
Chinee Apple (Ziziphus mauritiana): A Comprehensive Review of Its Weediness, Ecological Impacts and Management Approaches




Abstract
:1. Introduction
2. Historical and Global Significance of Ziziphus mauritiana Fruit Trees
3. Botany, Biology, and Ecology of Ziziphus mauritiana
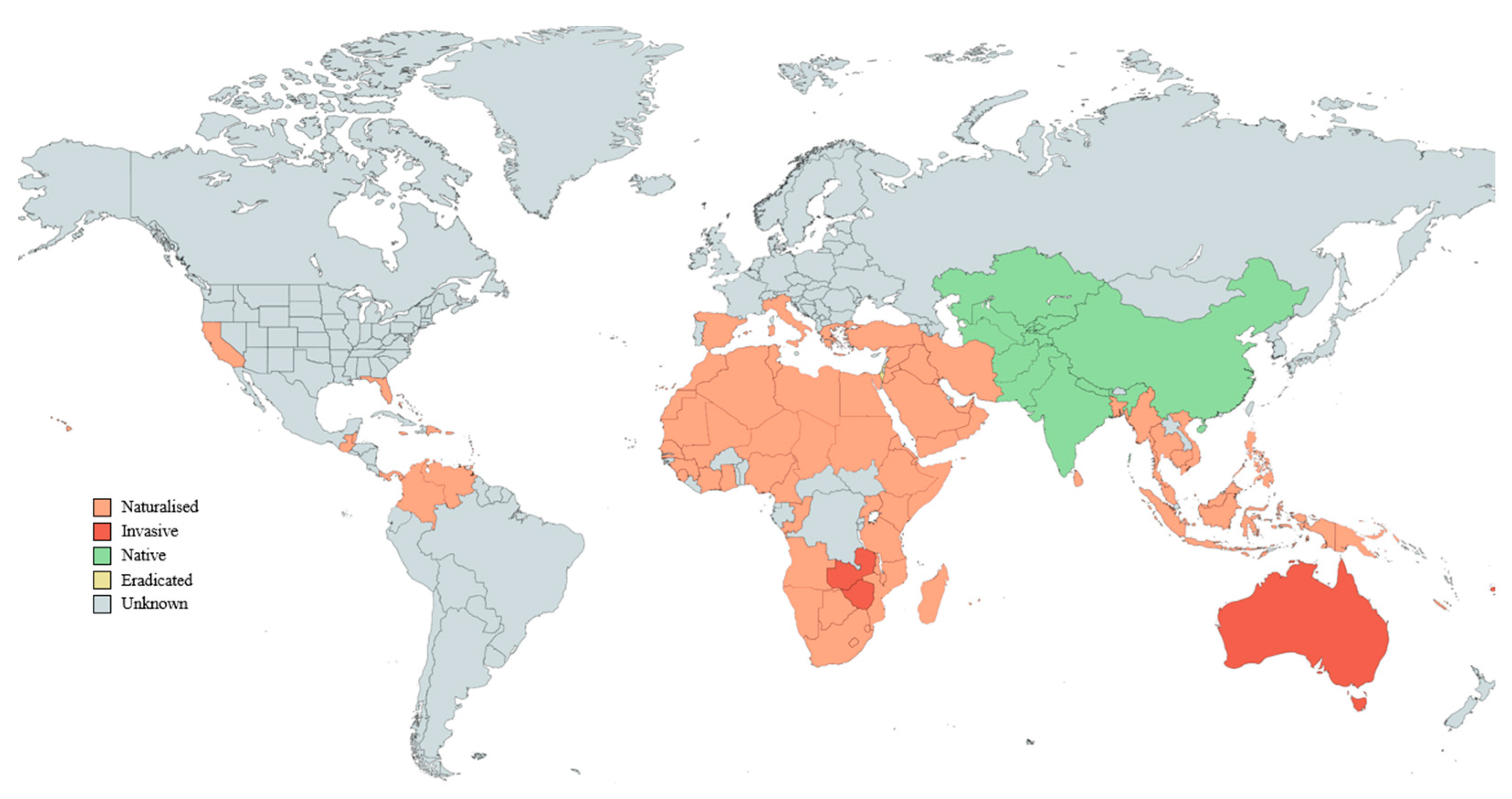
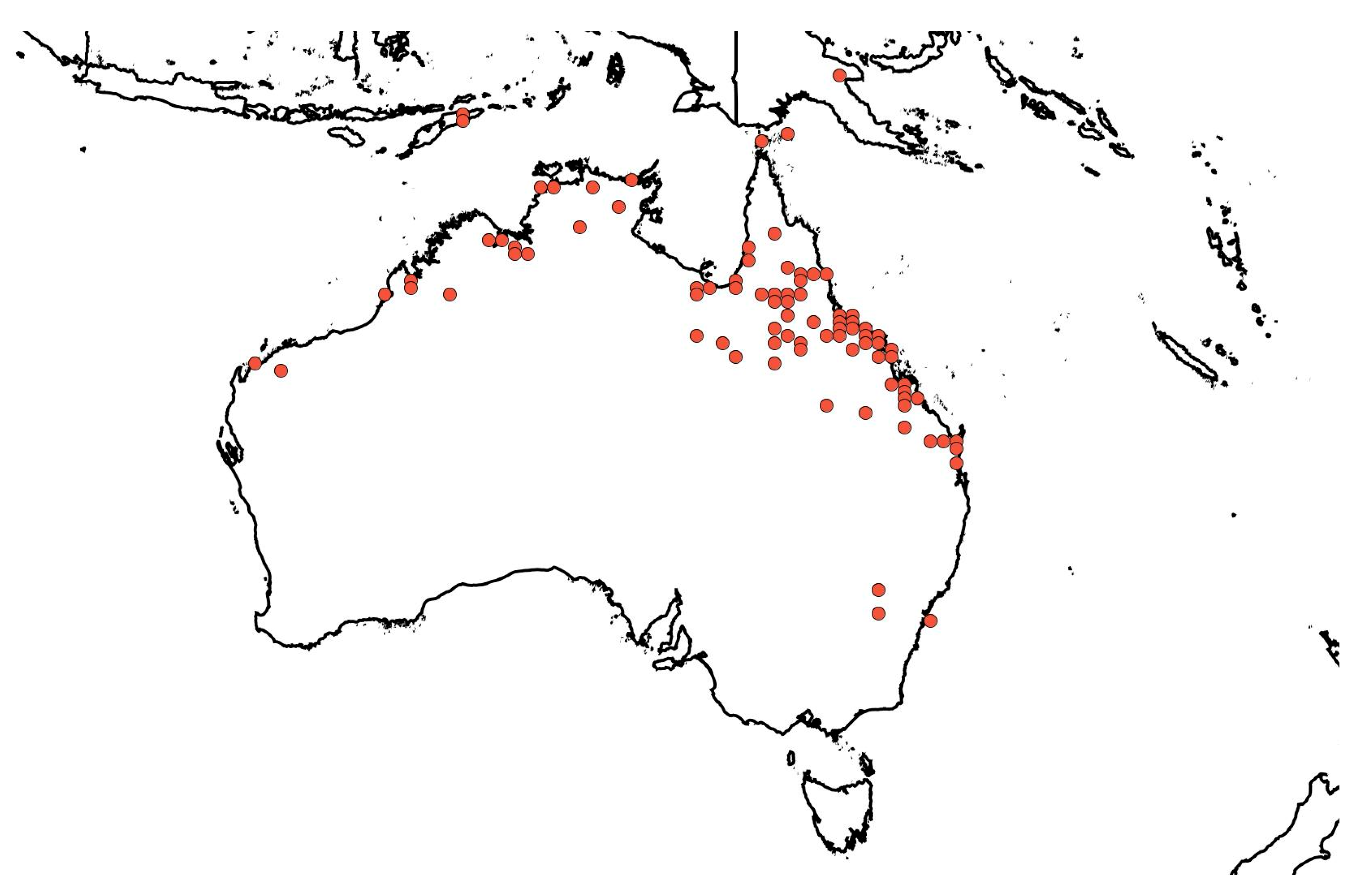
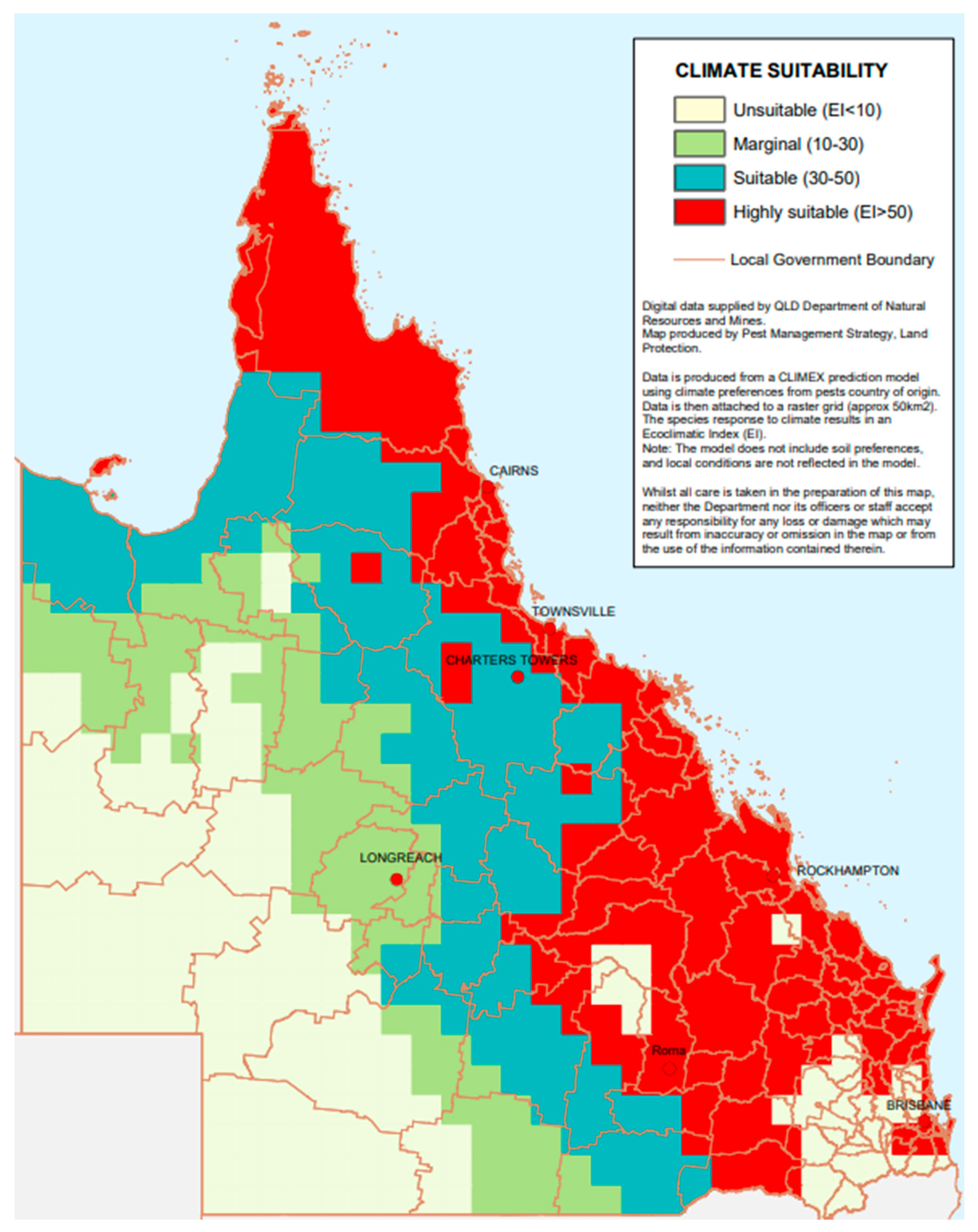
4. Native and Naturalised Distribution of Ziziphus mauritiana
5. Agricultural, Environmental, and Economic Impacts of Ziziphus mauritiana
6. Management of Invasive Ziziphus mauritiana in Australia
6.1. Manual and Mechanical Control Methods
6.2. Chemical Control Methods
6.3. Biological Control Methods
7. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Grice, A.C. Seed Production, Dispersal and Germination in Cryptostegia grandiflora and Ziziphus mauritiana, Two Invasive Shrubs in Tropical Woodlands of Northern Australia. Aust. J. Ecol. 1996, 21, 324–331. [Google Scholar] [CrossRef]
- Grice, A.C.; Radford, I.J.; Abbott, B.N. Regional and Landscape-Scale Patterns of Shrub Invasion in Tropical Savannas. Biol. Invasions 2000, 2, 187–205. [Google Scholar] [CrossRef]
- Bebawi, F.F.; Campbell, S.D.; Mayer, R.J. Seed Bank Persistence and Germination of Chinee Apple (Ziziphus mauritiana Lam.). Rangel. J. 2016, 38, 17–25. [Google Scholar] [CrossRef]
- Liang, T.; Sun, W.; Ren, H.; Ahmad, I.; Vu, N.; Huang, M.; Huang, J. Genetic Diversity of Ziziphus mauritiana Germplasm Based on SSR Markers and Ploidy Level Estimation. Planta 2019, 249, 1875–1887. [Google Scholar] [CrossRef] [PubMed]
- O’Brien, C.O.; Campbell, S.; Vogler, W.; Galea, V.J. Evaluation of Di-Bak® Herbicide Capsule System for Control of Chinee Apple (Ziziphus mauritiana) in North Queensland. In Proceedings of the 22nd Australasian Weeds Conference, Adelaide, SA, Australia, 25–29 September 2022; Council of Australasian Weed Societies Inc. (CAWS): Adelaide, SA, Australia, 2022. [Google Scholar]
- Muhammad, N.; Luo, Z.; Yang, M.; Liu, Z.; Liu, M. The Nutritional, Medicinal, and Drought-Resistance Properties of Ziziphus Mill. Make It an Important Candidate for Alleviating Food Insecurity in Arid Regions—A Case of Pakistan. Horticulturae 2022, 8, 867. [Google Scholar] [CrossRef]
- Dhileepan, K. Biological Control of Ziziphus mauritiana (Rhamnaceae): Feasibility, Prospective Agents, and Research Gaps. Ann. Appl. Biol. 2017, 170, 287–300. [Google Scholar] [CrossRef]
- Nyoka, B.I. Biosecurity in Forestry: A Case Study on the Status of Invasive Forest Species in South Africa; Forestry Department of Food and Agriculture Organisation (FAO): Rome, Italy, 2003. [Google Scholar]
- Weber, E. Invasive Plant Species of the World: A Reference Guide to Environmental Weeds, 2nd ed.; CABI International: Wallingford, UK, 2003; p. 548. [Google Scholar]
- Grice, A.C. Weeds and the Monitoring of Biodiversity in Australian Rangelands. Austral Ecol. 2004, 29, 51–58. [Google Scholar] [CrossRef]
- Ani, O.; Onu, O.; Okoro, G.; Uguru, M. Chapter 2: Overview of Biological Methods of Weed Control. In Biological Approaches for Controlling Weeds; Radhakrishnan, R., Ed.; InTechOpen: London, England, 2018; pp. 5–16. [Google Scholar]
- Vitelli, J.S. Options for Effective Weed Management. Trop. Grassl 2000, 34, 280–294. [Google Scholar]
- Grice, A.C. Ecology in the Management of Indian Jujube (Ziziphus mauritiana). Weed Sci. 1998, 46, 467–474. [Google Scholar] [CrossRef]
- Ward-Fear, G.; Brown, G.P.; Pearson, D.J.; Shine, R. An Invasive Tree Facilitates the Persistence of Native Rodents on an Over-Grazed Floodplain in Tropical Australia. Austral Ecol. 2017, 42, 385–393. [Google Scholar] [CrossRef]
- Laguna, J.M.; Reside, A.E.; Kutt, A.; Grice, A.C.; Buosi, P.; Vanderduys, E.P.; Taylor, M.; Schwarzkopf, L. Conserving the Endangered Black-Throated Finch Southern Subspecies: What Do We Need to Know? Emu—Austral Ornithol. 2019, 119, 331–345. [Google Scholar] [CrossRef]
- Northern Territory Department of Environment, Parks and Water Security. Chinee Apple Management Guide. Available online: https://nt.gov.au/__data/assets/pdf_file/0019/231418/chinee-apple-management-guide.pdf (accessed on 9 August 2020).
- Abbas, T.; Zahir, Z.A.; Naveed, M.; Kremer, R.J. Limitations of Existing Weed Control Practices Necessitate Development of Alternative Techniques Based on Biological Approaches. Adv. Agron. 2018, 147, 239–280. [Google Scholar] [CrossRef]
- Moss, S.R. Non-Chemical Methods for Weed Control: Benefits and Limitations. In Proceedings of the 17th Australasian Weeds Conference, Christchurch, New Zealand, 26–30 September 2010; New Zealand Plant Protection Society: Karekare, New Zealand, 2010. [Google Scholar]
- O’Brien, C.J.; Mellor, V.; Galea, V.J. Controlling Woody Weed Chinese Elm (Celtis sinensis Pers.) with Stem-Implanted Herbicide Capsules. Plants 2022, 11, 444. [Google Scholar] [CrossRef] [PubMed]
- Marzuza, I.M.; Musemwa, L.; Mapurazi, S.; Matsika, P.; Munyati, V.T.; Ndhleve, S. Future Prospects of Ziziphus mauritiana in Alleviating Household Food Insecurity and Illness in Arid and Semi-Arid Areas: A Review. World Dev. Perspect. 2017, 5, 1–6. [Google Scholar] [CrossRef]
- Sharif, M.; Naz, A.; Farooq, U.; Razzaq, K.; Sher, M.A.; Khan, M.A.; Waheed, U.; Hayder, Q.; Naz, N.; Ahmad, S.; et al. A Comprehensive Investigation of Novel Ber (Ziziphus mauritiana) Products From South Punjab, Pakistan. J. Bioresour. Manag. 2022, 9, 15–31. [Google Scholar]
- Grice, A.C. The Biology of Australian Weeds—39. Ziziphus mauritiana Lam. Plant Prot. Q. 2002, 17, 2–11. [Google Scholar]
- Meghwal, P.R.; Khan, M.A.; Tewari, J.C. Growing Ber (Ziziphus mauritiana Lam.) for Sustainable Income and Employment in Arid and Semi-Arid Regions; ICAR—Central Arid Zone Research Institute (CAZRI): Jodhpur, India, 2007. [Google Scholar]
- Janick, J.; Paull, R.E. The Encyclopaedia of Fruit and Nuts; CABI: Oxfordshire, UK, 2008; p. 617. [Google Scholar]
- Mishra, T.; Paice, A.G.; Bhatia, A. Chapter 87: Use of Seeds of Malay Apple (Ziziphus mauritiana) and Related Species in Health and Disease. In Nuts and Seeds in Health and Disease Prevention; Preedy, V.R., Watson, R.R., Patel, V.B., Eds.; Academic Press: Cambridge, MA, USA, 2011; pp. 733–739. [Google Scholar]
- Pareek, S.; Yahia, E.M. Postharvest Biology and Technology of Ber Fruit. In Horticultural Reviews; Janick, J., Ed.; John Wiley & Sons: Hoboken, NJ, USA, 2013; pp. 201–231. [Google Scholar]
- Tel-Zur, N.; Schneider, B. Floral Biology of Ziziphus mauritiana (Rhamnaceae). Sex. Plant Reprod. 2009, 22, 73–85. [Google Scholar] [CrossRef]
- Pareek, S.; Mukherjee, S.; Paliwal, R. Floral Biology of Ber—A Review. Agric. Revolut. 2007, 28, 277–282. [Google Scholar]
- Verma, S.S.; Verma, R.P.; Verma, S.K.; Yadav, A.L.; Verma, A.K. Responses of Ber (Ziziphus mauritiana Lamk.) Varieties to Different Level of Salinity. Int. J. Curr. Microbiol. App. Sci. 2018, 7, 580–591. [Google Scholar] [CrossRef]
- Clifford, S.C.; Arndt, S.K.; Corlett, J.E.; Joshi, S.; Sankhla, N.; Popp, M.; Jones, H.G. The Role of Solute Accumulation, Osmotic Adjustment and Changes in Cell Wall Elasticity in Drought Tolerance in Ziziphus mauritiana (Lamk.). J. Exp. Bot. 1998, 49, 967–977. [Google Scholar] [CrossRef]
- Bhatt, M.J.; Patel, A.D.; Bhatti, P.M.; Pandey, A.N. Effect of Soil Salinity on Growth, Water Status and Nutrient Accumulation in Seedlings of Ziziphus mauritiana (Rhamnaceae). J. Fruit Ornam. Plant Res. 2008, 16, 383–401. [Google Scholar]
- Prakash, O.; Usmani, S.; Singh, R.; Singh, N.; Gupta, A.; Ved, A. A Panoramic View on Phytochemical, Nutritional and Therapeutic Attributes of Ziziphus mauritiana Lam.: A Comprehensive Review. Phytother. Res. 2020, 35, 63–77. [Google Scholar] [CrossRef]
- Naaz, F.; Agari, N.; Singh, A. Medicinal Properties of Ziziphus mauritiana: A Review Article. Int. J. Pharm. Pharm. Res. 2020, 19, 219–230. [Google Scholar]
- Dahiru, D.; Sini, J.M.; John-Africa, L. Antidiarrheal Activity of Ziziphus mauritiana Root Extract in Rodents. Afr. J. Biotechnol. 2006, 5, 941–945. [Google Scholar]
- Maaiden, E.E.; Kharrassi, Y.E.; Qarah, N.A.S.; Essamadi, A.K.; Moustaid, K.; Nasser, B. Genus Ziziphus: A Comprehensive Review on Ethnopharmacological, Phyto-chemical and Pharmacological Properties. J. Ethnopharmacol. 2020, 259, 112950. [Google Scholar] [CrossRef] [PubMed]
- Butt, S.Z.; Hussain, S.; Munawar, K.S. Phytochemistry of Ziziphus mauritiana; Its Nutritional and Pharmaceutical Potential. Sci Inq. Rev. 2021, 5, 1–15. [Google Scholar] [CrossRef]
- Morton, J. Indian Jujube. In Fruits of Warm Climates; Morton, J.F., Ed.; Echo Point Books & Media: Brattleboro, VT, USA, 1987; pp. 272–275. [Google Scholar]
- Spinoni, J.; Barbosa, P.; Cherlet, M.; Forzieri, G.; McCormick, N.; Naumann, G.; Vogt, J.V.; Dosio, A. How Will the Progressive Global Increase of Arid Areas Affect Population and Land-Use in the 21st Century? Glob Planet Change 2021, 205, 103597. [Google Scholar] [CrossRef]
- Crawford, R.; Shan, F.; McCarthy, A. Chinese Jujube: A Developing Industry in Australia. Acta Hortic. 2013, 993, 29–36. [Google Scholar] [CrossRef]
- Johnstone, R. Development of the Chinese Jujube Industry in Australia; Australian Government Rural Industries Research and Development Corporation: Barton, ACT, Australia, 2014. [Google Scholar]
- Johnstone, R.M.; Shan, F. Chinese Jujube Industry Takes Root in Western Australia. Acta Hortic. 2016, 1116, 31–34. [Google Scholar] [CrossRef]
- Johnstone, R. Overcoming Barriers to Development of the Australian Jujube Industry; AgriFutures Australia: New South Wales, Australia, 2017. [Google Scholar]
- ABC Landline. Jujube Expansion: Australia’s Biggest Kiwi Grower Invests in Jujubes. Available online: https://www.abc.net.au/news/rural/programs/landline/2023-05-07/jujube-expansion:-australias-biggest-kiwi-fruit/102314704 (accessed on 12 July 2023).
- Pasieznik, N. Ziziphus Mauritiana (Jujube); CABI Compendium: Wallingford, UK, 2007. [Google Scholar]
- Dight, I. Burdekin Water Quality Improvement Plan; Burdekin Solutions Ltd.: Townsville, Australia, 2009. [Google Scholar]
- Arndt, S.K.; Wanek, W.; Clifford, S.C.; Popp, M. Contrasting Adaptations to Drought Stress in Field-Grown Ziziphus mauritiana and Prunus persica Trees: Water Relations, Osmotic Adjustment and Carbon Isotope Composition. Aust. J. Plant Physiol. 2000, 27, 985–996. [Google Scholar] [CrossRef]
- Atlas of Florida Plants: Institute for Systematic Botany. Ziziphus mauritiana. Available online: https://florida.plantatlas.usf.edu/Plant.aspx?id=255 (accessed on 3 May 2023).
- CABI. Forestry Compendium; CABI: Wallingford, UK, 2005. [Google Scholar]
- Orwa, C.; Mutua, A.; Kindt, R.; Jamnadass, R.; Simons, A. Agroforestree Database: A Tree Reference and Selection Guide (Version 4.0); World Agroforestry Centre: Kenya, Africa, 2009. [Google Scholar]
- Pacific Islands Ecosystems at Risk (PIER). Available online: https://www.hear.org.au/pier/index.html (accessed on 5 May 2023).
- Atlas of Living Australia (ALA). Available online: https://biocache.ala.org.au/occurrences/search?q=lsid%3Ahttps%3A%2F%2Fid.biodiversity.org.au%2Fnode%2Fapni%2F2894258#tab_mapView (accessed on 6 May 2023).
- Pest Management Strategy. Available online: https://www.daf.qld.gov.au/__data/assets/pdf_file/0007/50389/IPA-Chinee-Apple-PM.pdf (accessed on 8 May 2023).
- Kriticos, D.J.; Morin, L.; Leriche, A.; Anderson, R.C.; Caley, P. Combining a Climatic Niche Model of an Invasive Fungus with its Host Species Distributions to Identify Risks to Natural Assets: Puccinia psidii Sensu Lato in Australia. PLoS ONE 2013, 8, e64479. [Google Scholar] [CrossRef] [PubMed]
- Byeon, D.H.; Jung, S.; Lee, W.H. Review of CLIMEX and MaxEnt for Studying Species Distribution in South Korea. J. Asia-Pac. Biodivers 2018, 11, 325–333. [Google Scholar] [CrossRef]
- Kour, S.; Khurma, U.; Brodie, G.; Singh, S. Modelling the Potential Global Distribution of Suitable Habitat for the Biological Control Agent Heterorhabditis indica. Ecol. Evol. 2022, 12, e8997. [Google Scholar] [CrossRef] [PubMed]
- Jones, R.; Alemseged, Y.; Medd, R.; Vere, D. The Distribution, Density and Economic Impact of Weeds in the Australian Annual Winter Cropping System; CRC for Weed Management Systems: Glen Osmond, Australia, 2000. [Google Scholar]
- Walker, P.T. Crop Losses: The Need to Quantify the Effect of Pests, Disease and Weeds on Agricultural Production. Agric Ecosyst Env. 1983, 9, 119–158. [Google Scholar] [CrossRef]
- Grice, A.C. Post-Fire Regrowth and Survival of the Invasive Tropical Shrubs Cryptostegia grandiflora and Ziziphus mauritiana. Aust. J. Ecol. 1997, 22, 49–55. [Google Scholar] [CrossRef]
- Queensland Department of Agriculture and Fisheries. Restricted Plants of Queensland. Available online: https://www.daf.qld.gov.au/__data/assets/pdf_file/0004/383818/restricted-invasive-plants.pdf (accessed on 16 June 2023).
- Northern Territory Department of Environment, Parks and Water Security. Weed Management Plan for Chinee Apple 2012–2022. Available online: https://nt.gov.au/__data/assets/pdf_file/0020/231419/chinee-apple-weed-management-plan.PDF (accessed on 16 June 2023).
- Western Australia Department of Primary Industries and Regional Development. Ziziphus mauritiana Lam. Available online: https://www.agric.wa.gov.au/organisms/119159 (accessed on 16 June 2023).
- Queensland Government. Area Management Plans (AMPs). Available online: https://www.qld.gov.au/environment/land/management/vegetation/clearing-approvals/area-management-plans (accessed on 14 August 2023).
- Queensland Government. Legislation: Vegetation Management Act 1999. Available online: https://www.stateoftheenvironment.des.qld.gov.au/biodiversity/management-responses/legislation/vegetation-management-act-1999#:~:text=The%20Vegetation%20Management%20Act%201999,prevents%20loss%20of%20biodiversity (accessed on 14 August 2023).
- Queensland Department of Natural Resources and Mines. Area Management Plan for the Control of Pest Plants in the Dry Tropics Region; Queensland Government: Brisbane, Australia, 2015. [Google Scholar]
- Martin-Forés, I.; Guerin, G.R.; Lowe, A.J. Weed Abundance is Positively Correlated With Native Plant Diversity in Grasslands of Southern Australia. PLoS ONE 2017, 12, e0178681. [Google Scholar] [CrossRef]
- Fridley, J.D.; Stachowicz, J.J.; Naeem, S.; Sax, D.F.; Seabloom, E.W.; Smith, M.D.; Stohlgren, T.J.; Tilman, D.; Von Holle, B. The Invasion Paradox: Reconciling Pattern and Process in Species Invasions. Ecology 2007, 88, 3–17. [Google Scholar] [CrossRef]
- Zull, A.; Cacho, O.; Lawes, R. A Matrix Model for the Management of Perennial Weeds in the North Queensland Rangelands System: Application to Ziziphus mauritiana (Lam). In Proceedings of the 15th Australian Weeds Conference, Adelaide, SA, Australia, 24–28 September 2006; Weed Management Society of South Australia: Torrens Park, SA, Australia, 2006; pp. 671–674. [Google Scholar]
- Zull, A.F.; Cacho, O.J.; Lawes, R.A. Optimising the Control of Rangeland Woody Weeds. In Proceedings of the 17th Australasian Weeds Conference, Christchurch, New Zealand, 26–30 September 2010; New Zealand Plant Protection Society: Karekare, New Zealand, 2010; pp. 414–417. [Google Scholar]
- Zull, A.F.; Cacho, O.J.; Lawes, R.A. Optimising Woody-Weed Control. In Proceedings of the 53rd Australian Agricultural and Resource Economics Society (AARES) Annual Conference, Cairns, Queensland, Australia, 11–13 February 2009; Australian Agricultural and Resource Economics Society: Vermont, VIC, Australian, 2009. [Google Scholar]
- Zull, A.F.; Lawes, R.A.; Cacho, O.J. Optimal Frequency for Woody Weed Management for North Queensland Grazing Properties: An Economic Perspective. In Proceedings of the 16th Australian Weeds Conference, Cairns, Queensland, Australia, 18–22 May 2008; Queensland Weed Society: Clifford Gardens, QLD, Australian, 2008. [Google Scholar]
- DiTomaso, J.M.; Brooks, M.L.; Allen, E.B.; Minnich, R.; Rice, P.M.; Kyser, G.B. Control of Invasive Weeds with Prescribed Burning. Weed Technol. 2006, 20, 535–548. [Google Scholar] [CrossRef]
- Queensland Department of Agriculture, Fisheries and Forestry. Fire: Management of Native and Invasive Woody Weeds in the Northern and Southern Gulf. Available online: https://futurebeef.com.au/wp-content/uploads/fire_factsheet_web1.pdf (accessed on 20 June 2023).
- Grice, A.C.; Brown, J.R. Fire and Population Ecology of Invasive Shrubs in the Tropical Woodlands. In Frontiers of Population Biology, Canberra; Floyd, R.B., Sheppard, A.W., De Barro, P.J., Eds.; CSIRO Publishing: Melbourne, Australia, 1996; pp. 589–597. [Google Scholar]
- Nobel, J.C.; Grice, A.C.; MacLeod, N.D.; Mulle, W.J. Integration of Prescribed Fire and Sub-Lethal Chemical Defoliation for Controlling Shrub Populations in Australian Semi-Arid Woodlands. In Proceedings of the First International Weed Control Conference, Monash University, Melbourne, Australia, 17–21 February 1992; pp. 362–364. [Google Scholar]
- Queensland Government. Chinee Apple (Ziziphus mauritiana). Available online: https://www.daf.qld.gov.au/__data/assets/pdf_file/0008/52766/chinee-apple.pdf (accessed on 23 June 2023).
- Western Australia Department of Primary Industries and Regional Development. Chinee Apple Control. Available online: https://www.agric.wa.gov.au/herbicides/chinee-apple-control (accessed on 23 June 2023).
- Chauhan, B.S.; Abugho, S.B. Effect of Growth Stage on the Efficacy of Postemergence Herbicides on Four Weed Species of Direct-Seeded Rice. Sci. World J. 2012, 2012, 123071. [Google Scholar] [CrossRef]
- Galea, V.J. Use of Stem Implanted Bioherbicide Capsules to Manage an Infestation of Parkinsonia aculeata in Northern Australia. Plants 2021, 10, 1909. [Google Scholar] [CrossRef]
- Galea, V.J. Stem-Implanted Capsules—A Targeted, Efficient and Sustainable Method for Delivery of Synthetic and Biological Agents to Plantation Crops. In Proceedings of the 2nd International Plantation Conference (IPC2021), Online, 9–10 March 2021; Faculty of Plantation and Agrotechnology, Universiti Teknologi MARA: Shah Alam, Malaysia, 2021. [Google Scholar]
- Morin, L. Progress in Biological Control of Weeds with Plant Pathogens. Annu. Rev. Phytopathol. 2020, 58, 201–223. [Google Scholar] [CrossRef]
- Goulter, K.C.; Galea, V.J.; Riikonen, P. Encapsulated Dry Herbicides: A Novel Approach for Control of Trees. In Proceedings of the 21st Australasian Weeds Conference, Sydney, New South Wales, Australia, 9–13 September 2018; The Weed Society of New South Wales Inc.: Muswellbrook, NSW, Australian, 2018. [Google Scholar]
- Limbongan, A.A.; Campbell, S.D.; Galea, V.J. Novel Encapsulated Herbicide Delivery Mechanism: Its Efficacy in Mimosa Bush (Vachellia farnesiana). Plants 2021, 10, 2505. [Google Scholar] [CrossRef]
- Hasan, M.; Ahmad-Hamdani, M.S.; Rosli, A.M.; Hamdan, H. Bioherbicides: An Eco-Friendly Tool for Sustainable Weed Management. Plants 2021, 10, 1212. [Google Scholar] [CrossRef] [PubMed]
- Mirzaee, M.R. An Overview of Jujube (Ziziphus jujuba) Diseases. Arch. Phytopathol. 2014, 47, 82–89. [Google Scholar] [CrossRef]
- Jamadar, M.M.; Balikai, R.A.; Sataraddi, A.R. Status of Diseases on Ber (Ziziphus mauritiana Lamarck) in India and their Management Options. Acta Hortic. 2009, 840, 383–390. [Google Scholar] [CrossRef]
- Mungate, P.N.P.; Masocha, M.; Dube, T. Modelling the Distribution of the Invasive Ziziphus mauritiana Along Road Corridors. Afr. J. Ecol. 2018, 57, 122–129. [Google Scholar] [CrossRef]
- Harding, D.P.; Raizada, M.N. Controlling Weeds with Fungi, Bacteria and Viruses: A Review. Front. Plant. Sci. 2015, 6, 659. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
 |  |  |  |  |
| FRUIT | SEED | LEAVES | BARK | ROOTS |
| Blood Disorders Constipation Dyspepsia Fevers Flatulence Hyperdipsia Indigestion Laxative Leprosy Ulcers Vomiting Wounds | Abdominal Pains Asthma Cough Diarrhea Encephalopathy Insomnia Nausea Ophthalmopathy Rheumatism Sedative | Asthma Blood Diseases Boils Conjunctivitis Diarrhoea Dysentery Fever Gum Bleeding Liver Issues Smallpox Stomatitis Syphilitic Ulcers Tuberculosis Typhoid Fever Wounds | Boils Diarrhoea Dysentery Gingivitis Ulcers | Analgesic Anti-Allergic Anti-Inflammatory Cephalalgia Dysentery Fever Gout Rheumatism Ulcers Wounds |
| Fixed Costs (ha−1) | Variable Costs (ha−1) | Total Cost (ha−1) | |
|---|---|---|---|
| No Control | AUD$0 | AUD$0 | AUD$0 |
| Burning | AUD$15 | AUD$0 | AUD$15 |
| Chemical | AUD$37.50 | AUD$112.50 | AUD$150 |
| Mechanical * | AUD$50 | AUD$50 | AUD$100 |
| TRADE NAME | ACTIVE INGREDIENT(S) | APPLICATION RATE | ||
|---|---|---|---|---|
| BASAL BARK & CUT-STUMP APPLICATION | Access® | Triclopyr (240 g/L) + Picloram (120 g/L) | 1 L/60 L Diesel | GROUP 4 (Disruptors of Plant Cell Growth) |
| Invader® 600 | Triclopyr (600 g/L) | 1 L/60 L Diesel | ||
| Garlon® 600 | ||||
| Redeem® 600 | ||||
| Acclaim® | Fluroxypyr (200 g/L) | 3 L/100 L Diesel | ||
| Flagship® 200 | ||||
| Starane® Advanced | Fluroxypyr (333 g/L) | 1.8 L/100 L Diesel | ||
| Comet® 400 | Fluroxypyr (400 g/L) | 1.5 L/100 L Diesel | ||
| Decoy® 400 | ||||
| HIGH VOLUME SPRAY | Fightback® | Triclopyr (300 g/L) + Picloram (100 g/L) | 350 mL/100 L Water | |
| Conqueror® | ||||
| Grazon® Extra | Triclopyr (300 g/L) + Picloram (100 g/L) + Aminopyralid (8 g/L) | 350 mL/ 100 L Water | ||
| SOIL APPLICATION | TordonTM Granules | Picloram (20 g/kg) | 35 to 45 g/m2 |
| JAN | FEB | MAR | APR | MAY | JUNE | JULY | AUG | SEP | OCT | NOV | DEC | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| FLOWERING |  |  |  |  |  |  | ||||||
| FRUITING |  |  |  |  |  |  |  |  | ||||
| GERMINATION |  |  | ||||||||||
| FOLIAR SPRAY |  |  |  |  |  |  |  | |||||
| BASAL BARK |  |  |  |  |  |  |  |  |  |  |  |  |
| CUT-STUMP |  |  |  |  |  |  |  |  |  |  |  |  |
| MANUAL |  |  |  |  |  |  |  |  |  |  |  |  |
 Growth & Reproductive Events;
Growth & Reproductive Events;  Most Suitable Control Option;
Most Suitable Control Option;  Least Suitable Control Option.
Least Suitable Control Option.| Prospective Agent(s) | PHYTOPHAGUS INSECTS | |
| Seed-Feeding Weevil | Aubeus himalayanus | |
| Leaf-Feeding Crambid Moth | Synclera univocalis | |
| Leaf-Feeding Gracillariid Moth | Phyllonorycter iochrysis | |
| Leaf-Galling Midge | Phyllodiplosis jujubae | |
| Stem-Galling Midge | Silvestrina jujubae | |
| PHYTOPHAGUS MITES | ||
| Shoot-Galling Mite | Aceria cernuus | |
| Shoot-Galling Mite | Larvacarus transitans | |
| FUNGAL PATHOGENS | ||
| Leaf Rust | Phakopsora zizyphi-vulgaris | |
| Powdery Mildew | Pseudoidium ziziphi | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
O’Brien, C.J.; Campbell, S.; Young, A.; Vogler, W.; Galea, V.J. Chinee Apple (Ziziphus mauritiana): A Comprehensive Review of Its Weediness, Ecological Impacts and Management Approaches. Plants 2023, 12, 3213. https://doi.org/10.3390/plants12183213
O’Brien CJ, Campbell S, Young A, Vogler W, Galea VJ. Chinee Apple (Ziziphus mauritiana): A Comprehensive Review of Its Weediness, Ecological Impacts and Management Approaches. Plants. 2023; 12(18):3213. https://doi.org/10.3390/plants12183213
Chicago/Turabian StyleO’Brien, Ciara J., Shane Campbell, Anthony Young, Wayne Vogler, and Victor J. Galea. 2023. "Chinee Apple (Ziziphus mauritiana): A Comprehensive Review of Its Weediness, Ecological Impacts and Management Approaches" Plants 12, no. 18: 3213. https://doi.org/10.3390/plants12183213
APA StyleO’Brien, C. J., Campbell, S., Young, A., Vogler, W., & Galea, V. J. (2023). Chinee Apple (Ziziphus mauritiana): A Comprehensive Review of Its Weediness, Ecological Impacts and Management Approaches. Plants, 12(18), 3213. https://doi.org/10.3390/plants12183213