
Phenotypic and Genotypic Diversity of Roots Response to Salt in Durum Wheat Seedlings




Abstract
1. Introduction
2. Results
2.1. Salt Effect on Durum Wheat Shoots Traits
2.2. Salt Effect on the Root System
2.3. Correlation among Traits
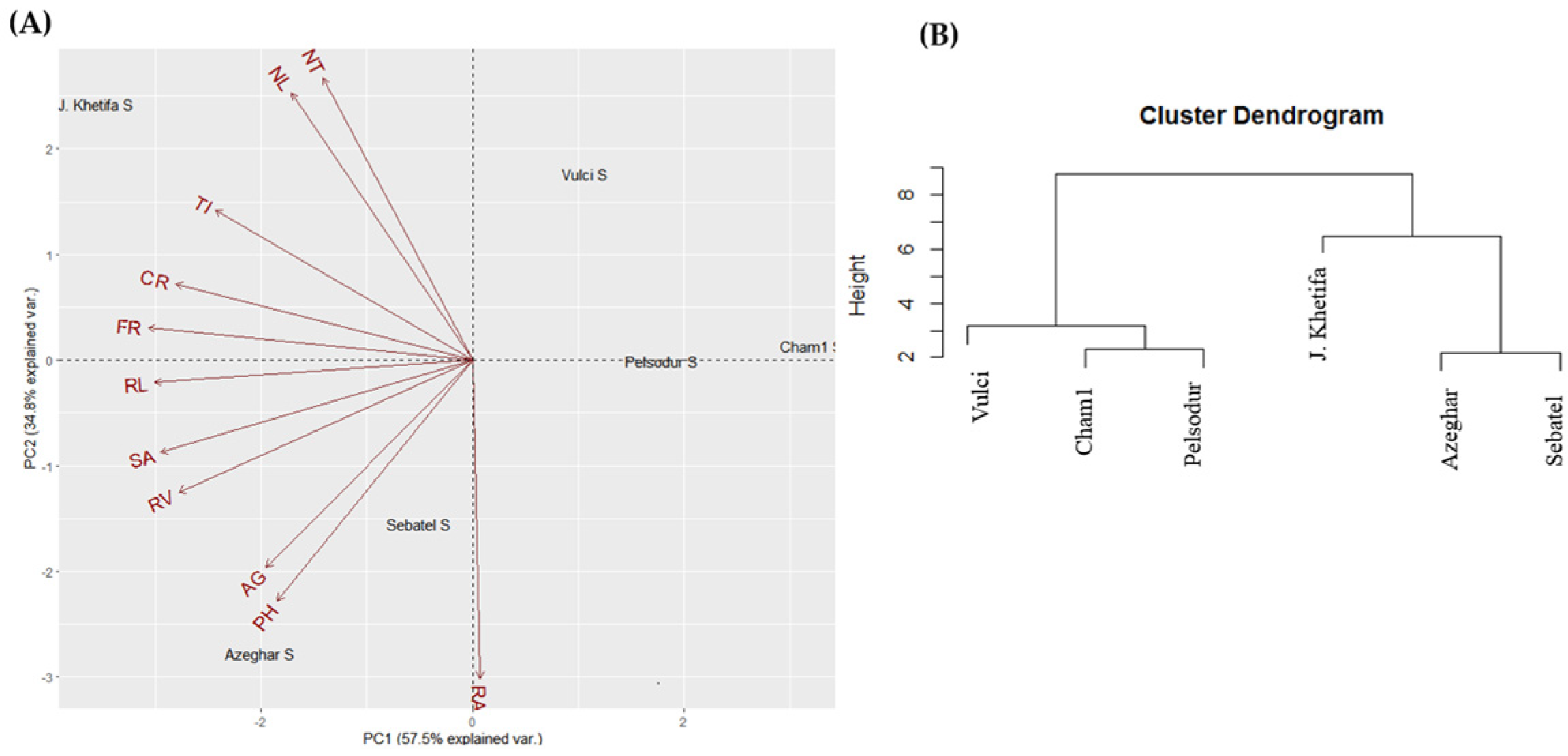
2.4. Principal Components Analysis (PCA)
2.5. Genetic Diversity Based on SSR Markers
2.6. More Detailed Analysis of Genomic Regions Linked to Selected SSRs Markers and In Silico Candidate Gene Expression Analysis
3. Discussion
4. Materials and Methods
4.1. Plant Material
4.2. Pot Experiment Condition and Design
4.3. Roots Phenotyping
4.4. DNA Extraction, PCR Amplifications, and Gel Electrophoresis
4.5. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
References
- Food and Agriculture Organization of the United Nations. FAO 2022 Global Map of Salt-Affected Soils. Fao Soils Portal. Available online: https://www.fao.org/soils-portal/data-hub/soil-maps-and-databases/global-map-of-salt-affected-soils/en/ (accessed on 10 August 2022).
- Urbanavičiūtė, I.; Bonfiglioli, L.; Pagnotta, M.A. One hundred candidate genes and their roles in drought and salt tolerance in wheat. Int. J. Mol. Sci. 2021, 22, 6378. [Google Scholar] [CrossRef] [PubMed]
- D’Odorico, P.; Bhattachan, A.; Davis, K.F.; Ravi, S.; Runyan, C.W. Global desertification: Drivers and feedbacks. Adv. Water Resour. 2013, 51, 326–344. [Google Scholar] [CrossRef]
- Daneshmand, H.; Alaghmand, S.; Camporese, M.; Talei, A.; Yeh, P.J.F.; Daly, E. Long-Term Impacts of Partial Afforestation on Water and Salt Dynamics of an Intermittent Catchment under Climate Change. Water 2020, 12, 1067. [Google Scholar] [CrossRef]
- El-Ramady, H.; Faizy, S.; Amer, M.M.; Elsakhawy, T.A.; Omara, A.E.-D.; Eid, Y.; Brevik, E. Management of Salt-Affected Soils: A Photographic Mini-Review. Environ. Biodivers. Soil Secur. 2022, 6, 61–79. [Google Scholar] [CrossRef]
- Corwin, D.L. Climate change impacts on soil salinity in agricultural areas. Eur. J. Soil Sci. 2021, 72, 842–862. [Google Scholar] [CrossRef]
- Daliakopoulos, I.N.; Tsanis, I.K.; Koutroulis, A.; Kourgialas, N.N.; Varouchakis, A.E.; Karatzas, G.P.; Ritsema, C.J. The threat of soil salinity: A European scale review. Sci. Total Environ. 2016, 573, 727–739. [Google Scholar] [CrossRef]
- Evelin, H.; Devi, T.S.; Gupta, S.; Kapoor, R. Mitigation of salinity stress in plants by arbuscular mycorrhizal symbiosis: Current understanding and new challenges. Front. Plant Sci. 2019, 10, 470. [Google Scholar] [CrossRef]
- Tedeschi, A. Irrigated Agriculture on Saline Soils: A Perspective. Agronomy 2020, 10, 1630. [Google Scholar] [CrossRef]
- MUNNS, R. Physiological processes limiting plant growth in saline soils: Some dogmas and hypotheses. Plant. Cell Environ. 1993, 16, 15–24. [Google Scholar] [CrossRef]
- Wang, J.; Zhu, J.; Zhang, Y.; Fan, F.; Li, W.; Wang, F.; Zhong, W.; Wang, C.; Yang, J. Comparative transcriptome analysis reveals molecular response to salinity stress of salt-tolerant and sensitive genotypes of indica rice at seedling stage. Sci. Rep. 2018, 8, 2085. [Google Scholar] [CrossRef]
- Qadir, M.; Quillérou, E.; Nangia, V.; Murtaza, G.; Singh, M.; Thomas, R.J.; Drechsel, P.; Noble, A.D. Economics of salt-induced land degradation and restoration. Nat. Resour. Forum 2014, 38, 282–295. [Google Scholar] [CrossRef]
- Safdarian, M.; Askari, H.; Shariati, J.V.; Nematzadeh, G. Transcriptional responses of wheat roots inoculated with Arthrobacter nitroguajacolicus to salt stress. Sci. Rep. 2019, 9, 1792. [Google Scholar] [CrossRef]
- Majeed, A.; Muhammad, Z.; Islam, S.; Ahmad, H. Salinity imposed stress on principal cereal crops and employing seed priming as a sustainable management approach. Acta Ecol. Sin. 2019, 39, 280–283. [Google Scholar] [CrossRef]
- Munns, R.; James, R.A.; Läuchli, A. Approaches to increasing the salt tolerance of wheat and other cereals. J. Exp. Bot. 2006, 57, 1025–1043. [Google Scholar] [CrossRef]
- Mansour, E.; Moustafa, E.S.A.; Desoky, E.S.M.; Ali, M.M.A.; Yasin, M.A.T.; Attia, A.; Alsuhaibani, N.; Tahir, M.U.; El-Hendawy, S. Multidimensional Evaluation for Detecting Salt Tolerance of Bread Wheat Genotypes Under Actual Saline Field Growing Conditions. Plants 2020, 9, 1324. [Google Scholar] [CrossRef] [PubMed]
- Attia, H.; Arnaud, N.; Karray, N.; Lachaâl, M. Long-term effects of mild salt stress on growth, ion accumulation and superoxide dismutase expression of Arabidopsis rosette leaves. Physiol. Plant. 2008, 132, 293–305. [Google Scholar] [CrossRef]
- West, G.; Inzé, D.; Beemster, G.T.S. Cell Cycle Modulation in the Response of the Primary Root of Arabidopsis to Salt Stress. Plant Physiol. 2004, 135, 1050–1058. [Google Scholar] [CrossRef]
- Julkowska, M.M.; Koevoets, I.T.; Mol, S.; Hoefsloot, H.; Feron, R.; Tester, M.A.; Keurentjes, J.J.B.; Korte, A.; Haring, M.A.; De Boer, G.J.; et al. Genetic components of root architecture remodeling in response to salt stress. Plant Cell 2017, 29, 3198–3213. [Google Scholar] [CrossRef]
- Kitomi, Y.; Hanzawa, E.; Kuya, N.; Inoue, H.; Hara, N.; Kawai, S.; Kanno, N.; Endo, M.; Sugimoto, K.; Yamazaki, T.; et al. Root angle modifications by the DRO1 homolog improve rice yields in saline paddy fields. Proc. Natl. Acad. Sci. USA 2020, 117, 21242–21250. [Google Scholar] [CrossRef] [PubMed]
- Sun, F.; Zhang, W.; Hu, H.; Li, B.; Wang, Y.; Zhao, Y.; Li, K.; Liu, M.; Li, X. Salt modulates gravity signaling pathway to regulate growth direction of primary roots in arabidopsis. Plant Physiol. 2007, 146, 178–188. [Google Scholar] [CrossRef]
- Fusi, R.; Rosignoli, S.; Lou, H.; Sangiorgi, G.; Bovina, R.; Pattem, J.K.; Borkar, A.N.; Lombardi, M.; Forestan, C.; Milner, S.G.; et al. Root angle is controlled by EGT1 in cereal crops employing an antigravitropic mechanism. Proc. Natl. Acad. Sci. USA 2022, 119, e2201350119. [Google Scholar] [CrossRef]
- El Hassouni, K.; Alahmad, S.; Belkadi, B.; Filali-Maltouf, A.; Hickey, L.T.; Bassi, F.M. Root system architecture and its association with yield under different water regimes in Durum wheat. Crop Sci. 2018, 58, 2331–2346. [Google Scholar] [CrossRef]
- Wasson, A.P.; Richards, R.A.; Chatrath, R.; Misra, S.C.; Prasad, S.V.S.; Rebetzke, G.J.; Kirkegaard, J.A.; Christopher, J.; Watt, M. Traits and selection strategies to improve root systems and water uptake in water-limited wheat crops. J. Exp. Bot. 2012, 63, 3485–3498. [Google Scholar] [CrossRef]
- Urbanaviči, I.; Bonfiglioli, L.; Pagnotta, M.A. Diversity in Root Architecture of Durum Wheat at Stem Elongation under Drought Stress. Agronomy 2022, 12, 1329. [Google Scholar] [CrossRef]
- Watt, M.; Moosavi, S.; Cunningham, S.C.; Kirkegaard, J.A.; Rebetzke, G.J.; Richards, R.A. A rapid, controlled-environment seedling root screen for wheat correlates well with rooting depths at vegetative, but not reproductive, stages at two field sites. Ann. Bot. 2013, 112, 447–455. [Google Scholar] [CrossRef]
- Gioia, T.; Galinski, A.; Lenz, H.; Müller, C.; Lentz, J.; Heinz, K.; Briese, C.; Putz, A.; Fiorani, F.; Watt, M.; et al. GrowScreen-PaGe, a non-invasive, high-throughput phenotyping system based on germination paper to quantify crop phenotypic diversity and plasticity of root traits under varying nutrient supply. Funct. Plant Biol. 2016, 44, 76–93. [Google Scholar] [CrossRef]
- Nagel, K.A.; Putz, A.; Gilmer, F.; Heinz, K.; Fischbach, A.; Pfeifer, J.; Faget, M.; Blossfeld, S.; Ernst, M.; Dimaki, C.; et al. GROWSCREEN-Rhizo is a novel phenotyping robot enabling simultaneous measurements of root and shoot growth for plants grown in soil-filled rhizotrons. Funct. Plant Biol. 2012, 39, 891–904. [Google Scholar] [CrossRef]
- Canales, F.J.; Nagel, K.A.; Müller, C.; Rispail, N.; Prats, E. Deciphering Root Architectural Traits Involved to Cope with Water Deficit in Oat. Front. Plant Sci. 2019, 10, 1558. [Google Scholar] [CrossRef] [PubMed]
- Soriano, J.M.; Alvaro, F. Discovering consensus genomic regions in wheat for root-related traits by QTL meta-analysis. Sci. Rep. 2019, 9, 10537. [Google Scholar] [CrossRef] [PubMed]
- Sanguineti, M.C.; Li, S.; MacCaferri, M.; Corneti, S.; Rotondo, F.; Chiari, T.; Tuberosa, R. Genetic dissection of seminal root architecture in elite durum wheat germplasm. Ann. Appl. Biol. 2007, 151, 291–305. [Google Scholar] [CrossRef]
- Canè, M.A.; Maccaferri, M.; Nazemi, G.; Salvi, S.; Francia, R.; Colalongo, C.; Tuberosa, R. Association mapping for root architectural traits in durum wheat seedlings as related to agronomic performance. Mol. Breed. 2014, 34, 1629–1645. [Google Scholar] [CrossRef] [PubMed]
- Iannucci, A.; Marone, D.; Russo, M.A.; De Vita, P.; Miullo, V.; Ferragonio, P.; Blanco, A.; Gadaleta, A.; Mastrangelo, A.M. Mapping QTL for Root and Shoot Morphological Traits in a Durum Wheat × T. dicoccum Segregating Population at Seedling Stage. Int. J. Genom. 2017, 2017, 6876393. [Google Scholar] [CrossRef]
- Maccaferri, M.; Stefanelli, S.; Rotondo, F.; Tuberosa, R.; Sanguineti, M.C. Relationships among durum wheat accessions. I. Comparative analysis of SSR, AFLP, and phenotypic data. Genome 2007, 50, 373–384. [Google Scholar] [CrossRef] [PubMed]
- Mondini, L.; Noorani, A.; Pagnotta, M.A. Assessing Plant Genetic Diversity by Molecular Tools. Diversity 2009, 1, 19–35. [Google Scholar] [CrossRef]
- Nei, M. Genetic Distance between Populations. Am. Nat. 1972, 106, 283–292. [Google Scholar] [CrossRef]
- Schneider, H.M.; Lor, V.S.N.; Hanlon, M.T.; Perkins, A.; Kaeppler, S.M.; Borkar, A.N.; Bhosale, R.; Zhang, X.; Rodriguez, J.; Bucksch, A.; et al. Root angle in maize influences nitrogen capture and is regulated by calcineurin B-like protein (CBL)-interacting serine/threonine-protein kinase 15 (ZmCIPK15). Plant. Cell Environ. 2022, 45, 837–853. [Google Scholar] [CrossRef] [PubMed]
- Pariyar, S.R.; Nagel, K.A.; Lentz, J.; Galinski, A.; Wilhelm, J.; Putz, A.; Adels, S.; Heinz, K.; Frohberg, C.; Watt, M. Variation in Root System Architecture among the Founder Parents of Two 8-way MAGIC Wheat Populations for Selection in Breeding. Agronomy 2021, 11, 2452. [Google Scholar] [CrossRef]
- Houshmand, S.; Arzani, A.; Maibody, S.A.M.; Feizi, M. Evaluation of salt-tolerant genotypes of durum wheat derived from in vitro and field experiments. F. Crop. Res. 2005, 91, 345–354. [Google Scholar] [CrossRef]
- Katerji, N.; Van Hoorn, J.W.; Hamdy, A.; Mastrorilli, M.; Oweis, T. Salt tolerance analysis of chickpea, faba bean and durum wheat varieties: I. Chickpea and faba bean. Agric. Water Manag. 2005, 72, 177–194. [Google Scholar] [CrossRef]
- Chamekh, Z.; Ayed, S.; Sahli, A.; Ayadi, S.; Hammemi, Z.; Jallouli, S.; Trifa, Y.; Amara, H.; Othmani, A.; Rezgui, M.; et al. Effect of salt stress on the flag leaf area and yield components in twenty five durum wheat genotypes (Triticum turgidum ssp. durum). J. New Sci. 2014, 6, 15–23. [Google Scholar]
- Ojeda, V.; Pérez-Ruiz, J.M.; González, M.; Nájera, V.A.; Sahrawy, M.; Serrato, A.J.; Geigenberger, P.; Cejudo, F.J. NADPH Thioredoxin Reductase C and Thioredoxins Act Concertedly in Seedling Development. Plant Physiol. 2017, 174, 1436–1448. [Google Scholar] [CrossRef]
- Li, S.; Chen, N.; Li, F.; Mei, F.; Wang, Z.; Cheng, X.; Kang, Z.; Mao, H. Characterization of wheat homeodomain-leucine zipper family genes and functional analysis of TaHDZ5-6A in drought tolerance in transgenic Arabidopsis. BMC Plant Biol. 2020, 20, 1–23. [Google Scholar] [CrossRef]
- Harris, J.C.; Sornaraj, P.; Taylor, M.; Bazanova, N.; Baumann, U.; Lovell, B.; Langridge, P.; Lopato, S.; Hrmova, M. Molecular interactions of the γ-clade homeodomain-leucine zipper class I transcription factors during the wheat response to water deficit. Plant Mol. Biol. 2016, 90, 435–452. [Google Scholar] [CrossRef] [PubMed]
- Ke, S.; Liu, S.; Luan, X.; Xie, X.M.; Hsieh, T.F.; Zhang, X.Q. Mutation in a putative glycosyltransferase-like gene causes programmed cell death and early leaf senescence in rice. Rice 2019, 12, 7. [Google Scholar] [CrossRef] [PubMed]
- Lu, F.; Cui, Y.; Zhang, J.; Du, X.; Hu, Y.; Yan, Y.; Mostofa, G.; Bajguz, A.; Yang, J.; Duan, W.; et al. Genome-Wide Identification and Characterisation of Wheat MATE Genes Reveals Their Roles in Aluminium Tolerance. Int. J. Mol. Sci. 2022, 23, 4418. [Google Scholar] [CrossRef]
- Nabi, R.B.S.; Tayade, R.; Imran, Q.M.; Hussain, A.; Shahid, M.; Yun, B.W. Functional Insight of Nitric-Oxide Induced DUF Genes in Arabidopsis thaliana. Front. Plant Sci. 2020, 11, 1041. [Google Scholar] [CrossRef] [PubMed]
- He, X.; Hou, X.; Shen, Y.; Huang, Z. TaSRG, a wheat transcription factor, significantly affects salt tolerance in transgenic rice and Arabidopsis. FEBS Lett. 2011, 585, 1231–1237. [Google Scholar] [CrossRef]
- Voss-Fels, K.P.; Robinson, H.; Mudge, S.R.; Richard, C.; Newman, S.; Wittkop, B.; Stahl, A.; Friedt, W.; Frisch, M.; Gabur, I.; et al. VERNALIZATION1 Modulates Root System Architecture in Wheat and Barley. Mol. Plant 2018, 11, 226. [Google Scholar] [CrossRef]
- Alvarez-Buylla, E.R.; García-Ponce, B.; de la Sánchez, M.P.; Espinosa-Soto, C.; García-Gómez, M.L.; Piñeyro-Nelson, A.; Garay-Arroyo, A. MADS-box genes underground becoming mainstream: Plant root developmental mechanisms. New Phytol. 2019, 223, 1143–1158. [Google Scholar] [CrossRef]
- Castilhos, G.; Lazzarotto, F.; Spagnolo-Fonini, L.; Bodanese-Zanettini, M.H.; Margis-Pinheiro, M. Possible roles of basic helix-loop-helix transcription factors in adaptation to drought. Plant Sci. 2014, 223, 1–7. [Google Scholar] [CrossRef]
- Yoo, C.M.; Wen, J.; Motes, C.M.; Sparks, J.A.; Blancaflor, E.B. A Class I ADP-Ribosylation Factor GTPase-Activating Protein Is Critical for Maintaining Directional Root Hair Growth in Arabidopsis. Plant Physiol. 2008, 147, 1659–1674. [Google Scholar] [CrossRef]
- Alahmad, S.; El Hassouni, K.; Bassi, F.M.; Dinglasan, E.; Youssef, C.; Quarry, G.; Aksoy, A.; Mazzucotelli, E.; Juhász, A.; Able, J.A.; et al. A major root architecture QTL responding to water limitation in durum wheat. Front. Plant Sci. 2019, 10, 436. [Google Scholar] [CrossRef] [PubMed]
- Suetsugu, N.; Higa, T.; Kong, S.G.; Wada, M. PLASTID MOVEMENT IMPAIRED1 and PLASTID MOVEMENT IMPAIRED1-RELATED1 Mediate Photorelocation Movements of Both Chloroplasts and Nuclei. Plant Physiol. 2015, 169, 1155–1167. [Google Scholar] [CrossRef] [PubMed]
- Qi, T.; Wang, J.; Sun, Q.; Day, B.; Guo, J.; Ma, Q. TaARPC3, contributes to wheat resistance against the stripe rust fungus. Front. Plant Sci. 2017, 8, 1245. [Google Scholar] [CrossRef]
- Pandian, B.A.; Sathishraj, R.; Djanaguiraman, M.; Prasad, P.V.V.; Jugulam, M. Role of Cytochrome P450 Enzymes in Plant Stress Response. Antioxidants 2020, 9, 454. [Google Scholar] [CrossRef] [PubMed]
- Buey, R.M.; Ledesma-Amaro, R.; Balsera, M.; de Pereda, J.M.; Revuelta, J.L. Increased riboflavin production by manipulation of inosine 5′-monophosphate dehydrogenase in Ashbya gossypii. Appl. Microbiol. Biotechnol. 2015, 99, 9577–9589. [Google Scholar] [CrossRef]
- Meena, M.; Divyanshu, K.; Kumar, S.; Swapnil, P.; Zehra, A.; Shukla, V.; Yadav, M.; Upadhyay, R.S. Regulation of L-proline biosynthesis, signal transduction, transport, accumulation and its vital role in plants during variable environmental conditions. Heliyon 2019, 5, e02952. [Google Scholar] [CrossRef] [PubMed]
- Singh, A.K.; Kumar, R.; Pareek, A.; Sopory, S.K.; Singla-Pareek, S.L. Overexpression of rice CBS domain containing protein improves salinity, oxidative, and heavy metal tolerance in transgenic tobacco. Mol. Biotechnol. 2012, 52, 205–216. [Google Scholar] [CrossRef] [PubMed]
- Lawlor, D.W.; Paul, M.J. Source/sink interactions underpin crop yield: The case for trehalose 6-phosphate/SnRK1 in improvement of wheat. Front. Plant Sci. 2014, 5, 418. [Google Scholar] [CrossRef]
- Wang, Y.; Ma, H. Step-wise and lineage-specific diversification of plant RNA polymerase genes and origin of the largest plant-specific subunits. New Phytol. 2015, 207, 1198–1212. [Google Scholar] [CrossRef]
- Krattinger, S.G.; Kang, J.; Bräunlich, S.; Boni, R.; Chauhan, H.; Selter, L.L.; Robinson, M.D.; Schmid, M.W.; Wiederhold, E.; Hensel, G.; et al. Abscisic acid is a substrate of the ABC transporter encoded by the durable wheat disease resistance gene Lr34. New Phytol. 2019, 223, 853–866. [Google Scholar] [CrossRef] [PubMed]
- Gietler, M.; Nykiel, M.; Orzechowski, S.; Fettke, J.; Zagdańska, B. Protein carbonylation linked to wheat seedling tolerance to water deficiency. Environ. Exp. Bot. 2017, 137, 84–95. [Google Scholar] [CrossRef]
- Fucile, G.; Falconer, S.; Christendat, D. Evolutionary Diversification of Plant Shikimate Kinase Gene Duplicates. PLoS Genet. 2008, 4, e1000292. [Google Scholar] [CrossRef] [PubMed]
- Trusov, Y.; Botella, J.R. Plant G-proteins come of age: Breaking the bond with animal models. Front. Chem. 2016, 4, 24. [Google Scholar] [CrossRef] [PubMed]
- Kato, Y.; Sakamoto, W. FtsH protease in the thylakoid membrane: Physiological functions and the regulation of protease activity. Front. Plant Sci. 2018, 9, 855. [Google Scholar] [CrossRef] [PubMed]
- Sengupta, D.; Naik, D.; Reddy, A.R. Plant aldo-keto reductases (AKRs) as multi-tasking soldiers involved in diverse plant metabolic processes and stress defense: A structure-function update. J. Plant Physiol. 2015, 179, 40–55. [Google Scholar] [CrossRef] [PubMed]
- Tejos, R.; Rodriguez-Furlán, C.; Adamowski, M.; Sauer, M.; Norambuena, L.; Friml, J. PATELLINS are regulators of auxin-mediated PIN1 relocation and plant development in Arabidopsis thaliana. J. Cell Sci. 2018, 131, jcs204198. [Google Scholar] [CrossRef]
- Zhang, J.; Xiao, J.; Li, Y.; Su, B.; Xu, H.; Shan, X.; Song, C.; Xie, J.; Li, R. PDM3, a pentatricopeptide repeat-containing protein, affects chloroplast development. J. Exp. Bot. 2017, 68, 5615–5627. [Google Scholar] [CrossRef]
- Gahlaut, V.; Jaiswal, V.; Kumar, A.; Gupta, P.K. Transcription factors involved in drought tolerance and their possible role in developing drought tolerant cultivars with emphasis on wheat (Triticum aestivum L.). Theor. Appl. Genet. 2016, 129, 2019–2042. [Google Scholar] [CrossRef]
- Furutani, M.; Hirano, Y.; Nishimura, T.; Nakamura, M.; Taniguchi, M.; Suzuki, K.; Oshida, R.; Kondo, C.; Sun, S.; Kato, K.; et al. Polar recruitment of RLD by LAZY1-like protein during gravity signaling in root branch angle control. Nat. Commun. 2020, 11, 1–3. [Google Scholar] [CrossRef]
- Grzesiak, M.T.; Hordyńska, N.; Maksymowicz, A.; Grzesiak, S.; Szechyńska-Hebda, M. Variation among Spring Wheat (Triticum aestivum L.) Genotypes in Response to the Drought Stress. II—Root System Structure. Plants 2019, 8, 584. [Google Scholar] [CrossRef] [PubMed]
- Almeida, D.M.; Gregorio, G.B.; Oliveira, M.M.; Saibo, N.J.M. Five novel transcription factors as potential regulators of OsNHX1 gene expression in a salt tolerant rice genotype. Plant Mol. Biol. 2016, 93, 61–77. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.J.; Yuan, S.; Zhang, N.H.; Lei, T.; Duan, H.G.; Liang, H.G.; Lin, H.H. Effect of water stress on photosystem 2 in two wheat cultivars. Biol. Plant. 2006, 50, 597–602. [Google Scholar] [CrossRef]
- Nigro, D.; Blanco, A.; Piarulli, L.; Signorile, M.A.; Colasuonno, P.; Blanco, E.; Simeone, R. Fine Mapping and Candidate Gene Analysis of Pm36, a Wild Emmer-Derived Powdery Mildew Resistance Locus in Durum Wheat. Int. J. Mol. Sci. 2022, 23, 13659. [Google Scholar] [CrossRef] [PubMed]
- Chen, M.; Markham, J.E.; Dietrich, C.R.; Jaworski, J.G.; Cahoon, E.B. Sphingolipid Long-Chain Base Hydroxylation Is Important for Growth and Regulation of Sphingolipid Content and Composition in Arabidopsis. Plant Cell 2008, 20, 1862–1878. [Google Scholar] [CrossRef]
- Mondini, L.; Nachit, M.; Porceddu, E.; Pagnotta, M.A. Identification of SNP mutations in DREB1, HKT1, and WRKY1 genes involved in drought and salt stress tolerance in durum wheat (Triticum turgidum L. var durum). Omi. A J. Integr. Biol. 2012, 16, 178–187. [Google Scholar] [CrossRef]
- Mondini, L.; Nachit, M.M.; Pagnotta, M.A. Allelic variants in durum wheat (Triticum turgidum L. var. durum) DREB genes conferring tolerance to abiotic stresses. Mol. Genet. Genom. 2015, 290, 531–544. [Google Scholar] [CrossRef]
- GitHub. Taiyun/Corrplot: A visual Exploratory Tool on Correlation Matrix. Available online: https://github.com/taiyun/corrplot (accessed on 23 December 2021).
- Vincent, Q.V. GitHub—Vqv/Ggbiplot: A Biplot Based on ggplot2. Available online: https://github.com/vqv/ggbiplot (accessed on 23 December 2021).
- Lewis, P.O. Genetic Data Analysis: Computer Program for the Analysis of Allelic Data. Version 1.0 (d16c). Available online: https://plewis.github.io/software/ (accessed on 20 December 2022).
- Peakall, R.; Smouse, P.E. GENALEX 6: Genetic analysis in Excel. Population genetic software for teaching and research. Mol. Ecol. Notes 2006, 6, 288–295. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genotype | Seedling Length, cm | Number of Leaves | Number of Tillers | ||||||
|---|---|---|---|---|---|---|---|---|---|
| Control | Salt | Diff. (%) | Control | Salt | Diff. (%) | Control | Salt | Diff. (%) | |
| Azeghar | 34.6 ± 3.2 b | 40.5 ± 1.6 a | 16.96 ns | 5.0 ± 0.0 c | 6.7 ± 0.6 c | 33.3 ns | 1.0 ± 0.0 c | 1.0 ± 0.0 b | 0.0 ns |
| Cham1 | 37.1 ± 0.6 ab | 31.3 ± 0.8 bc | −15.71 ns | 6.0 ± 1.7 c | 6.3 ± 1.2 c | 5.6 ns | 2.3 ± 0.6 b | 1.0 ± 0.0 b | −57.1 ** |
| J. Khetifa | 42.8 ± 1.1 a | 33.1 ± 2.6 b | −22.74 * | 9.3 ± 0.6 a | 14.0 ±0.0 a | 50.0 *** | 3.0 ± 0.0 ab | 3.3 ± 0.6 a | 11.1 ns |
| Pelsodur | 34.0 ± 1.5 b | 29.2 ± 2.2 c | −14.18 ns | 8.0 ± 0.0 ab | 7.3 ± 1.5 c | −8.3 ns | 3.3 ± 0.6 a | 1.3 ± 0.6 b | −60.0 *** |
| Sebatel | 36.4 ± 2.8 b | 34.8 ± 0.8 b | −4.40 ns | 5.7 ± 0.6 c | 7.3 ± 0.6 c | 29.4 ns | 1.0 ± 0.0 c | 1.0 ± 0.0 b | 0.0 ns |
| Vulci | 35.2 ± 7.0 b | 27.6 ± 2.4 c | −21.76 ns | 6.7 ± 1.5 bc | 11.3 ± 1.2 b | 70.0 *** | 2.3 ± 0.6 b | 2.7 ± 0.6 a | 14.3 ns |
| Genotype | ** | *** | *** | ||||||
| Treatment | *** | *** | ** | ||||||
| G × T | ** | *** | *** | ||||||
| ANOVA | Root Volume | Root Length | Tips | Root Surface Area |
| Genotype | * | * | * | ** |
| Treatments | *** | *** | *** | *** |
| G × T | ns | ns | * | ns |
| ANOVA | Root Angle | Crossings | Forks | Average Root Diameter |
| Genotype | *** | * | ** | ns |
| Treatments | *** | * | ** | ns |
| G × T | *** | ns | ns | ns |
| Trait | Treat | Genotype | |||||
|---|---|---|---|---|---|---|---|
| Azeghar | Cham1 | J. Khetifa | Pelsodur | Sebatel | Vulci | ||
| Root volume (cm3) | control | 0.80 ± 0.28 a | 0.57 ± 0.15 a | 0.57 ± 0.04 a | 0.61 ± 0.19 a | 0.68 ± 0.18 a | 0.48 ± 0.18 a |
| salt | 1.28 ± 0.24 a | 0.57 ± 0.02 c | 1.07 ± 0.34 ab | 0.68 ± 0.06 bc | 0.99 ± 0.15 abc | 0.77 ± 0.29 bc | |
| Diff. (%) | 59.6 * | 0.0 ns | 88.8 ** | 12.4 ns | 45.1 ns | 61.5 ns | |
| Root Length (cm) | control | 684 ± 106 a | 333 ± 73 c | 475 ± 104 bc | 521 ± 80 abc | 578 ± 55 ab | 530 ± 120 ab |
| salt | 826 ± 52 ab | 603 ± 23 b | 890 ± 192 a | 590 ± 7 b | 737 ± 200 ab | 627 ± 155 b | |
| Diff. (%) | 20.6 ns | 81.2 * | 87.3 ** | 13.4 ns | 27.6 ns | 18.2 ns | |
| Number of tips | control | 1243 ± 287 a | 633 ± 94 a | 1188 ± 406 a | 1214 ± 225 a | 1326 ± 507 a | 1272 ± 353 a |
| salt | 1510 ± 119 b | 1362 ± 21 b | 2399 ± 143 a | 1401 ± 265 b | 1480 ± 352 b | 1311 ± 309 b | |
| Diff. (%) | 21.5 ns | 115.3 * | 101.9 *** | 15.4 ns | 11.6 ns | 3.0 ns | |
| Root surface area (cm2) | control | 81.4 ± 7.8 a | 48.5 ± 11.8 b | 57.7 ± 4.2 b | 62.8 ± 14.5 ab | 69.9 ± 11.6 ab | 56.4 ± 16.5 b |
| salt | 114.9 ± 14 a | 65.5 ± 0.0 c | 109 ± 28 ab | 71.1 ± 2.6 bc | 95.3 ± 20.3 abc | 77.2 ± 21.7 bc | |
| Diff. (%) | 41.2 * | 35.1 ns | 89.0 ** | 13.2 ns | 36.3 ns | 37.0 ns | |
| Root angle (°) | control | 110.1 ± 4.4 a | 109.4 ± 7.4 a | 90.1 ± 3.3 b | 79.7 ± 1.3 c | 106.1 ± 2.3 a | 84.9 ± 2.8 bc |
| salt | 117.0 ± 7.9 a | 104.5 ± 3.2 bc | 95.4 ± 4.3 d | 109.5 ± 6.6 ab | 115.3 ± 2.7 a | 96.7 ± 2.9 cd | |
| Diff. (%) | 6.2 ns | −4.5 ns | 6.0 ns | 37.3 *** | 8.7 * | 13.9 * | |
| Crossings | control | 956 ± 487 a | 421 ± 162 a | 942 ± 204 a | 887 ± 62 a | 857 ± 136 a | 859 ± 218 a |
| salt | 1056 ± 97 abc | 512 ± 315 c | 1543 ± 423 a | 981 ± 241 abc | 1122 ± 503 ab | 916 ± 266 bc | |
| Diff. (%) | 10.5 ns | 21.8 ns | 63.9 * | 10.6 ns | 30.9 ns | 6.6 ns | |
| Forks | control | 4990 ± 914 a | 2774 ± 542 b | 4651 ± 511 a | 4736 ± 531 a | 4587 ± 897 a | 4152 ± 931 ab |
| salt | 6606 ± 143 ab | 3547 ± 242 c | 8180 ± 1987 a | 4801 ± 183 bc | 6100 ± 2120 abc | 5111 ± 1201 bc | |
| 32.4 ns | 27.9 ns | 75.9 * | 1.4 ns | 33.0 ns | 23.1 ns | ||
| Mean diameter (mm) | control | 0.39 ± 0.1 ab | 0.46 ± 0.01 a | 0.39 ± 0.06 ab | 0.38 ± 0.03 ab | 0.38 ± 0.05 ab | 0.33 ± 0.3 b |
| salt | 0.44 ± 0.03 a | 0.35 ± 0.01 b | 0.39 ± 0.03 ab | 0.39 ± 0.02 ab | 0.42 ± 0.03 ab | 0.39 ± 0.06 ab | |
| Diff. (%) | 12.8 ns | −23.9 * | −1.7 ns | 1.3 ns | 8.7 ns | 17.0 ns | |
| Locus | Na | I | Ho | He | uHe | PIC | Size, bp |
|---|---|---|---|---|---|---|---|
| cfa2086 | 5 | 1.561 | 0.000 | 0.778 | 0.848 | 0.744 | 223–299 |
| wms5 | 3 | 1.011 | 0.000 | 0.611 | 0.667 | 0.536 | 180–190 |
| gwm234a | 2 | 0.451 | 0.000 | 0.278 | 0.303 | 0.240 | 214–218 |
| gwm234b | 2 | 0.693 | 0.000 | 0.500 | 0.545 | 0.375 | 242–256 |
| wmc727a | 2 | 0.679 | 0.833 | 0.486 | 0.530 | 0.368 | 88–96 |
| wmc727b | 2 | 0.451 | 0.000 | 0.278 | 0.303 | 0.240 | null-231 |
| cfa2257 | 5 | 1.424 | 0.333 | 0.722 | 0.788 | 0.680 | 104–156 |
| wms205 | 2 | 0.693 | 1.000 | 0.500 | 0.545 | 0.375 | 157–170 |
| gwm427 | 4 | 1.330 | 0.000 | 0.722 | 0.788 | 0.672 | 201–256 |
| gwm573.2 | 5 | 1.424 | 1.000 | 0.722 | 0.788 | 0.680 | 181–240 |
| gwm636 | 3 | 1.011 | 0.000 | 0.611 | 0.667 | 0.536 | 110–131 |
| gwm459a | 3 | 1.011 | 0.000 | 0.611 | 0.667 | 0.536 | 131–153 |
| gwm499b | 4 | 1.330 | 0.000 | 0.722 | 0.788 | 0.672 | 147–197 |
| mean | 3.2 | 1.005 | 0.244 | 0.580 | 0.633 |
| Locus | Allele | Frequency | Found in |
|---|---|---|---|
| cfa2086 | 223 | 1 | Sebatel |
| cfa2086 | 299 | 1 | Pelsodur |
| cfa2086 | 269 | 1 | Cham |
| cfa2086 | 277 | 1 | J. Khetifa |
| wms5 | 185 | 1 | Cham |
| gwm234a | 214 | 1 | J. Khetifa |
| wmc727a | 1 | 1 | Pelsodur |
| cfa2257 | 121 | 1 | Cham |
| cfa2257 | 142 | 0.5 | Azeghar |
| cfa2257 | 104 | 0.5 | Azeghar |
| gwm427 | 237 | 1 | Sebatel |
| gwm427 | 201 | 1 | Azeghar |
| gwm573.2 | 226 | 0.5 | Pelsodur |
| gwm573.2 | 181 | 0.5 | Azeghar |
| gwm636 | 131 | 1 | Pelsodur |
| gwm459a | 131 | 1 | Vulci |
| gwm499b | 147 | 1 | Sebatel |
| gwm499b | 154 | 1 | J. Khetifa |
| Marker | Primer Forward (5′-3′) | Primer Reverse (5′-3′) | Chr. | AT (°C) |
|---|---|---|---|---|
| cfa2086 | TCTACTTTCAGGGCACCTCG | TCTCTCCAAACCTCCCTGTAA | 2A | 56 |
| gwm573.2 | AAGAGATAACATGCAAGAAA | TTCAAATATGTGGGAACTAC | 7B | 45 |
| wmc727 | CATAATCAGGACAGCCGCAC | TAGTGGCCTGATGTATCTAGTTGG | 5A | 55 |
| wms205 | CGACCCGGTTCACTTCAG | AGTCGCCGTTGTATAGTGCC | 5A | 56 |
| wms5 | GCCAGCTACCTCGATACAACTC | AGAAAGGGCCAGGCTAGTAGT | 3A | 56 |
| gmw459 | ATGGAGTGGTCACACTTTGAA | AGCTTCTCTGACCAACTTCTCG | 6A | 54 |
| gwm234 | GAGTCCTGATGTGAAGCTGTTG | CTCATTGGGGTGTGTACGTG | 5B | 55 |
| gwm427 | AAACTTAGAACTGTAATTTCAGA | AGTGTGTTCATTTGACAGTT | 6A | 45 |
| gwm499 | ACTTGTATGCTCCATTGATTGG | GGGAGTGGAAACTGCATAA | 5B | 52 |
| gwm637 | AAAGAGGTCTGCCGCTAACA | TATACGGTTTTGTGAGGGGG | 4A | 55 |
| cfa2257 | GATACAATAGGTGCCTCCGC | CCATTATGTAAATGCTTCTGTTTGA | 7A | 49 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Urbanavičiūtė, I.; Bonfiglioli, L.; Pagnotta, M.A. Phenotypic and Genotypic Diversity of Roots Response to Salt in Durum Wheat Seedlings. Plants 2023, 12, 412. https://doi.org/10.3390/plants12020412
Urbanavičiūtė I, Bonfiglioli L, Pagnotta MA. Phenotypic and Genotypic Diversity of Roots Response to Salt in Durum Wheat Seedlings. Plants. 2023; 12(2):412. https://doi.org/10.3390/plants12020412
Chicago/Turabian StyleUrbanavičiūtė, Ieva, Luca Bonfiglioli, and Mario A. Pagnotta. 2023. "Phenotypic and Genotypic Diversity of Roots Response to Salt in Durum Wheat Seedlings" Plants 12, no. 2: 412. https://doi.org/10.3390/plants12020412
APA StyleUrbanavičiūtė, I., Bonfiglioli, L., & Pagnotta, M. A. (2023). Phenotypic and Genotypic Diversity of Roots Response to Salt in Durum Wheat Seedlings. Plants, 12(2), 412. https://doi.org/10.3390/plants12020412