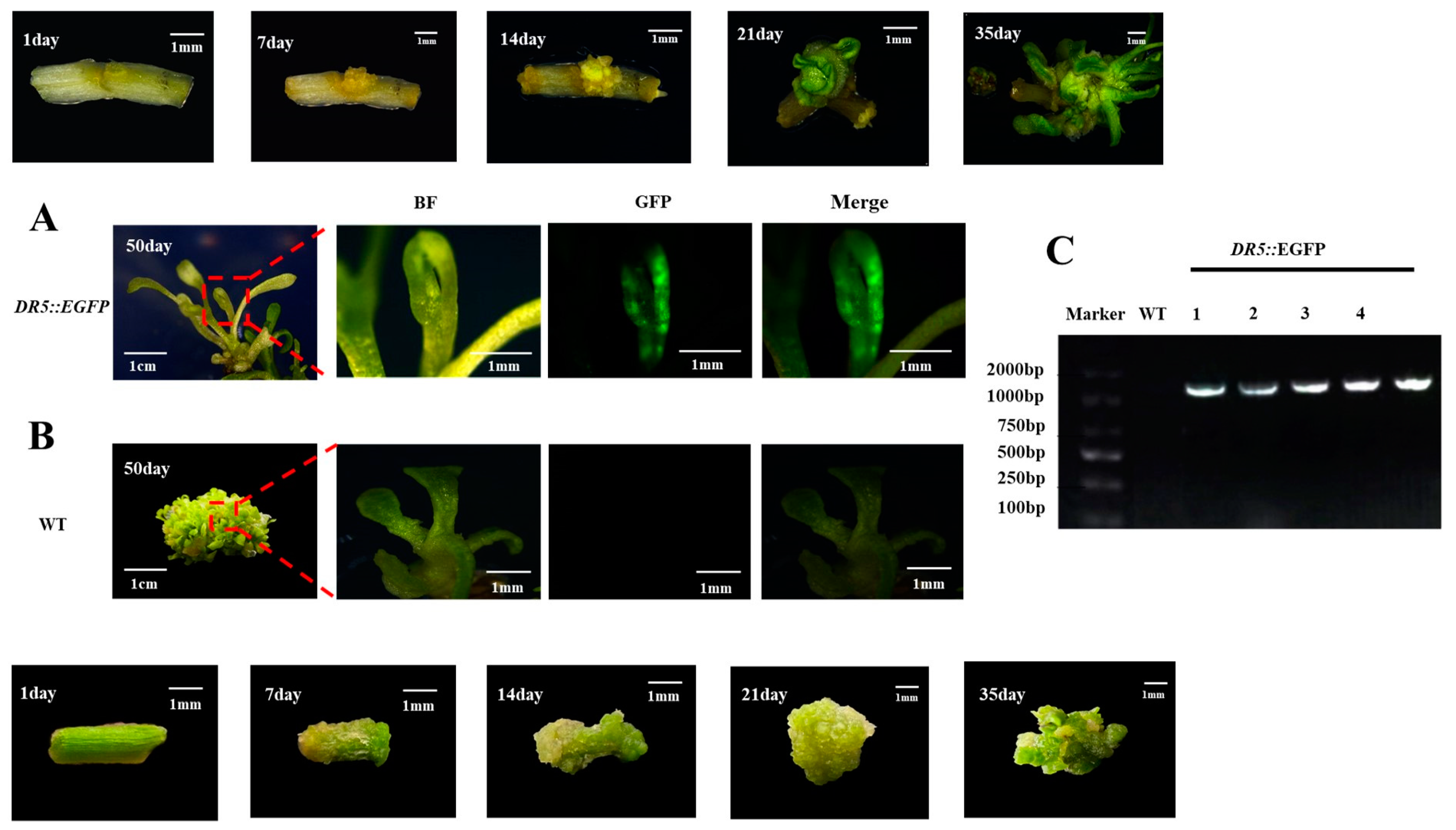
3.3. Genetic Transformation Steps
3.4. Identification of Transgenic Watercress
4. Discussion
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Clemente, M.; Miguel, M.D.; Felipe, K.B.; Gribner, C.; Moura, P.F.; Rigoni, A.G.R.; Fernandes, L.C.; Carvalho, J.L.S.; Hartmann, I.; Piltz, M.T.; et al. Acute and sub-acute oral toxicity studies of standardized extract of Nasturtium officinale in Wistar rats. Regul. Toxicol. Pharmacol. 2019, 108, 104443. [Google Scholar] [CrossRef]
- Lee, T.H.; Khan, Z.; Subedi, L.; Kim, S.Y.; Lee, K.R. New bis-thioglycosyl-1, 1′-disulfides from Nasturtium officinale R. Br. and their anti-neuroinflammatory effect. Bioorganic Chem. 2019, 86, 501–506. [Google Scholar] [CrossRef]
- Li, K.; Lin, L.; Wang, J.; Xia, H.; Liang, D.; Wang, X.; Liao, M.A.; Wang, L.; Liu, L.; Chen, C.; et al. Hyperaccumulator straw improves the cadmium phytoextraction efficiency of emergent plant Nasturtium officinale. Environ. Monit. Assess. 2017, 189, 374. [Google Scholar] [CrossRef] [PubMed]
- Ogita, S.; Usui, M.; Shibutani, N.; Kato, Y. A simple shoot multiplication procedure using internode explants, and its application for particle bombardment and Agrobacterium-mediated transformation in watercress. J. Plant Res. 2009, 122, 455–463. [Google Scholar] [CrossRef] [PubMed]
- Vissenberg, K.; Claeijs, N.; Balcerowicz, D.; Schoenaers, S. Hormonal regulation of root hair growth and responses to the environment in Arabidopsis. J. Exp. Bot. 2020, 71, 2412–2427. [Google Scholar] [CrossRef] [PubMed]
- Xue, C.; Li, W.; Shen, R.; Lan, P. Impacts of iron on phosphate starvation-induced root hair growth in Arabidopsis. Plant Cell Environ. 2023, 46, 215–238. [Google Scholar] [CrossRef]
- Tanaka, H.; Dhonukshe, P.; Brewer, P.B.; Friml, J. Spatiotemporal asymmetric auxin distribution: A means to coordinate plant development. Cell. Mol. Life Sci. CMLS 2006, 63, 2738–2754. [Google Scholar] [CrossRef]
- Tanaka, N.; Kato, M.; Tomioka, R.; Kurata, R.; Fukao, Y.; Aoyama, T.; Maeshima, M. Characteristics of a root hair-less line of Arabidopsis thaliana under physiological stresses. J. Exp. Bot. 2014, 65, 1497–1512. [Google Scholar] [CrossRef]
- Kleine-Vehn, J.; Wabnik, K.; Martinière, A.; Łangowski, Ł.; Willig, K.; Naramoto, S.; Friml, J. Recycling, clustering, and endocytosis jointly maintain PIN auxin carrier polarity at the plasma membrane. Mol. Syst. Biol. 2011, 7, 540. [Google Scholar] [CrossRef]
- Zhang, Y.; Xu, F.; Ding, Y.; Du, H.; Zhang, Q.; Dang, X.; Cao, Y.; Dodd, I.C.; Xu, W. Abscisic acid mediates barley rhizosheath formation under mild soil drying by promoting root hair growth and auxin response. Plant Cell Environ. 2021, 44, 1935–1945. [Google Scholar] [CrossRef]
- Tabesh, M.; Sh, M.E.; Etemadi, M.; Naddaf, F.; Heidari, F. The antibacterial activity of Nasturtium officinale extract on common oral pathogenic bacteria. Niger. J. Clin. Pract. 2022, 25, 1466–1475. [Google Scholar] [PubMed]
- He, F.; Nair, G.R.; Soto, C.S.; Chang, Y.; Hsu, L.; Ronzone, E.; DeGrado, W.F.; Binns, A.N. Molecular basis of ChvE function in sugar binding, sugar utilization, and virulence in Agrobacterium tumefaciens. J. Bacteriol. 2009, 191, 5802–5813. [Google Scholar] [CrossRef] [PubMed]
- Sadeghi, H.; Azarmehr, N.; Razmkhah, F. The hydroalcoholic extract of watercress attenuates protein oxidation, oxidative stress, and liver damage after bile duct ligation in rats. J. Cell. Biochem. 2019, 120, 14875–14884. [Google Scholar] [CrossRef] [PubMed]
- El-Bayoumy, K.; Stoner, G. Use of Freeze-dried Watercress for Detoxification of Carcinogens and Toxicants in Smokers: Implications of the Findings and Potential Opportunities. Cancer Prev. Res. 2022, 15, 139–141. [Google Scholar] [CrossRef]
- Schulze, H.; Hornbacher, J.; Wasserfurth, P.; Reichel, T.; Günther, T.; Krings, U.; Krüger, K.; Hahn, A.; Papenbrock, J.; Schuchardt, J.P. Immunomodulating Effect of the Consumption of Watercress (Nasturtium officinale) on Exercise-Induced Inflammation in Humans. Foods 2021, 10, 1774. [Google Scholar] [CrossRef] [PubMed]
- Bong, S.J.; Jeon, J.; Park, Y.J.; Kim, J.K.; Park, S.U. Identification and analysis of phenylpropanoid biosynthetic genes and phenylpropanoid accumulation in watercress (Nasturtium officinale R. Br.). Biotech 2020, 10, 260. [Google Scholar] [CrossRef] [PubMed]
- Wielanek, M.; Królicka, A.; Bergier, K.; Gajewska, E.; Skłodowska, M. Transformation of Nasturtium officinale, Barbarea verna and Arabis caucasica for hairy roots and glucosinolate-myrosinase system production. Biotechnol. Lett. 2009, 31, 917–921. [Google Scholar] [CrossRef]
- Jeon, J.; Bong, S.J.; Park, J.S.; Park, Y.-K.; Arasu, M.V.; Al-Dhabi, N.A.; Park, S.U. De novo transcriptome analysis and glucosinolate profiling in watercress (Nasturtium officinale R. Br.). BMC Genom. 2020, 18, 401. [Google Scholar] [CrossRef]
- Hibbert, L.; Taylor, G. Improving phosphate use efficiency in the aquatic crop watercress (Nasturtium officinale). Hortic. Res. 2022, 9, uhac011. [Google Scholar] [CrossRef]
- Staudt, G. The Effect of myo-Inositol on the Growth of Callus Tissue in Vitis. J. Plant Physiol. 1984, 116, 161–166. [Google Scholar] [CrossRef]
- Biffen, M.; Hanke, D.E. Reduction in the level of intracellular myo-inositol in cultured soybean (Glycine max) cells inhibits cell division. Biochem. J. 1990, 265, 809–814. [Google Scholar] [CrossRef] [PubMed]
- Freitas, E.; Aires, A.; de Santos Rosa, E.A.; Saavedra, M.J. Antibacterial activity and synergistic effect between watercress extracts, 2-phenylethyl isothiocyanate and antibiotics against 11 isolates of Escherichia coli from clinical and animal source. Lett. Appl. Microbiol. 2013, 57, 266–273. [Google Scholar] [CrossRef] [PubMed]
- Simmons, C.W.; VanderGheynst, J.S.; Upadhyaya, S.K. A model of Agrobacterium tumefaciens vacuum infiltration into harvested leaf tissue and subsequent in planta transgene transient expression. Biotechnol. Bioeng. 2008, 102, 965–970. [Google Scholar] [CrossRef] [PubMed]
- Tague, B.W.; Mantis, J. In planta Agrobacterium-mediated transformation by vacuum infiltration. Arab. Protoc. 2006, 323, 215–223. [Google Scholar]
- Voutsina, N.; Payne, A.C.; Hancock, R.D.; Clarkson, G.J.J.; Rothwell, S.D.; Chapman, M.A.; Taylor, G. Characterization of the watercress (Nasturtium officinale R. Br.; Brassicaceae) transcriptome using RNASeq and identification of candidate genes for important phytonutrient traits linked to human health. BMC Genom. 2016, 17, 378. [Google Scholar]
- Martin, R.E.; Marzol, E.; Estevez, J.M.; Muday, G.K. Ethylene signaling increases reactive oxygen species accumulation to drive root hair initiation in Arabidopsis. Development 2022, 149, dev200487. [Google Scholar] [CrossRef]
- Park, N.I.; Kim, J.K.; Park, W.T.; Cho, J.W.; Lim, Y.P.; Park, S.U. An efficient protocol for genetic transformation of watercress (Nasturtium officinale) using Agrobacterium rhizogenes. Mol. Biol. Rep. 2011, 38, 4947–4953. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Number | 6-BA mg/L | TDZ mg/L | 2,4-D mg/L | Number of Inoculated Stem Segments | Callus Rate (%) |
|---|---|---|---|---|---|
| CK | - | - | - | 300 | 0.00 ± 0.00 a |
| 1-1 | 2 | 1 | 1.5 | 300 | 80.32 ± 1.53 b |
| 1-2 | 2 | 1 | 2 | 300 | 73.67 ± 2.08 c |
| 1-3 | 2 | 1 | 2.5 | 300 | 64.67 ± 1.53 d |
| 2-1 | 2 | 1.5 | 1.5 | 300 | 84.33 ± 3.05 b |
| 2-2 | 2 | 1.5 | 2 | 300 | 79.67 ± 1.53 c |
| 2-3 | 2 | 1.5 | 2.5 | 300 | 73.68 ± 2.08 d |
| 3-1 | 3 | 1 | 1.5 | 300 | 92.67 ± 1.46 b |
| 3-2 | 3 | 1 | 2 | 300 | 84.67 ± 1.45 c |
| 3-3 | 3 | 1 | 2.5 | 300 | 76.33 ± 2.52 d |
| 4-1 | 3 | 1.5 | 1.5 | 300 | 94.34 ± 1.53 b |
| 4-2 | 3 | 1.5 | 2 | 300 | 89.00 ± 2.65 c |
| 4-3 | 3 | 1.5 | 2.5 | 300 | 85.67 ± 3.07 d |
| 5-1 | 4 | 1 | 1.5 | 300 | 91.67 ± 0.58 b |
| 5-2 | 4 | 1 | 2 | 300 | 86.01 ± 1.00 c |
| 5-3 | 4 | 1 | 2.5 | 300 | 80.33 ± 1.53 d |
| 6-1 | 4 | 1.5 | 1.5 | 300 | 98.00 ± 2.00 b |
| 6-2 | 4 | 1.5 | 2 | 300 | 95.01 ± 1.00 c |
| 6-3 | 4 | 1.5 | 2.5 | 300 | 92.67 ± 2.08 d |
| Number | 6-BA mg/L | TDZ mg/L | 2,4-D mg/L | NAA mg/L | Number of Inoculated Calli | Number of Buds |
|---|---|---|---|---|---|---|
| CK | - | - | - | - | 300 | 0 a |
| 1 | 2 | 1 | - | - | 300 | 1422 b |
| 2 | 2 | 2 | - | - | 300 | 2267 c |
| 3 | 2 | 3 | - | - | 300 | 3697 d |
| 4 | 3 | 1 | - | - | 300 | 2788 e |
| 5 | 3 | 2 | - | - | 300 | 4059 f |
| 6 | 3 | 3 | - | - | 300 | 5111 g |
| 7 | 5 | 1 | - | - | 300 | 2453 h |
| 8 | 5 | 2 | - | - | 300 | 1930 i |
| 9 | 5 | 3 | - | - | 300 | 1151 j |
| Number | NAA mg/L | Rooting Rate | Rooting Number (Root/Plant) | Root Length (cm) |
|---|---|---|---|---|
| CK | - | 100% | 21.67 ± 0.58 a | 6.30 ± 0.1 a |
| 1 | 0.3 | 100% | 35.67 ± 0.58 b | 5.33 ± 0.25 b |
| 2 | 0.5 | 100% | 30.00 ± 1 c | 3.90 ± 0.36 c |
| 3 | 0.7 | 100% | 27.00 ± 1 d | 1.63 ± 0.15 d |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ran, J.; Ding, Q.; Shen, Y.; Gao, Z.; Wang, G.; Gao, Y.; Ma, X.; Hou, X. Construction of an Efficient Genetic Transformation System for Watercress (Nasturtium officinale W. T. Aiton). Plants 2023, 12, 4149. https://doi.org/10.3390/plants12244149
Ran J, Ding Q, Shen Y, Gao Z, Wang G, Gao Y, Ma X, Hou X. Construction of an Efficient Genetic Transformation System for Watercress (Nasturtium officinale W. T. Aiton). Plants. 2023; 12(24):4149. https://doi.org/10.3390/plants12244149
Chicago/Turabian StyleRan, Jiajun, Qiang Ding, Yunlou Shen, Zhanyuan Gao, Guangpeng Wang, Yue Gao, Xiaoqing Ma, and Xilin Hou. 2023. "Construction of an Efficient Genetic Transformation System for Watercress (Nasturtium officinale W. T. Aiton)" Plants 12, no. 24: 4149. https://doi.org/10.3390/plants12244149
APA StyleRan, J., Ding, Q., Shen, Y., Gao, Z., Wang, G., Gao, Y., Ma, X., & Hou, X. (2023). Construction of an Efficient Genetic Transformation System for Watercress (Nasturtium officinale W. T. Aiton). Plants, 12(24), 4149. https://doi.org/10.3390/plants12244149