
The Cell Wall Characterization of Brown Alga Cladosiphon okamuranus during Growth

 ,
,  , ,
, , 
Abstract
:1. Introduction
2. Results
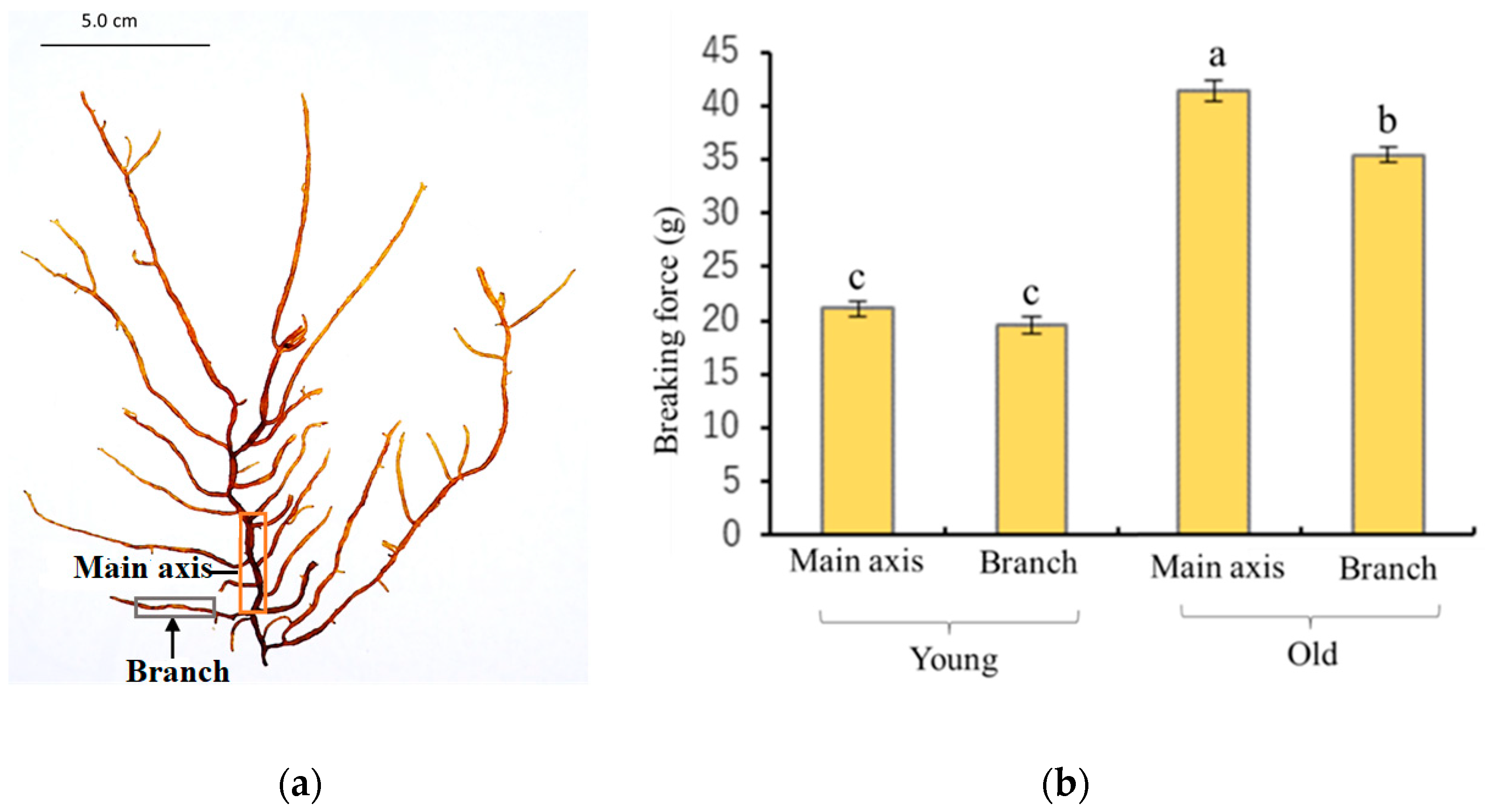
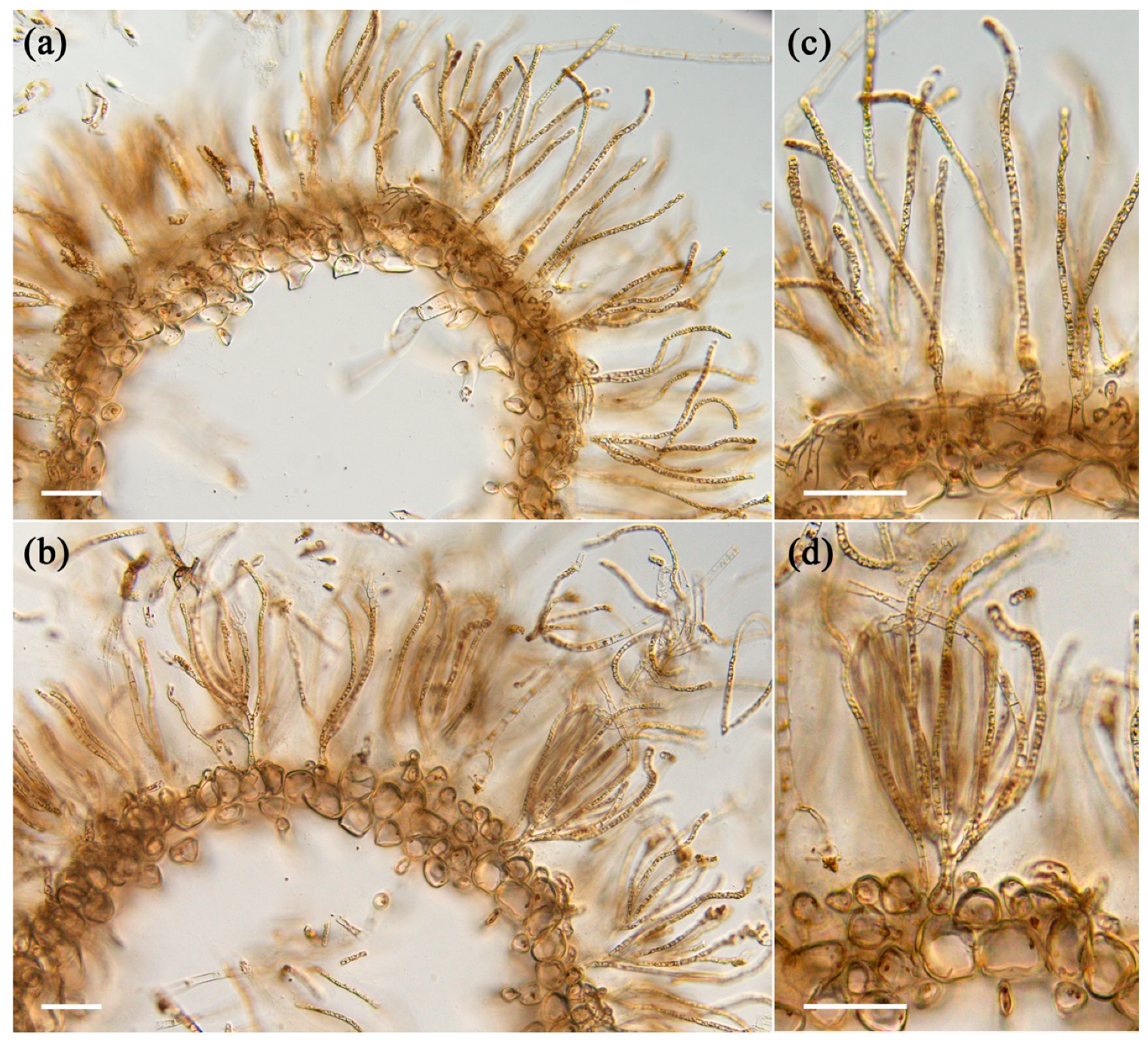
2.1. Physical Characteristics of Young and Old Thalli
2.2. Yield of AIR
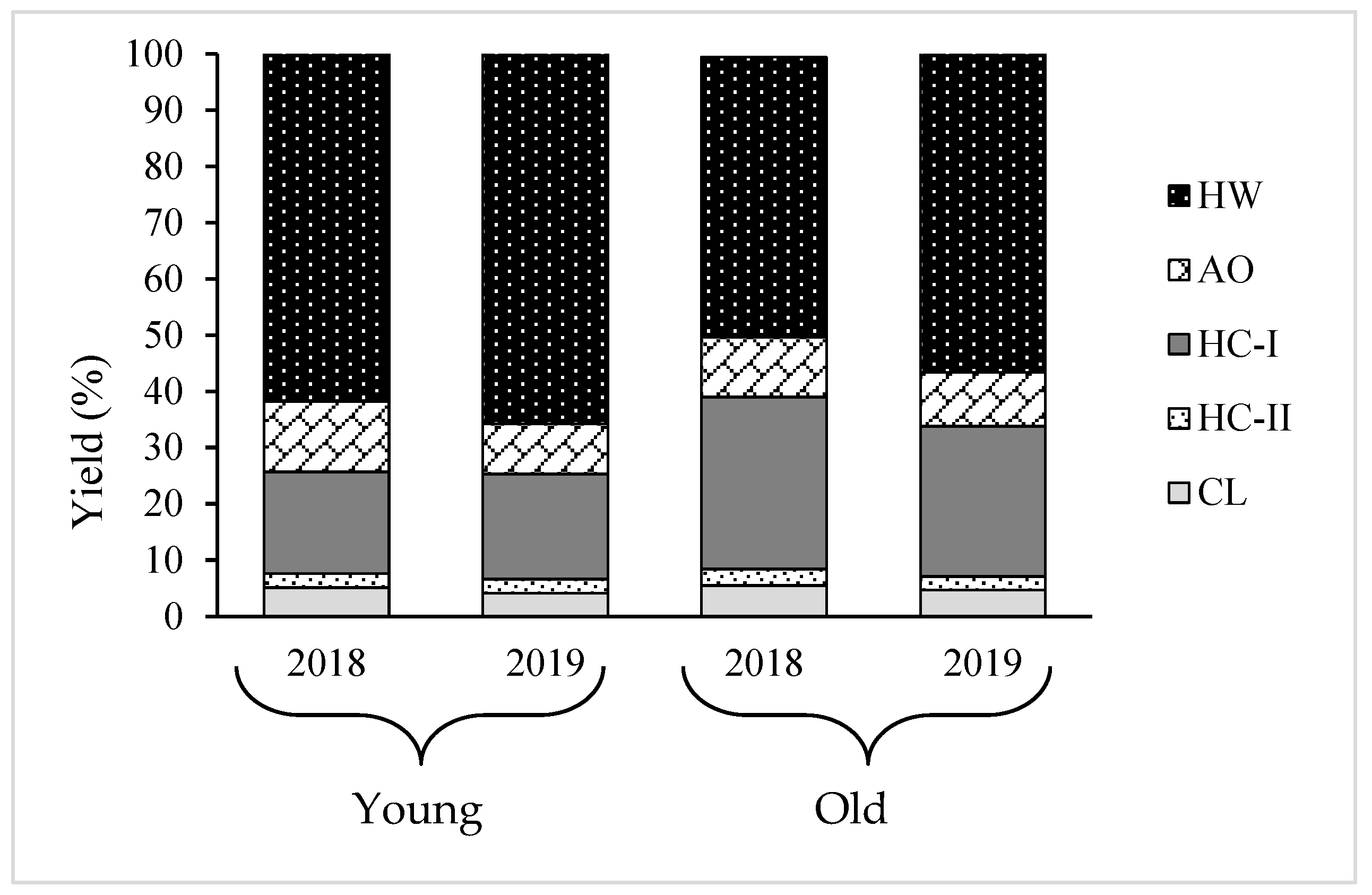
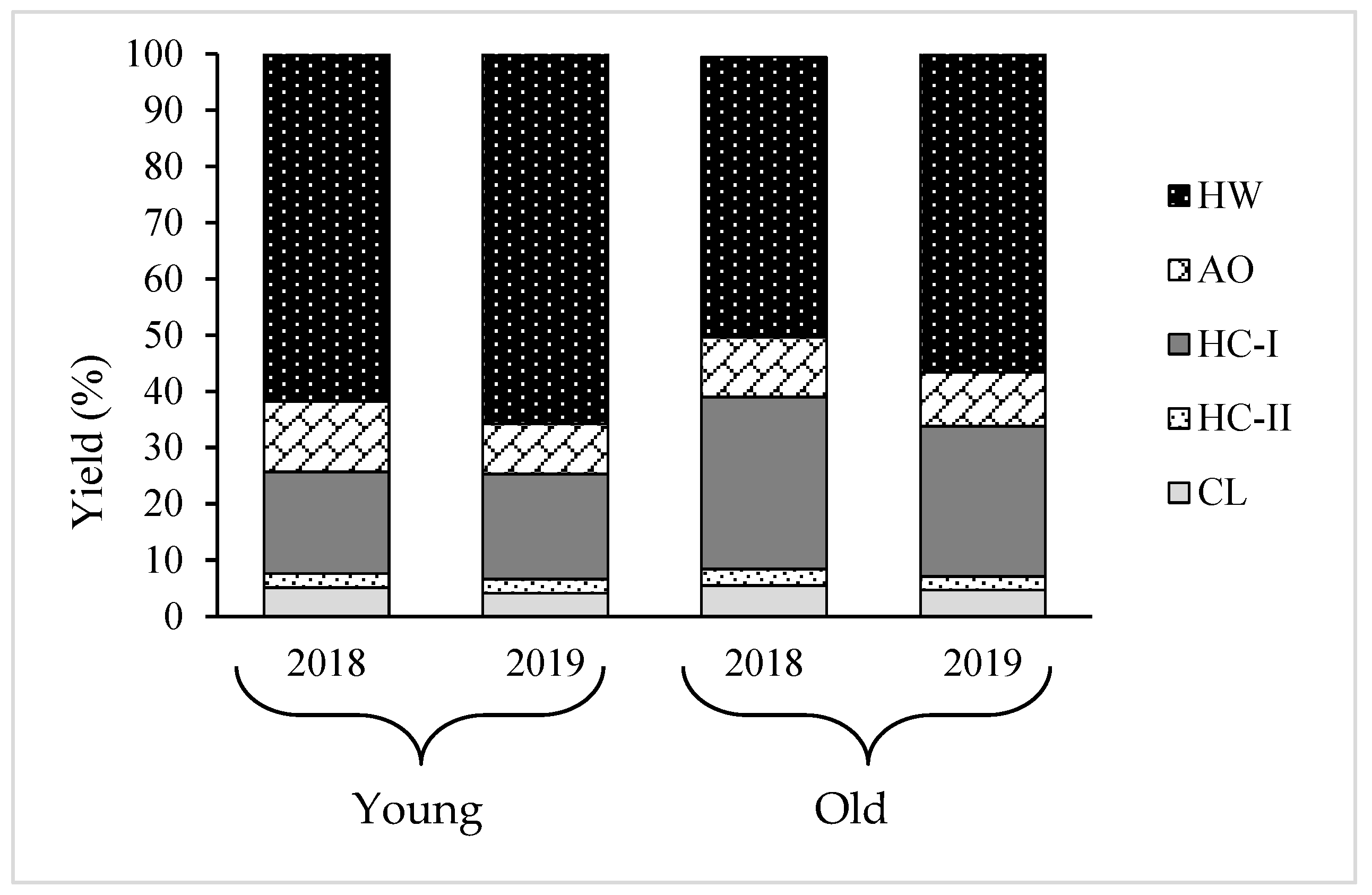
2.3. Yield and Composition of Different Cell Wall Fractions
2.4. Sugar Composition Analysis
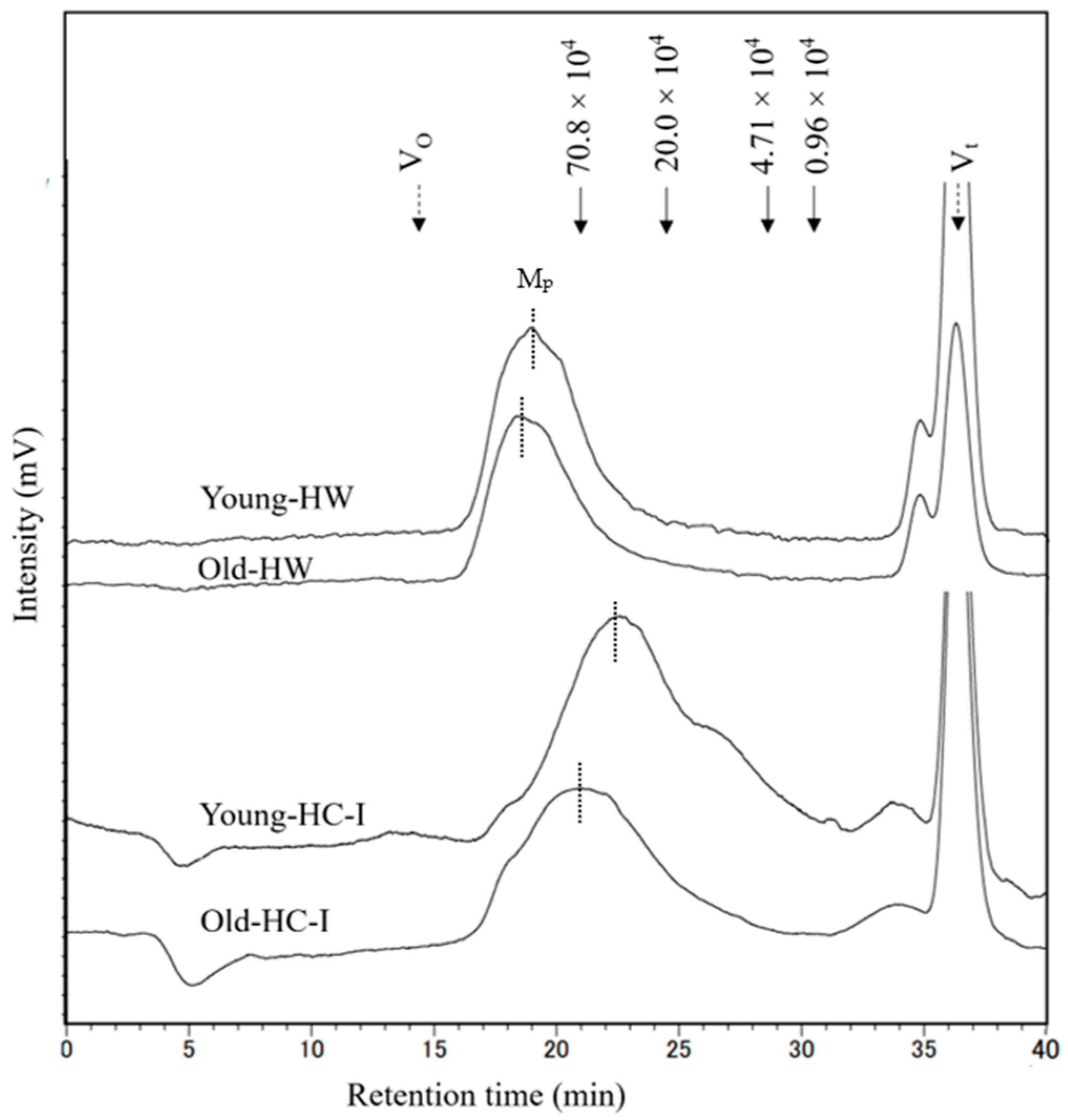
2.5. Molecular Weight Distribution
2.6. Anion Exchange Chromatography
3. Discussion
4. Materials and Methods
4.1. Algal Sample
4.2. Measurement of Moisture Content
4.3. Measurement of Tensile Strength
4.4. Light Microscopy
4.5. Preparation of Alcohol Insoluble Residue (AIR)
4.6. Fractionation of Cell Wall Polysaccharides
4.7. Chemical Composition Analysis
4.8. Sugar Composition Analysis
4.9. Determination of Molecular Weight (MW)
4.10. Anion Exchange Chromatography
4.11. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Popper, Z.A.; Michel, G.; Hervé, C.; Domozych, D.S.; Willats, W.G.; Tuohy, M.G.; Kloareg, B.; Stengel, D.B. Evolution and diversity of plant cell walls: From algae to flowering plants. Annu. Rev. Plant Biol. 2011, 62, 567–590. [Google Scholar] [CrossRef] [PubMed]
- Zhang, B.; Gao, Y.; Zhang, L.; Zhou, Y. The plant cell wall: Biosynthesis, construction, and functions. J. Integr. Plant Biol. 2021, 63, 251–272. [Google Scholar] [CrossRef]
- Chaiklahan, R.; Srinorasing, T.; Chirasuwan, N.; Tamtin, M.; Bunnag, B. The potential of polysaccharide extracts from Caulerpa lentillifera waste. Int. J. Biol. Macromol. 2020, 161, 1021–1028. [Google Scholar] [CrossRef]
- Silberfeld, T.; Leigh, J.W.; Verbruggen, H.; Cruaud, C.; de Reviers, B.; Rousseau, F. A multi-locus time-calibrated phylogeny of the brown algae (Heterokonta, Ochrophyta, Phaeophyceae): Investigating the evolutionary nature of the ‘brown algal crown radiation. Mol. Phylogenet. Evol. 2010, 56, 659–674. [Google Scholar] [CrossRef]
- Michel, G.; Tonon, T.; Scornet, D.; Cock, J.M.; Kloareg, B. The cell wall polysaccharide metabolism of the brown alga Ectocarpus siliculosus. Insights into the evolution of extracellular matrix polysaccharides in Eukaryotes. New Phytol. 2010, 188, 82–97. [Google Scholar] [CrossRef]
- Deniaud-Bouët, E.; Kervarec, N.; Michel, G.; Tonon, T.; Kloareg, B.; Hervé, C. Chemical and enzymatic fractionation of cell walls from Fucales: Insights into the structure of the extracellular matrix of brown algae. Ann. Bot. 2014, 114, 1203–1216. [Google Scholar] [CrossRef]
- Salgado, L.T.; Cinelli, L.P.; Viana, N.B.; Tomazetto de Carvalho, R.; de Souza Mourao, P.A.; Teixeira, V.L.; Farina, M.; Filho, A.G.M.A. Avanadium bromoperoxidase catalyzes the formation of high-molecular-weight complexes between brown algal phenolic substances and alginates. J. Phycol. 2009, 45, 193–202. [Google Scholar] [CrossRef]
- Siméon, A.; Kridi, S.; Kloareg, B.; Hervé, C. Presence of exogenous sulfate is mandatory for tip growth in the brown alga Ectocarpus subulatus. Front. Plant Sci. 2020, 11, 1277. [Google Scholar] [CrossRef]
- le Bail, A.A.; Billoud, B.; le Panse, S.; Chenivesse, S.; Charrier, B. ETOILE egulates developmental patterning in the filamentous brown alga Ectocarpus siliculosus. Plant Cell 2011, 23, 1666–1678. [Google Scholar] [CrossRef]
- Berger, F.; Taylor, A.; Brownlee, C. Cell fate determination by the cell wall in early fucus development. Science 1994, 263, 1421–1423. [Google Scholar] [CrossRef]
- Herburger, K.; Ryan, L.M.; Popper, Z.A.; Holzinger, A. Localisation and substrate specificities of transglycanases in charophyte algae relate to development and morphology. J. Cell Sci. 2018, 131, jcs203208. [Google Scholar] [CrossRef] [PubMed]
- Zvyagintseva, T.N.; Shevchenko, N.M.; Chizhov, A.O.; Krupnova, T.N.; Sundukova, E.V.; Isakov, V.V. Water-soluble polysaccharides of some far-eastern brown seaweeds. Distribution, structure, and their dependence on the developmental conditions. J. Exp. Mar. Biol. Ecol. 2003, 294, 1–13. [Google Scholar] [CrossRef]
- Awanthi, M.G.G.; Umosa, M.; Yuguchi, Y.; Oku, H.; Kitahara, K.; Ito, M.; Tanaka, A.; Konishi, T. Fractionation and characterization of cell wall polysaccharides from the brown alga Cladosiphon okamuranus. Carbohydr. Res. 2023, 523, 108722. [Google Scholar] [CrossRef] [PubMed]
- Tsuji, M.; Sudou, Y.; Enoki, M.; Tako, M.; Konishi, T. Variation in the contents and structure of fucoidan from cultivated Cladosiphon okamuranus; Tokida. Bull. Appl. Glycosci. 2013, 3, 248–252. [Google Scholar] [CrossRef]
- Starko, S.; Mansfield, S.D.; Martone, P.T. Cell wall chemistry and tissue structure underlie shifts in material properties of a perennial kelp. Eur. J. Phycol. 2018, 53, 307–317. [Google Scholar] [CrossRef]
- Krumhansl, K.A.; Demes, K.W.; Carrington, E.; Harley, C.D.G. Divergent growth strategies between red algae and kelps influence biomechanical properties. Am. J. Bot. 2015, 102, 1938–1944. [Google Scholar] [CrossRef]
- Martone, P.T. Kelp versus coralline: Cellular basis for mechanical strength in the wave-swept seaweed Calliarthron (Corallinaceae, Rhodophyta). J. Phycol. 2007, 43, 882–891. [Google Scholar] [CrossRef]
- Schiener, P.; Black, K.D.; Stanley, M.S.; Green, D.H. The seasonal variation in the chemical composition of the kelp species Laminaria digitata, Laminaria hyperborea, Saccharina latissima and Alaria esculenta. J. Appl. Phycol. 2015, 27, 363–373. [Google Scholar] [CrossRef]
- Deniaud-Bouët, E.; Hardouin, K.; Potin, P.; Kloareg, B.; Hervé, C. A review about brown algal cell walls and fucose-containing sulfated polysaccharides: Cell wall context, biomedical properties and key research challenges. Carbohydr. Polym. 2017, 175, 395–408. [Google Scholar] [CrossRef]
- Albersheim, P.; Darvill, A.; Roberts, K.; Sederoff, R.; Staehelin, A. Plant Cell Walls; Garland Science: New York, NY, USA, 2009; pp. 365–407. [Google Scholar]
- Kim, J.H.; Kim, J.S.; Wi, S.G.; Mun, S.P.; Chung, B.Y. The cell wall characterization at immature and mature stages of Arabidopsis thaliana L. Agric. Chem. Biotechnol. 2004, 47, 11–14. [Google Scholar]
- Burgert, I.; Fratzl, I. Plants control the properties and actuation of their organs through the orientation of cellulose fibrils in their cell walls. Integr. Comp. Biol. 2009, 49, 69–79. [Google Scholar] [CrossRef]
- Turner, S.R.; Somerville, C.R. Collapsed xylem phenotype of arabidopsis identifies mutants deficient in cellulose deposition in the secondary cell wall. Plant Cell 1997, 9, 689–701. [Google Scholar] [CrossRef]
- Revilla, G.; Zarra, I. Changes in the molecular weight distribution of the hemicellulosic polysaccharides from rice coleoptiles growing under different conditions. J. Exp. Bot. 1987, 38, 1818–1825. [Google Scholar] [CrossRef]
- Dubois, M.; Gilles, K.A.; Hamilton, J.K.; Rebers, P.A.; Smith, F. Colorimetric method for determination of sugars and related substances. Anal. Chem. 1956, 28, 350–356. [Google Scholar] [CrossRef]
- Blumenkrantz, N.; Asboehansen, G. New method for quantitative of uronic acids determination. Anal. Biochem. 1973, 89, 484–489. [Google Scholar] [CrossRef]
- Hillis, W.E.; Swain, T. The phenolic constituents of Prunus domestica. II.—The analysis of tissues of the Victoria plum tree. J. Sci. Food Agric. 1959, 10, 135–144. [Google Scholar] [CrossRef]
- Konishi, T.; Nakata, I.; Miyagi, Y.; Tako, M. Extraction of β-1,3 xylan from green seaweed, Caulerpa lentillifera. J. Appl. Glycosci. 2012, 59, 161–163. [Google Scholar] [CrossRef]
- Kato, Y.; Matsukura, J. Carbohydrate composition of major leaf vegetables: Carbohydrate composition of vegetable food (Part I). Bull. Fac. Educ. Hirosaki 1994, 7, 61–71. [Google Scholar]
- Peng, L.; Hocart, C.H.; Redmond, J.W.; Williamson, R.E. Fractionation of carbohydrates in Arabidopsis root cell walls shows that three radial swelling loci are specifically involved in cellulose production. Planta 2000, 211, 406–414. [Google Scholar] [CrossRef]
- Shiroma, R.; Konishi, T.; Uechi, S.; Tako, M. Structural study of fucoidan from the brown seaweed Hizikia fusiformis. Food Sci. Technol. Res. 2008, 14, 176–182. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Harvested Year | Yield of AIR * | Moisture Content * | |
|---|---|---|---|
| Young | 2018 | 2.8 | 93.2 |
| 2019 | 3.2 | 92.9 | |
| Old | 2018 | 5.5 | 91.4 |
| 2019 | 5.0 | 91.9 |
| Fraction | Yield a | Total Sugar | UA b | SO3− | Protein | Polyphenol | |
|---|---|---|---|---|---|---|---|
| HW | 62.4 | 49.4 | 24.5 | 24.5 | 8.9 | 2.9 | |
| AO | 12.5 | 59.5 | 34.3 | 3.5 | 6.6 | 1.7 | |
| Young | HC-I | 18.1 | 49.9 | 22.3 | 6.4 | 31.5 | 8.4 |
| HC-II | 2.5 | 92.3 | 7.8 | 1.9 | 8.1 | 5.6 | |
| CL | 5.1 | 100.0 | tr | tr | tr | tr | |
| HW | 49.8 | 56.2 | 22.9 | 21.7 | 14.5 | 3.9 | |
| AO | 10.6 | 64.8 | 30.5 | 6.1 | 8.0 | 2.1 | |
| Old | HC-I | 30.6 | 62.9 | 21.1 | 9.9 | 22.9 | 7.3 |
| HC-II | 2.9 | 80.1 | 13.7 | 5.9 | 10.0 | 2.4 | |
| CL | 5.5 | 100.0 | tr | tr | tr | tr |
| Fraction | Yield a | Total Sugar | UA b | SO3− | Protein | Polyphenol | |
|---|---|---|---|---|---|---|---|
| HW | 66.7 | 55.3 | 25.6 | 15.7 | 15.0 | 4.3 | |
| AO | 8.9 | 59.2 | 30.7 | 3.8 | 7.4 | 2.4 | |
| Young | HC-I | 18.7 | 67.4 | 21.8 | 6.7 | 21.9 | 6.8 |
| HC-II | 2.5 | 114.0 | 9.8 | 2.9 | 5.1 | 1.2 | |
| CL | 4.1 | 100.0 | tr | tr | tr | tr | |
| HW | 58.5 | 61.6 | 23.7 | 20.6 | 12.1 | 4.5 | |
| AO | 9.6 | 61.6 | 31.4 | 10.4 | 8.4 | 2.6 | |
| Old | HC-I | 26.7 | 65.3 | 21.2 | 8.5 | 18.9 | 5.3 |
| HC-II | 2.4 | 112.1 | 11.5 | 11.5 | 6.8 | 2.5 | |
| CL | 4.7 | 100.0 | tr | tr | tr | tr |
| Fraction | Neutral Sugar | UA | SO3− | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Fuc | Gal | Glc | Man | Xyl | GlcA | GalA/GulA | ManA | ||||
| HW | 1.0 | tr | tr | - | tr | 0.3 | tr | - | 0.7 | ||
| AO | 1.0 | 0.1 | tr | tr | 0.1 | tr | 0.3 | 0.3 | 0.1 | ||
| Young | HC-I | 1.0 | tr | 0.1 | tr | 0.2 | 0.3 | tr | - | 0.2 | |
| HC-II | 1.0 | - | 10.8 | 0.4 | 0.6 | 0.9 | - | - | 0.6 | ||
| CL | 1.0 | - | 55.9 | - | 3.7 | - | - | - | - | ||
| HW | 1.0 | tr | tr | - | tr | 0.3 | tr | - | 0.6 | ||
| AO | 1.0 | tr | - | - | tr | tr | 0.3 | 0.1 | 0.2 | ||
| Old | HC-I | 1.0 | tr | tr | - | 0.1 | 0.3 | tr | - | 0.3 | |
| HC-II | 1.0 | - | 1.3 | 0.1 | 0.3 | 0.4 | - | - | 0.4 | ||
| CL | 1.0 | - | 26.7 | - | 0.8 | - | - | - | - | ||
| Fraction | Neutral Sugar | UA | SO3− | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Fuc | Gal | Glc | Man | Xyl | GlcA | GalA/GulA | ManA | ||||
| HW | 1.0 | tr | 0.1 | - | - | 0.3 | tr | - | 0.5 | ||
| AO | 1.0 | tr | tr | 0.1 | 0.1 | tr | 0.2 | 0.2 | 0.1 | ||
| Young | HC-I | 1.0 | tr | tr | tr | tr | 0.3 | tr | - | 0.2 | |
| HC-II | 1.0 | - | 12.7 | 3.1 | 3.1 | 2.0 | - | - | 1.4 | ||
| CL | 1.0 | 0.8 | 71.5 | - | - | - | - | - | - | ||
| HW | 1.0 | tr | 0.1 | - | tr | 0.3 | tr | - | 0.7 | ||
| AO | 1.0 | tr | tr | - | tr | tr | 0.2 | 0.2 | 0.4 | ||
| Old | HC-I | 1.0 | tr | 0.1 | - | 0.1 | 0.3 | tr | - | 0.5 | |
| HC-II | 1.0 | - | 4.3 | 0.1 | 0.6 | 0.7 | - | - | 1.7 | ||
| CL | 1.0 | - | 18.9 | - | 1.2 | - | - | - | - | ||
| Fraction | Fuc | Gal | Glc | Man | Xyl | GlcA | SO3− | |
|---|---|---|---|---|---|---|---|---|
| HW-FT | 1.0 | tr | 0.6 | - | - | 0.3 | 0.4 | |
| Young | HW-NaCl | 1.0 | tr | tr | tr | tr | 0.2 | 0.4 |
| HC-I-NaCl | 1.0 | 0.1 | 0.1 | 0.1 | tr | 0.4 | 0.5 | |
| HW-FT | 1.0 | - | 0.3 | - | - | 0.3 | 0.4 | |
| Old | HW-NaCl | 1.0 | tr | tr | tr | tr | 0.2 | 0.3 |
| HC-I-NaCl | 1.0 | tr | tr | 0.1 | tr | 0.2 | 0.6 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Miwa, Y.; Awanthi, M.G.G.; Soga, K.; Tanaka, A.; Ito, M.; Numata, Y.; Sato, Y.; Konishi, T. The Cell Wall Characterization of Brown Alga Cladosiphon okamuranus during Growth. Plants 2023, 12, 3274. https://doi.org/10.3390/plants12183274
Miwa Y, Awanthi MGG, Soga K, Tanaka A, Ito M, Numata Y, Sato Y, Konishi T. The Cell Wall Characterization of Brown Alga Cladosiphon okamuranus during Growth. Plants. 2023; 12(18):3274. https://doi.org/10.3390/plants12183274
Chicago/Turabian StyleMiwa, Yuka, Mahanama Geegana Gamage Awanthi, Kouichi Soga, Atsuko Tanaka, Michihiro Ito, Yuichiro Numata, Yoichi Sato, and Teruko Konishi. 2023. "The Cell Wall Characterization of Brown Alga Cladosiphon okamuranus during Growth" Plants 12, no. 18: 3274. https://doi.org/10.3390/plants12183274
APA StyleMiwa, Y., Awanthi, M. G. G., Soga, K., Tanaka, A., Ito, M., Numata, Y., Sato, Y., & Konishi, T. (2023). The Cell Wall Characterization of Brown Alga Cladosiphon okamuranus during Growth. Plants, 12(18), 3274. https://doi.org/10.3390/plants12183274