
Starch Properties and Morphology of Eight Floury Endosperm Mutants in Rice



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
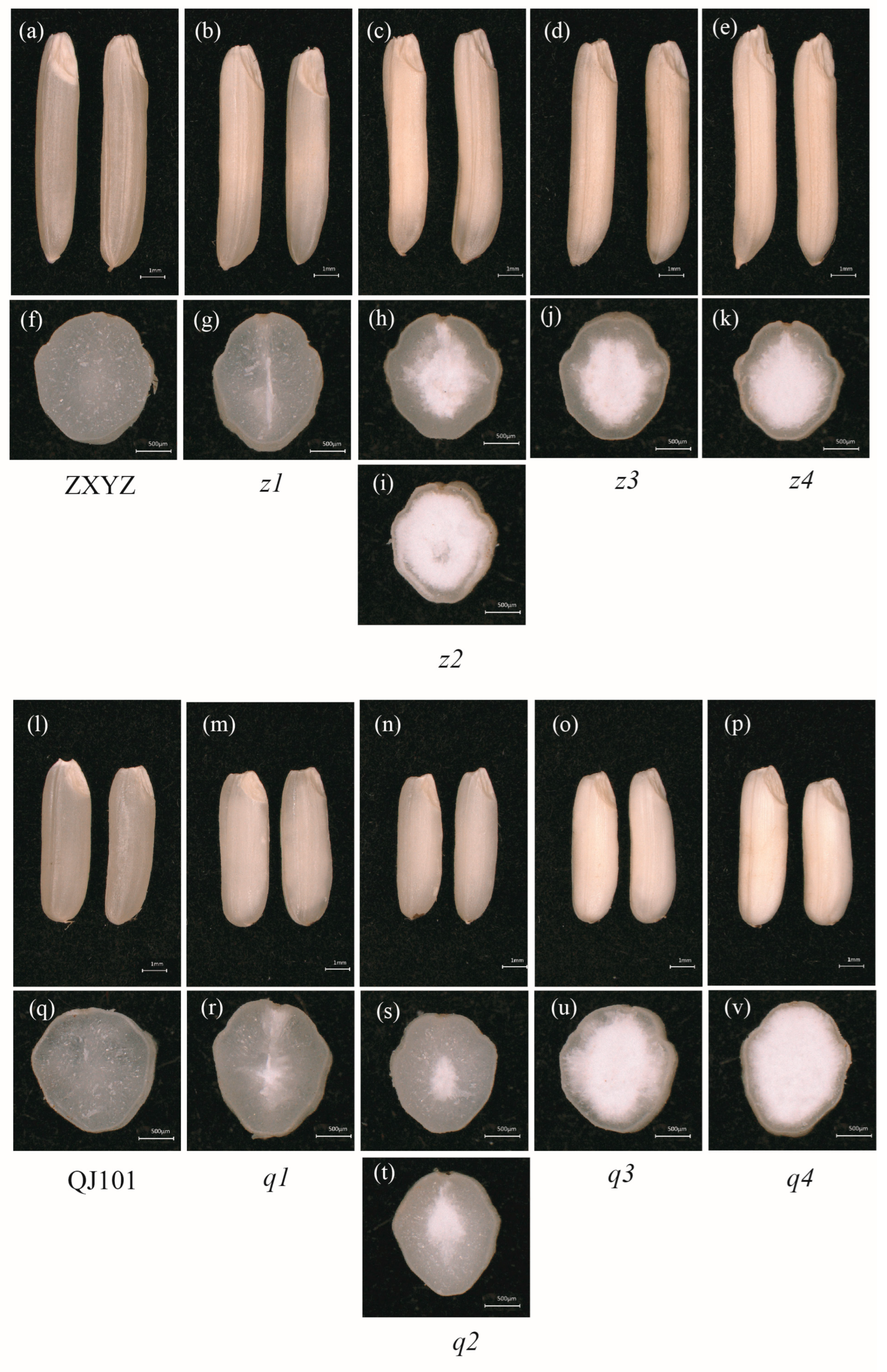
2.1. Mutants Exhibited Varying Degrees of Floury Endosperm
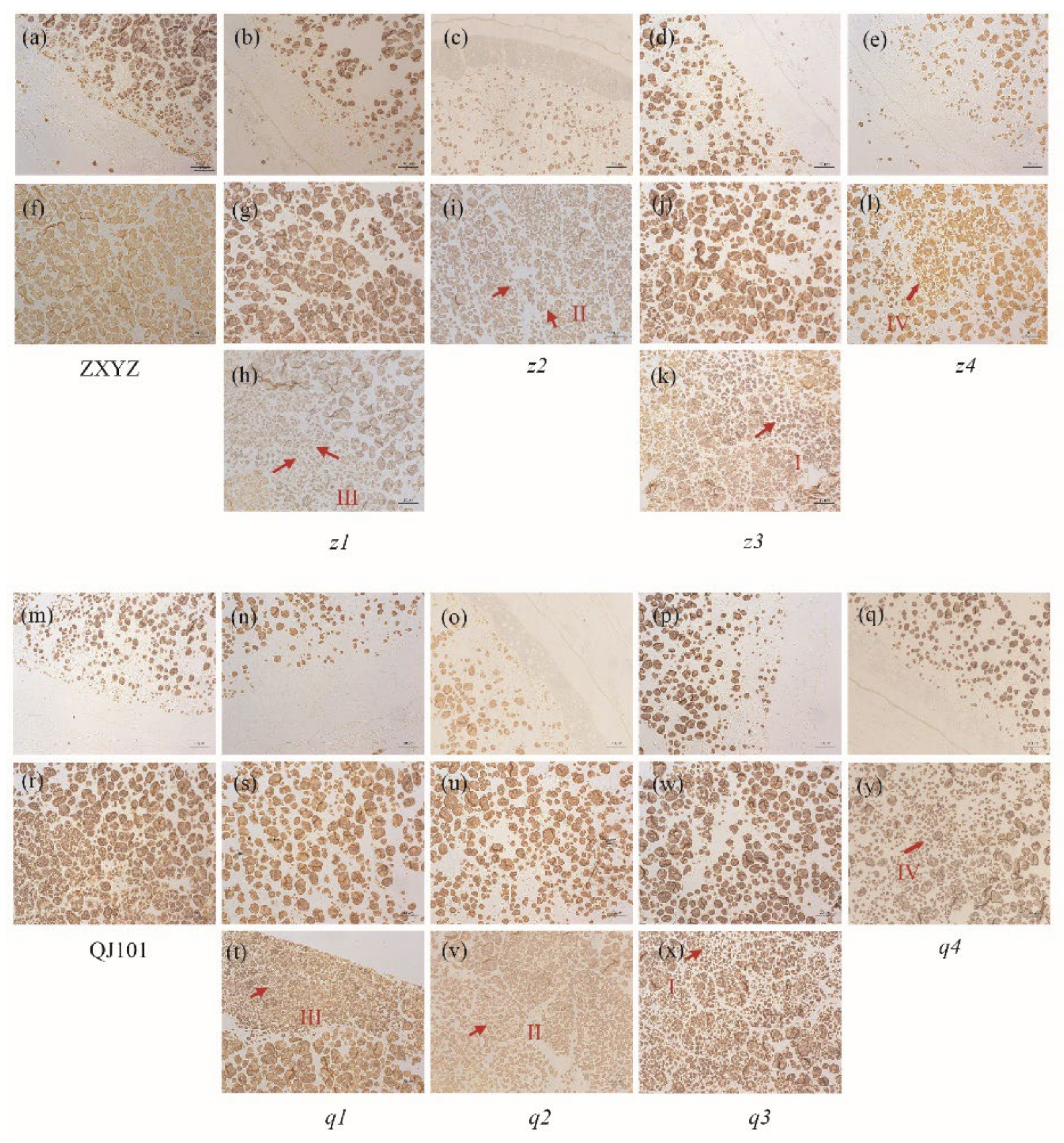
2.2. Mutants Show Abnormal Development of Starch Granules and Amyloplasts
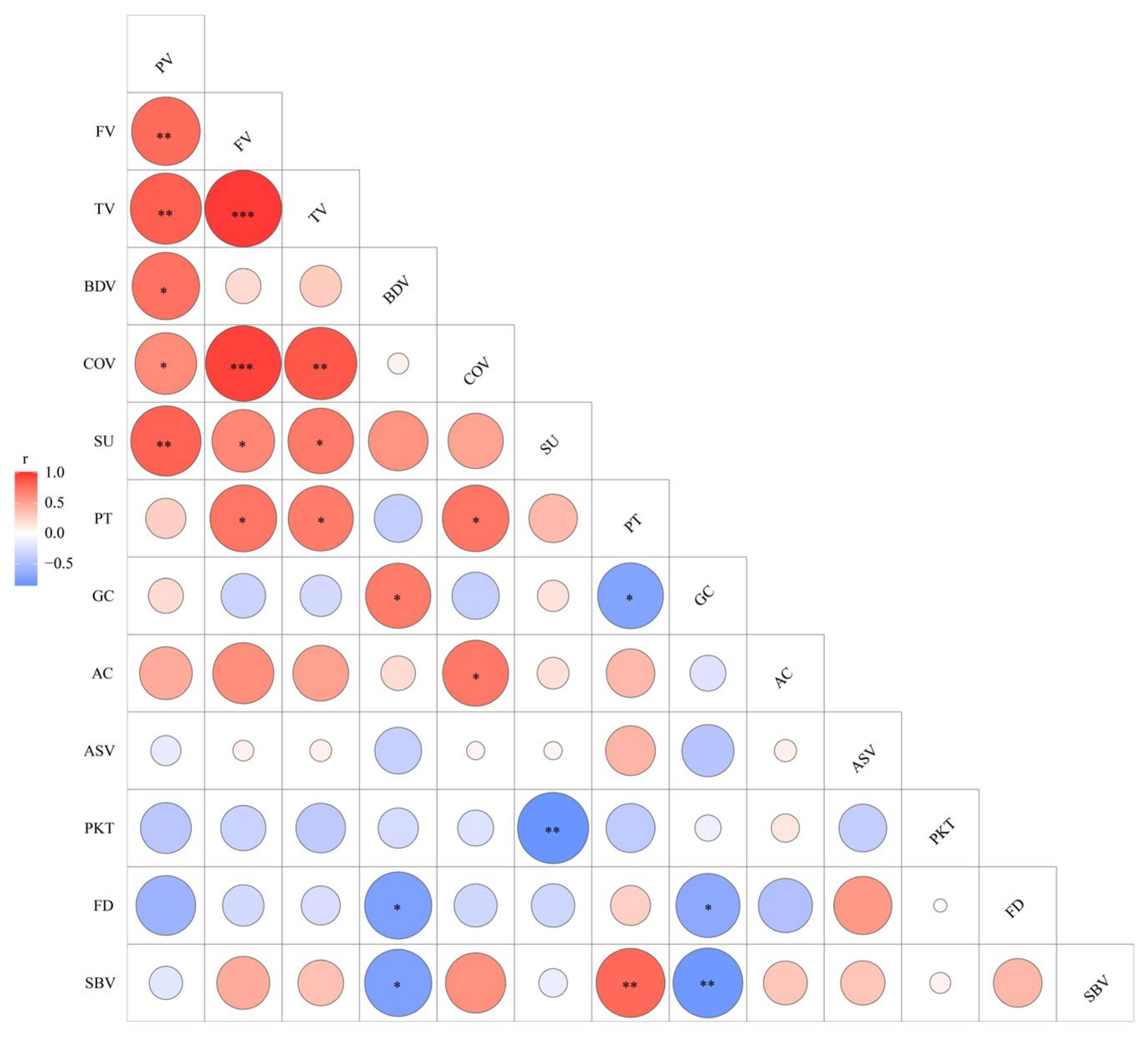
2.3. Mutants Showed Significant Changes of CEQs
2.4. Starch Swelling in Urea (SU) Solution Dramatically Changed in z2 and z4
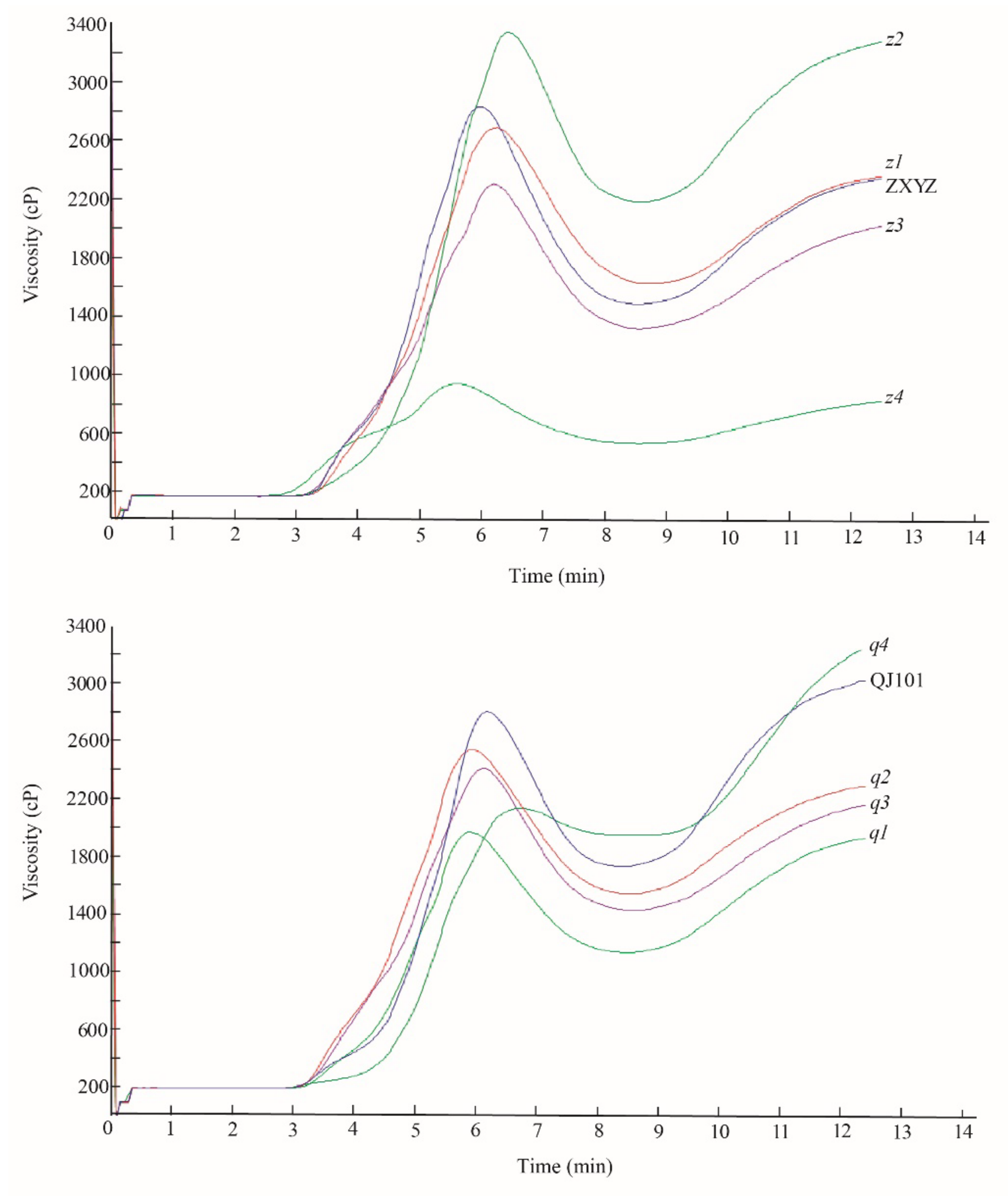
2.5. Paste viscosity of Milled Rice Flour Dramatically Changed in z2 and z4
3. Discussion
3.1. Eight Floury Endosperm Mutants Exhibited Wide Variation in Starch Properties
3.2. The Morphology of Starch Granules Affect CEQs
3.3. Certain Genes but May Not PUL Affect CEQs in These Floury Mutants
4. Materials and Methods
4.1. Plant Materials
4.2. Microscopy
4.3. Determination of RVA Profiles
4.4. Determination of CEQ
4.5. Determination of Starch Swelling in Urea Solution
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Zhao, D.S.; Zhang, C.Q.; Li, Q.F.; Liu, Q.Q. Genetic control of grain appearance quality in rice. Biotechnol. Adv. 2022, 60, 108014. [Google Scholar] [CrossRef]
- Siebenmorgen, T.J.; Grigg, B.C.; Lanning, S.B. Impacts of preharvest factors during kernel development on rice quality and functionality. Annu. Rev. Food Sci. Technol. 2013, 4, 101–115. [Google Scholar] [CrossRef]
- Li, Y.B.; Fan, C.C.; Xing, Y.Z.; Yun, P.; Luo, L.J.; Yan, B.; Peng, B.; Xie, W.B.; Wang, G.W.; Li, X.H.; et al. Chalk5 encodes a vacuolar H+-translocating pyrophosphatase influencing grain chalkiness in rice. Nat. Genet. 2014, 46, 398–404. [Google Scholar] [CrossRef]
- Wu, B.; Yun, P.; Zhou, H.; Xia, D.; Gu, Y.; Li, P.B.; Yao, J.L.; Zhou, Z.Q.; Chen, J.X.; Liu, R.J.; et al. Natural variation in WHITE-CORE RATE 1 regulates redox homeostasis in rice endosperm to affect grain quality. Plant Cell 2022, 34, 1912–1932. [Google Scholar] [CrossRef]
- Tuncel, A.; Kawaguchi, J.; Ihara, Y.; Matsusaka, H.; Nishi, A.; Nakamura, T.; Kuhara, S.; Hirakawa, H.; Nakamura, Y.; Cakir, B.; et al. The rice endosperm ADP-glucose pyrophosphorylase large subunit is essential for optimal catalysis and allosteric regulation of the heterotetrameric enzyme. Plant Cell Physiol. 2014, 55, 1169–1183. [Google Scholar] [CrossRef]
- Xu, Y.; Lin, Q.P.; Li, X.F.; Wang, F.Q.; Chen, Z.H.; Wang, J.; Li, W.Q.; Fan, F.J.; Tao, Y.J.; Jiang, Y.J.; et al. Fine-tuning the amylose content of rice by precise base editing of the Wx gene. Plant Biotechnol. J. 2021, 19, 11–13. [Google Scholar] [CrossRef]
- Matsushima, R.; Maekawa, M.; Kusano, M.; Kondo, H.; Fujita, N.; Kawagoe, Y.; Sakamoto, W. Amyloplast-localized SUBSTANDARD STARCH GRAIN4 protein influences the size of starch grains in rice endosperm. Plant Physiol. 2014, 164, 623–636. [Google Scholar] [CrossRef] [PubMed]
- Matsushima, R.; Maekawa, M.; Kusano, M.; Tomita, K.; Kondo, H.; Nishimura, H.; Crofts, N.; Fujita, N.; Sakamoto, W. Amyloplast membrane protein SUBSTANDARD STARCH GRAIN6 controls starch grain size in rice endosperm. Plant Physiol. 2016, 170, 1445–1459. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.H.; Liu, F.; Ren, Y.L.; Wang, Y.L.; Liu, X.; Long, W.H.; Wang, D.; Zhu, J.P.; Zhu, X.P.; Jing, R.N.; et al. GOLGI TRANSPORT 1B regulates protein export from the endoplasmic reticulum in rice endosperm cells. Plant Cell 2016, 28, 2850–2865. [Google Scholar] [CrossRef] [PubMed]
- Zhu, J.P.; Ren, Y.L.; Zhang, Y.Y.; Yang, J.; Duan, E.C.; Wang, Y.L.; Liu, F.; Wu, M.M.; Pan, T.; Wang, Y.F.; et al. Subunit E isoform 1 of vacuolar H+-ATPase OsVHA enables post-Golgi trafficking of rice seed storage proteins. Plant Physiol. 2021, 187, 2192–2208. [Google Scholar] [CrossRef]
- Hao, Y.Y.; Wang, Y.L.; Wu, M.M.; Zhu, X.P.; Teng, X.; Sun, Y.L.; Zhu, J.P.; Zhang, Y.Y.; Jing, R.N.; Lei, J.; et al. The nuclear-localized PPR protein OsNPPR1 is important for mitochondrial function and endosperm development in rice. J. Exp. Bot. 2019, 70, 4705–4720. [Google Scholar] [CrossRef] [PubMed]
- Wu, M.M.; Ren, Y.L.; Cai, M.H.; Wang, Y.L.; Zhu, S.S.; Zhu, J.P.; Hao, Y.Y.; Teng, X.; Zhu, X.P.; Jing, R.N.; et al. Rice FLOURY ENDOSPERM10 encodes a pentatricopeptide repeat protein that is essential for the trans-splicing of mitochondrial nad1 intron 1 and endosperm development. New Phytol. 2019, 223, 736–750. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.; Cai, X.L.; Wang, Z.Y.; Hong, M.M. An interaction between a MYC protein and an EREBP protein is involved in transcriptional regulation of the rice Wx gene. J. Biol. Chem. 2003, 278, 47803–47811. [Google Scholar] [CrossRef] [PubMed]
- Chung, T.; Kim, C.S.; Nguyen, H.N.; Meeley, R.B.; Larkins, B.A. The maize zmsmu2 gene encodes a putative RNA-splicing factor that affects protein synthesis and RNA processing during endosperm development. Plant Physiol. 2007, 144, 821–835. [Google Scholar] [CrossRef]
- Morita, S.; Wada, H.; Matsue, Y. Countermeasures for heat damage in rice grain quality under climate change. Plant Prod. Sci. 2013, 19, 1–11. [Google Scholar] [CrossRef]
- Wang, L.Q.; Liu, W.J.; Xu, Y.; He, Y.Q.; Luo, L.J.; Xing, Y.Z.; Xu, C.G.; Zhang, Q.F. Genetic basis of 17 traits and viscosity parameters characterizing the eating and cooking quality of rice grain. Theor. Appl. Genet. 2007, 115, 463–476. [Google Scholar] [CrossRef]
- Perez, C.M.; Juliano, B.O. Indicators of eating quality for non-waxy rices. Food Chem. 1979, 4, 185–195. [Google Scholar] [CrossRef]
- Wang, Z.Y.; Zheng, F.Q.; Shen, G.Z.; Gao, J.P.; Snustad, D.P.; Li, M.G.; Zhang, J.L.; Hong, M. The amylose content in rice endosperm is related to the post-transcriptional regulation of the waxy gene. Plant J. 1995, 7, 613–622. [Google Scholar] [CrossRef]
- Khush, G.S.; Paule, C.M.; De La Cruz, N.M. Rice grain quality evaluation and improvement at IRRI. In Proceedings of the Workshop on Chemical Aspects of Rice Grain Quality, IRRI, Hanoi, Vietnam, 23–25 October 1978. [Google Scholar]
- Fitzgerald, M.A.; McCouch, S.R.; Hall, R.D. Not just a grain of rice: The quest for quality. Trends Plant Sci. 2009, 14, 133–139. [Google Scholar] [CrossRef]
- Bao, J.S.; Corke, H.; Sun, M. Nucleotide diversity in starch synthase IIa and validation of single nucleotide polymorphisms in relation to starch gelatinization temperature and other physicochemical properties in rice (Oryza sativa L.). Theor. Appl. Genet. 2006, 113, 1171–1183. [Google Scholar] [CrossRef]
- Fan, C.C.; Yu, X.Q.; Xing, Y.Z.; Xu, C.G.; Luo, L.J.; Zhang, Q.T. The main effects, epistatic effects and environmental interactions of QTLs on the cooking and eating quality of rice in a doubled-haploid line population. Theor. Appl. Genet. 2005, 110, 1445–1452. [Google Scholar] [CrossRef]
- Zhang, A.P.; Gao, Y.; Li, Y.Y.; Ruan, B.P.; Yang, S.L.; Liu, C.L.; Zhang, B.; Jiang, H.Z.; Fang, G.N.; Ding, S.L.; et al. Genetic analysis for cooking and eating quality of super rice and fine mapping of a novel locus qgc10 for gel consistency. Front Plant Sci. 2020, 11, 342. [Google Scholar] [CrossRef] [PubMed]
- Su, Y.; Rao, Y.C.; Hu, S.K.; Yang, Y.L.; Gao, Z.Y.; Zhang, G.H.; Liu, J.; Hu, J.; Yan, M.X.; Dong, G.J.; et al. Map-based cloning proves qGC-6, a major QTL for gel consistency of japonica/indica cross, responds by Waxy in rice (Oryza sativa L.). Theor. Appl. Genet. 2011, 123, 859–867. [Google Scholar] [CrossRef] [PubMed]
- Tran, N.A.; Daygon, V.D.; Resurreccion, A.P.; Cuevas, R.P.; Corpuz, H.M.; Fitzgerald, M.A. A single nucleotide polymorphism in the Waxy gene explains a significant component of gel consistency. Theor. Appl. Genet. 2011, 123, 519–525. [Google Scholar] [CrossRef] [PubMed]
- Gao, Z.Y.; Zeng, D.L.; Cheng, F.M.; Tian, Z.X.; Guo, L.B.; Su, Y.; Yan, M.X.; Jiang, H.; Dong, G.J.; Huang, Y.C.; et al. ALK, the key gene for gelatinization temperature, is a modifier gene for gel consistency in rice. J. Integr. Plant Biol. 2011, 53, 756–765. [Google Scholar]
- Fitzgerald, M.A.; Martin, M.; Ward, R.M.; Park, W.D.; Shead, H.J. Viscosity of rice flour: A rheological and biological study. J. Agric. Food Chem. 2003, 51, 2295–2299. [Google Scholar] [CrossRef]
- Ohtsubo, K.; Nakamura, S. Evaluation of palatability of cooked rice. In Advances in International Rice Research, 6th ed.; Ohtsubo, K.I., Nakamura, S., Eds.; INTECH: Zagreb, Croatia, 2017; Volume 3, pp. 91–110. [Google Scholar]
- Buenafe, R.; Kumanduri, V.; Sreenivasulu, N. Deploying viscosity and starch polymer properties to predict cooking and eating quality models: A novel breeding tool to predict texture. Carbohydr. Polym. 2021, 260, 117766. [Google Scholar] [CrossRef]
- Pang, Y.L.; Ali, J.; Wang, X.Q.; Franje, N.J.; Revilleza, J.E.; Xu, J.L.; Li, Z.K. Relationship of rice grain amylose, gelatinization temperature and pasting properties for breeding better eating and cooking quality of rice varieties. PLoS ONE 2016, 11, e168483. [Google Scholar] [CrossRef]
- Yan, C.J.; Tian, Z.X.; Fang, Y.W.; Yang, Y.C.; Li, J.; Zeng, S.Y.; Gu, S.L.; Xu, C.W.; Tang, S.Z.; Gu, M.H. Genetic analysis of starch paste viscosity parameters in glutinous rice (Oryza sativa L.). Theor. Appl. Genet. 2011, 122, 63–76. [Google Scholar] [CrossRef]
- Tian, Z.X.; Qian, Q.; Liu, Q.Q.; Yan, M.X.; Liu, X.F.; Yan, C.J.; Liu, G.F.; Gao, Z.Y.; Tang, S.Z.; Zeng, D.L.; et al. Allelic diversities in rice starch biosynthesis lead to a diverse array of rice eating and cooking qualities. Proc. Natl. Acad. Sci. USA 2009, 106, 21760–21765. [Google Scholar] [CrossRef]
- Sui, J.M.; Li, X.; Yan, S.; Yan, C.J.; Zhang, R. Studies on the rice RVA profile characteristics and its correlation with the quality. Sci. Agric. Sin. 2005, 38, 657–663. [Google Scholar]
- Hsu, Y.C.; Tseng, M.C.; Wu, Y.P.; Ying, L.M.; Jin, W.F.; Kang, H.K.; Le, H.Y.; Rong, L.Y. Genetic factors responsible for eating and cooking qualities of rice grains in a recombinant inbred population of an inter-subspecific cross. Mol. Breed. 2014, 34, 655–673. [Google Scholar] [CrossRef] [PubMed]
- Chun, A.; Song, J.; Kim, K.; Lee, H. Quality of head and chalky rice and deterioration of eating quality by chalky rice. J. Crop Sci. Biotechnol. 2009, 12, 239–244. [Google Scholar] [CrossRef]
- Nishi, A.; Nakamura, Y.; Tanaka, N.; Satoh, H. Biochemical and genetic analysis of the effects of amylose-extender mutation in rice endosperm. Plant Physiol. 2001, 127, 459–472. [Google Scholar] [CrossRef]
- Youngjun, M.; Ji-Ung, J. The use of floury endosperm mutants to develop rice cultivars suitable for dry milling. Plant Biotechnol. Rep. 2020, 14, 185–191. [Google Scholar]
- Ryoo, N.; Yu, C.; Park, C.S.; Baik, M.Y.; Park, I.M.; Cho, M.H.; Bhoo, S.H.; An, G.; Hahn, T.R.; Jeon, J.S. Knockout of a starch synthase gene OsSSIIIa/Flo5 causes white-core floury endosperm in rice (Oryza sativa L.). Plant Cell Rep. 2007, 26, 1083–1095. [Google Scholar] [CrossRef]
- You, X.M.; Zhang, W.W.; Hu, J.L.; Jing, R.N.; Cai, Y.; Feng, Z.M.; Kong, F.; Zhang, J.; Yan, H.G.; Chen, W.W.; et al. FLOURY ENDOSPERM15 encodes a glyoxalase I involved in compound granule formation and starch synthesis in rice endosperm. Plant Cell Rep. 2019, 38, 345–359. [Google Scholar] [CrossRef]
- Lei, J.; Teng, X.; Wang, Y.F.; Jiang, X.K.; Zhao, H.H.; Zheng, X.M.; Ren, Y.L.; Dong, H.; Wang, Y.L.; Duan, R.C.; et al. Plastidic pyruvate dehydrogenase complex E1 component subunit Alpha1 is involved in galactolipid biosynthesis required for amyloplast development in rice. Plant Biotechnol. J. 2022, 20, 437–453. [Google Scholar] [CrossRef]
- Wang, E.T.; Wang, J.J.; Zhu, X.D.; Hao, W.; Wang, L.Y.; Li, Q.; Zhang, L.X.; He, W.; Lu, B.R.; Lin, H.X.; et al. Control of rice grain-filling and yield by a gene with a potential signature of domestication. Nat. Genet. 2008, 40, 1370–1374. [Google Scholar] [CrossRef]
- Liu, F.; Ren, Y.L.; Wang, Y.H.; Peng, C.; Zhou, K.N.; Lv, J.; Guo, X.P.; Zhang, X.; Zhong, M.S.; Zhao, S.L.; et al. OsVPS9A functions cooperatively with OsRAB5A to regulate post-Golgi dense vesicle-mediated storage protein trafficking to the protein storage vacuole in rice endosperm cells. Mol. Plant 2013, 6, 1918–1932. [Google Scholar] [CrossRef]
- Teng, X.; Zhong, M.S.; Zhu, X.P.; Wang, C.M.; Ren, Y.L.; Wang, Y.L.; Zhang, H.; Jiang, L.; Wang, D.; Hao, Y.Y.; et al. FLOURY ENDOSPERM16 encoding a NAD-dependent cytosolic malate dehydrogenase plays an important role in starch synthesis and seed development in rice. Plant Biotechnol. J. 2019, 17, 1914–1927. [Google Scholar] [CrossRef] [PubMed]
- Zhu, X.P.; Teng, X.; Wang, Y.L.; Hao, Y.Y.; Jing, R.N.; Wang, Y.F.; Liu, Y.; Zhu, J.P.; Wu, M.M.; Zhong, M.S.; et al. FLOURY ENDOSPERM11 encoding a plastid heat shock protein 70 is essential for amyloplast development in rice. Plant Sci. 2018, 277, 89–99. [Google Scholar] [CrossRef]
- Zhang, L.; Ren, Y.L.; Lu, B.Y.; Yang, C.Y.; Feng, Z.M.; Liu, Z.; Chen, J.; Ma, W.W.; Wang, Y.; Yu, X.W.; et al. FLOURY ENDOSPERM7 encodes a regulator of starch synthesis and amyloplast development essential for peripheral endosperm development in rice. J. Exp. Bot. 2016, 67, 633–647. [Google Scholar] [CrossRef] [PubMed]
- Matsushima, R.; Maekawa, M.; Fujita, N.; Sakamoto, W. A rapid, direct observation method to isolate mutants with defects in starch grain morphology in rice. Plant Cell Physiol. 2010, 51, 728–741. [Google Scholar] [CrossRef] [PubMed]
- Myers, A.M.; James, M.G.; Lin, Q.; Yi, G.; Stinard, P.S.; Hennen-Bierwagen, T.A.; Becraft, P.W. Maize opaque5 encodes monogalactosyldiacylglycerol synthase and specifically affects galactolipids necessary for amyloplast and chloroplast function. Plant Cell 2011, 23, 2331–2347. [Google Scholar] [CrossRef]
- Yun, M.S.; Kawagoe, Y. Septum formation in amyloplasts produces compound granules in the rice endosperm and is regulated by plastid division proteins. Plant Cell Physiol. 2010, 51, 1469–1479. [Google Scholar] [CrossRef]
- Kawagoe, Y. The characteristic polyhedral, sharp-edged shape of compound-type starch granules in rice endosperm is achieved via the septum-like structure of the amyloplast. J. Appl. Glycosci. 2013, 60, 29–36. [Google Scholar] [CrossRef]
- Toyosawa, Y.; Kawagoe, Y.; Matsushima, R.; Crofts, N.; Ogawa, M.; Fukuda, M.; Kumamaru, T.; Okazaki, Y.; Kusano, M.; Saito, K.; et al. Deficiency of Starch Synthase IIIa and IVb alters starch granule morphology from polyhedral to spherical in rice endosperm. Plant Physiol. 2016, 170, 1255–1270. [Google Scholar] [CrossRef]
- Jia, L.; Ding, X.Y.; Wang, P.R.; Ding, X.J. Rice RVA profile characteristics and correlation with the physical/chemical quality. Acta Agron. Sin. 2008, 34, 790–794. [Google Scholar] [CrossRef]
- Sun, M.M.; Abdula, S.E.; Lee, H.J.; Cho, Y.C.; Han, L.Z.; Koh, H.J.; Cho, Y.G. Molecular aspect of good eating quality formation in Japonica rice. PLoS ONE 2011, 6, e18385. [Google Scholar] [CrossRef]
- Peng, C.; Wang, Y.H.; Liu, F.; Ren, Y.L.; Zhou, K.N.; Lv, J.; Zheng, M.; Zhao, S.L.; Zhang, L.; Wang, C.; et al. FLOURY ENDOSPERM6 encodes a CBM48 domain-containing protein involved in compound granule formation and starch synthesis in rice endosperm. Plant J. 2014, 77, 917–930. [Google Scholar] [CrossRef] [PubMed]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hao, Y.; Huang, F.; Gao, Z.; Xu, J.; Zhu, Y.; Li, C. Starch Properties and Morphology of Eight Floury Endosperm Mutants in Rice. Plants 2023, 12, 3541. https://doi.org/10.3390/plants12203541
Hao Y, Huang F, Gao Z, Xu J, Zhu Y, Li C. Starch Properties and Morphology of Eight Floury Endosperm Mutants in Rice. Plants. 2023; 12(20):3541. https://doi.org/10.3390/plants12203541
Chicago/Turabian StyleHao, Yuanyuan, Fudeng Huang, Zhennan Gao, Junfeng Xu, Ying Zhu, and Chunshou Li. 2023. "Starch Properties and Morphology of Eight Floury Endosperm Mutants in Rice" Plants 12, no. 20: 3541. https://doi.org/10.3390/plants12203541
APA StyleHao, Y., Huang, F., Gao, Z., Xu, J., Zhu, Y., & Li, C. (2023). Starch Properties and Morphology of Eight Floury Endosperm Mutants in Rice. Plants, 12(20), 3541. https://doi.org/10.3390/plants12203541