Comparison of Policosanol Profiles of the Sprouts of Wheat Mutant Lines and the Effect of Differential LED Lights on Selected Lines

 , , ,
, , ,  , and
, and
Abstract
:1. Introduction
2. Results and Discussion
2.1. Identification of Policosanols in the Wheat Sprout Samples Using GC–MS Analysis
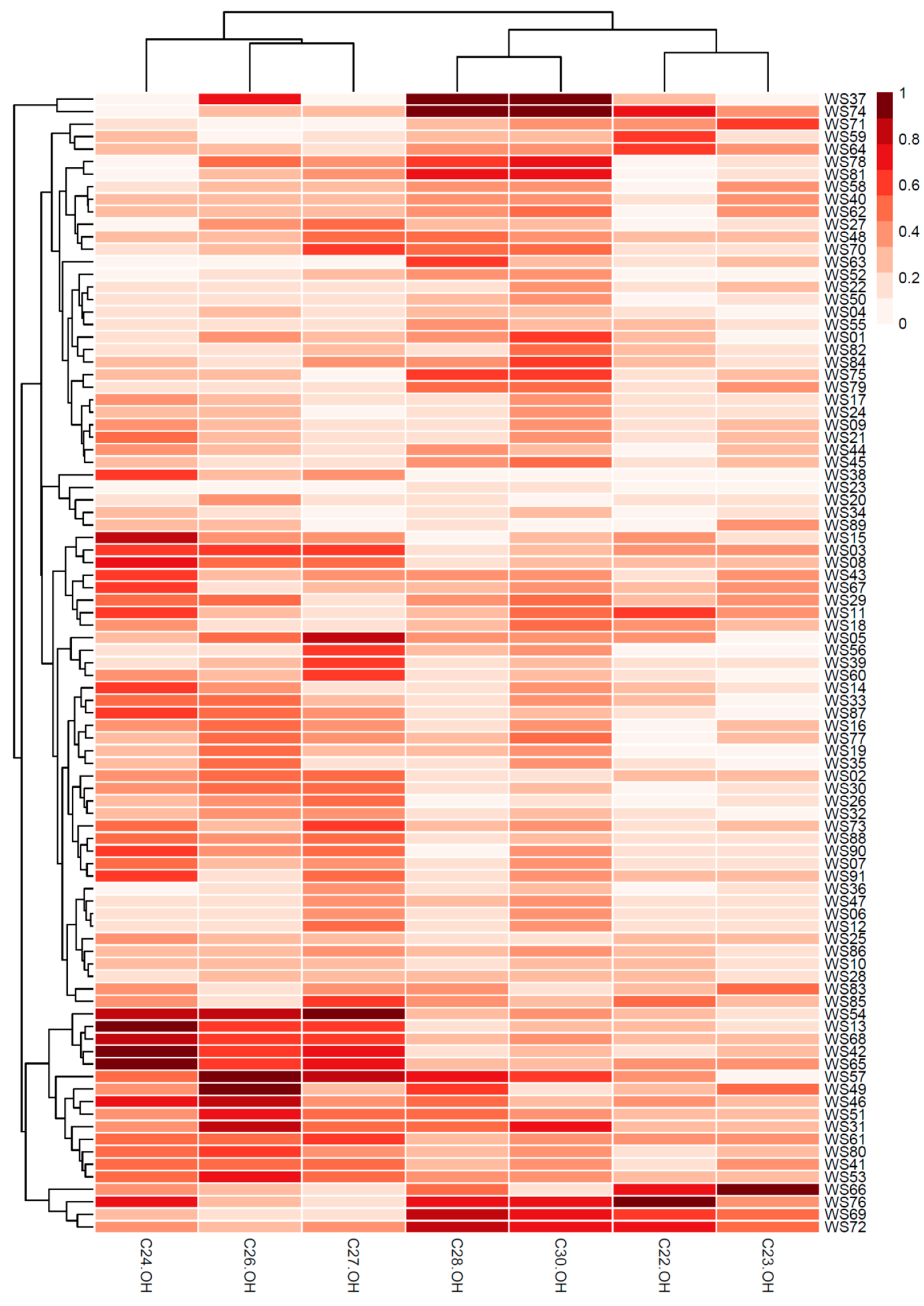
2.2. Comparison of Policosanol Contents in Different Wheat Sprout Samples
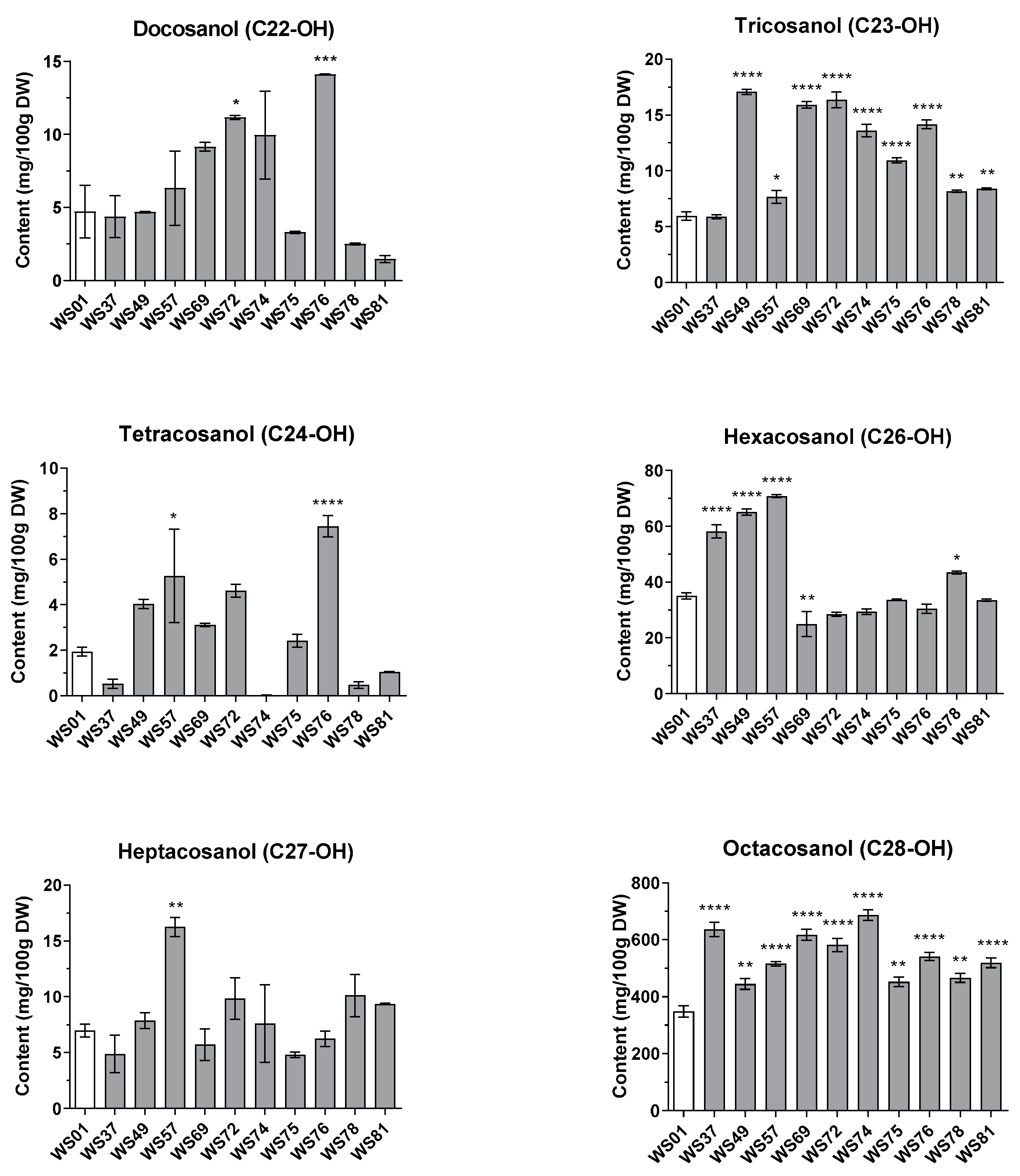
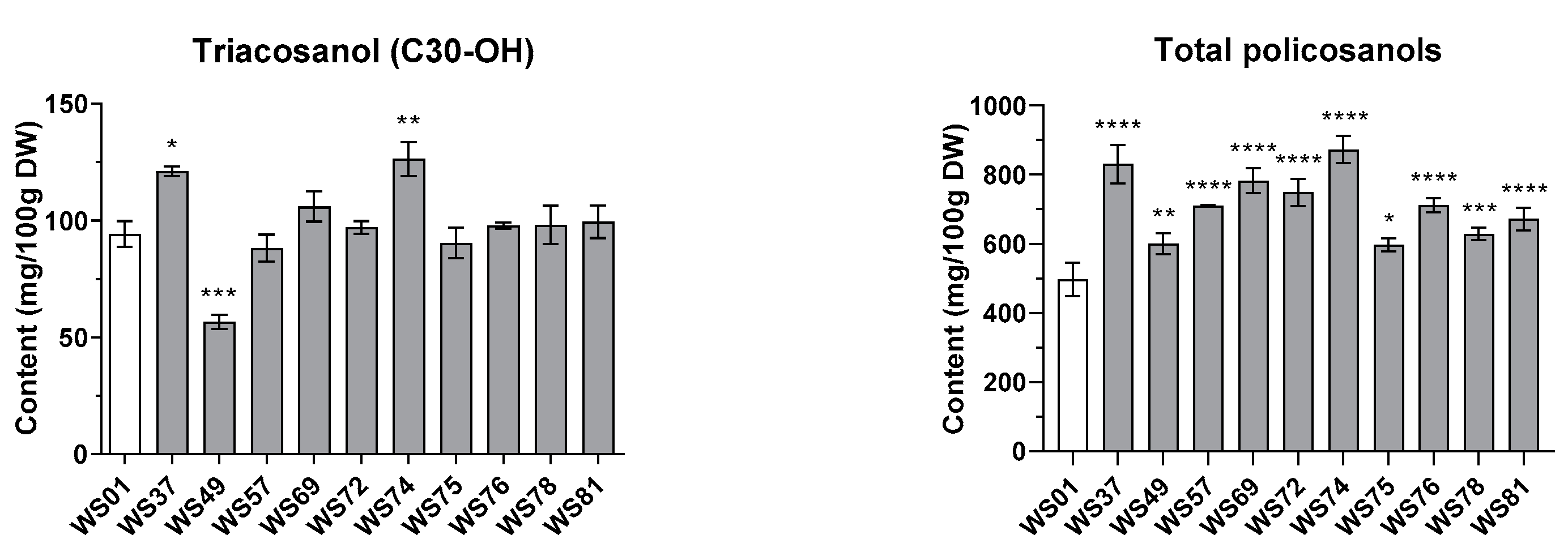
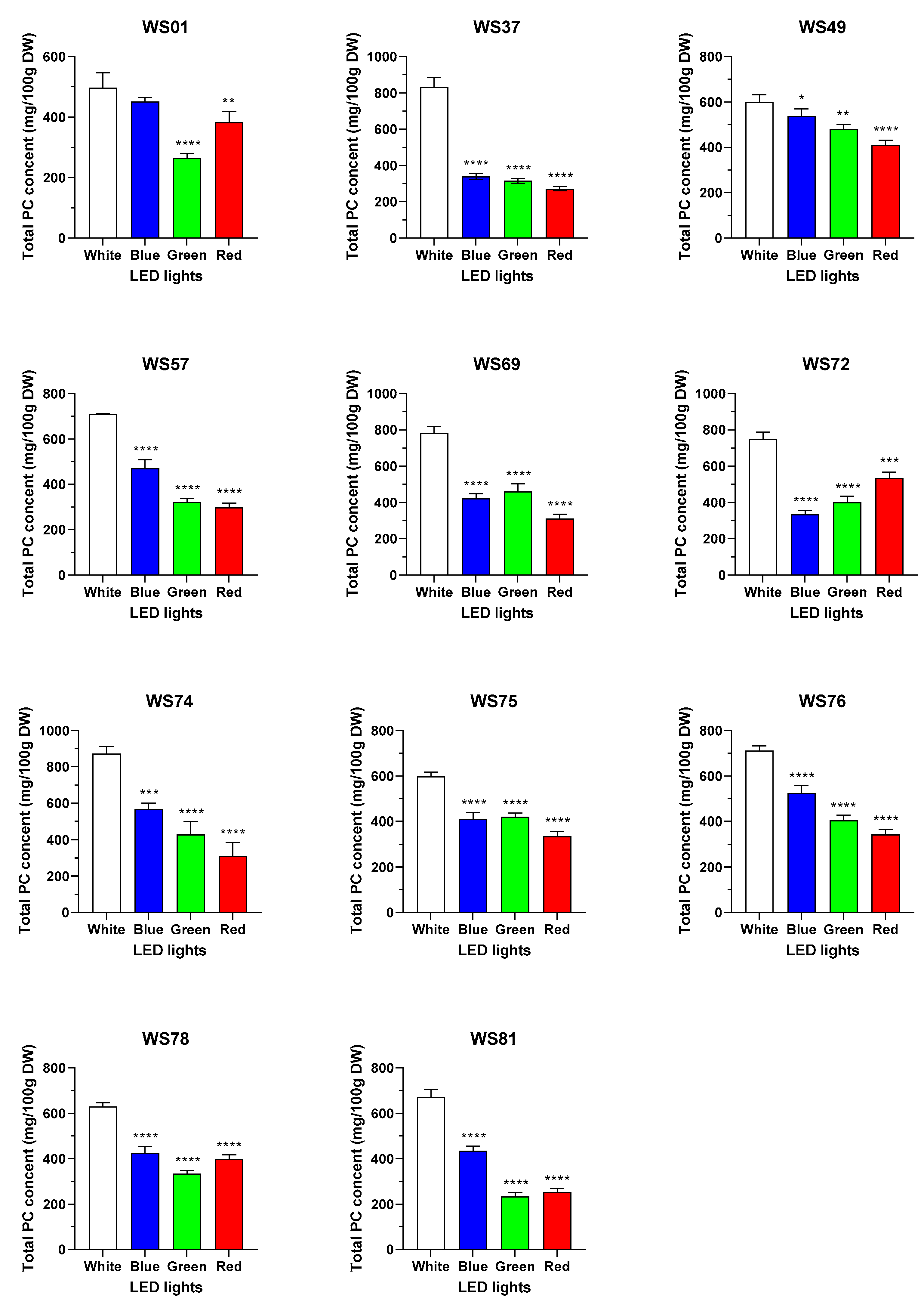
2.3. Effects of Different LED Conditions on the Policosanol Content in the Sprouts of Wheat Mutant Lines
3. Discussions
4. Materials and Methods
4.1. Plant Materials
4.2. Sample Preparation
4.3. GC-MS Analysis
4.4. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Shewry, P.R.; Hey, S.J. The contribution of wheat to human diet and health. Food Energy Secur. 2015, 4, 178–202. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.; Sang, S. Phytochemicals in whole grain wheat and their health-promoting effects. Mol. Nutr. Food Res. 2017, 61, 1600852. [Google Scholar] [CrossRef] [PubMed]
- Geng, J.; Li, J.; Zhu, F.; Chen, X.; Du, B.; Tian, H.; Li, J. Plant sprout foods: Biological activities, health benefits, and bioavailability. J. Food Biochem. 2022, 46, e13777. [Google Scholar] [CrossRef] [PubMed]
- Miyahira, R.F.; Lopes, J.O.; Antunes, A.E.C. The use of sprouts to improve the nutritional value of food products: A brief review. Plant Foods Hum. Nutr. 2021, 76, 143–152. [Google Scholar] [CrossRef] [PubMed]
- Han, A.; Hong, M.J.; Nam, B.; Kim, B.; Park, H.H.; Baek, I.; Kil, Y.; Nam, J.; Jin, C.H.; Kim, J. Comparison of Flavonoid Profiles in Sprouts of Radiation Breeding Wheat Lines (Triticum aestivum L.). Agronomy 2020, 10, 1489. [Google Scholar] [CrossRef]
- Ra, J.E.; Woo, S.Y.; Lee, K.S.; Lee, M.J.; Kim, H.Y.; Ham, H.M.; Chung, I.M.; Kim, D.H.; Lee, J.H.; Seo, W.D. Policosanol profiles and adenosine 5’-monophosphate-activated protein kinase (AMPK) activation potential of Korean wheat seedling extracts according to cultivar and growth time. Food Chem. 2020, 317, 126388. [Google Scholar] [CrossRef]
- Muthusamy, M.; Kim, J.H.; Kim, S.H.; Kim, J.Y.; Heo, J.W.; Lee, H.; Lee, K.S.; Seo, W.D.; Park, S.; Kim, J.A.; et al. Changes in Beneficial C-glycosylflavones and Policosanol Content in Wheat and Barley Sprouts Subjected to Differential LED Light Conditions. Plants 2020, 9, 1502. [Google Scholar] [CrossRef]
- Yadav, A.; Singh, S. Effect of exogenous phytohormone treatment on antioxidant activity, enzyme activity and phenolic content in wheat sprouts and identification of metabolites of control and treated samples by UHPLC-MS analysis. Food Res. Int. 2023, 169, 112811. [Google Scholar] [CrossRef]
- Bonfili, L.; Amici, M.; Cecarini, V.; Cuccioloni, M.; Tacconi, R.; Angeletti, M.; Fioretti, E.; Keller, J.N.; Eleuteri, A.M. Wheat sprout extract-induced apoptosis in human cancer cells by proteasomes modulation. Biochimie 2009, 91, 1131–1144. [Google Scholar] [CrossRef]
- Peryt, B.; Szymczyk, T.; Lesca, P. Mechanism of antimutagenicity of wheat sprout extracts. Mutat. Res. 1992, 269, 201–215. [Google Scholar] [CrossRef]
- Han, B.K.; Yoon, H.; Kim, K.H.; Shin, E.C.; Ko, K.S.; Lee, H.S.; Kim, Y.J. Inhibitory effects of wheat sprouts extract on RANKL-induced osteoclast differentiation via suppressing MAPK and NFATc1 signaling pathways. J. Med. Food 2023, 26, 480–488. [Google Scholar] [CrossRef] [PubMed]
- Lim, J.Y.; Yun, D.H.; Lee, J.H.; Kwon, Y.B.; Lee, Y.M.; Lee, D.H.; Kim, D.K. Extract of Triticum aestivum sprouts suppresses acetaminophen-induced hepatotoxicity in mice by inhibiting oxidative stress. Molecules 2021, 26, 6336. [Google Scholar] [CrossRef] [PubMed]
- Brighenti, V.; Venturelli, A.; Caroli, C.; Anceschi, L.; Gjikolaj, M.; Durante, C.; Pellati, F. An innovative method for the extraction and HPLC analysis of bioactive policosanols from non-psychoactive Cannabis sativa L. J. Pharm. Biomed. Anal. 2023, 234, 115547. [Google Scholar] [CrossRef] [PubMed]
- Safari, S.; Mirazi, N.; Ahmadi, N.; Asadbegi, M.; Nourian, A.; Rashno, M.; Komaki, A. Policosanol protects against Alzheimer’s disease-associated spatial cognitive decline in male rats: Possible involved mechanisms. Psychopharmacol 2023, 240, 755–767. [Google Scholar] [CrossRef] [PubMed]
- Ma, C.; Feng, Y.; Li, X.; Sun, L.; He, Z.; Gan, J.; He, M.; Zhang, X.; Chen, X. Potential therapeutic effects of policosanol from insect wax on Caenorhabditis elegans Models of Parkinson’s disease. J. Neuroimmune Pharmacol. 2023, 18, 127–144. [Google Scholar] [CrossRef]
- Sun, L.; Li, X.; Ma, C.; He, Z.; Zhang, X.; Wang, C.; Zhao, M.; Gan, J.; Feng, Y. Improving Effect of the policosanol from Ericerus pela Wax on Learning and Memory Impairment Caused by scopolamine in Mice. Foods 2022, 11, 2095. [Google Scholar] [CrossRef]
- Varady, K.A.; Wang, Y.; Jones, P.J. Role of policosanols in the prevention and treatment of cardiovascular disease. Nutr. Rev. 2003, 61, 376–383. [Google Scholar] [CrossRef]
- Barbagallo, C.M.; Cefalù, A.B.; Noto, D.; Averna, M.R. Role of Nutraceuticals in Hypolipidemic Therapy. Front. Cardiovasc. Med. 2015, 2, 22. [Google Scholar] [CrossRef]
- Pirro, M.; Vetrani, C.; Bianchi, C.; Mannarino, M.R.; Bernini, F.; Rivellese, A.A. Joint position statement on "Nutraceuticals for the treatment of hypercholesterolemia" of the Italian Society of Diabetology (SID) and of the Italian Society for the Study of Arteriosclerosis (SISA). Nutr. Metab Cardiovasc. Dis. 2017, 27, 2–17. [Google Scholar] [CrossRef]
- EFSA Panel on Dietetic Products, Nutrition and Allergies (NDA). Scientific opinion on the substantiation of health claims related to policosanols from sugar cane wax and maintenance of normal blood LDL-cholesterol concentrations (ID 1747, 1748, 1864, 1951, 1954, 4693) and maintenance of normal blood HDL-cholesterol concentrations (ID 1747, 1748, 1864, 1951, 1954, 4693) pursuant to Article 13(1) of Regulation (EC) No 1924/2006. EFSA J. 2011, 9, 2255. [Google Scholar]
- Nam, D.E.; Yun, J.M.; Kim, D.; Kim, O.K. Policosanol attenuates cholesterol synthesis via AMPK activation in hypercholesterolemic rats. J. Med. Food 2019, 22, 1110–1117. [Google Scholar] [CrossRef] [PubMed]
- Zhai, Z.; Niu, K.M.; Liu, H.; Lin, C.; Tu, Y.; Liu, Y.; Cai, L.; Ouyang, K.; Liu, J. Policosanol alleviates hepatic lipid accumulation by regulating bile acids metabolism in C57BL6/mice through AMPK-FXR-TGR5 cross-talk. J. Food Sci. 2021, 86, 5466–5478. [Google Scholar] [CrossRef] [PubMed]
- Kim, K.M.; Kim, C.H.; Cho, K.H.; Jang, W.G. Policosanol attenuates Pi-induced calcification via AMPK-mediated INSIGs expression in rat VSMCs. Clin. Exp. Pharmacol. Physiol. 2021, 48, 1336–1345. [Google Scholar] [CrossRef]
- Kim, K.M.; Lim, Y.J.; Jang, W.G. Policosanol stimulates osteoblast differentiation via adenosine monophosphate-activated protein kinase-mediated expression of insulin-induced genes 1 and 2. Cells 2023, 12, 1863. [Google Scholar] [CrossRef] [PubMed]
- Seo, W.D.; Yuk, H.J.; Curtis-Long, M.J.; Jang, K.C.; Lee, J.H.; Han, S.I.; Kang, H.W.; Nam, M.H.; Lee, S.J.; Lee, J.H.; et al. Effect of the growth stage and cultivar on policosanol profiles of barley sprouts and their adenosine 5′-monophosphate-activated protein kinase activation. J. Agric. Food Chem. 2013, 61, 1117–1123. [Google Scholar] [CrossRef]
- Lee, H.G.; Woo, S.Y.; Ahn, H.J.; Yang, J.Y.; Lee, M.J.; Kim, H.Y.; Song, S.Y.; Lee, J.H.; Seo, W.D. Comparative analysis of policosanols related to growth times from the seedlings of various Korean oat (Avena sativa L.) cultivars and screening for adenosine 5′-monophosphate-activated protein kinase (AMPK) activation. Plants 2022, 11, 1844. [Google Scholar] [CrossRef]
- Thoma, F.; Somborn-Schulz, A.; Schlehuber, D.; Keuter, V.; Deerberg, G. Effects of Light on Secondary Metabolites in Selected Leafy Greens: A Review. Front. Plant Sci. 2020, 11, 497. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Wen, K.S.; Ruan, X.; Zhao, Y.X.; Wei, F.; Wang, Q. Response of Plant Secondary Metabolites to Environmental Factors. Molecules 2018, 23, 762. [Google Scholar] [CrossRef]
- Liu, K.; Harrison, M.T.; Yan, H.; Liu, D.L.; Meinke, H.; Hoogenboom, G.; Wang, B.; Peng, B.; Guan, K.; Jaegermeyr, J.; et al. Silver lining to a climate crisis in multiple prospects for alleviating crop waterlogging under future climates. Nat. Commun. 2023, 14, 765. [Google Scholar] [CrossRef]
- Alrifai, O.; Hao, X.; Marcone, M.F.; Tsao, R. Current review of the modulatory effects of LED lights on photosynthesis of secondary metabolites and future perspectives of microgreen vegetables. J. Agric. Food Chem. 2019, 67, 6075–6090. [Google Scholar] [CrossRef]
- Sathasiva, R.; Park, S.U.; Kim, J.K.; Park, Y.J.; Kim, M.C.; Nguyen, B.V.; Lee, S.Y. Metabolic profiling of primary and secondary metabolites in Kohlrabi (Brassica oleracea var. gongylodes) sprouts exposed to different light-emitting diodes. Plants 2023, 12, 1296. [Google Scholar] [CrossRef]
- Tuan, P.A.; Thwe, A.A.; Kim, Y.B.; Kim, J.K.; Kim, S.J.; Lee, S.; Chung, S.O.; Park, S.U. Effects of white, blue, and red light-emitting diodes on carotenoid biosynthetic gene expression levels and carotenoid accumulation in sprouts of tartary buckwheat (Fagopyrum tataricum gaertn.). J. Agric. Food Chem. 2013, 61, 12356–12361. [Google Scholar] [CrossRef] [PubMed]
- Dou, H.; Niu, G.; Gu, M.; Masabni, J.G. Effects of light quality on growth and phytonutrient accumulation of herbs under controlled environments. Horticulturae 2017, 3, 36. [Google Scholar] [CrossRef]
- Hasan, M.M.; Bashir, T.; Ghosh, R.; Lee, S.K.; Bae, H. An overview of LEDs’ effects on the production of bioactive compounds and crop quality. Molecules 2017, 22, 1420. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Kubota, C. Effects of supplemental light quality on growth and phytochemicals of baby leaf lettuce. Environ. Exp. Bot. 2009, 67, 59–64. [Google Scholar] [CrossRef]
- Brazaityte, A.; Sakalauskiene, S.; Samuoliene, G.; Jankauskiene, J.; Viršile, A.; Novičkovas, A.; Sirtautas, R.; Miliauskiene, J.; Vaštakaite, V.; Dabašinskas, L.; et al. The effects of LED illumination spectra and intensity on carotenoid content in Brassicaceae microgreens. Food Chem. 2015, 173, 600–606. [Google Scholar] [CrossRef]
- Samuoliene, G.; Viršile, A.; Brazaityte, A.; Jankauskiene, J.; Sakalauskiene, S.; Vaštakaite, V.; Novičkovas, A.; Viškeliene, A.; Sasnauskas, A.; Duchovskis, P. Blue light dosage affects carotenoids and tocopherols in microgreens. Food Chem. 2017, 228, 50–56. [Google Scholar] [CrossRef]
- Wang, Y.; Wang, M.; Sun, Y.; Wang, Y.; Li, T.; Chai, G.; Jiang, W.; Shan, L.; Li, C.; Xiao, E.; et al. FAR5, a fatty acyl-coenzyme A reductase, is involved in primary alcohol biosynthesis of the leaf blade cuticular wax in wheat (Triticum aestivum L.). J. Exp. Bot. 2015, 66, 1165–1178. [Google Scholar] [CrossRef]
- Wang, M.; Wu, H.; Xu, J.; Li, C.; Wang, Y.; Wang, Z. Five fatty acyl-coenzyme a reductases are involved in the biosynthesis of primary alcohols in Aegilops tauschii leaves. Front Plant. Sci. 2017, 8, 1012. [Google Scholar] [CrossRef]
- Nam, B.; So, Y.; Kim, H.; Kim, J.; Jin, C.; Han, A. A new monoterpene from the leaves of a radiation mutant cultivar of Perilla frutescens var. crispa with inhibitory activity on LPS-induced NO production. Molecules 2017, 22, 1471. [Google Scholar] [CrossRef]
- Irmak, S.; Dunford, N.T.; Milligan, J. Policosanol contents of beeswax, sugar cane and wheat extracts. Food Chem. 2006, 95, 312–318. [Google Scholar] [CrossRef]
- Wu, T.-T.; Charles, A.L.; Huang, T.-C. Determination of the contents of the main biochemical compounds of Adlay (Coxi lachrymal-jobi). Food Chem. 2007, 104, 1509–1515. [Google Scholar] [CrossRef]
- Harrabi, S.; Boukhchina, S.; Mayer, P.M.; Kallel, H. Policosanol distribution and accumulation in developing corn kernels. Food Chem. 2009, 115, 918–923. [Google Scholar] [CrossRef]
- Cherif, A.O.; Ben Messaouda, M.; Kaabi, B.; Boukhchina, S.; Pepe, C.; Kallel, H. Comparison of the concentrations of long-chain alcohols (policosanol) in three Tunisian peanut varieties (Arachis hypogaea L.). J. Agric. Food Chem. 2010, 58, 12143–12148. [Google Scholar] [CrossRef] [PubMed]
- Yuka, H.J.; Ryua, H.W.; Kim, D.-Y.; Park, M.H.; Seo, W.D.; Jeong, S.H.; Sei-Ryang Oh, S.-R. Comparison of flavonoid and policosanol profiles in Korean winter-spinach (Spinacia oleracea L.) cultivated in different regions. Food Chem. 2019, 279, 202–208. [Google Scholar] [CrossRef] [PubMed]
- Chai, G.; Li, C.; Xu, F.; Li, Y.; Shi, X.; Wang, Y.; Wang, Z. Three endoplasmic reticulum-associated fatty acyl-coenzyme a reductases were involved in the production of primary alcohols in hexaploid wheat (Triticum aestivum L.). BMC Plant. Biol. 2018, 18, 41. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Sun, Y.; You, Q.; Luo, W.; Wang, C.; Zhao, S.; Chai, G.; Li, T.; Shi, X.; Li, C.; et al. Three Fatty Acyl-Coenzyme A Reductases, BdFAR1, BdFAR2 and BdFAR3, are involved in cuticular wax primary alcohol biosynthesis in Brachypodium distachyon. Plant Cell Physiol. 2018, 59, 527–543. [Google Scholar] [CrossRef]
- Singh, D.K.; Li, L.; Porter, T.D. Policosanol inhibits cholesterol synthesis in hepatoma cells by activation of AMP-kinase. J. Pharmacol. Exp. Ther. 2006, 318, 1020–1026. [Google Scholar] [CrossRef]
- Ding, Y.Y.; Fang, Y.; Pan, Y.; Lan, J.; Xu, T.; Zhang, W.; Mao, H.; Gu, Z.; Chen, X.; Shen, Q. Orally administered octacosanol improves liver insulin resistance in high-fat diet-fed mice through the reconstruction of the gut microbiota structure and inhibition of the TLR4/NF-κB inflammatory pathway. Food Funct. 2023, 14, 769–786. [Google Scholar] [CrossRef]
- Bai, J.; Yang, T.; Zhou, Y.; Xu, W.; Han, S.; Guo, T.; Zhu, L.; Qin, D.; Luo, Y.; Hu, Z.; et al. Octacosanol modifies obesity, expression profile and inflammation response of hepatic tissues in high-fat diet mice. Foods 2022, 11, 1606. [Google Scholar] [CrossRef]
- Miao, S.T.; Lu, Q.S.; Zhou, Y.J.; Chang, Y.N.; Xu, T.; Zhu, M.Y. Oral administration of octacosanol modulates the gut bacteria and protects the intestinal barrier in ulcerative colitis mice. J. Food Biochem. 2022, 46, e14284. [Google Scholar] [CrossRef]
- Zhou, Y.; Cao, F.; Wu, Q.; Luo, Y.; Guo, T.; Han, S.; Huang, M.; Hu, Z.; Bai, J.; Luo, F.; et al. Dietary supplementation of octacosanol improves exercise-induced fatigue and its molecular mechanism. J. Agric. Food Chem. 2021, 69, 7603–7618. [Google Scholar] [CrossRef]
- Su, J.; Liu, B.; Liao, J.; Yang, Z.; Lin, C.; Oka, Y. Coordination of Cryptochrome and phytochrome signals in the regulation of plant light responses. Agronomy 2017, 7, 25. [Google Scholar] [CrossRef]
- Wang, Q.; Zuo, Z.; Wang, X.; Gu, L.; Yoshizumi, T.; Yang, Z.; Yang, L.; Liu, Q.; Liu, W.; Han, Y.J.; et al. Photoactivation and inactivation of Arabidopsis cryptochrome 2. Science 2016, 354, 343–347. [Google Scholar] [CrossRef] [PubMed]
- Kopsell, D.A.; Sams, C.E. Increases in shoot tissue pigments, glucosinolates, and mineral elements in sprouting broccoli after exposure to short-duration blue light from light emitting diodes. J. Am. Soc. Hortic. Sci. 2013, 138, 31–37. [Google Scholar] [CrossRef]
- Samuolienė, G.; Sirtautas, R.; Brazaitytė, A.; Duchovskis, P. LED lighting and seasonality effects antioxidant properties of baby leaf lettuce. Food Chem. 2012, 134, 1494–1499. [Google Scholar] [CrossRef] [PubMed]
- Lefsrud, M.G.; Kopsell, D.A.; Kopsell, D.E.; Curran-Celentano, J. Irradiance levels affect growth parameters and carotenoid pigments in kale and spinach grown in a controlled environment. Physiol. Plant 2006, 127, 624–631. [Google Scholar] [CrossRef]
- Charron, C.S.; Sams, C.E. Glucosinolate content and myrosinase activity in rapid-cycling Brassica oleracea grown in a controlled environment. J. Amer. Soc. Hort. Sci. 2004, 129, 321–330. [Google Scholar] [CrossRef]
- Shin, O.H.; Kim, D.Y.; Seo, Y.W. Effects of different depth of grain colour on antioxidant capacity during water imbibition in wheat (Triticum aestivum L). J. Sci. Food Agric. 2017, 97, 2750–2758. [Google Scholar] [CrossRef]
- Galili, T.; O’Callaghan, A.; Sidi, J.; Sievert, C. Heatmaply: An R package for creating interactive cluster heatmaps for online publishing. Bioinformatics 2018, 34, 1600–1602. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| tR (min) | PC-TMS Derivatives 1 Mass (m/z) | Molecular Weight (m/z) | Molecular Formula | Identification |
|---|---|---|---|---|
| 3.57 | 355.3 [M–15]+ | 370.3 [M]+ | C20H42O | Eicosanol |
| 4.20 | 369.4 [M–15]+ | 384.4 [M]+ | C21H44O | Heneicosanol |
| 5.02 | 383.4 [M–15]+ | 398.4 [M]+ | C22H46O | Docosanol |
| 5.96 | 397.4 [M–15]+ | 412.4 [M]+ | C23H48O | Tricosanol |
| 7.38 | 411.4 [M–15]+ | 426.4 [M]+ | C24H50O | Tetracosanol |
| 11.30 | 439.5 [M–15]+ | 454.5 [M]+ | C26H54O | Hexacosanol |
| 12.83 | 453.5 [M–15]+ | 468.5 [M]+ | C27H56O | Heptacosanol |
| 13.95 | 467.5 [M–15]+ | 482.5 [M]+ | C28H58O | Octacosanol |
| 16.43 | 495.5 [M–15]+ | 510.5 [M]+ | C30H62O | Triacontanol |
| Cultivars | Mutant Lines | Policosanol Content (mg/100 g Dry Weight) a,b | |||||||
|---|---|---|---|---|---|---|---|---|---|
| C22-OH c | C23-OH c | C24-OH c | C26-OH c | C27-OH c | C28-OH c | C30-OH c | Total PC | ||
| Woori-mil × D-7 (original; WS01) | 4.72 ± 3.13 | 5.94 ± 0.64 | 1.94 ± 0.33 | 35.02 ± 1.99 | 6.98 ± 1.03 | 348.57 ± 34.81 | 94.34 ± 9.37 | 497.50 ± 51.30 | |
| WS02 | 4.19 ± 3.46 | 11.05 ± 3.94 | 4.31 ± 3.03 | 42.49 ± 10.64 | 10.75 ± 5.22 | 247.91 ± 32.35 | 49.94 ± 25.61 | 370.63 ± 84.25 | |
| WS03 | 5.99 ± 2.33 | 13.39 ± 3.08 | 6.89 ± 2.65 | 46.89 ± 9.27 | 11.99 ± 5.85 | 212.92 ± 14.99 | 63.09 ± 15.34 | 361.17 ± 53.50 | |
| WS04 | 2.81 ± 0.70 | 7.24 ± 0.94 | 1.88 ± 0.40 | 29.97 ± 6.20 | 5.65 ± 1.00 | 268.99 ± 17.92 | 60.02 ± 12.46 | 376.56 ± 39.62 | |
| WS05 | 6.02 ± 3.48 | 6.75 ± 0.97 | 3.07 ± 1.87 | 41.35 ± 8.54 | 16.98 ± 18.85 | 321.24 ± 34.29 | 77.08 ± 17.67 | 472.48 ± 85.67 | |
| WS06 | 3.49 ± 2.63 | 7.911.93 | 1.921.89 | 22.93 ± 2.87 | 10.15 ± 0.62 | 223.92 ± 19.73 | 69.33 ± 9.55 | 339.65 ± 39.22 | |
| WS07 | 3.71 ± 0.91 | 9.10 ± 0.82 | 5.65 ± 0.93 | 31.49 ± 4.23 | 10.17 ± 0.89 | 233.01 ± 26.73 | 67.32 ± 10.39 | 360.46 ± 44.90 | |
| WS08 | 4.33 ± 0.37 | 10.29 ± 1.36 | 7.10 ± 0.98 | 43.15 ± 5.09 | 10.60 ± 1.29 | 227.97 ± 12.36 | 58.02 ± 9.66 | 361.45 ± 31.13 | |
| WS09 | 3.68 ± 1.92 | 11.21 ± 1.38 | 4.08 ± 2.18 | 29.01 ± 3.20 | 5.80 ± 1.60 | 249.19 ± 13.68 | 76.01 ± 7.44 | 378.98 ± 31.39 | |
| WS10 | 5.03 ± 1.92 | 8.44 ± 0.59 | 2.32 ± 1.01 | 29.02 ± 2.27 | 7.10 ± 4.29 | 228.27 ± 14.28 | 65.21 ± 7.02 | 345.39 ± 31.39 | |
| WS11 | 8.34 ± 2.69 | 12.75 ± 1.37 | 6.69 ± 1.86 | 32.47 ± 1.96 | 6.47 ± 1.54 | 283.89 ± 25.47 | 86.73 ± 6.81 | 437.34 ± 41.70 | |
| WS12 | 3.26 ± 0.60 | 8.69 ± 0.81 | 2.10 ± 0.44 | 23.41 ± 1.52 | 11.09 ± 4.04 | 214.56 ± 18.32 | 68.22 ± 5.39 | 331.32 ± 31.11 | |
| WS13 | 4.39 ± 1.10 | 9.51 ± 0.63 | 9.73 ± 1.06 | 52.22 ± 3.74 | 12.52 ± 2.50 | 214.25 ± 11.12 | 62.62 ± 2.42 | 365.25 ± 22.58 | |
| WS14 | 4.10 ± 2.10 | 9.24 ± 0.14 | 5.93 ± 0.41 | 35.02 ± 2.10 | 6.41 ± 1.23 | 234.20 ± 24.78 | 73.44 ± 6.40 | 368.34 ± 37.15 | |
| WS15 | 6.18 ± 0.85 | 8.18 ± 0.75 | 8.82 ± 1.25 | 39.30 ± 2.92 | 9.01 ± 3.83 | 182.72 ± 31.71 | 62.24 ± 9.76 | 316.46 ± 51.07 | |
| WS16 | 1.06 ± 0.24 | 10.76 ± 1.07 | 3.48 ± 0.46 | 42.65 ± 2.96 | 9.38 ± 0.64 | 248.19 ± 22.15 | 68.76 ± 4.29 | 384.27 ± 31.81 | |
| WS17 | 2.92 ± 1.32 | 8.65 ± 0.57 | 3.90 ± 2.07 | 30.11 ± 1.52 | 5.30 ± 1.64 | 240.05 ± 28.98 | 73.86 ± 3.70 | 364.79 ± 39.80 | |
| WS18 | 6.46 ± 2.43 | 12.00 ± 1.25 | 4.25 ± 2.09 | 27.38 ± 1.60 | 6.67 ± 0.49 | 264.94 ± 24.32 | 78.53 ± 6.25 | 400.23 ± 38.45 | |
| WS19 | 2.13 ± 0.26 | 6.75 ± 0.48 | 3.15 ± 0.14 | 43.46 ± 2.66 | 7.89 ± 1.19 | 270.22 ± 27.60 | 74.98 ± 7.16 | 408.57 ± 39.49 | |
| WS20 | 3.43 ± 0.24 | 9.56 ± 0.83 | 2.06 ± 0.09 | 38.58 ± 1.54 | 6.50 ± 0.95 | 222.01 ± 20.38 | 37.65 ± 2.16 | 319.79 ± 26.20 | |
| WS21 | 3.16 ± 1.68 | 11.73 ± 0.98 | 4.84 ± 1.98 | 32.11 ± 0.99 | 5.68 ± 0.82 | 248.80 ± 23.24 | 74.36 ± 3.87 | 380.67 ± 33.56 | |
| WS22 | 3.86 ± 2.18 | 11.02 ± 1.02 | 1.53 ± 0.25 | 26.90 ± 1.05 | 5.98 ± 3.09 | 241.09 ± 29.59 | 73.14 ± 4.35 | 363.51 ± 41.52 | |
| WS23 | 1.31 ± 1.28 | 6.31 ± 5.47 | 0.23 ± 0.20 | 19.11 ± 16.56 | 3.33 ± 5.14 | 205.24 ± 178.40 | 53.64 ± 46.83 | 289.17 ± 253.88 | |
| WS24 | 2.67 ± 0.61 | 8.83 ± 0.41 | 3.04 ± 0.09 | 29.91 ± 1.03 | 4.81 ± 0.89 | 242.92 ± 27.91 | 69.96 ± 4.80 | 362.14 ± 35.74 | |
| WS25 | 4.18 ± 0.71 | 10.34 ± 0.72 | 3.58 ± 0.23 | 33.03 ± 1.11 | 7.06 ± 0.46 | 224.44 ± 9.35 | 56.60 ± 3.17 | 339.23 ± 15.75 | |
| WS26 | 1.95 ± 0.35 | 8.31 ± 0.68 | 3.03 ± 0.13 | 37.80 ± 1.95 | 11.19 ± 4.43 | 183.26 ± 17.87 | 56.61 ± 4.45 | 302.14 ± 29.85 | |
| WS27 | 2.37 ± 2.42 | 10.11 ± 0.80 | 0.94 ± 0.49 | 39.59 ± 1.31 | 10.51 ± 4.38 | 312.25 ± 13.62 | 60.60 ± 10.97 | 436.37 ± 34.00 | |
| WS28 | 4.80 ± 1.39 | 8.47 ± 0.79 | 1.52 ± 0.21 | 28.63 ± 1.43 | 6.75 ± 0.67 | 283.63 ± 28.49 | 60.79 ± 5.78 | 394.59 ± 38.75 | |
| WS29 | 5.33 ± 0.11 | 14.71 ± 1.25 | 5.36 ± 1.57 | 43.33 ± 1.30 | 5.62 ± 1.46 | 333.42 ± 30.30 | 80.04 ± 6.35 | 487.81 ± 42.35 | |
| WS30 | 2.37 ± 1.13 | 9.09 ± 0.81 | 4.51 ± 1.28 | 44.15 ± 2.22 | 10.79 ± 1.04 | 211.24 ± 10.40 | 65.61 ± 4.95 | 347.76 ± 21.82 | |
| WS31 | 4.35 ± 0.68 | 12.07 ± 0.36 | 4.64 ± 1.61 | 60.96 ± 2.13 | 10.82 ± 3.30 | 390.60 ± 36.83 | 98.94 ± 7.69 | 582.37 ± 52.60 | |
| WS32 | 3.18 ± 0.01 | 7.43 ± 0.36 | 3.10 ± 0.20 | 37.94 ± 2.06 | 9.79 ± 2.11 | 202.02 ± 14.89 | 63.54 ± 1.07 | 326.99 ± 20.70 | |
| WS33 | 5.16 ± 0.91 | 7.67 ± 0.65 | 5.47 ± 1.58 | 45.34 ± 2.74 | 7.64 ± 0.88 | 213.42 ± 12.63 | 69.59 ± 3.46 | 354.29 ± 22.86 | |
| WS34 | 1.36 ± 0.59 | 9.97 ± 0.60 | 2.64 ± 0.83 | 26.81 ± 0.82 | 3.76 ± 0.48 | 224.64 ± 19.59 | 59.22 ± 3.72 | 328.41 ± 26.63 | |
| WS35 | 3.39 ± 1.93 | 7.21 ± 0.52 | 3.42 ± 0.20 | 42.70 ± 1.95 | 6.03 ± 1.17 | 223.93 ± 26.32 | 67.28 ± 2.58 | 353.96 ± 34.67 | |
| WS36 | 2.11 ± 1.11 | 8.62 ± 0.41 | 0.90 ± 0.11 | 23.35 ± 0.50 | 9.33 ± 3.12 | 209.56 ± 22.28 | 62.47 ± 2.26 | 316.33 ± 29.79 | |
| WS37 | 4.37 ± 2.50 | 5.88 ± 0.34 | 0.53 ± 0.35 | 58.12 ± 4.12 | 4.88 ± 2.91 | 636.02 ± 43.59 | 121.20 ± 3.70 | 830.99 ± 57.51 | |
| WS38 | 1.84 ± 1.61 | 6.78 ± 6.00 | 6.80 ± 5.93 | 29.65 ± 25.83 | 8.52 ± 7.62 | 130.34 ± 114.73 | 40.44 ± 35.37 | 224.38 ± 197.10 | |
| WS39 | 3.42 ± 0.05 | 8.67 ± 0.33 | 2.24 ± 0.21 | 29.21 ± 0.89 | 12.15 ± 5.67 | 239.92 ± 19.22 | 60.83 ± 5.32 | 356.43 ± 31.68 | |
| WS40 | 2.93 ± 1.69 | 13.44 ± 0.63 | 2.49 ± 3.32 | 31.93 ± 1.35 | 7.55 ± 3.46 | 372.22 ± 24.89 | 75.52 ± 11.07 | 506.08 ± 46.41 | |
| WS41 | 3.65 ± 1.33 | 13.39 ± 0.49 | 5.46 ± 1.80 | 45.97 ± 1.44 | 11.73 ± 5.76 | 290.41 ± 23.28 | 73.77 ± 1.49 | 444.37 ± 35.60 | |
| WS42 | 3.18 ± 0.05 | 10.78 ± 0.23 | 10.59 ± 0.32 | 49.75 ± 0.13 | 15.39 ± 1.00 | 234.83 ± 26.43 | 59.00 ± 2.41 | 383.53 ± 30.57 | |
| WS43 | 3.77 ± 2.35 | 12.58 ± 0.28 | 6.23 ± 1.76 | 33.30 ± 0.86 | 9.53 ± 3.43 | 338.65 ± 25.86 | 67.91 ± 7.65 | 471.95 ± 42.19 | |
| WS44 | 2.24 ± 0.14 | 12.46 ± 0.38 | 4.26 ± 2.33 | 29.30 ± 0.40 | 5.82 ± 1.05 | 324.63 ± 24.16 | 64.26 ± 5.46 | 442.97 ± 33.94 | |
| WS45 | 3.00 ± 0.14 | 11.54 ± 0.34 | 3.29 ± 2.13 | 25.95 ± 0.31 | 5.54 ± 1.44 | 317.86 ± 12.59 | 78.58 ± 2.36 | 445.77 ± 19.32 | |
| WS46 | 5.76 ± 0.10 | 11.46 ± 0.27 | 7.10 ± 0.57 | 63.67 ± 0.99 | 9.75 ± 3.40 | 395.49 ± 20.94 | 65.42 ± 8.83 | 558.65 ± 35.10 | |
| WS47 | 3.04 ± 0.07 | 9.28 ± 0.19 | 1.49 ± 0.18 | 26.71 ± 0.94 | 10.18 ± 4.17 | 291.95 ± 19.80 | 70.68 ± 0.47 | 413.34 ± 25.84 | |
| WS48 | 4.33 ± 0.13 | 11.87 ± 0.33 | 2.35 ± 0.23 | 31.40 ± 0.56 | 10.54 ± 5.92 | 417.83 ± 30.78 | 67.92 ± 11.63 | 546.24 ± 49.57 | |
| WS49 | 4.68 ± 0.09 | 17.07 ± 0.41 | 4.03 ± 0.36 | 65.12 ± 1.88 | 7.87 ± 1.21 | 445.23 ± 33.53 | 56.78 ± 5.16 | 600.78 ± 42.64 | |
| WS50 | 2.40 ± 1.09 | 10.00 ± 0.04 | 1.30 ± 0.15 | 26.84 ± 0.67 | 5.98 ± 1.36 | 279.45 ± 26.89 | 71.29 ± 4.22 | 397.24 ± 34.42 | |
| WS51 | 5.24 ± 0.10 | 10.86 ± 0.85 | 4.17 ± 0.54 | 57.39 ± 1.88 | 10.36 ± 3.37 | 403.45 ± 26.90 | 68.34 ± 3.02 | 559.80 ± 36.67 | |
| WS52 | 2.04 ± 0.16 | 7.75 ± 0.34 | 0.65 ± 0.08 | 23.36 ± 0.69 | 8.37 ± 2.73 | 324.61 ± 22.25 | 75.97 ± 3.06 | 442.75 ± 29.30 | |
| WS53 | 4.20 ± 0.11 | 12.31 ± 0.17 | 5.52 ± 0.30 | 54.09 ± 0.36 | 11.51 ± 0.82 | 340.89 ± 24.48 | 69.13 ± 1.76 | 497.64 ± 27.99 | |
| WS54 | 4.30 ± 0.13 | 9.96 ± 0.28 | 8.57 ± 1.80 | 60.26 ± 0.36 | 18.91 ± 0.42 | 271.48 ± 42.23 | 71.55 ± 0.22 | 445.03 ± 45.45 | |
| WS55 | 4.03 ± 0.13 | 9.35 ± 0.18 | 1.80 ± 0.17 | 25.73 ± 1.06 | 5.73 ± 0.45 | 357.13 ± 27.72 | 64.08 ± 0.97 | 467.84 ± 30.68 | |
| WS56 | 2.25 ± 0.24 | 5.58 ± 0.38 | 1.65 ± 0.42 | 26.90 ± 0.36 | 13.11 ± 2.32 | 278.15 ± 20.14 | 72.06 ± 2.81 | 399.69 ± 26.67 | |
| WS57 | 6.32 ± 4.38 | 7.66 ± 1.00 | 5.27 ± 3.56 | 70.85 ± 0.91 | 16.25 ± 1.49 | 515.98 ± 13.87 | 88.27 ± 9.94 | 710.60 ± 35.15 | |
| WS58 | 2.37 ± 0.09 | 14.26 ± 0.65 | 1.66 ± 0.38 | 31.34 ± 0.56 | 7.80 ± 4.51 | 328.16 ± 24.62 | 71.32 ± 2.71 | 456.90 ± 33.52 | |
| WS59 | 8.77 ± 0.32 | 8.20 ± 0.32 | 2.73 ± 0.23 | 18.95 ± 0.54 | 6.46 ± 0.78 | 296.17 ± 22.91 | 61.64 ± 2.26 | 402.93 ± 27.36 | |
| WS60 | 3.17 ± 0.01 | 7.15 ± 0.19 | 4.08 ± 0.17 | 32.73 ± 0.70 | 12.12 ± 5.33 | 225.04 ± 14.92 | 63.22 ± 3.97 | 347.50 ± 25.28 | |
| WS61 | 6.28 ± 0.22 | 12.93 ± 0.05 | 4.99 ± 0.21 | 44.79 ± 0.56 | 12.08 ± 3.25 | 287.80 ± 40.92 | 73.70 ± 0.79 | 442.58 ± 46.00 | |
| WS62 | 1.86 ± 0.04 | 12.91 ± 0.18 | 2.46 ± 0.80 | 28.47 ± 0.51 | 7.91 ± 3.96 | 334.24 ± 26.90 | 79.09 ± 1.18 | 466.95 ± 33.58 | |
| WS63 | 2.80 ± 0.10 | 10.56 ± 0.12 | 0.00 ± 0.00 | 15.32 ± 1.84 | 4.35 ± 4.45 | 456.98 ± 20.46 | 66.07 ± 5.03 | 556.09 ± 32.00 | |
| WS64 | 9.68 ± 1.61 | 14.38 ± 0.37 | 3.02 ± 0.17 | 32.92 ± 0.23 | 5.74 ± 1.70 | 350.95 ± 25.99 | 70.70 ± 9.45 | 487.40 ± 39.52 | |
| WS65 | 5.61 ± 0.87 | 13.36 ± 0.30 | 10.37 ± 1.11 | 51.54 ± 0.42 | 15.02 ± 0.95 | 269.75 ± 27.57 | 61.73 ± 8.60 | 427.39 ± 39.80 | |
| WS66 | 9.96 ± 0.16 | 26.47 ± 0.28 | 3.84 ± 0.39 | 33.61 ± 0.49 | 6.58 ± 1.92 | 377.49 ± 26.68 | 50.85 ± 2.96 | 508.80 ± 32.90 | |
| WS67 | 3.98 ± 2.84 | 14.67 ± 1.23 | 6.48 ± 2.72 | 25.89 ± 1.34 | 7.93 ± 3.84 | 284.14 ± 20.94 | 75.83 ± 5.57 | 418.92 ± 38.49 | |
| WS68 | 4.46 ± 0.11 | 11.87 ± 0.12 | 8.78 ± 2.54 | 49.24 ± 1.15 | 12.87 ± 3.56 | 273.23 ± 15.29 | 67.80 ± 2.35 | 428.24 ± 25.13 | |
| WS69 | 9.15 ± 0.51 | 15.91 ± 0.52 | 3.11 ± 0.12 | 24.93 ± 7.79 | 5.73 ± 2.45 | 617.49 ± 33.86 | 106.11 ± 11.28 | 782.42 ± 56.53 | |
| WS70 | 2.94 ± 0.11 | 9.81 ± 0.27 | 1.14 ± 0.24 | 29.41 ± 0.58 | 12.48 ± 4.07 | 394.60 ± 21.04 | 80.69 ± 8.81 | 531.06 ± 35.13 | |
| WS71 | 6.54 ± 0.08 | 17.19 ± 0.10 | 1.65 ± 0.18 | 18.66 ± 0.96 | 3.91 ± 1.34 | 305.97 ± 27.26 | 72.56 ± 1.78 | 426.48 ± 31.70 | |
| WS72 | 11.16 ± 0.24 | 16.36 ± 1.22 | 4.62 ± 0.48 | 28.51 ± 1.25 | 9.84 ± 3.21 | 581.83 ± 40.42 | 97.13 ± 4.56 | 749.44 ± 51.38 | |
| WS73 | 2.88 ± 0.13 | 10.83 ± 0.32 | 4.72 ± 1.51 | 30.06 ± 0.39 | 12.45 ± 3.47 | 292.07 ± 28.02 | 76.03 ± 2.51 | 429.03 ± 36.35 | |
| WS74 | 9.94 ± 5.21 | 13.61 ± 0.96 | -0.11 ± 0.16 | 29.41 ± 1.71 | 7.61 ± 6.01 | 686.38 ± 32.75 | 126.40 ± 12.48 | 873.24 ± 59.28 | |
| WS75 | 3.29 ± 0.13 | 10.93 ± 0.42 | 2.41 ± 0.49 | 33.63 ± 0.47 | 4.80 ± 0.43 | 452.24 ± 29.04 | 90.48 ± 11.34 | 597.79 ± 42.31 | |
| WS76 | 14.10 ± 0.07 | 14.16 ± 0.68 | 7.46 ± 0.82 | 30.43 ± 2.78 | 6.24 ± 1.22 | 541.15 ± 24.90 | 97.94 ± 2.30 | 711.48 ± 32.77 | |
| WS77 | 2.13 ± 0.15 | 11.33 ± 0.67 | 3.38 ± 0.19 | 43.32 ± 1.29 | 8.54 ± 0.79 | 304.42 ± 45.03 | 78.93 ± 9.35 | 452.04 ± 57.48 | |
| WS78 | 2.50 ± 0.12 | 8.16 ± 0.18 | 0.48 ± 0.25 | 43.42 ± 0.87 | 10.13 ± 3.27 | 466.44 ± 27.62 | 98.22 ± 14.23 | 629.35 ± 46.55 | |
| WS79 | 3.91 ± 0.04 | 12.59 ± 0.08 | 1.31 ± 0.30 | 27.26 ± 0.57 | 6.31 ± 3.68 | 431.67 ± 23.67 | 79.59 ± 11.16 | 562.65 ± 39.49 | |
| WS80 | 3.71 ± 0.19 | 11.10 ± 0.25 | 5.06 ± 0.30 | 47.12 ± 0.80 | 8.62 ± 0.41 | 301.20 ± 20.37 | 75.28 ± 2.23 | 452.10 ± 24.55 | |
| WS81 | 1.47 ± 0.42 | 8.38 ± 0.15 | 1.05 ± 0.05 | 33.53 ± 0.79 | 9.34 ± 0.11 | 518.96 ± 28.74 | 99.52 ± 12.11 | 672.24 ± 42.37 | |
| WS82 | 5.02 ± 0.16 | 9.40 ± 0.46 | 1.55 ± 0.12 | 25.94 ± 0.94 | 8.34 ± 2.66 | 219.84 ± 190.47 | 86.35 ± 6.56 | 356.45 ± 201.36 | |
| WS83 | 5.10 ± 0.17 | 16.55 ± 0.38 | 3.80 ± 2.22 | 27.04 ± 0.71 | 9.28 ± 0.87 | 319.58 ± 24.37 | 51.83 ± 8.12 | 433.19 ± 36.85 | |
| WS84 | 4.97 ± 3.19 | 9.01 ± 1.33 | 2.50 ± 3.88 | 25.47 ± 0.97 | 9.87 ± 0.52 | 322.80 ± 36.83 | 87.87 ± 4.65 | 462.50 ± 51.37 | |
| WS85 | 7.08 ± 2.74 | 11.22 ± 0.22 | 3.86 ± 3.21 | 21.74 ± 0.52 | 12.16 ± 0.80 | 351.35 ± 11.36 | 66.75 ± 3.57 | 474.16 ± 22.43 | |
| WS86 | 4.46 ± 1.34 | 10.07 ± 0.13 | 2.52 ± 2.24 | 29.74 ± 0.63 | 8.60 ± 1.39 | 294.76 ± 39.32 | 71.77 ± 1.85 | 421.93 ± 46.89 | |
| Geumkang (commercial; WS87) | 3.53 ± 0.06 | 7.28 ± 1.32 | 6.42 ± 2.39 | 44.13 ± 2.13 | 9.17 ± 3.05 | 209.00 ± 9.87 | 67.18 ± 4.58 | 346.71 ± 23.40 | |
| WS88 | 2.83 ± 0.07 | 8.34 ± 0.55 | 5.68 ± 0.72 | 37.39 ± 1.96 | 10.32 ± 8.79 | 196.84 ± 21.90 | 60.47 ± 5.12 | 321.87 ± 39.11 | |
| WS89 | 1.38 ± 1.31 | 13.49 ± 11.69 | 2.63 ± 2.55 | 28.63 ± 24.79 | 3.26 ± 3.07 | 199.37 ± 173.05 | 45.40 ± 39.52 | 294.16 ± 255.98 | |
| WS90 | 3.05 ± 0.42 | 8.48 ± 1.56 | 5.86 ± 0.54 | 34.82 ± 3.87 | 10.94 ± 6.37 | 173.55 ± 152.08 | 72.57 ± 6.92 | 309.27 ± 171.76 | |
| Cheongwoo (commercial; WS91) | 4.81 ± 0.85 | 10.43 ± 0.86 | 6.28 ± 0.73 | 26.14 ± 1.18 | 10.74 ± 2.66 | 217.67 ± 17.69 | 67.29 ± 2.73 | 343.36 ± 26.70 | |
| Cultivars | Mutant Lines | Policosanol Content (mg/100 g Dry Weight) a,b | |||||||
|---|---|---|---|---|---|---|---|---|---|
| C22-OH c | C23-OH c | C24-OH c | C26-OH c | C27-OH c | C28-OH c | C30-OH c | Total PC | ||
| Woori-mil × D-7 (original; WS01) | ND d | 3.89 ± 0.33 | 0.99 ± 0.85 | 21.62 ± 2.09 | 3.77 ± 0.43 | 394.04 ± 9.94 | 26.74 ± 1.91 | 451.06 ± 15.55 | |
| WS37 | ND | 3.55 ± 0.40 | ND | 13.76 ± 1.13 | 2.68 ± 0.37 | 297.20 ± 11.32 | 22.03 ± 1.96 | 339.21 ± 15.12 | |
| WS49 | ND | 6.91 ± 0.89 | 4.00 ± 0.31 | 53.91 ± 5.31 | 5.39 ± 0.62 | 448.52 ± 23.34 | 18.20 ± 1.70 | 536.94 ± 32.17 | |
| WS57 | ND | 6.49 ± 0.49 | ND | 41.48 ± 4.15 | 3.98 ± 0.56 | 393.50 ± 30.34 | 25.17 ± 2.83 | 470.63 ± 38.37 | |
| WS69 | 4.31 ± 0.54 | 6.88 ± 0.71 | 3.12 ± 0.50 | 15.10 ± 1.28 | 3.19 ± 0.16 | 360.84 ± 21.47 | 28.90 ± 2.68 | 422.35 ± 27.34 | |
| WS72 | 0.99 ± 0.28 | 5.24 ± 0.60 | 1.64 ± 0.36 | 11.06 ± 1.22 | 2.59 ± 0.43 | 292.60 ± 17.32 | 20.20 ± 2.30 | 334.33 ± 22.51 | |
| WS74 | 2.76 ± 0.87 | 9.03 ± 0.80 | 3.45 ± 1.60 | 18.43 ± 1.61 | 4.25 ± 0.58 | 496.20 ± 27.75 | 33.42 ± 2.61 | 567.54 ± 35.81 | |
| WS75 | 2.14 ± 0.23 | 6.59 ± 0.87 | 3.26 ± 0.32 | 17.99 ± 1.45 | 2.38 ± 0.39 | 353.03 ± 22.60 | 24.71 ± 2.37 | 410.01 ± 28.24 | |
| WS76 | 5.36 ± 2.18 | 4.68 ± 0.76 | 5.54 ± 1.28 | 25.32 ± 2.73 | 3.74 ± 0.48 | 445.55 ± 21.85 | 34.98 ± 5.27 | 525.16 ± 34.53 | |
| WS78 | ND | 3.61 ± 0.26 | ND | 27.37 ± 2.73 | 3.89 ± 0.15 | 363.45 ± 21.40 | 27.21 ± 4.76 | 425.54 ± 29.29 | |
| WS81 | 1.32 ± 0.49 | 4.79 ± 0.60 | 2.67 ± 0.12 | 19.45 ± 1.95 | 3.12 ± 0.11 | 377.09 ± 15.30 | 27.23 ± 2.36 | 435.68 ± 20.92 | |
| Cultivars | Mutant Lines | Policosanol Content (mg/100 g Dry Weight) a,b | |||||||
|---|---|---|---|---|---|---|---|---|---|
| C22-OH c | C23-OH c | C24-OH c | C26-OH c | C27-OH c | C28-OH c | C30-OH c | Total PC | ||
| Woori-mil × D-7 (original; WS01) | 2.51 ± 0.56 | 8.77 ± 1.22 | 3.84 ± 0.77 | 9.79 ± 1.86 | 1.47 ± 0.32 | 169.31 ± 8.92 | 9.40 ± 0.94 | 205.08 ± 14.59 | |
| WS37 | ND d | 10.60 ± 0.97 | 1.41 ± 0.22 | 8.36 ± 0.56 | 1.62 ± 0.10 | 208.41 ± 8.77 | 14.69 ± 0.83 | 245.10 ± 11.44 | |
| WS49 | 2.21 ± 0.22 | 10.5 ± 1.6 | 4.82 ± 1.02 | 36.18 ± 3.83 | 3.33 ± 0.44 | 309.97 ± 11.27 | 12.84 ± 1.51 | 379.82 ± 19.88 | |
| WS57 | 1.06 ± 0.12 | 12.1 ± 1.3 | 2.77 ± 0.18 | 20.97 ± 1.64 | 2.03 ± 0.31 | 202.37 ± 9.73 | 11.01 ± 0.87 | 252.31 ± 14.19 | |
| WS69 | ND | 8.2 ± 1.2 | ND | 8.31 ± 2.94 | ND | 289.79 ± 27.30 | 12.70 ± 3.89 | 318.96 ± 35.30 | |
| WS72 | 3.25 ± 1.00 | 15.4 ± 1.2 | 2.57 ± 0.15 | 11.37 ± 1.11 | 1.96 ± 0.29 | 256.28 ± 23.17 | 18.87 ± 1.92 | 309.73 ± 28.85 | |
| WS74 | ND | 9.0 ± 1.2 | 4.65 ± 1.21 | ND | ND | 268.88 ± 45.96 | 14.14 ± 1.56 | 296.70 ± 49.90 | |
| WS75 | 7.30 ± 1.39 | 13.9 ± 1.1 | 4.03 ± 0.80 | 16.29 ± 1.91 | ND | 270.53 ± 13.58 | 19.82 ± 1.34 | 331.87 ± 20.11 | |
| WS76 | 2.45 ± 0.12 | 14.5 ± 1.6 | 3.95 ± 1.14 | 15.95 ± 1.42 | 1.78 ± 0.36 | 262.53 ± 13.74 | 17.30 ± 2.31 | 318.48 ± 20.66 | |
| WS78 | 3.79 ± 0.21 | 7.8 ± 0.6 | 2.32 ± 0.57 | 15.24 ± 0.88 | 2.18 ± 0.36 | 216.93 ± 9.61 | 13.93 ± 0.66 | 262.18 ± 12.93 | |
| WS81 | 1.22 ± 0.02 | 11.8 ± 1.4 | 2.01 ± 0.23 | 5.91 ± 1.22 | ND | 145.81 ± 10.33 | 8.81 ± 1.29 | 175.54 ± 14.54 | |
| Cultivars | Mutant Lines | Policosanol Content (mg/100 g Dry Weight) a,b | |||||||
|---|---|---|---|---|---|---|---|---|---|
| C22-OH c | C23-OH c | C24-OH c | C26-OH c | C27-OH c | C28-OH c | C30-OH c | Total PC | ||
| Woori-mil × D-7 (original; WS01) | 0.60 ± 0.16 | 15.31 ± 1.97 | 2.27 ± 0.64 | 17.11 ± 5.10 | 3.03 ± 0.77 | 324.80 ± 25.77 | 19.50 ± 3.32 | 382.61 ± 37.74 | |
| WS37 | ND d | 9.33 ± 0.88 | 1.36 ± 0.13 | 10.01 ± 1.04 | 1.24 ± 0.16 | 234.17 ± 9.81 | 16.11 ± 0.22 | 272.22 ± 12.23 | |
| WS49 | 0.63 ± 0.03 | 10.31 ± 1.03 | 3.96 ± 0.22 | 40.08 ± 3.71 | 3.46 ± 0.34 | 338.52 ± 14.53 | 13.31 ± 1.11 | 410.27 ± 20.97 | |
| WS57 | 0.82 ± 0.15 | 10.67 ± 0.81 | 2.20 ± 0.20 | 22.13 ± 2.44 | 2.31 ± 0.17 | 246.80 ± 15.17 | 13.24 ± 1.13 | 298.16 ± 20.06 | |
| WS69 | ND | 12.47 ± 1.58 | 1.75 ± 0.33 | 10.33 ± 2.33 | 2.52 ± 1.14 | 270.68 ± 17.79 | 14.27 ± 0.91 | 312.02 ± 24.09 | |
| WS72 | ND | 22.27 ± 2.06 | 2.93 ± 0.47 | 20.45 ± 1.28 | 2.96 ± 0.33 | 455.58 ± 25.94 | 29.88 ± 3.73 | 534.07 ± 33.79 | |
| WS74 | ND | 9.17 ± 0.77 | ND | 4.19 ± 2.24 | 4.74 ± 2.36 | 268.94 ± 50.45 | 24.28 ± 19.81 | 311.32 ± 75.64 | |
| WS75 | 1.82 ± 0.12 | 15.81 ± 1.83 | 3.59 ± 0.10 | 14.79 ± 1.18 | 2.08 ± 0.23 | 280.17 ± 15.95 | 17.61 ± 2.65 | 335.86 ± 22.05 | |
| WS76 | 1.67 ± 0.07 | 14.55 ± 1.47 | 3.30 ± 0.58 | 14.30 ± 0.82 | 2.14 ± 0.66 | 292.10 ± 18.02 | 16.59 ± 1.57 | 344.64 ± 23.18 | |
| WS78 | ND | 7.69 ± 0.62 | 2.56 ± 0.30 | 29.98 ± 2.74 | 4.20 ± 0.81 | 333.04 ± 13.82 | 21.31 ± 1.66 | 398.78 ± 19.94 | |
| WS81 | 0.66 ± 0.09 | 9.14 ± 0.81 | 2.32 ± 0.17 | 11.14 ± 1.52 | 1.44 ± 0.12 | 215.82 ± 12.17 | 12.54 ± 1.21 | 253.06 ± 16.10 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Han, A.-R.; Choi, E.; Park, J.; Jo, S.-H.; Hong, M.J.; Kim, J.-B.; Ryoo, G.-H.; Jin, C.H. Comparison of Policosanol Profiles of the Sprouts of Wheat Mutant Lines and the Effect of Differential LED Lights on Selected Lines. Plants 2023, 12, 3377. https://doi.org/10.3390/plants12193377
Han A-R, Choi E, Park J, Jo S-H, Hong MJ, Kim J-B, Ryoo G-H, Jin CH. Comparison of Policosanol Profiles of the Sprouts of Wheat Mutant Lines and the Effect of Differential LED Lights on Selected Lines. Plants. 2023; 12(19):3377. https://doi.org/10.3390/plants12193377
Chicago/Turabian StyleHan, Ah-Reum, Euna Choi, Jisu Park, Sang-Hee Jo, Min Jeong Hong, Jin-Baek Kim, Ga-Hee Ryoo, and Chang Hyun Jin. 2023. "Comparison of Policosanol Profiles of the Sprouts of Wheat Mutant Lines and the Effect of Differential LED Lights on Selected Lines" Plants 12, no. 19: 3377. https://doi.org/10.3390/plants12193377
APA StyleHan, A.-R., Choi, E., Park, J., Jo, S.-H., Hong, M. J., Kim, J.-B., Ryoo, G.-H., & Jin, C. H. (2023). Comparison of Policosanol Profiles of the Sprouts of Wheat Mutant Lines and the Effect of Differential LED Lights on Selected Lines. Plants, 12(19), 3377. https://doi.org/10.3390/plants12193377