
Unveiling Diversity for Quality Traits in the Indian Landraces of Horsegram [Macrotyloma uniflorum (Lam.) Verdc.]
 , ,
, ,  ,
,
Abstract
:1. Introduction
2. Results
2.1. Nutritional Composition
2.2. Correlation Analysis
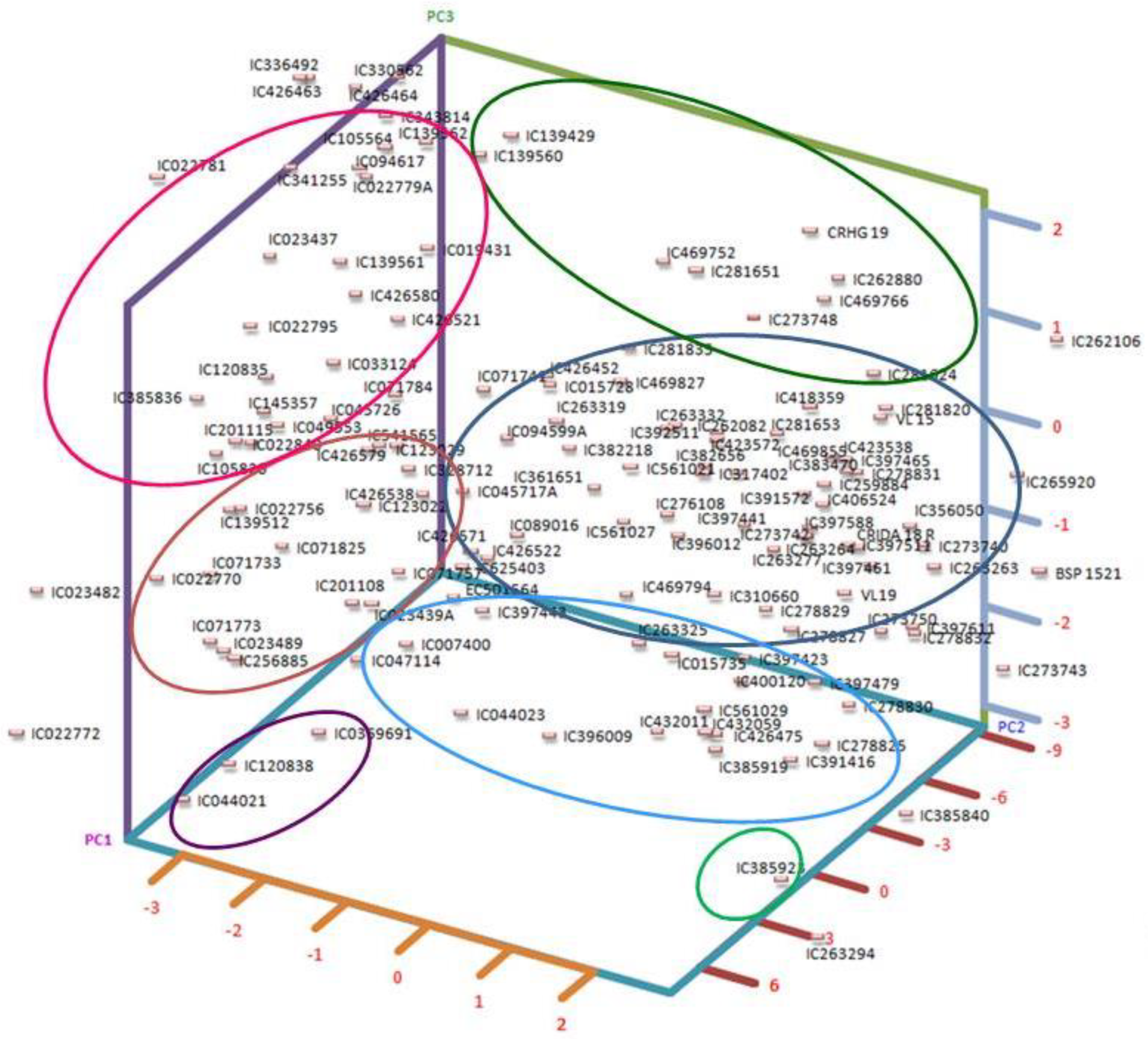
2.3. Principal Component Analysis
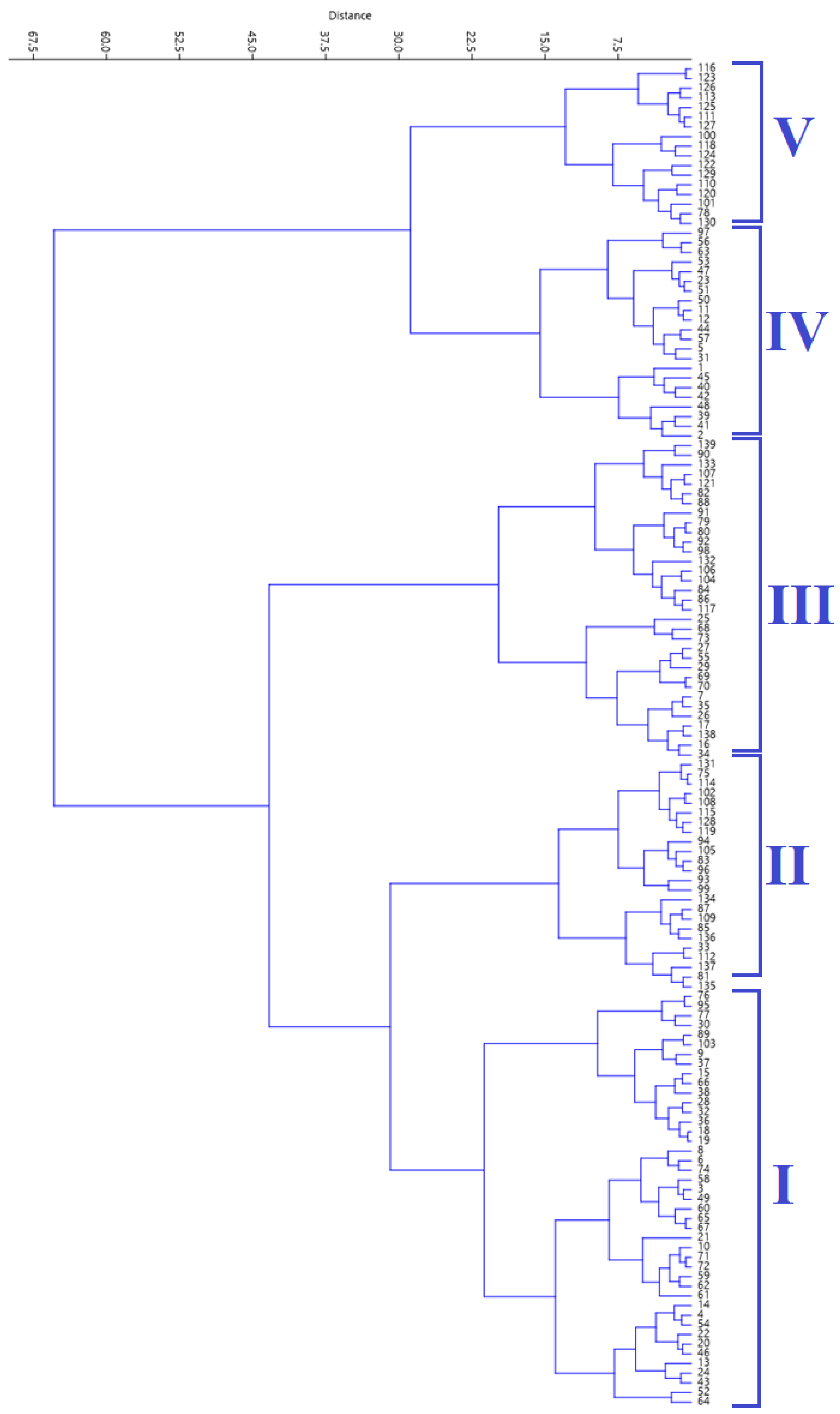
2.4. Hierarchical Clustering Analysis
3. Discussion
3.1. Nutritional Analysis
3.1.1. Total Protein Content
3.1.2. Total Starch Content
3.1.3. Total Soluble Sugar Content
3.1.4. Total Phenol Content
3.1.5. Total Phytic Acid Content
3.2. Multivariate Analysis and Identification of Promising Germplasms
4. Materials and Methods
4.1. Plant Material
4.2. Methodology
4.2.1. Estimation of Total Proteins
4.2.2. Estimation of Total Soluble Sugars, Starch, Phenols, and Phytic Acid
4.2.3. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Poos, M.I.; Costello, R.; Carlson-Newberry, S.J. The Role of Protein and Amino Acids in Sustaining and Enhancing Performance. In Committee on Military Nutrition Research: Activity Report December 1, 1994 through May 31, 1999; National Academies Press: Washington, DC, USA, 1999; pp. 49–52. Available online: https://www.ncbi.nlm.nih.gov/books/NBK224683/ (accessed on 16 August 2023).
- Richter, C.K.; Skulas-Ray, A.C.; Champagne, C.M.; Kris-Etherton, P.M. Plant protein and animal proteins: Do they differentially affect cardiovascular disease risk? Adv. Nutr. 2015, 6, 712–728. [Google Scholar] [CrossRef] [PubMed]
- Naghshi, S.; Sadeghi, O.; Willett, W.C.; Esmaillzadeh, A. Dietary intake of total, animal, and plant proteins and risk of all cause, cardiovascular, and cancer mortality: Systematic review and dose-response meta-analysis of prospective cohort studies. BMJ 2020, 370, m2412. [Google Scholar] [CrossRef] [PubMed]
- FAOSTAT Statistical Database 2021. Food and Agriculture Organization of the United Nations, Rome. Available online: https://www.fao.org/faostat/en/#data/QCL (accessed on 27 September 2023).
- Government of India. Agricultural Statistics at a Glance 2022. Directorate of Economics & Statistics, Department of Agriculture & Farmers Welfare, Ministry of Agriculture & Farmers Welfare, Government of India. Available online: https://agricoop.nic.in/Documents/CWWGDATA/Agricultural_Statistics_at_a_Glance_2022_0.pdf (accessed on 10 June 2023).
- Fuller, D.Q.; Murphy, C. The origins and early dispersal of horsegram (Macrotyloma uniflorum), a major crop of ancient India. Genet. Resour. Crop Evol. 2018, 65, 285–305. [Google Scholar] [CrossRef]
- Bravo, L.; Siddhuraju, P.; Saura-Calixto, F. Composition of underexploited Indian pulse; Comparison with common legumes. Food Chem. 1999, 64, 185–192. [Google Scholar] [CrossRef]
- Dwivedi, M.; Vasantha, K.Y.; Sreerama, Y.N.; Haware, D.J.; Singh, R.P.; Sattur, A.P. Kaulath, a new fungal fermented food from horse gram. J. Food Sci. Technol. 2015, 52, 8371–8376. [Google Scholar] [CrossRef]
- Ghumman, A.; Kaur, A.; Singh, N.; Singh, B. Effect of feed moisture and extrusion temperature on protein digestibility and extrusion behaviour of lentil and horsegram. Food Sci. Technol 2016, 70, 349–357. [Google Scholar] [CrossRef]
- Lalitha, N.; Singh, S.A. Preparation of horsegram protein concentrate with improved protein quality, in vitro digestibility and available lysine. J. Food Sci. Technol. 2020, 57, 2554–2560. [Google Scholar] [CrossRef]
- Longvah, T.; Ananthan, I.; Bhaskarachary, K.; Venkaiah, K. Indian Food Composition Tables; National Institute of Nutrition, Indian Council of Medical Research: Hyderabad, India, 2017; pp. 2–58. [Google Scholar]
- Sreerama, Y.N.; Sashikala, V.B.; Pratape, V.M.; Singh, V. Nutrients and antinutrients in cowpea and horse gram flours in comparison to chickpea flour: Evaluation of their flour functionality. Food Chem. 2012, 131, 462–468. [Google Scholar] [CrossRef]
- Reddy, A.M.; Kumar, S.G.; Jyothsnakumari, G.; Thimmanaik, S.; Sudhakar, C. Lead induced changes in antioxidant metabolism of horsegram (Macrotyloma uniflorum (Lam.) Verdc.) and bengalgram(Cicer arietinum L.). Chemosphere 2005, 60, 97–104. [Google Scholar] [CrossRef]
- All India Coordinated Research Project for Dryland Agriculture; Annual Report 2017–2018; ICAR—Central Research Institute for Dryland Agriculture, Indian Council of Agricultural Research: Hyderabad, India, 2018; p. 304.
- Hoover, R.; Zhou, Y. In vitro and in vivo hydrolysis of legume starches by an amylase and resistant starch formation in legumes-A review. Carbohydr. Polym. 2003, 54, 401–417. [Google Scholar] [CrossRef]
- Ofuya, Z.M.; Akhidue, V. The role of pulses in human nutrition: A review. J. Appl. Sci. Environ. Manag. 2005, 9, 99–104. [Google Scholar] [CrossRef]
- Burešová, I.; Sedláčková, I.; Faměra, O.; Lipavský, J. Effect of growing conditions on starch and protein content in triticale grain and amylose content in starch. Plant Soil Environ. 2010, 56, 99–104. [Google Scholar] [CrossRef]
- Padhi, S.R.; Bartwal, A.; John, R.; Tripathi, K.; Gupta, K.; Wankhede, D.P.; Mishra, G.P.; Kumar, S.; Archak, S.; Bhardwaj, R. Evaluation and multivariate analysis of cowpea [Vigna unguiculata (L.) walp] germplasm for selected nutrients—Mining for nutri-dense accessions. Front. Sustain. Food Syst. 2022, 6, 182–193. [Google Scholar]
- Bachmann, M.; Keller, F. Metabolism of the raffinose family oligosaccharides in leaves of Ajuga reptans L. (inter-and intracellular compartmentation). Plant Physiol. 1995, 109, 991–998. [Google Scholar] [CrossRef] [PubMed]
- Elango, D.; Rajendran, K.; Van der Laan, L.; Sebastiar, S.; Raigne, J.; Thaiparambil, N.A.; El Haddad, N.; Raja, B.; Wang, W.; Ferela, A.; et al. Raffinose family oligosaccharides: Friend or foe for human and plant health? Front. Plant Sci. 2022, 13, 829118. [Google Scholar] [CrossRef] [PubMed]
- Salvi, P.; Varshney, V.; Majee, M. Raffinose family oligosaccharides (RFOs): Role in seed vigor and longevity. Biosci. Rep. 2022, 42, BSR20220198. [Google Scholar] [CrossRef]
- Amarowicz, R.; Shahidi, F. Antioxidant activity of broad bean seed extract and its phenolic composition. J. Funct. Foods 2017, 38, 656–662. [Google Scholar] [CrossRef]
- Rahman, M.M.; Rahaman, M.S.; Islam, M.R.; Rahman, F.; Mithi, F.M.; Alqahtani, T.; Almikhlafi, M.A.; Alghamdi, S.Q.; Alruwaili, A.S.; Hossain, M.S.; et al. Role of Phenolic Compounds in Human Disease: Current Knowledge and Future Prospects. Molecules 2021, 27, 233. [Google Scholar] [CrossRef]
- Okafor, J.N.; Rautenbauch, F.; Meyer, M.; Le Roes-Hill, M.; Harris, T.; Jideani, V.A. Phenolic content, antioxidant, cytotoxic and antiproliferative effects of fractions of Vigna subterraenea (L.) verdc from Mpumalanga, South Africa. Heliyon 2021, 7, e08397. [Google Scholar] [CrossRef]
- Bhartiya, A.; Aditya, J.P.; Pal, R.S.; Bajeli, J. Agromorphological, nutritional and antioxidant properties in horsegram [Macrotyloma uniflorum (Lam.) Verdc.] germplasm collection from diverse altitudinal range of north western himalayan hills of India. Vegetos 2017, 30, 4473. [Google Scholar] [CrossRef]
- Stanisavljević, N.S.; Ilić, M.D.; Jovanović, Ž.S.; Čupić, T.; Dabić-Zagorac, D.; Natić, M.; TešićZ, L.; Radovic, S.S. Identification of seed coat phenolic compounds from differently colored pea varieties and characterization of their antioxidant activity. Arch. Biol. Sci. 2015, 67, 829–840. [Google Scholar] [CrossRef]
- Rodríguez Madrera, R.; Campa Negrillo, A.; Suárez Valles, B.; Ferreira Fernández, J.J. Phenolic content and antioxidant activity in seeds of common bean (Phaseolus vulgaris L.). Foods 2021, 10, 864. [Google Scholar] [CrossRef] [PubMed]
- Gupta, R.K.; Gangoliya, S.S.; Singh, N.K. Reduction of phytic acid and enhancement of bioavailable micronutrients in food grains. J. Food Sci. Technol. 2015, 52, 676–684. [Google Scholar] [CrossRef] [PubMed]
- Perera, D.; Devkota, L.; Garnier, G.; Panozzo, J.; Dhital, S. Hard-to-cook phenomenon in common legumes: Chemistry, mechanisms and utilisation. Food Chem. 2023, 415, 135743. [Google Scholar] [CrossRef] [PubMed]
- Feizollahi, E.; Mirmahdi, R.S.; Zoghi, A.; Zijlstra, R.T.; Roopesh, M.S.; Vasanthan, T. Review of the beneficial and anti-nutritional qualities of phytic acid, and procedures for removing it from food products. Food Res. Int. 2021, 143, 110284. [Google Scholar] [CrossRef]
- Sharma, H.; Singh, S.; Shamshad, M.; Padhy, A.K.; Kaur, R.; Kashyap, L.; Srivastava, P.; Mavi, G.S.; Kaur, S.; Sharma, A.; et al. Variability in iron, zinc, phytic acid and protein content in pre-breeding wheat germplasm under different water regimes. Plant Growth Regul. 2022, 100, 531–543. [Google Scholar] [CrossRef]
- Bhagyawant, S.S.; Gautam, A.K.; Narvekar, D.T.; Gupta, N.; Bhadkaria, A.; Srivastava, N.; Upadhyaya, H.D. Biochemical diversity evaluation in chickpea accessions employing mini-core collection. Physiol. Mol. Biol. Plants 2018, 24, 1165–1183. [Google Scholar] [CrossRef]
- Singh, J.; Kanaujia, R.; Srivastava, A.K.; Dixit, G.P.; Singh, N.P. Genetic variability for iron and zinc as well as antinutrients affecting bioavailability in black gram (Vigna mungo (L.) Hepper). J. Food Sci. Technol. 2017, 54, 1035–1042. [Google Scholar] [CrossRef]
- Duhan, A.; Khetarpaul, N.; Bishnoi, S. Changes in phytates and HCl extractability of calcium, phosphorus, and iron of soaked, dehulled, cooked, and sprouted pigeon pea cultivar (UPAS-120). Plant Foods Hum. Nutr. 2002, 57, 275–284. [Google Scholar] [CrossRef]
- Thavarajah, P.; See, C.T.; Vandenberg, A. Phytic acid and Fe and Zn concentration in lentil (Lens culinaris L.) seeds is influenced by temperature during seed filling period. Food Chem. 2010, 122, 254–259. [Google Scholar] [CrossRef]
- Dhole, V.J.; Reddy, K.S. Genetic variation for phytic acid content in mungbean (Vigna radiata L. Wilczek). Crop J. 2015, 3, 157–162. [Google Scholar] [CrossRef]
- Saba, I.; Sofi, P.A.; Zeerak, N.A.; Bhat, M.A.; Mir, R.R. Characterisation of a core set of common bean (Phaseolus vulgaris L.) germplasm for seed quality traits. SABRAO J. Breed. Genet. 2016, 48, 359–376. [Google Scholar]
- Vishakha, S.; Vibha, B. Effects of processing on phytic acid, iron and its bioavailability of Macrotyloma uniflorum. Food Sci. Res. J. 2017, 8, 128–131. [Google Scholar]
- Koo, S.; Jeon, M.; Lee, Y.; Kim, H.; Kang, B.; Go, J.; Baek, I.; Yun, H.; Choi, M. Screening of soybean germplasm with high starch content. Korean J. Breed. Sci. 2014, 46, 52–57. [Google Scholar] [CrossRef]
- Dhungana, S.K.; Kulkarni, K.P.; Kim, M.; Ha, B.K.; Kang, S.; Song, J.T.; Shin, D.H.; Lee, J.D. Environmental stability and correlation of soybean seed starch with protein and oil contents. Plant Breed. Biotechnol. 2017, 5, 293–303. [Google Scholar] [CrossRef]
- Shen, S.; Hou, H.; Ding, C.; Bing, D.J.; Lu, Z.X. Protein content correlates with starch morphology, composition and physicochemical properties in field peas. Can. J. Plant Sci. 2016, 96, 404–412. [Google Scholar] [CrossRef]
- Rolletschek, H.; Nguyen, T.H.; Häusler, R.E.; Rutten, T.; Göbel, C.; Feussner, I.; Radchuk, R.; Tewes, A.; Claus, B.; Klukas, C.; et al. Antisense inhibition of the plastidial glucose-6-phosphate/phosphate translocator in Vicia seeds shifts cellular differentiation and promotes protein storage. Plant J. 2007, 51, 468–484. [Google Scholar] [CrossRef]
- Zeng, Z.; Li, Y.; Yang, R.; Liu, C.; Hu, X.; Luo, S.; Gong, E.; Ye, J. The relationship between reducing sugars and phenolic retention of brown rice after enzymatic extrusion. J. Cereal Sci. 2017, 74, 244–249. [Google Scholar] [CrossRef]
- Horwitz, W. Official Methods of Analysis of AOAC International. Volume I, Agricultural Chemicals, Contaminants; Horwitz, W., Ed.; AOAC International: Gaithersburg, MD, USA, 2010; p. 1997. [Google Scholar]
- DuBois, M.; Gilles, K.A.; Hamilton, J.K.; Rebers, P.T.; Smith, F. Colorimetric method for determination of sugars and related substances. Anal. Chem. 1956, 28, 350–356. [Google Scholar] [CrossRef]
- Singleton, V.L.; Orthofer, R.; Lamuela-Raventós, R.M. Analysis of total phenols and other oxidation substrates and antioxidants by means of folin-ciocalteu reagent. Meth. Enzymol. 1999, 299, 152–178. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2023; Available online: https://www.R-project.org/ (accessed on 1 August 2023).

{kind=link}
{kind=link}
| Estimate of Variability | Protein (g/100 g) | Starch (g/100 g) | Total Soluble Sugars (g/100 g) | Phytic Acid (mg/g) | Phenols (mg GAE/g) |
|---|---|---|---|---|---|
| Median | 23.8 | 29.9 | 5.47 | 10.2 | 7.01 |
| Mean | 23.7 | 29.8 | 5.61 | 10.2 | 7.00 |
| Standard deviation | 0.99 | 1.36 | 2.50 | 4.36 | 1.71 |
| Coefficient of variation % | 4.16 | 4.56 | 44.6 | 42.9 | 24.5 |
| Minimum value | 21.8 | 26.2 | 0.86 | 1.07 | 3.38 |
| Accession with minimum value | IC139429 | IC0369691 | IC022772 | BSP 15-1 | IC023482 |
| Maximum value | 26.7 | 33.00 | 12.10 | 21.2 | 11.3 |
| Accession with maximum value | IC023482 | IC426463 | IC265920 | IC071733 | IC278825 |
| IFCT 2017 | 21.7 | 47.9 | 1.99 * | 3.39 | 3.32 |
| Trait | Protein | Starch | Sugars | Phytic Acid | Phenols |
|---|---|---|---|---|---|
| Protein | 1.00 | ||||
| Starch | −0.61 ** | 1.00 | |||
| Sugars | −0.33 ** | 0.02 | 1.00 | ||
| Phytic Acid | 0.14 | 0.07 | −0.05 | 1.00 | |
| Phenols | −0.26 ** | −0.08 | 0.70 ** | −0.10 | 1.00 |
| Properties | HCA Cluster Mean | Overall Mean | ||||
|---|---|---|---|---|---|---|
| I | II | III | IV | V | ||
| Number of accessions | 43 | 24 | 33 | 22 | 17 | |
| Protein (g/100 g) | 23.4 | 24.0 | 23.7 | 23.3 | 24.7 | 23.7 |
| Starch (g/100 g) | 29.9 | 29.6 | 29.8 | 30.5 | 29.3 | 29.8 |
| Sugar (g/100 g) | 7.15 | 3.50 | 4.96 | 8.02 | 2.9 | 5.61 |
| Phytic acid (mg/g) | 9.36 | 10.1 | 4.86 | 14.8 | 16.5 | 10.2 |
| Phenol mg (GAE/g) | 8.17 | 5.65 | 6.65 | 8.32 | 4.91 | 7.00 |
| SN | Trait/s | Accessions Identified as Superior |
|---|---|---|
| 1 | Low phytic acid (mg/g) | BSP 15-1 (1.07), IC262106 (1.66), IC281653 (1.88), IC019431 (2.83), IC281820 (3.41) |
| 2 | Low phenols (mg GAE/g) | IC023482 (3.38), IC022781 (3.76), IC022795 (3.81), IC105564 (3.85), IC139512 (3.93) |
| 3 | High sugars (g/100 g) | IC265920 (12.1), IC273743 (10.7), IC397511 (10.2), IC262106 (9.64), BSP 15-1 (9.53) |
| 4 | High protein (g/100 g) | IC023482 (26.70), IC256885 (26.2), IC044021 (26.1), IC139512 (26), IC022795 (25.8) |
| 5 | High starch (g/100 g) | IC426463 (33.00), IC273748 (33.0), IC343814 (32.6), IC469766 (32.4), IC418359 (32.2) |
| 6 | Moderate protein, low phenols, low sugars, high phytic acid | IC022781 (23.4 g/100 g, 3.76 mg GAE/g, 1.60 g/100 g, 10.1 mg/g) |
| 7 | High protein, low phenols, low sugars, high phytic acid | IC023482 (26.70 g/100 g, 3.38 mg GAE/g, 1.31 g/100 g, 14.2 mg/g) |
| 8 | High protein, low phenols, low sugars, moderate phytic acid | IC022795 (25.8 g/100 g, 3.81 mg GAE/g, 2.46 g/100 g, 5.21 mg/g) |
| 9 | High protein, low phenols, low sugars, low phytic acid | IC139512 (26.0 g/100 g, 3.93 mg GAE/g, 3.60 g/100 g, 3.93 mg/g) |
| IC019431 (24.2 g/100 g, 4.98 mg GAE/g, 3.64 g/100 g, 2.83 mg/g |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kumari, M.; Padhi, S.R.; Chourey, S.K.; Kondal, V.; Thakare, S.S.; Negi, A.; Gupta, V.; Arya, M.; Yasin, J.K.; Singh, R.; et al. Unveiling Diversity for Quality Traits in the Indian Landraces of Horsegram [Macrotyloma uniflorum (Lam.) Verdc.]. Plants 2023, 12, 3803. https://doi.org/10.3390/plants12223803
Kumari M, Padhi SR, Chourey SK, Kondal V, Thakare SS, Negi A, Gupta V, Arya M, Yasin JK, Singh R, et al. Unveiling Diversity for Quality Traits in the Indian Landraces of Horsegram [Macrotyloma uniflorum (Lam.) Verdc.]. Plants. 2023; 12(22):3803. https://doi.org/10.3390/plants12223803
Chicago/Turabian StyleKumari, Manju, Siddhant Ranjan Padhi, Sushil Kumar Chourey, Vishal Kondal, Swapnil S. Thakare, Ankita Negi, Veena Gupta, Mamta Arya, Jeshima Khan Yasin, Rakesh Singh, and et al. 2023. "Unveiling Diversity for Quality Traits in the Indian Landraces of Horsegram [Macrotyloma uniflorum (Lam.) Verdc.]" Plants 12, no. 22: 3803. https://doi.org/10.3390/plants12223803
APA StyleKumari, M., Padhi, S. R., Chourey, S. K., Kondal, V., Thakare, S. S., Negi, A., Gupta, V., Arya, M., Yasin, J. K., Singh, R., Bharadwaj, C., Kumar, A., Bhatt, K. C., Bhardwaj, R., Rana, J. C., Joshi, T., & Riar, A. (2023). Unveiling Diversity for Quality Traits in the Indian Landraces of Horsegram [Macrotyloma uniflorum (Lam.) Verdc.]. Plants, 12(22), 3803. https://doi.org/10.3390/plants12223803