
Slow Release of GA3 Hormone from Polymer Coating Overcomes Seed Dormancy and Improves Germination
 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
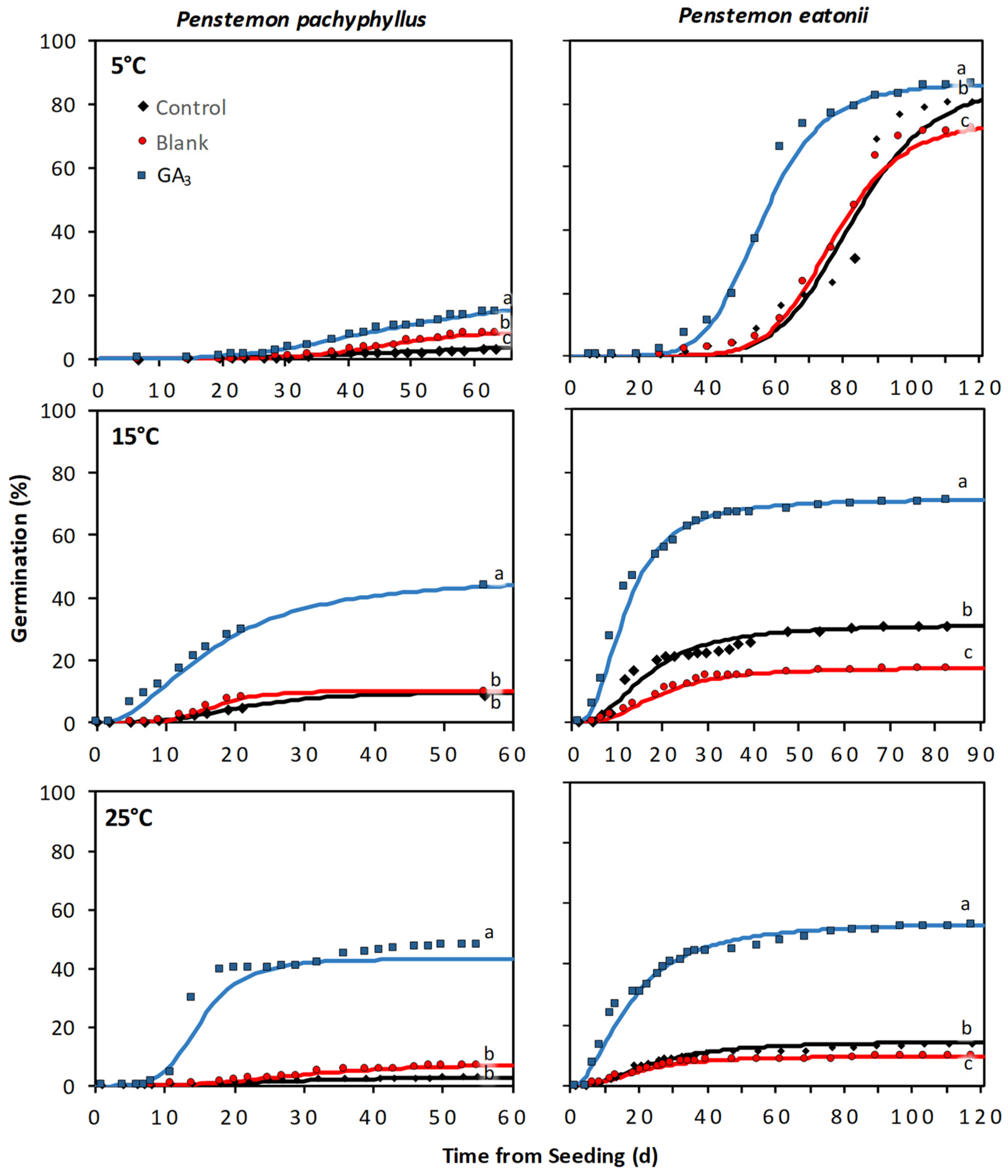
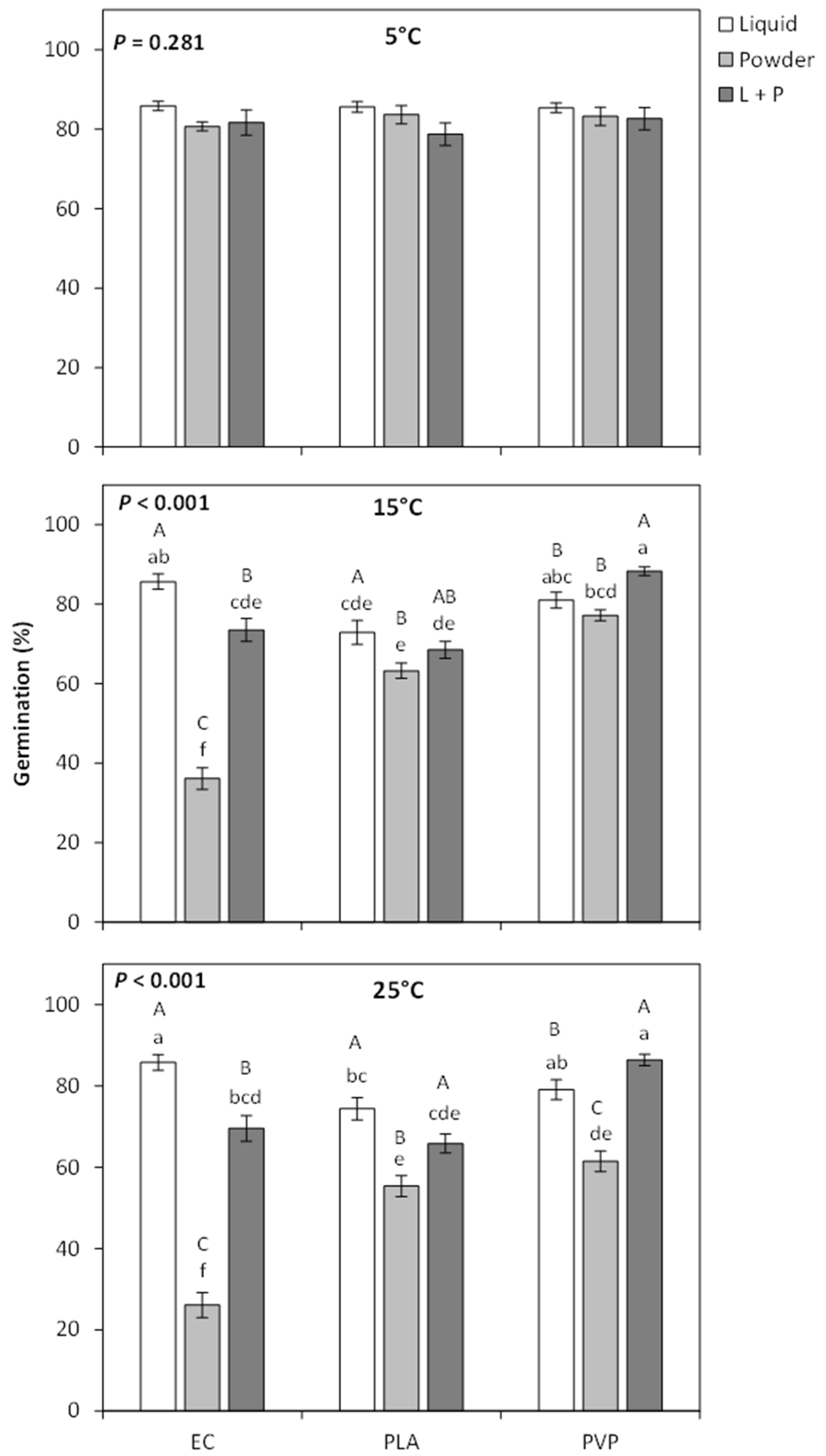
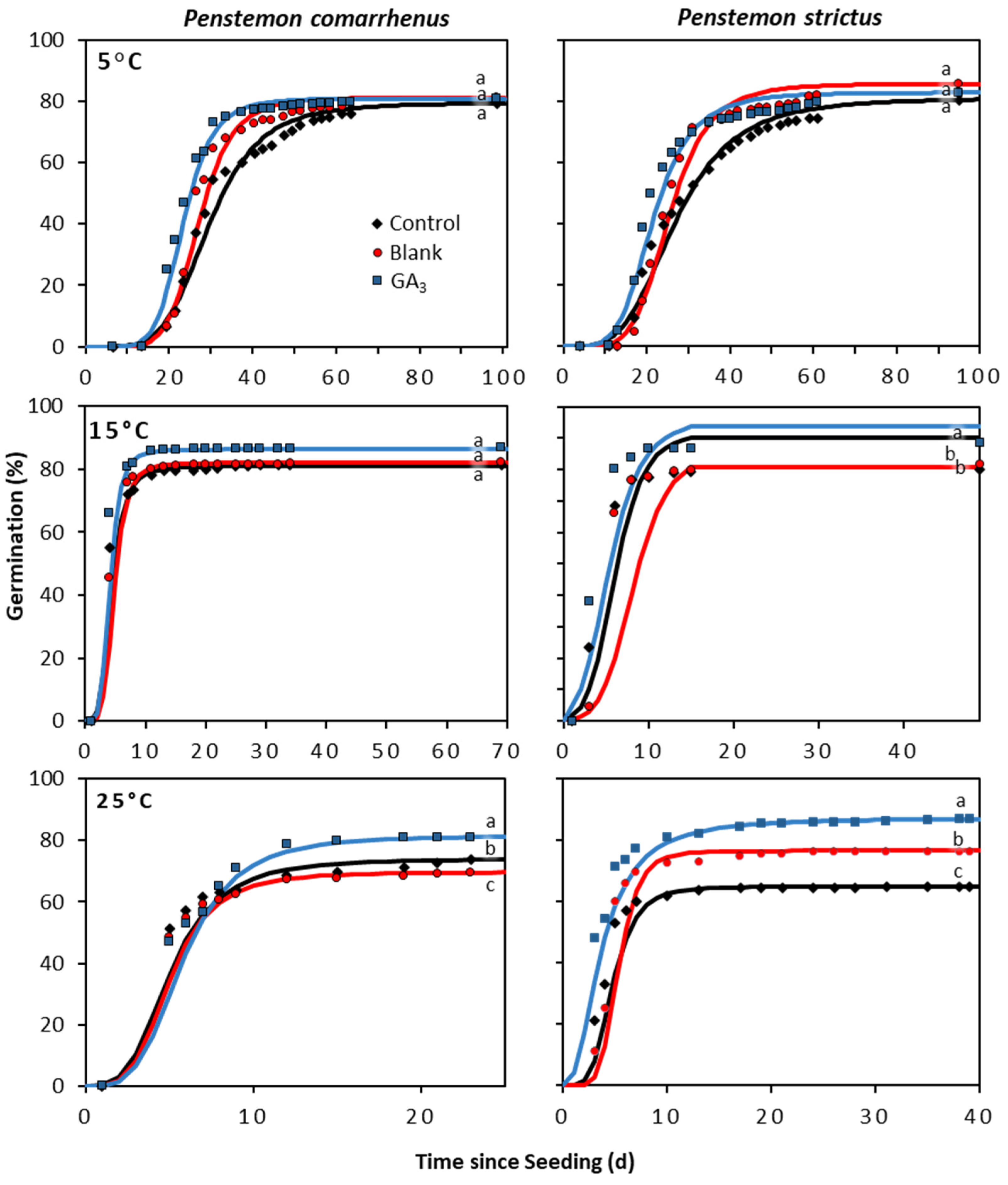
2. Results
2.1. Seed Coating Development on Penstemon palmeri
2.2. Evaluation of a GA3 Seed Coating on Additional Penstemon Species
3. Discussion
4. Materials and Methods
4.1. Study Species
4.2. Seed Coating Development on Penstemon palmeri
4.3. Evaluation of a GA3 Seed Coating on Additional Penstemon Species
4.4. Statistical Analysis
5. Patents
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ecosystems Division. The Restoration Initiative Year in Review 2019; Ecosystems Division: Millbrook, NY, USA, 2020. [Google Scholar]
- Gann, G.D.; McDonald, T.; Walder, B.; Aronson, J.; Nelson, C.R.; Jonson, J.; Hallett, J.G.; Eisenberg, C.; Guariguata, M.R.; Liu, J.; et al. International Principles and Standards for the Practice of Ecological Restoration. Second Edition. Restor. Ecol. 2019, 27, S1–S46. [Google Scholar] [CrossRef]
- Merritt, D.J.; Turner, S.R.; Clarke, S.; Dixon, K.W. Seed Dormancy and Germination Stimulation Syndromes for Australian Temperate Species. Aust. J. Bot. 2007, 55, 336–344. [Google Scholar] [CrossRef]
- Griscom, B.W.; Lomax, G.; Kroeger, T.; Fargione, J.E.; Adams, J.; Almond, L.; Bossio, D.; Cook-Patton, S.C.; Ellis, P.W.; Kennedy, C.M.; et al. We Need Both Natural and Energy Solutions to Stabilize Our Climate. Glob. Chang. Biol. 2019, 25, 1889–1890. [Google Scholar] [CrossRef] [PubMed]
- Hardegree, S.P.; Sheley, R.L.; Duke, S.E.; James, J.J.; Boehm, A.R.; Flerchinger, G.N. Temporal Variability in Microclimatic Conditions for Grass Germination and Emergence in the Sagebrush Steppe. Rangel. Ecol. Manag. 2016, 69, 123–128. [Google Scholar] [CrossRef]
- Leverkus, A.B.; Levy, L.; Andivia, E.; Annighöfer, P.; de Cuyper, B.; Ivetic, V.; Lazdina, D.; Löf, M.; Villar-Salvador, P. Restoring Oak Forests through Direct Seeding or Planting: Protocol for a Continental-Scale Experiment. PLoS ONE 2021, 16, e0259552. [Google Scholar] [CrossRef]
- Shackelford, N.; Paterno, G.B.; Winkler, D.E.; Erickson, T.E.; Leger, E.A.; Svejcar, L.N.; Breed, M.F.; Faist, A.M.; Harrison, P.A.; Curran, M.F.; et al. Drivers of Seedling Establishment Success in Dryland Restoration Efforts. Nat. Ecol. Evol. 2021, 5, 1283–1290. [Google Scholar] [CrossRef]
- James, J.J.; Svejcar, T.J.; Rinella, M.J. Demographic Processes Limiting Seedling Recruitment in Arid Grassland Restoration. J. Appl. Ecol. 2011, 48, 961–969. [Google Scholar] [CrossRef]
- Pedrini, S.; Balestrazzi, A.; Madsen, M.D.; Bhalsing, K.; Hardegree, S.P.; Dixon, K.W.; Kildisheva, O.A. Seed Enhancement: Getting Seeds Restoration-Ready. Restor. Ecol. 2020, 28, S266–S275. [Google Scholar] [CrossRef]
- Kildisheva, O.A.; Erickson, T.E.; Merritt, D.J.; Dixon, K.W. Setting the Scene for Dryland Recovery: An Overview and Key Findings from a Workshop Targeting Seed-Based Restoration. Restor. Ecol. 2016, 24, S36–S42. [Google Scholar] [CrossRef]
- Kildisheva, O.A.; Dixon, K.W.; Silveira, F.A.O.; Chapman, T.; Di Sacco, A.; Mondoni, A.; Turner, S.R.; Cross, A.T. Dormancy and Germination: Making Every Seed Count in Restoration. Restor. Ecol. 2020, 28, S256–S265. [Google Scholar] [CrossRef]
- Commander, L.E.; Golos, P.J.; Miller, B.P.; Merritt, D.J. Seed Germination Traits of Desert Perennials. Plant Ecol. 2017, 218, 1077–1091. [Google Scholar] [CrossRef]
- Baskin, C.C.; Baskin, J.M. Seeds: Ecology, Biogeography and Evolution of Dormancy and Germination; Academic Press: San Diego, CA, USA, 2001. [Google Scholar]
- Baskin, J.M.; Baskin, C.C. A Classification System for Seed Dormancy. Seed Sci. Res. 2004, 14, 1–16. [Google Scholar] [CrossRef]
- Finch-Savage, W.E.; Leubner-Metzger, G. Seed Dormancy and the Control of Germination. New Phytol. 2006, 171, 501–523. [Google Scholar] [CrossRef] [PubMed]
- Xu, D.; Cao, H.; Fang, W.; Pan, J.; Chen, J.; Zhang, J.; Shen, W. Linking Hydrogen-Enhanced Rice Aluminum Tolerance with the Reestablishment of GA/ABA Balance and MiRNA-Modulated Gene Expression: A Case Study on Germination. Ecotoxicol. Environ. Saf. 2017, 145, 303–312. [Google Scholar] [CrossRef]
- Shu, K.; Zhou, W.; Chen, F.; Luo, X.; Yang, W. Abscisic Acid and Gibberellins Antagonistically Mediate Plant Development and Abiotic Stress Responses. Front. Plant Sci. 2018, 9, 416. [Google Scholar] [CrossRef]
- Gupta, R.; Chakrabarty, S.K. Gibberellic Acid in Plant: Still a Mystery Unresolved. Plant Signal Behav. 2013, 8, e25504. [Google Scholar] [CrossRef]
- González-López, Ó.; Casquero, P.A. Effects of GA3 Pregerminative Treatment on Gentiana lutea L. Var. Aurantiaca Germination and Seedlings Morphology. Sci. World J. 2014, 2014, 751279. [Google Scholar] [CrossRef]
- Christensen, C.T.; Zotarelli, L.; Haynes, K.G.; Kelly, C.E. Comparative Evaluation of the Effects of Gibberellic Acid Concentrations on Dormancy Break in Tubers of Solanum Chacoense. Horttechnology 2020, 30, 76–81. [Google Scholar] [CrossRef]
- Ma, H.Y.; Zhao, D.D.; Ning, Q.R.; Wei, J.P.; Li, Y.; Wang, M.M.; Liu, X.L.; Jiang, C.J.; Liang, Z.W. A Multi-Year Beneficial Effect of Seed Priming with Gibberellic Acid-3 (GA3) on Plant Growth and Production in a Perennial Grass, Leymus chinensis. Sci. Rep. 2018, 8, 13214. [Google Scholar] [CrossRef]
- da Silva, A.L.B.R.; de Barros, M.F.L.; Foshee, W.; Candian, J.S.; Diaz-Perez, J.C. Priming Strategies for Parsley Seedling Production. HortScience 2022, 57, 1086–1091. [Google Scholar] [CrossRef]
- Tarquis, A.M.; Bradford, K.J. Prehydration and Priming Treatments That Advance Germination Also Increase the Rate of Deterioration of Lettuce Seeds. J. Exp. Bot. 1992, 43, 307–317. [Google Scholar] [CrossRef]
- Parera, C.A.; Cantliffe, D.J. Presowing Seed Priming. Hortic. Rev. (Am. Soc. Hortic. Sci.) 2010, 16, 109–141. [Google Scholar] [CrossRef]
- Taylor, A.; Allen, P.; Bennett, M.; Bradford, K.; Burris, J.; Misra, M. Seed Enhancements. Seed Sci. Res. 1998, 8, 245–256. [Google Scholar] [CrossRef]
- Farooq, M.; Usman, M.; Nadeem, F.; Rehman, H.U.; Wahid, A.; Basra, S.M.A.; Siddique, K.H.M. Seed Priming in Field Crops: Potential Benefits, Adoption and Challenges. Crop Pasture Sci. 2019, 70, 731–771. [Google Scholar] [CrossRef]
- Fabrissin, I.; Sano, N.; Seo, M.; North, H.M. Ageing Beautifully: Can the Benefits of Seed Priming Be Separated from a Reduced Lifespan Trade-Off? J. Exp. Bot. 2021, 72, 2312–2333. [Google Scholar] [CrossRef] [PubMed]
- Kamaly, N.; Fredman, G.; Fojas, J.J.R.; Subramanian, M.; Choi, W.I.; Zepeda, K.; Vilos, C.; Yu, M.; Gadde, S.; Wu, J.; et al. Targeted Interleukin-10 Nanotherapeutics Developed with a Microfluidic Chip Enhance Resolution of Inflammation in Advanced Atherosclerosis. ACS Nano 2016, 10, 5280–5292. [Google Scholar] [CrossRef]
- Langer, R.S.; Peppas, N.A. Present and Future Applications of Biomaterials in Controlled Drug Delivery Systems. Biomaterials 1981, 2, 201–214. [Google Scholar] [CrossRef]
- Liechty, W.B.; Kryscio, D.R.; Slaughter, B.V.; Peppas, N.A. Polymers for Drug Delivery Systems. Annu. Rev. Chem. Biomol. Eng. 2010, 1, 149–173. [Google Scholar] [CrossRef]
- Duncan, R. The Dawning Era of Polymer Therapeutics. Nat. Rev. Drug Discov. 2003, 2, 347–360. [Google Scholar] [CrossRef]
- Duncan, R. Polymer Conjugates as Anticancer Nanomedicines. Nat. Rev. Cancer 2006, 6, 688–701. [Google Scholar] [CrossRef]
- Roberts, W.C. Pulmonary Talc Granulomas, Pulmonary Fibrosis, and Pulmonary Hypertension Resulting from Intravenous Injection of Talc-Containing Drugs Intended for Oral Use. Bayl. Univ. Med. Cent. Proc. 2002, 15, 260–261. [Google Scholar] [CrossRef]
- Akalin, G.O.; Pulat, M. Controlled Release Behavior of Zinc-Loaded Carboxymethyl Cellulose and Carrageenan Hydrogels and Their Effects on Wheatgrass Growth. J. Polym. Res. 2020, 27, 6. [Google Scholar] [CrossRef]
- Jiao, Q.; Bi, L.; Ren, Y.; Song, S.; Wang, Q.; Wang, Y.S. Advances in Studies of Tyrosine Kinase Inhibitors and Their Acquired Resistance. Mol. Cancer 2018, 17, 36. [Google Scholar] [CrossRef] [PubMed]
- Neri-Badang, M.C.; Chakraborty, S. Carbohydrate Polymers as Controlled Release Devices for Pesticides. J. Carbohydr. Chem. 2019, 38, 67–85. [Google Scholar] [CrossRef]
- Campos, E.V.R.; de Oliveira, J.L.; Fraceto, L.F.; Singh, B. Polysaccharides as Safer Release Systems for Agrochemicals. Agron. Sustain. Dev. 2015, 35, 47–66. [Google Scholar] [CrossRef]
- Wolfe, A.D.; Randle, C.P.; Datwyler, S.L.; Morawetz, J.J.; Arguedas, N.; Diaz, J. Phylogeny, Taxonomic Affinities, and Biogeography of Penstemon (Plantaginaceae) Based on ITS and CpDNA Sequence Data. Am. J. Bot. 2006, 93, 1699–1713. [Google Scholar] [CrossRef] [PubMed]
- Wessinger, C.A.; Freeman, C.C.; Mort, M.E.; Rausher, M.D.; Hileman, L.C. Wessinger2016_Multiplexed Shotgun Genotyping. Am. J. Bot. 2016, 103, 912–922. [Google Scholar] [CrossRef]
- Wolfe, A.D.; Blischak, P.D.; Kubatko, L.S. Phylogenetics of a Rapid, Continental Radiation: Diversification, Biogeography, and Circumscription of the Beardtongues (Penstemon; Plantaginaceae). bioRxiv 2021. preprint. [Google Scholar] [CrossRef]
- Kramer, A.T.; Fant, J.B.; Ashley, M.V. Influences of Landscape and Pollinators on Population Genetic Structure: Examples from Three Penstemon (Plantaginaceae) Species in the Great Basin. Am. J. Bot. 2011, 98, 109–121. [Google Scholar] [CrossRef]
- Wilson, P.; Castellanos, M.C.; Hogue, J.N.; Thomson, J.D.; Armbruster, W.S. A Multivariate Search for Pollination Syndromes among Penstemons. Oikos 2004, 104, 345–361. [Google Scholar] [CrossRef]
- Fenster, C.B.; Armbruster, W.S.; Wilson, P.; Dudash, M.R.; Thomson, J.D. Pollination Syndromes and Floral Specialization. Annu. Rev. Ecol. Evol. Syst. 2004, 35, 375–403. [Google Scholar] [CrossRef]
- Kratsch, H. Penstemons Are for Great Basin Gardens FS-13-35; Rocky Mountain Research Station: Reno, NV, USA, 2013.
- Dumroese, K.R.; Luna, T.; Pinto, J.R.; Landis, T.D. Natural Areas Association. Nat. Areas J. 2016, 36, 499–511. [Google Scholar] [CrossRef]
- Tilley, D.; Hulet, A.; Bushman, S.; Goebel, C.; Karl, J.; Love, S.; Wolf, M. When a Weed is not a Weed: Succession Management Using Early Seral Natives for Intermountain Rangeland Restoration. Rangelands 2022, 44, 270–280. [Google Scholar] [CrossRef]
- Barak, R.S.; Fant, J.B.; Kramer, A.T.; Skogen, K.A. Assessing the Value of Potential “Native Winners” for Restoration of Cheatgrass-Invaded Habitat. West. N. Am. Nat. 2015, 75, 58–69. [Google Scholar] [CrossRef]
- Allen, P.S.; Meyer, S.E. Temperature Requirements for Seed Germination of Three Penstemon Species. HortScience 1990, 25, 191–193. [Google Scholar] [CrossRef]
- Meyer, S.E. Habitat Correlated Variation in Firecracker Penstemon (Penstemon Eatonii Gray: Scrophulariaceae) Seed Germination Response. Bull. Torrey Bot. Club 1992, 119, 268–279. [Google Scholar] [CrossRef]
- Kramer, A.T.; Larkin, D.J.; Fant, J.B. Assessing Potential Seed Transfer Zones for Five Forb Species from the Great Basin Floristic Region, USA. Nat. Areas J. 2015, 35, 174–188. [Google Scholar] [CrossRef]
- Miller, B.P.; Sinclair, E.A.; Menz, M.H.M.; Elliott, C.P.; Bunn, E.; Commander, L.E.; Dalziell, E.; David, E.; Davis, B.; Erickson, T.E.; et al. A Framework for the Practical Science Necessary to Restore Sustainable, Resilient, and Biodiverse Ecosystems. Restor. Ecol. 2017, 25, 605–617. [Google Scholar] [CrossRef]
- Ladouceur, E.; Jiménez-Alfaro, B.; Marin, M.; De Vitis, M.; Abbandonato, H.; Iannetta, P.P.M.; Bonomi, C.; Pritchard, H.W. Native Seed Supply and the Restoration Species Pool. Conserv. Lett. 2018, 11, e12381. [Google Scholar] [CrossRef]
- Walck, J.L.; Hidayati, S.N.; Dixon, K.W.; Thompson, K.; Poschlod, P. Climate Change and Plant Regeneration from Seed. Glob. Chang. Biol. 2011, 17, 2145–2161. [Google Scholar] [CrossRef]
- Thomson, J.D.; Wilson, P. Explaining Evolutionary Shifts between Bee and Hummingbird Pollination: Convergence, Divergence, and Directionality. Int. J. Plant Sci. 2008, 169, 23–38. [Google Scholar] [CrossRef]
- Castellanos, M.C.; Wilson, P.; Thomson, J.D. Pollen Transfer by Hummingbirds and Bumblebees, and the Divergence of Pollination Modes in Penstemon. Evolution 2003, 57, 2742–2752. [Google Scholar] [PubMed]
- Rodríguez-Morgado, B.; Jiménez, P.C.; Moral, M.T.; Rubio, J.P. Effect of L-Lactic Acid from Whey Wastes on Enzyme Activities and Bacterial Diversity of Soil. Biol. Fertil. Soils 2017, 53, 389–396. [Google Scholar] [CrossRef]
- Adeleke, O.A. Premium Ethylcellulose Polymer Based Architectures at Work in Drug Delivery. Int. J. Pharm. X 2019, 1, 100023. [Google Scholar] [CrossRef] [PubMed]
- Franco, P.; De Marco, I. The Use of Poly(N-Vinyl Pyrrolidone) in the Delivery of Drugs: A Review. Polymers 2020, 12, 1114. [Google Scholar] [CrossRef]
- Soil Survey Staff Web Soil Survey. Available online: https://websoilsurvey.sc.egov.usda.gov/App/WebSoilSurvey.aspx (accessed on 30 January 2020).
- Sileshi, G.W. A Critique of Current Trends in the Statistical Analysis of Seed Germination and Viability Data. Seed Sci. Res. 2012, 22, 145–159. [Google Scholar] [CrossRef]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Larson, A.J.S.; Cartwright, M.M.; Jones, W.D.; Luce, K.; Chen, M.-Y.; Petersen, K.; Nelson, S.V.; Michaelis, D.J.; Madsen, M.D. Slow Release of GA3 Hormone from Polymer Coating Overcomes Seed Dormancy and Improves Germination. Plants 2023, 12, 4139. https://doi.org/10.3390/plants12244139
Larson AJS, Cartwright MM, Jones WD, Luce K, Chen M-Y, Petersen K, Nelson SV, Michaelis DJ, Madsen MD. Slow Release of GA3 Hormone from Polymer Coating Overcomes Seed Dormancy and Improves Germination. Plants. 2023; 12(24):4139. https://doi.org/10.3390/plants12244139
Chicago/Turabian StyleLarson, Alexandra J. S., Maureen M. Cartwright, Whitney D. Jones, Katrina Luce, Mei-Yu Chen, Kate Petersen, Shannon V. Nelson, David J. Michaelis, and Matthew D. Madsen. 2023. "Slow Release of GA3 Hormone from Polymer Coating Overcomes Seed Dormancy and Improves Germination" Plants 12, no. 24: 4139. https://doi.org/10.3390/plants12244139
APA StyleLarson, A. J. S., Cartwright, M. M., Jones, W. D., Luce, K., Chen, M.-Y., Petersen, K., Nelson, S. V., Michaelis, D. J., & Madsen, M. D. (2023). Slow Release of GA3 Hormone from Polymer Coating Overcomes Seed Dormancy and Improves Germination. Plants, 12(24), 4139. https://doi.org/10.3390/plants12244139