
The Patterns of Intraspecific Variations in Mass of Nectar Sugar along a Phylogeny Distinguish Native from Non-Native Plants in Urban Greenspaces in Southern England


Abstract
1. Introduction
2. Results
3. Discussion
4. Materials and Methods
4.1. Study Area
4.2. Data Collection
4.3. Phylogenetic Tree of All the Species
4.4. Data Analysis
Supplementary Materials
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hulme, P.E. Trade, transport and trouble: Managing invasive species pathways in an era of globalization. J. Appl. Ecol. 2009, 46, 10–18. [Google Scholar] [CrossRef]
- Yessoufou, K.; Ambani, A.E.; Elansary, H.O.; Gaoue, O.G. Alien woody plants are more versatile than native, but both share similar therapeutic redundancy in South Africa. PLoS ONE 2021, 16, e0260390. [Google Scholar] [CrossRef] [PubMed]
- van Kleunen, M.; Xu, X.; Yang, Q.; Maurel, N.; Zhang, Z.; Dawson, W.; Essl, F.; Kreft, H.; Pergl, J.; Pyšek, P.; et al. Economic use of plants is key to their naturalization success. Nat. Commun. 2020, 11, 3201. [Google Scholar] [CrossRef]
- Richardson, D.M.; Pysek, P.; Rejmanek, M.; Barbour, M.G.; Panetta, F.D.; West, C.J. Naturalization and invasion of alien plants: Concepts and definitions. Divers. Distrib. 2000, 6, 93–107. [Google Scholar] [CrossRef]
- Bezeng, S.B.; Davies, J.T.; Yessoufou, K.; Maurin, O.; Van der Bank, M. Revisiting Darwin’s naturalization conundrum: Ex-plaining invasion success of non-native trees and shrubs in southern. J. Ecol. 2015, 103, 871–879. [Google Scholar] [CrossRef]
- Baker, H.G. Characteristics and modes of origin of weeds. In The Genetics of Colonizing Species; Baker, H.G., Stebbins, G.L., Eds.; Academic Press: New York, NY, USA, 1965; pp. 147–169. [Google Scholar]
- Lau, J.A.; Funk, J.L. How ecological and evolutionary theory expanded the ‘ideal weed’ concept. Oecologia 2023, 200, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Uesugi, A.; Kessler, A. Herbivore exclusion drives the evolution of plant competitiveness via increased allelopathy. New Phytol. 2013, 198, 916–924. [Google Scholar] [CrossRef]
- Goodell, K.; Parker, I.M. Invasion of a dominant floral resource: Effects on the floral community and pollination of native plants. Ecology 2017, 98, 57–69. [Google Scholar] [CrossRef]
- Willis, C.G.; Ruhfel, B.R.; Primack, R.B.; Miller-Rushing, A.J.; Losos, J.B.; Davis, C.C. Favorable Climate Change Response Explains Non-Native Species’ Success in Thoreau’s Woods. PLoS ONE 2010, 5, e8878. [Google Scholar] [CrossRef]
- Pyšek, P.; Jarošík, V.; Chytrý, M.; Danihelka, J.; Kühn, I.; Pergl, J.; Tichý, L.; Biesmeijer, J.C.; Ellis, W.N.; Kunin, W.E.; et al. Successful invaders co-opt pollinators of native flora and accumulate insect pollinators with increasing residence time. Ecol. Monogr. 2011, 81, 277–293. [Google Scholar] [CrossRef]
- Johnson, C.A.; Dutt, P.; Levine, J.M. Competition for pollinators destabilizes plant coexistence. Nature 2022, 607, 721–725. [Google Scholar] [CrossRef] [PubMed]
- Adedoja, O.; Erckie, L.; Boatwright, J.S.; van Wyk, E.; Geerts, S. An invasive alien Proteaceae lures some, but not all nectar-feeding bird pollinators away from native Proteaceae in South African fynbos. Plant Biol. 2021, 23, 915–922. [Google Scholar] [CrossRef] [PubMed]
- Vanbergen, A.J.; Espíndola, A.; Aizen, M.A. Risks to pollinators and pollination from invasive alien species. Nat. Ecol. Evol. 2017, 2, 16–25. [Google Scholar] [CrossRef] [PubMed]
- Mack, R.N.; Simberloff, D.; Lonsdale, W.M.; Evans, H.; Clout, M.; Bazzaz, F.A. Biotic invasions: Causes, epidemiology, global consequences, and control. Ecol. Appl. 2000, 10, 689–710. [Google Scholar] [CrossRef]
- Albrecht, M.; Padrón, B.; Bartomeus, I.; Traveset, A.; San-Jose, L.M.; Peñalver-Alcázar, M.; Milá, B.; Gonzalez-Jimena, V.; Fitze, P.S. Consequences of plant invasions on compartmentalization and species’ roles in plant–pollinator networks. Proc. R. Soc. B Boil. Sci. 2014, 281, 20140773. [Google Scholar] [CrossRef] [PubMed]
- Herron-Sweet, C.R.; Lehnhoff, E.A.; Burkle, L.A.; Littlefield, J.L.; Mangold, J.M. Temporal- and density-dependent impacts of an invasive plant on pollinators and pollination services to a native plant. Ecosphere 2016, 7, e01233. [Google Scholar] [CrossRef]
- Jones, E.I.; Gomulkiewicz, R. Biotic Interactions, Rapid Evolution, and the Establishment of Introduced Species. Am. Nat. 2012, 179, E28–E36. [Google Scholar] [CrossRef]
- Vandepitte, K.; de Meyer, T.; Helsen, K.; van Acker, K.; Roldán-Ruiz, I.; Mergeay, J.; Honnay, O. Rapid genetic adaptation precedes the spread of an exotic plant species. Mol. Ecol. 2014, 23, 2157–2164. [Google Scholar] [CrossRef]
- Bossdorf, O.; Auge, H.; Lafuma, L.; Rogers, W.E.; Siemann, E.; Prati, D. Phenotypic and genetic differentiation between native and introduced plant populations. Oecologia 2005, 144, 1–11. [Google Scholar] [CrossRef]
- Powney, G.D.; Carvell, C.; Edwards, M.; Morris, R.K.A.; Roy, H.E.; Woodcock, B.A.; Isaac, N.J.B. Widespread losses of pollinating insects in Britain. Nat. Commun. 2019, 10, 118. [Google Scholar] [CrossRef]
- Sutherland, G.D.; Gass, C.L. Learning and remembering of spatial patterns by hummingbirds. Anim. Behav. 1995, 50, 1273–1286. [Google Scholar] [CrossRef]
- Irwin, R.E.; Brody, A.K. Nectar-robbing bumblebees reduce the fitness of Ipomopsis aggregata (Polemoniaceae). Ecology 1999, 80, 1703–1712. [Google Scholar] [CrossRef]
- Schemske, D.W.; Bradshaw, H.D. Pollinator preference and the evolution of floral traits in monkey flowers (Mimulus). Proc. Natl Acad. Sci. USA 1999, 96, 11910–11915. [Google Scholar] [CrossRef] [PubMed]
- Chalcoff, V.R.; Gleiser, G.; Ezcurra, C.; Aizen, M.A. Pollinator type and secondarily climate are related to nectar sugar composition across the angiosperms. Evol. Ecol. 2017, 31, 585–602. [Google Scholar] [CrossRef]
- Janeček, S.; Chmel, K.; Ewome, F.L.; Hrubá, K.; Klomberg, Y.; Kobe, I.N.; Kouede, R.D.; Mertens, J.E.J.; Njie, M.M.; Tropek, R. Differences in Nectar Traits between Ornithophilous and Entomophilous Plants on Mount Cameroon. Plants 2021, 10, 1161. [Google Scholar] [CrossRef]
- Tew, N.E.; Memmott, J.; Vaughan, I.P.; Bird, S.; Stone, G.N.; Potts, S.G.; Baldock, K.C.R. Quantifying nectar production by flowering plants in urban and rural landscapes. J. Ecol. 2021, 109, 1747–1757. [Google Scholar] [CrossRef]
- Tew, N.E.; Baldock, K.C.R.; Morten, J.M.; Bird, S.; Vaughan, I.P.; Memmott, J. A dataset of nectar sugar production for flowering plants found in urban green spaces. Ecol. Solut. Evid. 2023, 4, e1224. [Google Scholar] [CrossRef]
- Baldock, K.C.R.; Goddard, M.A.; Hicks, D.M.; Kunin, W.E.; Mitschunas, N.; Morse, H.; Osgathorpe, L.M.; Potts, S.G.; Robertson, K.M.; Scott, A.V.; et al. A systems approach reveals urban pollinator hotspots and conservation opportunities. Nat. Ecol. Evol. 2019, 3, 363–373. [Google Scholar] [CrossRef]
- Baldock, K.C.R.; Goddard, M.A.; Hicks, D.M.; Kunin, W.E.; Mitschunas, N.; Osgathorpe, L.M.; Potts, S.G.; Robertson, K.M.; Scott, A.V.; Stone, G.N.; et al. Where is the UK’s pollinator biodiversity? The importance of urban areas for flower-visiting insects. Proc. R. Soc. B Biol. Sci. 2015, 282, 20142849. [Google Scholar] [CrossRef]
- Dunnett, N.; Swanwick, C.; Woolley, H. Improving Urban Parks, Play Areas and Green Spaces; Report for Department for Transport, Local Government and the Regions: London, UK, 2002. [Google Scholar]
- Haq, S.M.A. Urban Green Spaces and an Integrative Approach to Sustainable Environment. J. Environ. Prot. 2011, 2, 601–608. [Google Scholar] [CrossRef]
- Tan, P.Y.; Wang, J.; Sia, A. Perspectives on five decades of the urban greening of Singapore. Cities 2013, 32, 24–32. [Google Scholar] [CrossRef]
- Yessoufou, K.; Sithole, M.; Elansary, H.O. Effects of urban green spaces on human perceived health improvements: Provision of green spaces is not enough but how people use them matters. PLoS ONE 2020, 15, e0239314. [Google Scholar] [CrossRef] [PubMed]
- Phogole, B.; Yessoufou, K. A Global Meta-Analysis of Effects of Green Infrastructure on COVID-19 Infection and Mortality Rates. medRxiv 2023. [Google Scholar] [CrossRef]
- Richardson, E.A.; Pearce, J.; Mitchell, R.; Kingham, S. Role of physical activity in the relationship between urban green space and health. Public Health 2013, 127, 318–324. [Google Scholar] [CrossRef]
- Maas, J.; Verheij, R.A.; Groenewegen, P.P.; de Vries, S.; Spreeuwenberg, P. Green space, urbanity, and health: How strong is the relation? J. Epidemiol. Community Health 2006, 60, 587–592. [Google Scholar] [CrossRef]
- de Vries, S.; van Dillen, S.M.E.; Groenewegen, P.P.; Spreeuwenberg, P. Streetscape greenery and health: Stress, social cohesion and physical activity as mediators. Soc. Sci. Med. 2013, 94, 26–33. [Google Scholar] [CrossRef]
- White, M.P.; Alcock, I.; Wheeler, B.W.; Depledge, M.H. Would You Be Happier Living in a Greener Urban Area? A Fixed-Effects Analysis of Panel Data. Psychol. Sci. 2013, 24, 920–928. [Google Scholar] [CrossRef]
- Diener, A.; Mudu, P. How can vegetation protect us from air pollution? A critical review on green spaces’ mitigation abilities for air-borne particles from a public health perspective—With implications for urban planning. Sci. Total. Environ. 2021, 796, 148605. [Google Scholar] [CrossRef]
- Li, Q. Effect of forest bathing trips on human immune function. Environ. Health Prev. Med. 2010, 15, 9–17. [Google Scholar] [CrossRef]
- Vivier, E.; Tomasello, E.; Baratin, M.; Walzer TUgolini, S. Functions of natural killer cells. Nat. Immunol. 2008, 9, 503–510. [Google Scholar] [CrossRef]
- Bringslimark, T.; Hartig, T.; Patil, G.G. The psychological benefits of indoor plants: A critical review of the experimental literature. J. Environ. Psychol. 2009, 29, 422–433. [Google Scholar] [CrossRef]
- Angioy, A.M.; Desogus, A.; Barbarossa, I.T.; Anderson, P.; Hansson, B.S. Extreme sensitivity in an olfactory system. Chem. Senses 2003, 28, 279–284. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Gibson, M.R.; Richardson, D.M.; Pauw, A. Can floral traits predict an invasive plant’s impact on native plant–pollinator communities? J. Ecol. 2012, 100, 1216–1223. [Google Scholar] [CrossRef]
- Waser, N.M. Interspecific pollen transfer and competition between co-occurring plant species. Oecologia 1978, 36, 223–236. [Google Scholar] [CrossRef]
- Brown, B.J.; Mitchell, R.J. Competition for pollination: Effects of pollen of an invasive plant on seed set of a native congener. Oecologia 2001, 129, 43–49. [Google Scholar] [CrossRef]
- Tscheulin, T.; Petanidou, T.; Potts, S.G.; Settele, J. The impact of Solanum elaeagnifolium, an invasive plant in the Mediterranean, on the flower visitation and seed set of the native co-flowering species Glaucium flavum. Plant Ecol. 2009, 205, 77–85. [Google Scholar] [CrossRef]
- Morales, C.L.; Traveset, A. A meta-analysis of impacts of alien vs. native plants on pollinator visitation and reproductive success of co-flowering native plants. Ecol. Lett. 2009, 12, 716–728. [Google Scholar] [CrossRef]
- Yessoufou, K.; Daru, B.H.; Muasya, A.M. Phylogenetic exploration of commonly used medicinal plants in South Africa. Mol. Ecol. Resour. 2015, 15, 405–413. [Google Scholar] [CrossRef]
- Canavan, S.; Richardson, D.M.; Visser, V.; Le Roux, J.J.; Vorontsova, M.S.; Wilson, J.R.U. The global distribution of bamboos: Assessing correlates of introduction and invasion. AoB Plants 2016, 9. [Google Scholar] [CrossRef]
- Yessoufou, K.; Ambani, A.E. Are Introduced Alien Species More Predisposed to Invasion in Recipient Environments if They Provide a Wider Range of Services to Humans? Diversity 2021, 13, 553. [Google Scholar] [CrossRef]
- Lockwood, J.L.; Cassey, P.; Blackburn, T. The role of propagule pressure in explaining species invasions. Trends Ecol. Evol. 2005, 20, 223–228. [Google Scholar] [CrossRef]
- Blackburn, T.M.; Duncan, R.P. Determinants of establishment success in introduced birds. Nature 2001, 414, 195–197. [Google Scholar] [CrossRef] [PubMed]
- Carlton, J.T. Pattern, process, and prediction in marine invasion ecology. Biol. Conserv. 1996, 78, 97–106. [Google Scholar] [CrossRef]
- Cruden, R.W.; Hermann, S.M.; Peterson, S. Patterns of nectar production and plant animal coevolution. In The Biology of Nectaries; Bentley, B., Elias, T., Eds.; Columbia University Press: New York, NY, USA, 1983; pp. 126–152. [Google Scholar]
- Pleasants, J.M. Nectar production in Ipomopsis aggregata (Polemoniaceae). Am. J. Bot. 1983, 70, 1468–1475. [Google Scholar] [CrossRef]
- Pyke, G.H.; Waser, N.M. The Production of Dilute Nectars by Hummingbird and Honeyeater Flowers. Biotropica 1981, 13, 260. [Google Scholar] [CrossRef]
- Lotz, C.N.; Schondube, J.E. Sugar preferences in nectar and fruit-eating birds: Behavioural patterns and physiological causes. Biotropica 2006, 38, 3–15. [Google Scholar] [CrossRef]
- Ornelas, J.F.; Ordano, M.; De-Nova, A.J.; Quintero, M.E.; Garland, T. Phylogenetic analysis of interspecific variation in nectar of hummingbird-visited plants. J. Evol. Biol. 2007, 20, 1904–1917. [Google Scholar] [CrossRef]
- Mitchell, R.J. Heritability of nectar traits: Why do we know so little? Ecology 2004, 85, 1527–1533. [Google Scholar] [CrossRef]
- Parachnowitsch, A.L.; Manson, J.S.; Sletvold, N. Evolutionary ecology of nectar. Ann. Bot. 2018, 123, 247–261. [Google Scholar] [CrossRef]
- Percival, M.S. Types of nectar in angiosperms. New Phytol. 1961, 60, 235–281. [Google Scholar] [CrossRef]
- Tavares, D.C.; Freitas, L.; Gaglianone, M.C. Nectar volume is positively correlated with flower size in hummingbird-visited flowers in the Brazilian Atlantic Forest. J. Trop. Ecol. 2016, 32, 335–339. [Google Scholar] [CrossRef]
- Faegri, K.; van der Pijl, L. The Principles of Pollination Ecology; Pergamon Press: Oxford, UK, 1979. [Google Scholar]
- Jin, Y.; Qian, H.V. PhyloMaker: An R package that can generate very large phylogenies for vascular plants. Ecography 2019, 42, 1353–1359. [Google Scholar] [CrossRef]
- Smith, S.A.; Brown, J.W. Constructing a broadly inclusive seed plant phylogeny. Am. J. Bot. 2018, 105, 302–314. [Google Scholar] [CrossRef] [PubMed]
- Zanne, A.E.; Tank, D.C.; Cornwell, W.K.; Eastman, J.M.; Smith, S.A.; FitzJohn, R.G.; McGlinn, D.J.; O’meara, B.C.; Moles, A.T.; Reich, P.B.; et al. Three keys to the radiation of angiosperms into freezing environments. Nature 2014, 506, 89–92. [Google Scholar] [CrossRef] [PubMed]
- R Development Core Team. R: A Language and Environment for Statistical Computing. 2023. Available online: https://www.r-project.org/ (accessed on 15 May 2021).
- Revell, L.J. phytools: An R package for phylogenetic comparative biology (and other things). Methods Ecol. Evol. 2012, 3, 217–223. [Google Scholar] [CrossRef]
- Felsenstein, J. Phylogenies and the Comparative Method. Am. Nat. 1985, 125, 1–15. [Google Scholar] [CrossRef]
- Garland, T., Jr.; Dickerman, A.W.; Janis, C.M.; Jones, J.A. Phyloge netic analysis of covariance by computer simulation. Syst. Biol. 1993, 42, 265–292. [Google Scholar] [CrossRef]
- Blomberg, S.P.; Garland, T., Jr.; Ives, A.R. Testing for phylogenetic signal in comparative data: Behavioural traits are more labile. Evolution 2003, 57, 717–745. [Google Scholar]
- Kemel, S.W.; Cowan, P.D.; Helmus, M.R.; Cornwell, W.K.; Morlon, H.; Ackerly, D.D.; Blomberg, S.P.; Webb, C.O. Picante: R tools for integrating phylogenies and ecology. Bioinformatics 2010, 26, 1463–1464. [Google Scholar] [CrossRef]


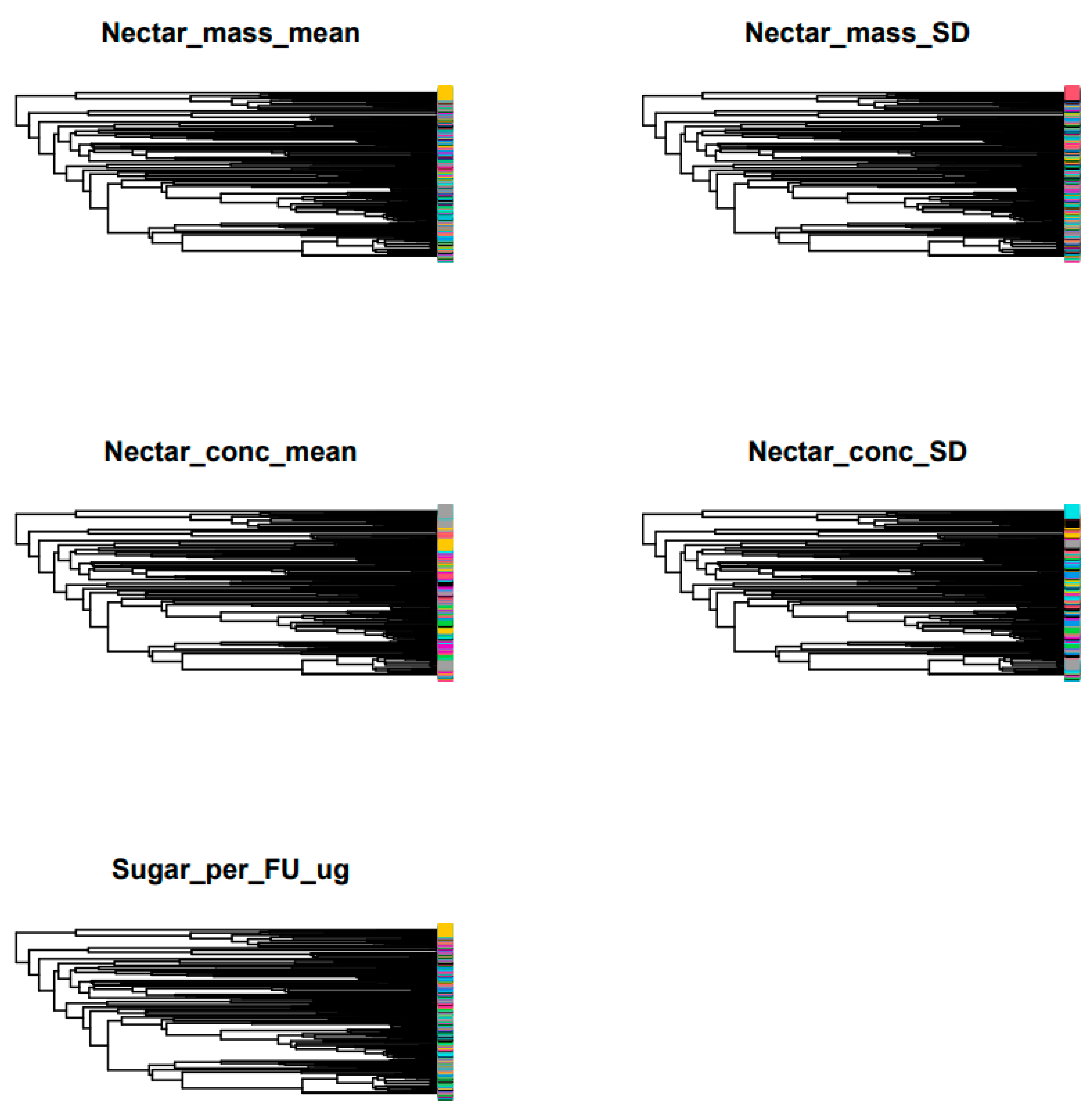{kind=link}
| Nectar Production | K | PIC.var.obs. | PIC var.rnd.mean | p-Value | PIC.var.Z |
|---|---|---|---|---|---|
| Nectar_mass_mean | 0.178063154 | 143,425.8953 | 443,998.8927 | 0.062 | −0.384427948 |
| Nectar_mass_SD | 0.223061818 | 41,689.07254 | 193,459.6056 | 0.003 ** | −0.55471627 |
| Nectar_conc_mean | 0.179022884 | 9.094378937 | 14.9308214 | 0.039 * | −1.421055868 |
| Nectar_conc_SD | 0.130033662 | 1.489374074 | 1.749948461 | 0.417 | −0.352449477 |
| Sugar_per_FU_ug | 0.110024302 | 640,764.1636 | 1,389,605.143 | 0.276 | −0.39843441 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yessoufou, K. The Patterns of Intraspecific Variations in Mass of Nectar Sugar along a Phylogeny Distinguish Native from Non-Native Plants in Urban Greenspaces in Southern England. Plants 2023, 12, 3270. https://doi.org/10.3390/plants12183270
Yessoufou K. The Patterns of Intraspecific Variations in Mass of Nectar Sugar along a Phylogeny Distinguish Native from Non-Native Plants in Urban Greenspaces in Southern England. Plants. 2023; 12(18):3270. https://doi.org/10.3390/plants12183270
Chicago/Turabian StyleYessoufou, Kowiyou. 2023. "The Patterns of Intraspecific Variations in Mass of Nectar Sugar along a Phylogeny Distinguish Native from Non-Native Plants in Urban Greenspaces in Southern England" Plants 12, no. 18: 3270. https://doi.org/10.3390/plants12183270
APA StyleYessoufou, K. (2023). The Patterns of Intraspecific Variations in Mass of Nectar Sugar along a Phylogeny Distinguish Native from Non-Native Plants in Urban Greenspaces in Southern England. Plants, 12(18), 3270. https://doi.org/10.3390/plants12183270