
Challenges and Opportunities behind the Use of Herbaria in Paleogenomics Studies

 , ,
, ,  , , , ,
, , , , 
Abstract
1. Introduction
1.1. History and State of the Art of Herbaria
1.2. Herbarium Genomics
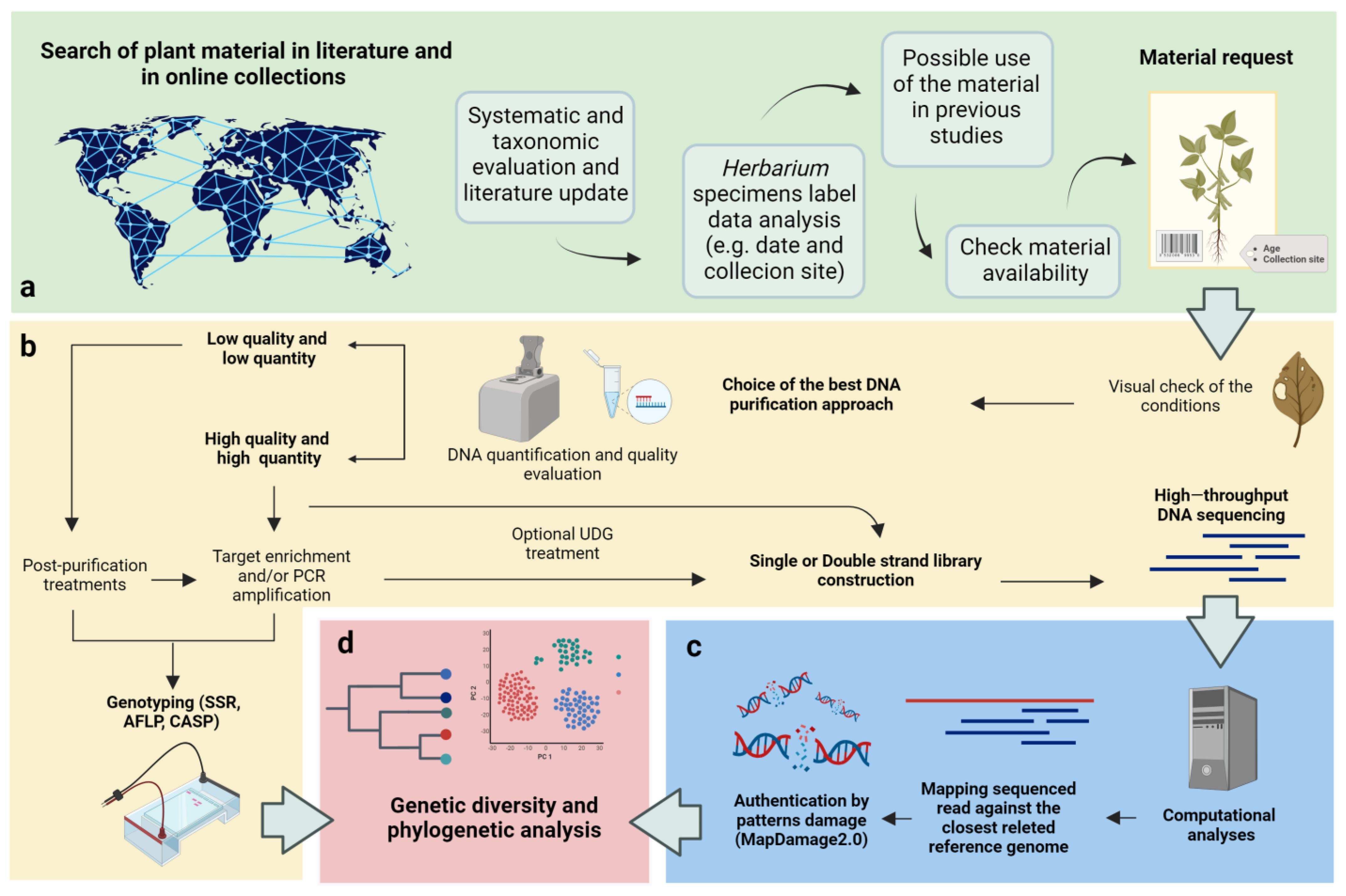
1.3. Challenges in the Use of Herbaria and Ancient Samples for Genomics Studies

{kind=link}
{kind=link}
{kind=link}
| DNA Extraction/Purification Protocol | Source of Plant Material | Timing of Sampling | Suitable for Extracting Herbarium DNA | Quality Evaluation Approach | Reference |
|---|---|---|---|---|---|
| CTAB, according to the protocol of Doyle and Doyle (1990) | Juncus and Luzula genera (Juncaceae) | * | Yes | PCR amplification | [43] |
| CTAB + pre-wash with a sorbitol-containing buffer | Lafoensia spp. | N.A. | Yes | PCR amplification | [44] |
| Modified CTAB | Agropyronjunceum (Gramineae), Poa juncifolia (Gramineae), Poa palustris, Triticum aestivum (Gramineae), Vicia faba (Fabaceae), Zea mays ssp. mays | N.A. | Yes | Restriction enzymes | [45] |
| Juncus and Luzula genera (Juncaceae) | N.A. | Yes, but may present CTAB contamination | PCR amplification | [43] | |
| N.A. | ≥60 years | Yes | PCR amplification | [46] | |
| Species from nine genera of the Papilionoideae | N.A. | Yes | PCR amplification | [47] | |
| DNeasy Plant Mini Kit (QIAGEN) | Juncus and Luzula genera (Juncaceae) | N.A. | Yes | PCR amplification | [43] |
| DNA extraction with phenol purification and liquid nitrogen | Juncus and Luzula genera (Juncaceae) | N.A. | No | PCR amplification | [43] |
| Long-term precipitation in isopropanol and CsCl gradient | Juncus and Luzula genera (Juncaceae) | N.A. | No | PCR amplification | [43] |
| Proteinase K and sodium dodecyl sulfate (SDS) | Scripus hattorianus | 1934 | Yes | PCR amplification | [48] |
| N-phenacylthiazolium bromide (PTB)—dithiothreitol (DTT) | Arabidopsis thaliana | Between 1839 and 1898 | Yes | NGS | [49] |
| Phenol-chloroform and silica spin column purification | Herbarium grape leaf tissue (unpublished data) | N.A. | Yes | PCR amplification | [50] |
| Polyvinylpyrrolidone PVP | genus Dalbergia | N.A. | Yes | PCR amplification | [51] |
| AMPure XP magnetic beads/PEG 8000-containing buffer | genus Scorzonera | Between 1920 and 1960 | Yes | PCR amplification | [52] |
2. Extraction and Purification of aDNA from Herbarium Specimens
3. DNA Amplification by PCR from Herbarium Specimens
4. Next-Generation Sequencing (NGS) and Genotyping on Herbarium Specimens to Disentangle Relevant Aspects of the Evolutionary History of a Species
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Bebber, D.P.; Carine, M.A.; Wood, J.R.I.; Wortley, A.H.; Harris, D.J.; Prance, G.T.; Davidse, G.; Paige, J.; Pennington, T.D.; Robson, N.K.B.; et al. Herbaria are a major frontier for species discovery. Proc. Natl. Acad. Sci. USA 2010, 107, 22169–22171. [Google Scholar] [CrossRef] [PubMed]
- Zunic, L.; Skrbo, A.; Dobraca, A. Historical Contribution of Pharmaceutics to Botany and Pharmacognosy Development. Mater. Socio Medica 2017, 29, 291–300. [Google Scholar] [CrossRef] [PubMed]
- Willis, C.G.; Ellwood, E.R.; Primack, R.B.; Davis, C.C.; Pearson, K.D.; Gallinat, A.S.; Yost, J.M.; Nelson, G.; Mazer, S.J.; Rossington, N.L.; et al. Old Plants, New Tricks: Phenological Research Using Herbarium Specimens. Trends Ecol. Evol. 2017, 32, 531–546. [Google Scholar] [CrossRef] [PubMed]
- Taylor, J.W.; Swann, E.C. DNA from Herbarium Specimens. In Ancient DNA: Recovery and Analysis of Genetic Material from Paleontological, Archaeological, Museum, Medical, and Forensic Specimens; Herrmann, B., Hummel, S., Eds.; Springer: New York, NY, USA, 1994; pp. 166–181. [Google Scholar] [CrossRef]
- Heberling, J.M. Herbaria as Big Data Sources of Plant Traits. Int. J. Plant Sci. 2022, 183, 87–118. [Google Scholar] [CrossRef]
- Von Engelhardt, D. Luca Ghini (1490–1556): Il Padre Fondatore Della Botanica Moderna Nel Contesto dei Rapporti Scientifici Europei del Sedicesimo Secolo; Museo: New York, NY, USA, 2011; Volume 27. [Google Scholar]
- Bellorini, C. The World of Plants in Renaissance Tuscany; Routledge: London, UK, 2016. [Google Scholar] [CrossRef]
- Cristofolini, G.; Mossetti, U. Pre-Linnean herbaria in Bologna: Some Newly Discovered Collections from the Time of Ulisse Aldrovandi. 1993. Available online: https://www.researchgate.net/publication/260179625 (accessed on 25 August 2023).
- Nepi, C. La “slegatura” dell’erbario di Andrea Cesalpino (1525–1603). Museol. Sci. Nuova Ser. 2007, 1, 50–54. [Google Scholar]
- Signorini, M.A. Sulle Piante Dipinte Dal Bachiacca Nello Scrittoio Di Cosimo I A Palazzo Vecchi. 1993. Available online: https://www.jstor.org/stable/27654359 (accessed on 25 August 2023).
- Stefanaki, A.; Porck, H.; Grimaldi, I.M.; Thurn, N.; Pugliano, V.; Kardinaal, A.; Salemink, J.; Thijsse, G.; Chavannes-Mazel, C.; Kwakkel, E.; et al. Breaking the silence of the 500-year-old smiling garden of everlasting flowers: The En Tibi book herbarium. PLoS ONE 2019, 14, e0217779. [Google Scholar] [CrossRef]
- Stearn, W.T. A New Photographic Record of the Linnaean Herbarium. Taxon 1961, 10, 16–19. [Google Scholar] [CrossRef]
- Müller-Wille, S. Linnaeus’ herbarium cabinet: A piece of furniture and its function. Endeavour 2006, 30, 60–64. [Google Scholar] [CrossRef]
- Thiers, B. The World’s Herbaria 2020: A Summary Report Based on Data from Index Herbariorum. 2021. Available online: http://sweetgum.nybg.org/science/ih/ (accessed on 25 August 2023).
- Thiers, B.M.; Tulig, M.C.; Watson, K.A. Digitization of The New York Botanical Garden Herbarium. Brittonia 2016, 68, 324–333. [Google Scholar] [CrossRef]
- Heberling, J.M.; Prather, L.A.; Tonsor, S.J. The Changing Uses of Herbarium Data in an Era of Global Change: An Overview Using Automated Content Analysis. BioScience 2019, 69, 812–822. [Google Scholar] [CrossRef]
- Nelson, G.; Paul, D.; Riccardi, G.; Mast, A. Five task clusters that enable efficient and effective digitization of biological collections. ZooKeys 2012, 209, 19–45. [Google Scholar] [CrossRef] [PubMed]
- Pont, C.; Wagner, S.; Kremer, A.; Orlando, L.; Plomion, C.; Salse, J. Paleogenomics: Reconstruction of plant evolutionary trajectories from modern and ancient DNA. Genome Biol. 2019, 20, 29. [Google Scholar] [CrossRef] [PubMed]
- Saville, A.C.; Martin, M.D.; Ristaino, J.B. Historic Historic late blight outbreaks caused by a widespread dominant lineage of Phytophthora infestans (Mont.) de Bary. PLoS ONE 2016, 11, e0168381. [Google Scholar] [CrossRef] [PubMed]
- Yoshida, K.; Schuenemann, V.J.; Cano, L.M.; Pais, M.; Mishra, B.; Sharma, R.; Lanz, C.; Martin, F.N.; Kamoun, S.; Krause, J.; et al. The rise and fall of the Phytophthora infestans lineage that triggered the Irish potato famine. eLife 2013, 2, e00731. [Google Scholar] [CrossRef]
- Muñoz-Rodríguez, P.; Carruthers, T.; Wood, J.R.I.; Williams, B.R.M.; Weitemier, K.; Kronmiller, B.; Goodwin, Z.; Sumadijaya, A.; Anglin, N.L.; Filer, D.; et al. A taxonomic monograph of Ipomoea integrated across phylogenetic scales. Nat. Plants 2019, 5, 1136–1144. [Google Scholar] [CrossRef]
- Bellucci, E.; Benazzo, A.; Xu, C.; Bitocchi, E.; Rodriguez, M.; Alseekh, S.; Di Vittori, V.; Gioia, T.; Neumann, K.; Cortinovis, G.; et al. Selection and adaptive introgression guided the complex evolutionary history of the European common bean. Nat. Commun. 2023, 14, 1908. [Google Scholar] [CrossRef]
- Cortinovis, G.; Di Vittori, V.; Bellucci, E.; Bitocchi, E.; Papa, R. Adaptation to novel environments during crop diversification. Curr. Opin. Plant Biol. 2020, 56, 203–217. [Google Scholar] [CrossRef]
- Damerval, C.; Ben Othman, W.; Manicacci, D.; Jabbour, F. Distribution area of the two floral morphs of Nigella damascena L. (Ranunculaceae): A diachronic study using herbarium specimens collected in France. Bot. Lett. 2018, 165, 396–403. [Google Scholar] [CrossRef]
- Davis, C.C.; Willis, C.G.; Connolly, B.; Kelly, C.; Ellison, A.M. Herbarium records are reliable sources of phenological change driven by climate and provide novel insights into species’ phenological cueing mechanisms. Am. J. Bot. 2015, 102, 1599–1609. [Google Scholar] [CrossRef]
- Myers, J.R.; Formiga, A.K.; Janick, J. Iconography of Beans and Related Legumes Following the Columbian Exchange. Front. Plant Sci. 2022, 13, 851029. [Google Scholar] [CrossRef]
- Bakker, F.T. Herbarium Genomics: Plant Archival DNA Explored. In Paleogenomics. Population Genomics; Springer: Cham, Switzerland, 2018; pp. 205–224. [Google Scholar] [CrossRef]
- Gutaker, R.M.; Burbano, H.A. Reinforcing plant evolutionary genomics using ancient DNA. Curr. Opin. Plant Biol. 2017, 36, 38–45. [Google Scholar] [CrossRef] [PubMed]
- Rosche, C.; Baasch, A.; Runge, K.; Brade, P.; Träger, S.; Parisod, C.; Hensen, I. Tracking population genetic signatures of local extinction with herbarium specimens. Ann. Bot. 2022, 129, 857–868. [Google Scholar] [CrossRef] [PubMed]
- Cozzolino, S.; Cafasso, D.; Pellegrino, G.; Musacchio, A.; Widmer, A. Genetic variation in time and space: The use of herbarium specimens to reconstruct patterns of genetic variation in the endangered orchid Anacamptis palustris. Conserv. Genet. 2006, 8, 629–639. [Google Scholar] [CrossRef]
- Bellucci, E.; Mario Aguilar, O.; Alseekh, S.; Bett, K.; Brezeanu, C.; Cook, D.; De la Rosa, L.; Delledonne, M.; Dostatny, D.F.; Ferreira, J.J.; et al. The INCREASE project: Intelligent Collections of food-legume genetic resources for European agrofood systems. Plant J. 2021, 108, 646–660. [Google Scholar] [CrossRef]
- Rocchetti, L.; Gioia, T.; Logozzo, G.; Brezeanu, C.; Pereira, L.G.; De la Rosa, L.; Marzario, S.; Pieri, A.; Fernie, A.R.; Alseekh, S.; et al. Towards the development, maintenance and standardized phenotypic characterization of single-seed-descent genetic re-sources for chickpea. Curr. Protoc. 2022, 2, e371. [Google Scholar] [CrossRef]
- Cortinovis, G.; Oppermann, M.; Neumann, K.; Graner, A.; Gioia, T.; Marsella, M.; Alseekh, S.; Fernie, A.R.; Papa, R.; Bellucci, E.; et al. Towards the Development, Maintenance, and Standardized Phenotypic Characterization of Single-Seed-Descent Genetic Resources for Common Bean. Curr. Protoc. 2021, 1, e133. [Google Scholar] [CrossRef]
- Kroc, M.; Tomaszewska, M.; Czepiel, K.; Bitocchi, E.; Oppermann, M.; Neumann, K.; Guasch, L.; Bellucci, E.; Alseekh, S.; Graner, A.; et al. Towards Development, Maintenance, and Standardized Phenotypic Characterization of Single-Seed-Descent Genetic Resources for Lupins. Curr. Protoc. 2021, 1, e191. [Google Scholar] [CrossRef]
- Guerra-García, A.; Gioia, T.; von Wettberg, E.; Logozzo, G.; Papa, R.; Bitocchi, E.; Bett, K.E. Intelligent characterization of lentil genetic resources: Evolutionary history, genetic diversity of germplasm, and the need for well-represented collections. Curr. Protoc. 2021, 1, e134. [Google Scholar] [CrossRef] [PubMed]
- Pääbo, S.; Poinar, H.; Serre, D.; Jaenicke-Després, V.; Hebler, J.; Rohland, N.; Kuch, M.; Krause, J.; Vigilant, L.; Hofreiter, M. Genetic Analyses from Ancient DNA. Annu. Rev. Genet. 2004, 38, 645–679. [Google Scholar] [CrossRef]
- Staats, M.; Cuenca, A.; Richardson, J.E.; Ginkel, R.V.-V.; Petersen, G.; Seberg, O.; Bakker, F.T. DNA Damage in Plant Herbarium Tissue. PLoS ONE 2011, 6, e28448. [Google Scholar] [CrossRef]
- Särkinen, T.; Staats, M.; Richardson, J.E.; Cowan, R.S.; Bakker, F.T. How to Open the Treasure Chest? Optimising DNA Extraction from Herbarium Specimens. PLoS ONE 2012, 7, e43808. [Google Scholar] [CrossRef] [PubMed]
- Bieker, V.C.; Barreiro, F.S.; Rasmussen, J.A.; Brunier, M.; Wales, N.; Martin, M.D. Metagenomic analysis of historical herbarium specimens reveals a postmortem microbial community. Mol. Ecol. Resour. 2020, 20, 1206–1219. [Google Scholar] [CrossRef] [PubMed]
- Kistler, L.; Bieker, V.C.; Martin, M.D.; Pedersen, M.W.; Madrigal, J.R.; Wales, N. Ancient Plant Genomics in Archaeology, Herbaria, and the Environment. Annu. Rev. Plant Biol. 2020, 71, 605–629. [Google Scholar] [CrossRef] [PubMed]
- Orlando, L.; Allaby, R.; Skoglund, P.; Der Sarkissian, C.; Stockhammer, P.W.; Ávila-Arcos, M.C.; Fu, Q.; Krause, J.; Willerslev, E.; Stone, A.C.; et al. Ancient DNA analysis. Nat. Rev. Methods Prim. 2021, 1, 14. [Google Scholar] [CrossRef]
- Weiß, C.L.; Schuenemann, V.J.; Devos, J.; Shirsekar, G.; Reiter, E.; Gould, B.A.; Stinchcombe, J.R.; Krause, J.; Burbano, H.A. Temporal patterns of damage and decay kinetics of DNA retrieved from plant herbarium specimens. R. Soc. Open Sci. 2016, 3, 160239. [Google Scholar] [CrossRef]
- Drábková, L.; Kirschner, J.; Vlĉek, Ĉ. Comparison of Seven DNA Extraction and Amplification Protocols in Historical Herbarium Specimens of Juncaceae. Plant Mol. Biol. Rep. 2002, 20, 161–175. [Google Scholar] [CrossRef]
- Inglis, P.W.; Pappas, M.D.C.R.; Resende, L.V.; Grattapaglia, D. Fast and inexpensive protocols for consistent extraction of high quality DNA and RNA from challenging plant and fungal samples for high-throughput SNP genotyping and sequencing applications. PLoS ONE 2018, 13, e0206085. [Google Scholar] [CrossRef]
- Rogers, S.O.; Bendich, A.J. Extraction of DNA from milligram amounts of fresh, herbarium and mummified plant tissues. Plant Mol. Biol. 1985, 5, 69–76. [Google Scholar] [CrossRef]
- Cota-Sánchez, J.H.; Remarchuk, K.; Ubayasena, K. Ready-to-use DNA extracted with a CTAB method adapted for herbarium specimens and mucilaginous plant tissue. Plant Mol. Biol. Rep. 2006, 24, 161–167. [Google Scholar] [CrossRef]
- Riahi, M.; Zarre, S.; Maassoumi, A.; Attar, F.; Osaloo, S.K. Methodology An inexpensive and rapid method for extracting papilionoid genomic DNA from herbarium specimens. Evolution 2010, 9, 1334–1342. [Google Scholar] [CrossRef]
- Sugita, N.; Ebihara, A.; Hosoya, T.; Jinbo, U.; Kaneko, S.; Kurosawa, T.; Nakae, M.; Yukawa, T. Non-destructive DNA extraction from herbarium specimens: A method particularly suitable for plants with small and fragile leaves. J. Plant Res. 2019, 133, 133–141. [Google Scholar] [CrossRef] [PubMed]
- Gutaker, R.M.; Reiter, E.; Furtwängler, A.; Schuenemann, V.J.; Burbano, H.A. Extraction of ultrashort DNA molecules from herbarium specimens. BioTechniques 2017, 62, 76–79. [Google Scholar] [CrossRef] [PubMed]
- Wales, N.; Kistler, L. Extraction of Ancient DNA from Plant Remains. In Ancient DNA: Methods and Protocols; Shapiro, B., Barlow, A., Heintzman, P., Hofreiter, M., Paijmans, J., Soares, A., Eds.; Methods in Molecular Biology; Humana Press: New York, NY, USA, 2019; Volume 1963. [Google Scholar] [CrossRef]
- Ribeiro, R.; Lovato, M. DNA extraction in fresh and herbarium specimens of Dalbergia Comparative analysis of different DNA extraction protocols in fresh and herbarium specimens of the genus Dalbergia. Genet. Mol. Res. 2007, 6, 173–187. [Google Scholar]
- Krinitsina, A.A.; Sizova, T.V.; Zaika, M.A.; Speranskaya, A.S.; Sukhorukov, A.P. A rapid and cost-effective method for DNA extraction from archival herbarium specimens. Biochemistry 2015, 80, 1478–1484. [Google Scholar] [CrossRef] [PubMed]
- Wang, W. A primer to the use of herbarium specimens in plant phylogenetics. Bot. Lett. 2018, 165, 404–408. [Google Scholar] [CrossRef]
- Doyle, J.J.; Dickson, E.E. Preservation of Plant Samples for DNA Restriction Endonuclease Analysis. Taxon 1987, 36, 715–722. [Google Scholar] [CrossRef]
- Jankowiak, K.; Buczkowska, K.; Szweykowska-Kulinska, Z. Successful extraction of DNA from 100-year-old herbarium specimens of the liverwort Bazzania trilobata. Taxon 2005, 54, 335–336. [Google Scholar] [CrossRef]
- Savolainen, V.; Cuénoud, P.; Spichiger, R.; Martinez, M.D.; Crèvecoeur, M.; Manen, J.F. Systematics and Evolution The use of herbarium specimens in DNA phylogenetics: Evaluation and improvement. Plant Syst. Evol. 1995, 197, 87–98. [Google Scholar] [CrossRef]
- Drábková, L.Z. DNA Extraction from Herbarium Specimens. Methods Mol. Biol. 2013, 1115, 69–84. [Google Scholar] [CrossRef]
- Freedman, J.; Dorp, V.; Brace, L. Destructive Sampling Natural Science Collections: An Overview for Museum Professionals and Researchers. 2018. Available online: http://www.natsca.org/article/2440 (accessed on 25 August 2023).
- Shepherd, L.D. A non-destructive DNA sampling technique for herbarium specimens. PLoS ONE 2017, 12, e0183555. [Google Scholar] [CrossRef]
- Gilbert, M.T.P.; Moore, W.; Melchior, L.; Worobey, M. DNA extraction from dry museum beetles without conferring external morphological damage. PLoS ONE 2007, 2, e272. [Google Scholar] [CrossRef] [PubMed]
- Santos, D.; Ribeiro, G.C.; Cabral, A.D.; Sperança, M.A. A non-destructive enzymatic method to extract DNA from arthropod specimens: Implications for morphological and molecular studies. PLoS ONE 2018, 13, e0192200. [Google Scholar] [CrossRef] [PubMed]
- Zeng, C.-X.; Hollingsworth, P.M.; Yang, J.; He, Z.-S.; Zhang, Z.-R.; Li, D.-Z.; Yang, J.-B. Genome skimming herbarium specimens for DNA barcoding and phylogenomics. Plant Methods 2018, 14, 43. [Google Scholar] [CrossRef] [PubMed]
- Marinček, P.; Wagner, N.D.; Tomasello, S. Ancient DNA extraction methods for herbarium specimens: When is it worth the effort? Appl. Plant Sci. 2022, 10, e11477. [Google Scholar] [CrossRef]
- Jobes, D.V.; Hurley, D.L.; Thien, L.B. Plant DNA Isolation: A Method to Efficiently Remove Polyphenolics, Polysaccharides, and RNA. Taxon 1995, 44, 379–386. [Google Scholar] [CrossRef]
- Hofreiter, M. Ancient DNA: Methods and Protocols; Shapiro, B., Barlow, A., Heintzman, P.D., Hofreiter, M., Paijmans, J.L.A., Soares, A.E.R., Eds.; Humana Press Incorporated: Totowa, NJ, USA, 2012; Volume 840. [Google Scholar]
- Thomas, M.; Gilbert, P.; Wilson, A.S.; Bunce, M.; Hansen, A.J.; Willerslev, E.; Shapiro, B.; Higham, T.F.G.; Richards, M.P.; O’connell, T.C.; et al. Ancient mito-chondrial DNA from hair. Curr. Biol. 2004, 14, R463–R464. [Google Scholar]
- Khanuja, S.P.; Shasany, A.K.; Darokar, M.; Kumar, S. Rapid Isolation of DNA from Dry and Fresh Samples of Plants Producing Large Amounts of Secondary Metabolites and Essential Oils. Plant Mol. Biol. Rep. 1999, 17, 74. [Google Scholar] [CrossRef]
- Lister, D.L.; Bower, M.A.; Howe, C.J.; Jones, M.K. Extraction and amplification of nuclear DNA from herbarium specimens of emmer wheat: A method for assessing DNA preservation by maximum amplicon length recovery. Taxon 2008, 57, 254–258. [Google Scholar] [CrossRef]
- Samarakoon, T.; Wang, S.Y.; Alford, M.H. Enhancing PCR Amplification of DNA from Recalcitrant Plant Specimens Using a Trehalose-Based Additive. Appl. Plant Sci. 2013, 1, 1200236. [Google Scholar] [CrossRef]
- Telle, S.; Thines, M. Amplification of cox2 (~620 bp) from 2 mg of up to 129 Years Old Herbarium Specimens, Comparing 19 Extraction Methods and 15 Polymerases. PLoS ONE 2008, 3, e3584. [Google Scholar] [CrossRef]
- Monroe, C.; Grier, C.; Kemp, B.M. Evaluating the efficacy of various thermo-stable polymerases against co-extracted PCR inhibitors in ancient DNA samples. Forensic Sci. Int. 2013, 228, 142–153. [Google Scholar] [CrossRef] [PubMed]
- Burrell, A.S.; Disotell, T.R.; Bergey, C.M. The use of museum specimens with high-throughput DNA sequencers. J. Hum. Evol. 2015, 79, 35–44. [Google Scholar] [CrossRef] [PubMed]
- Binladen, J.; Willerslev, E. Why study ancient DNA damage? J. Nord. Archaeol. Sci. 2010, 17, a012567. [Google Scholar]
- Pääbo, S.; Irwin, D.M.; Wilson, A.C. DNA damage promotes jumping between templates during enzymatic amplification. J. Biol. Chem. 1990, 265, 4718–4721. [Google Scholar] [CrossRef]
- Willerslev, E.; Cooper, A. Ancient DNA. Proc. R. Soc. B Boil. Sci. 2004, 272, 3–16. [Google Scholar] [CrossRef]
- Hofreiter, M. DNA sequences from multiple amplifications reveal artifacts induced by cytosine deamination in ancient DNA. Nucleic Acids Res. 2001, 29, 4793–4799. [Google Scholar] [CrossRef]
- Pruvost, M.; Grange, T.; Geigl, E.-M. Minimizing DNA contamination by using UNG-coupled quantitative real-time PCR on degraded DNA samples: Application to ancient DNA studies. BioTechniques 2005, 38, 569–575. [Google Scholar] [CrossRef] [PubMed]
- Champlot, S.; Berthelot, C.; Pruvost, M.; Bennett, E.A.; Grange, T.; Geigl, E.-M. An Efficient Multistrategy DNA Decontamination Procedure of PCR Reagents for Hypersensitive PCR Applications. PLoS ONE 2010, 5, e13042. [Google Scholar] [CrossRef]
- Knapp, M.; Clarke, A.C.; Horsburgh, K.A.; Matisoo-Smith, E.A. Setting the stage—Building and working in an ancient DNA laboratory. Ann. Anat.—Anat. Anz. 2011, 194, 3–6. [Google Scholar] [CrossRef]
- Beck, J.B.; Semple, J.C. Next-Generation Sampling: Pairing Genomics with Herbarium Specimens Provides Species-Level Signal in Solidago (Asteraceae). Appl. Plant Sci. 2015, 3, 1500014. [Google Scholar] [CrossRef]
- Hart, M.L.; Forrest, L.L.; Nicholls, J.A.; Kidner, C.A. Retrieval of hundreds of nuclear loci from herbarium specimens. Taxon 2016, 65, 1081–1092. [Google Scholar] [CrossRef]
- Rowe, K.C.; Singhal, S.; Macmanes, M.D.; Ayroles, J.F.; Morelli, T.L.; Rubidge, E.M.; Bi, K.; Moritz, C.C. Museum genomics: Low-cost and high-accuracy genetic data from historical specimens. Mol. Ecol. Resour. 2011, 11, 1082–1092. [Google Scholar] [CrossRef]
- Malenica, N.; Šimon, S.; Besendorfer, V.; Maletić, E.; Kontić, J.K.; Pejić, I. Whole genome amplification and microsatellite genotyping of herbarium DNA revealed the identity of an ancient grapevine cultivar. Sci. Nat. 2011, 98, 763–772. [Google Scholar] [CrossRef] [PubMed]
- Lambertini, C.; Frydenberg, J.; Gustafsson, M.H.G.; Brix, H. Herbarium specimens as a source of DNA for AFLP fingerprinting of Phragmites (Poaceae): Possibilities and limitations. Plant Syst. Evol. 2008, 272, 223–231. [Google Scholar] [CrossRef][Green Version]
- Roullier, C.; Benoit, L.; Mckey, D.B.; Lebot, V. Historical collections reveal patterns of diffusion of sweet potato in Oceania obscured by modern plant movements and recombination. Proc. Natl. Acad. Sci. USA 2013, 110, 2205–2210. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Li, Y.; Li, S.; Hu, N.; He, Y.; Pong, R.; Lin, D.; Lu, L.; Law, M. Comparison of next-generation sequencing systems. J. Biomed. Biotechnol. 2012, 2012, 251364. [Google Scholar] [CrossRef] [PubMed]
- Gansauge, M.-T.; Meyer, M. Single-stranded DNA library preparation for the sequencing of ancient or damaged DNA. Nat. Protoc. 2013, 8, 737–748. [Google Scholar] [CrossRef] [PubMed]
- Bennett, E.A.; Massilani, D.; Lizzo, G.; Daligault, J.; Geigl, E.-M.; Grange, T. Library construction for ancient genomics: Single strand or double strand? BioTechniques 2014, 56, 289–300. [Google Scholar] [CrossRef]
- Psonis, N.; Vassou, D.; Kafetzopoulos, D. Testing a series of modifications on genomic library preparation methods for ancient or degraded DNA. Anal. Biochem. 2021, 623, 114193. [Google Scholar] [CrossRef] [PubMed]
- Briggs, A.W.; Heyn, P. Preparation of Next-Generation Sequencing Libraries from Damaged DNA. Methods Mol. Biol. 2011, 840, 143–154. [Google Scholar] [CrossRef]
- Prüfer, K.; Stenzel, U.; Hofreiter, M.; Pääbo, S.; Kelso, J.; Green, R.E. Open Access METHOD Computational challenges in the analysis of ancient DNA. Genome Biol. 2010, 11. [Google Scholar] [CrossRef] [PubMed]
- Ginolhac, A.; Rasmussen, M.; Gilbert, M.T.P.; Willerslev, E.; Orlando, L. mapDamage: Testing for damage patterns in ancient DNA sequences. Bioinformatics 2011, 27, 2153–2155. [Google Scholar] [CrossRef] [PubMed]
- Jónsson, H.; Ginolhac, A.; Schubert, M.; Johnson, P.L.F.; Orlando, L. mapDamage2.0: Fast approximate Bayesian estimates of ancient DNA damage parameters. Bioinformatics 2013, 29, 1682–1684. [Google Scholar] [CrossRef] [PubMed]
- Trucchi, E.; Benazzo, A.; Lari, M.; Iob, A.; Vai, S.; Nanni, L.; Bellucci, E.; Bitocchi, E.; Raffini, F.; Xu, C.; et al. Ancient genomes reveal early Andean farmers selected common beans while preserving diversity. Nat. Plants 2021, 7, 123–128. [Google Scholar] [CrossRef]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Papalini, S.; Di Vittori, V.; Pieri, A.; Allegrezza, M.; Frascarelli, G.; Nanni, L.; Bitocchi, E.; Bellucci, E.; Gioia, T.; Pereira, L.G.; et al. Challenges and Opportunities behind the Use of Herbaria in Paleogenomics Studies. Plants 2023, 12, 3452. https://doi.org/10.3390/plants12193452
Papalini S, Di Vittori V, Pieri A, Allegrezza M, Frascarelli G, Nanni L, Bitocchi E, Bellucci E, Gioia T, Pereira LG, et al. Challenges and Opportunities behind the Use of Herbaria in Paleogenomics Studies. Plants. 2023; 12(19):3452. https://doi.org/10.3390/plants12193452
Chicago/Turabian StylePapalini, Simone, Valerio Di Vittori, Alice Pieri, Marina Allegrezza, Giulia Frascarelli, Laura Nanni, Elena Bitocchi, Elisa Bellucci, Tania Gioia, Luis Guasch Pereira, and et al. 2023. "Challenges and Opportunities behind the Use of Herbaria in Paleogenomics Studies" Plants 12, no. 19: 3452. https://doi.org/10.3390/plants12193452
APA StylePapalini, S., Di Vittori, V., Pieri, A., Allegrezza, M., Frascarelli, G., Nanni, L., Bitocchi, E., Bellucci, E., Gioia, T., Pereira, L. G., Susek, K., Tenaillon, M., Neumann, K., & Papa, R. (2023). Challenges and Opportunities behind the Use of Herbaria in Paleogenomics Studies. Plants, 12(19), 3452. https://doi.org/10.3390/plants12193452