
Overexpression of IlHMA2, from Iris lactea, Improves the Accumulation of and Tolerance to Cadmium in Tobacco


Abstract
:1. Introduction
2. Results
2.1. Overexpression of IlHMA2 Improved the Growth of Tobacco under Cd Exposure
2.2. Overexpression of IlHMA2 Reduced the Lipid Peroxidation and Overproduction of ROS in Tobacco under Cd Exposure
2.3. Overexpression of IlHMA2 Enhanced the Root-to-Shoot Cd Translocation in Tobacco under Cd Exposure
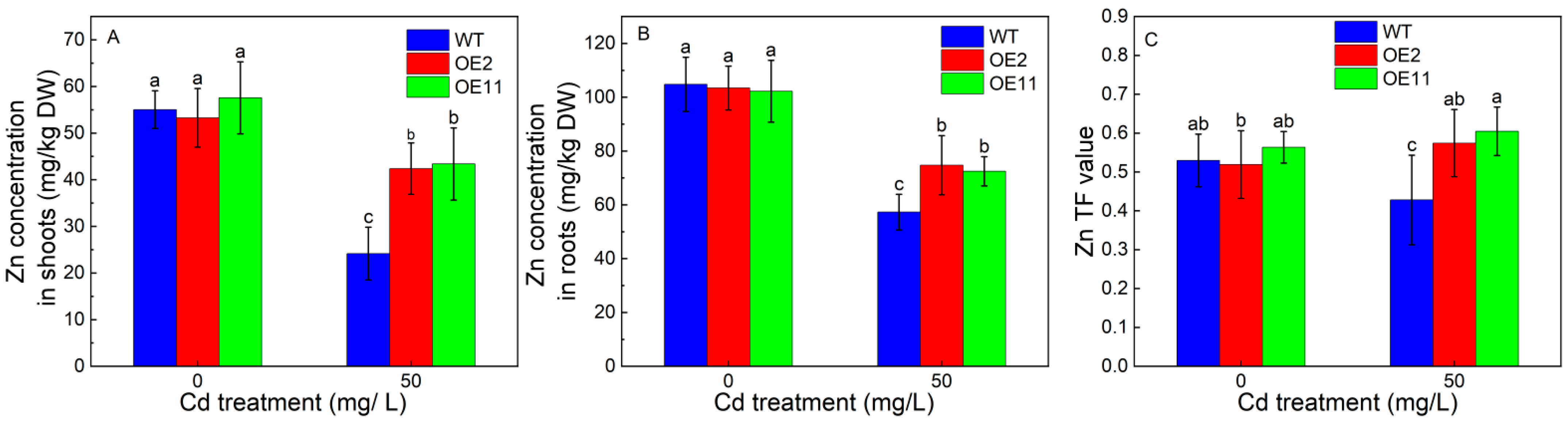
2.4. Overexpression of IlHMA2 Promoted the Accumulation of Zn in Tobacco under Cd Exposure
3. Discussion
3.1. Overexpression of IlHMA2 Contributed to Scavenging the ROS in Tobacco via Modulating Antioxidant Systems under Cd Contamination
3.2. Overexpression of IlHMA2 Facilitated the Root-to-Shoot Zn Translocation to Improve the Physiological Zn Status in Tobacco under Cd Contamination
3.3. Overexpression of IlHMA2 Enhances Cd Phytoremediation Efficiency in Tobacco
4. Materials and Methods
4.1. Plant Material and Growth Condition
4.2. Cloning and Vector Construction
4.3. Plant Transformation and Molecular Characterization
4.4. Cd treatments and Measurement of Growth Parameters for Transgenic Lines
4.5. Measurement of H2O2, MDA, and Antioxidant Enzymes for Transgenic Lines
4.6. Determination of Zn and Cd Concentration
4.7. Statistic Analysis
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Guo, Q.; Meng, L.; Humphreys, M.W.; Scullion, J.; Mur, L.A.J. Expression of FlHMA3, a P1B2-ATPase from Festulolium loliaceum, correlates with response to cadmium stress. Plant Physiol. Biochem. 2017, 112, 270–277. [Google Scholar] [CrossRef]
- Guo, Q.; Meng, L.; Zhang, Y.; Mao, P.C.; Tian, X.X.; Li, S.S.; Zhang, L. Antioxidative systems, metal ion homeostasis and cadmium distribution in Iris lactea exposed to cadmium stress. Ecotoxicol. Environ. Saf. 2017, 139, 50–55. [Google Scholar] [CrossRef] [PubMed]
- Shen, X.; Dai, M.; Yang, J.; Sun, L.; Tan, X.; Peng, C.; Ali, I.; Naz, I. A critical review on the phytoremediation of heavy metals from environment: Performance and challenges. Chemosphere 2022, 291, 132979. [Google Scholar] [CrossRef] [PubMed]
- Mendoza-Cózatl, D.G.; Jobe, T.O.; Hauser, F.; Schroeder, J.I. Long-distance transport, vacuolar sequestration, tolerance, and transcriptional responses induced by cadmium and arsenic. Curr. Opin. Plant Biol. 2011, 14, 554–562. [Google Scholar] [CrossRef] [PubMed]
- Colangelo, E.P.; Guerinot, M.L. Put the metal to the petal: Metal uptake and transport throughout plants. Curr. Opin. Plant Biol. 2006, 9, 322–330. [Google Scholar] [CrossRef]
- Grotz, N.; Guerinot, M.L. Molecular aspects of Cu, Fe and Zn homeostasis in plants. Biochim. Biophys. Acta 2006, 1763, 595–608. [Google Scholar] [CrossRef]
- Williams, L.E.; Mills, R.F. P1B-ATPases an ancient family of transition metal pumps with diverse functions in plants. Trends Plant Sci. 2005, 10, 491–502. [Google Scholar] [CrossRef] [PubMed]
- Wong, C.K.E.; Cobbett, C.S. HMA P-type ATPases are the major mechanism for root-to-shoot Cd translocation in Arabidopsis thaliana. New Phytol. 2009, 181, 71–78. [Google Scholar] [CrossRef]
- Axelsen, K.B.; Palmgren, M.G. Inventory of the superfamily of P-type ion pumps in Arabidopsis. Plant Physiol. 2001, 126, 696–706. [Google Scholar] [CrossRef] [PubMed]
- Eren, E.; Argüello, J.M. Arabidopsis HMA2, a divalent heavy metal-transporting P(IB)-type ATPase, is involved in cytoplasmic Zn2+ homeostasis. Plant Physiol. 2004, 136, 3712–3723. [Google Scholar] [CrossRef]
- Takahashi, R.; Ishimaru, Y.; Shimo, H.; Ogo, Y.; Senoura, T.; Nishizawa, N.K.; Nakanishi, H. The OsHMA2 transporter is involved in root-to-shoot translocation of Zn and Cd in rice. Plant Cell Environ. 2012, 35, 1948–1957. [Google Scholar] [CrossRef] [PubMed]
- Mills, R.F.; Peaston, K.A.; Runions, J.; Williams, L.E. HvHMA2, a P1B-ATPase from barley, is highly conserved among cereals and functions in Zn and Cd transport. PLoS ONE 2012, 7, e42640. [Google Scholar] [CrossRef] [PubMed]
- Tan, J.; Wang, J.; Chai, T.; Zhang, Y.X.; Feng, S.S.; Li, Y.; Zhao, H.J.; Liu, H.M.; Chai, X.P. Functional analyses of TaHMA2, a P1B -type ATPase in wheat. Plant Biotechnol. J. 2013, 11, 420–431. [Google Scholar] [CrossRef]
- Hanikenne, M.; Talke, I.N.; Haydon, M.J.; Lanz, C.; Nolte, A.; Motte, P.; Kroymann, J.; Weigel, D.; Krämer, U. Evolution of metal hyperaccumulation required cis-regulatory changes and triplication of HMA4. Nature 2008, 453, 391–395. [Google Scholar] [CrossRef] [PubMed]
- Verret, F.; Gravot, A.; Auroy, P.; Leonhardt, N.; David, P.; Nussaume, L.; Vavasseur, A.; Richaud, P. Overexpression of AtHMA4 enhances root-to-shoot translocation of zinc and cadmium and plant metal tolerance. FEBS Lett. 2004, 576, 306–312. [Google Scholar] [CrossRef]
- Migocka, M.; Papierniak, A.; Maciaszczyk-Dziubinska, E.; Posyniak, E.; Kosieradzka, A. Molecular and biochemical properties of two P1B2-ATPases, CsHMA3 and CsHMA4, from cucumber. Plant Cell Environ. 2015, 38, 1127–1141. [Google Scholar] [CrossRef]
- Zheng, S.W.; Liu, S.B.; Feng, J.H.; Wang, W.; Wang, Y.W.; Yu, Q.; Liao, Y.R.; Mo, Y.H.; Xu, Z.J.; Li, L.H.; et al. Overexpression of a stress response membrane protein gene OsSMP1 enhances rice tolerance to salt, cold and heavy metal stress. Environ. Exp. Bot. 2021, 182, 104327. [Google Scholar] [CrossRef]
- Fasani, E.; Manara, A.; Martini, F.; Furini, A.; DalCorso, G. The potential of genetic engineering of plants for the remediation of soils contaminated with heavy metals. Plant Cell Environ. 2018, 41, 1201–1232. [Google Scholar] [CrossRef]
- Guo, Q.; Tian, X.X.; Mao, P.C.; Meng, L. Functional characterization of IlHMA2, a P1B2-ATPase in Iris lactea response to Cd. Environ. Exp. Bot. 2019, 157, 131–139. [Google Scholar] [CrossRef]
- Cuypers, A.; Plusquin, M.; Remans, T.; Jozefczak, M.; Keunen, E.; Gielen, H.; Opdenakker, K.; Nair, A.R.; Munters, E.; Artois, T.J.; et al. Cadmium stress: An oxidative challenge. Biometals 2010, 23, 927–940. [Google Scholar] [CrossRef]
- Guo, Q.; Meng, L.; Mao, P.C.; Tian, X.X. An assessment of Agropyron cristatum tolerance to cadmium contaminated soil. Biol. Plant. 2014, 58, 174–178. [Google Scholar] [CrossRef]
- Tian, S.; Liang, S.; Qiao, K.; Wang, F.; Zhang, Y.; Chai, T. Co-expression of multiple heavy metal transporters changes the translocation, accumulation, and potential oxidative stress of Cd and Zn in rice (Oryza sativa). J. Hazard. Mater. 2019, 380, 120853. [Google Scholar] [CrossRef]
- Huang, Y.; Zu, L.; Zhang, M.; Yang, T.; Zhou, M.; Shi, C.; Shi, F.; Zhang, W. Tolerance and distribution of cadmium in an ornamental species Althaea rosea Cavan. Int. J. Phytoremediation. 2020, 22, 713–724. [Google Scholar] [CrossRef] [PubMed]
- Ekmekçi, Y.; Tanyolac, D.; Ayhan, B. Effects of cadmium on antioxidant enzyme and photosynthetic activities in leaves of two maize cultivars. J. Plant Physiol. 2008, 165, 600–611. [Google Scholar] [CrossRef] [PubMed]
- Anjum, A.S.; Tanveer, M.; Hussain, S.; Bao, M.C.; Wang, L.C.; Khan, I.; Ullah, E.; Tung, S.A.; Samad, R.A.; Shahzad, B. Cadmium toxicity in maize (Zea mays L.): Consequences on antioxidative systems, reactive oxygen species and cadmium accumulation. Environ. Sci. Pollut. Res. 2015, 22, 17022–17030. [Google Scholar] [CrossRef]
- Krantev, A.; Yordanova, R.; Janda, T.; Szalai, G.; Popova, L. Treatment with salicylic acid decreases the effect of cadmium on photosynthesis in maize plants. J. Plant Physiol. 2008, 165, 920–931. [Google Scholar] [CrossRef] [PubMed]
- Gill, S.S.; Tuteja, N. Reactive oxygen species and antioxidant machinery in abiotic stress tolerance in crop plants. Plant Physiol. Biochem. 2010, 48, 909–930. [Google Scholar] [CrossRef]
- Nazar, R.; Iqbal, N.; Masood, A.; Khan, M.I.R.; Syeed, S.; Khan, N.A. Cadmium toxicity in plants and role of mineral nutrients in its alleviation. Am. J. Plant Sci. 2012, 3, 1476–1489. [Google Scholar] [CrossRef]
- Zhang, H.; Yao, T.; Wang, Y.; Wang, J.; Song, J.; Cui, C.; Ji, G.; Cao, J.; Muhammad, S.; Ao, H.; et al. Trx CDSP32-overexpressing tobacco plants improves cadmium tolerance by modulating antioxidant mechanism. Plant Physiol. Biochem. 2023, 194, 524–532. [Google Scholar] [CrossRef]
- Yan, H.; Filardo, F.; Hu, X.; Zhao, X.; Fu, D. Cadmium stress alters the redox reaction and hormone balance in oilseed rape (Brassica napus L.) leaves. Environ. Sci. Pollut. Res. Int. 2016, 23, 3758–3769. [Google Scholar] [CrossRef]
- Krämer, U.; Talke, I.N.; Hanikenne, M. Transition metal transport. FEBS Lett. 2007, 581, 2263–2272. [Google Scholar] [CrossRef] [PubMed]
- Satoh-Nagasawa, N.; Mori, M.; Nakazawa, N.; Kawamoto, T.; Nagato, Y.; Sakurai, K.; Takahashi, H.; Watanabe, A.; Akagi, H. Mutations in rice (Oryza sativa) heavy metal ATPase 2 (OsHMA2) restrict the translocation of zinc and cadmium. Plant Cell Physiol. 2012, 53, 213–224. [Google Scholar] [CrossRef] [PubMed]
- Sinclair, S.A.; Senger, T.; Talke, I.N.; Cobbett, C.S.; Haydon, M.J.; Krämer, U. Systemic upregulation of MTP2- and HMA2-mediated Zn partitioning to the shoot supplements local Zn deficiency responses. Plant Cell 2018, 30, 2463–2479. [Google Scholar] [CrossRef] [PubMed]
- Guo, Q.; Tian, X.X.; Mao, P.C.; Meng, L. Overexpression of Iris lactea tonoplast Na+/H+ antiporter gene IlNHX confers improved salt tolerance in tobacco. Biol. Plant. 2020, 64, 50–57. [Google Scholar] [CrossRef]
- Li, C.; Mur, L.A.J.; Wang, Q.; Hou, X.; Zhao, C.; Chen, Z.; Wu, J.; Guo, Q. ROS scavenging and ion homeostasis is required for the adaptation of halophyte Karelinia caspia to high salinity. Front. Plant Sci. 2022, 13, 979956. [Google Scholar] [CrossRef]
- Liu, S.; Mo, X.; Sun, L.; Gao, L.; Su, L.; An, Y.; Zhou, P. MsDjB4, a HSP40 chaperone in alfalfa (Medicago sativa L.), improves alfalfa hairy root tolerance to aluminum stress. Plants 2023, 12, 2808. [Google Scholar] [CrossRef]
- Bates, L.S.; Waldren, R.P.; Teare, I.D. Rapid determination of free proline for water-stress studies. Plant Soil 1973, 39, 205–207. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cd Treatment (mg/L) | Lines | MDA (mmol/g FW) | H2O2 (μmol/g FW) | Proline (μg/g FW) |
|---|---|---|---|---|
| 0 | WT | 0.31 ± 0.03 c | 2.05 ± 0.07 b | 0.69 ± 0.02 b |
| OE2 | 0.31 ± 0.04 c | 2.16 ± 0.23 b | 0.74 ± 0.05 b | |
| OE11 | 0.28 ± 0.03 c | 1.95 ± 0.27 b | 0.71 ± 0.07 b | |
| 50 | WT | 1.3 ± 0.06 a | 3.82 ± 0.46 a | 0.75 ± 0.02 b |
| OE2 | 0.74 ± 0.09 b | 2.52 ± 0.58 b | 1.53 ± 0.17 a | |
| OE11 | 0.73 ± 0.05 b | 2.67 ± 0.57 b | 1.61 ± 0.33 a |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, C.; Wang, Q.; Hou, X.; Zhao, C.; Guo, Q. Overexpression of IlHMA2, from Iris lactea, Improves the Accumulation of and Tolerance to Cadmium in Tobacco. Plants 2023, 12, 3460. https://doi.org/10.3390/plants12193460
Li C, Wang Q, Hou X, Zhao C, Guo Q. Overexpression of IlHMA2, from Iris lactea, Improves the Accumulation of and Tolerance to Cadmium in Tobacco. Plants. 2023; 12(19):3460. https://doi.org/10.3390/plants12193460
Chicago/Turabian StyleLi, Cui, Qinghai Wang, Xincun Hou, Chunqiao Zhao, and Qiang Guo. 2023. "Overexpression of IlHMA2, from Iris lactea, Improves the Accumulation of and Tolerance to Cadmium in Tobacco" Plants 12, no. 19: 3460. https://doi.org/10.3390/plants12193460
APA StyleLi, C., Wang, Q., Hou, X., Zhao, C., & Guo, Q. (2023). Overexpression of IlHMA2, from Iris lactea, Improves the Accumulation of and Tolerance to Cadmium in Tobacco. Plants, 12(19), 3460. https://doi.org/10.3390/plants12193460