
Pitaya as a New Alternative Crop for Iberian Peninsula: Biology and Edaphoclimatic Requirements


 ,
,  , , and
, , and 
Abstract
:1. Introduction
2. Current Pitaya Distribution in Iberian Peninsula
3. Taxonomy and Botany
3.1. Family and Genera
3.2. Species and Cultivars
4. Edaphoclimatic Requirements
5. Reproductive Biology
5.1. Flowering
5.2. Pollination
6. Genetics, Xenia and Metaxenia
7. Physiology
8. Nutritional Characteristics and Uses
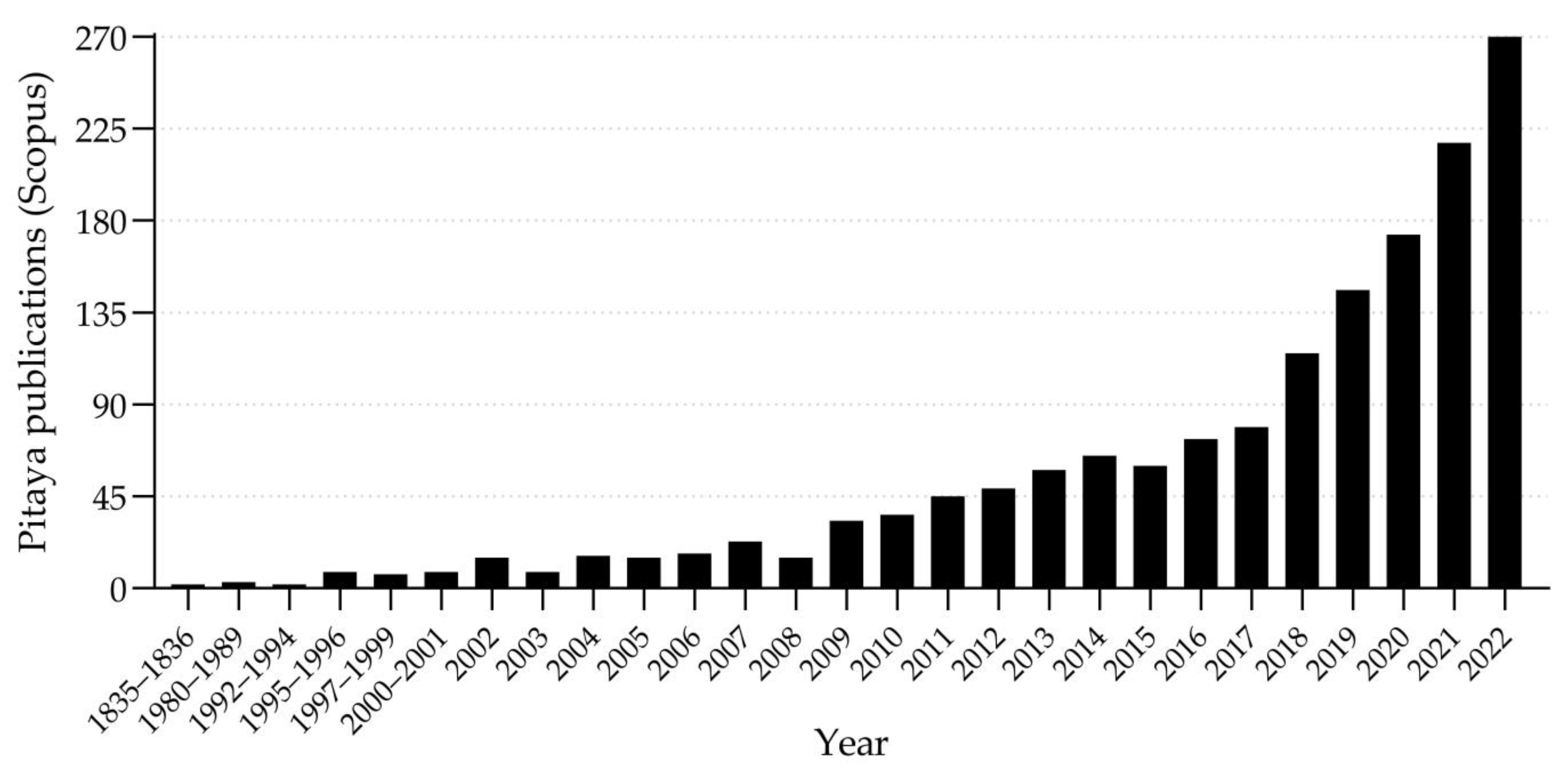
9. Previous Reviews and Published Scientific Articles
10. Future and Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Reis, P. O Olival em Portugal: DINÂMICAS, Tecnologias e Relação Com o Desenvolvimento Rural; Animar: Lisboa, Portugal, 2004. [Google Scholar]
- Viruel, J.; Le Galliot, N.; Pironon, S.; Nieto Feliner, G.; Suc, J.; Lakhal-Mirleau, F.; Juin, M.; Selva, M.; Bou Dagher Kharrat, M.; Ouahmane, L.; et al. A strong east-west Mediterranean divergence supports a new phylogeographic history of the carob tree (Ceratonia siliqua, Leguminosae) and multiple domestications from native populations. J. Biogeogr. 2020, 47, 460–471. [Google Scholar] [CrossRef]
- von Haselberg, C.D. Vegetative Growth and Flower and Fruit Development in Carob Tree (Ceratonia siliqua L.) with Special Emphasis on Environmental Conditions at Marginal Production Sites in South Portugal; Humboldt University of Berlin: Berlin, Germany, 2000. [Google Scholar]
- Duarte, A.; Fernandes, J.; Bernardes, J.; Miguel, G. Citrus as a Component of the Mediterranean Diet. J. Spat. Organ. Dyn. 2016, IV, 289–304. [Google Scholar]
- Naval, M.d.M.; Blasco, M.; Badenes, M.L. Eriobotrya japonica loquat. In Biotechnology of Fruit and Nut Crops; CAB International: Wallingford, UK, 2020; pp. 409–417. [Google Scholar]
- Ortiz-Hernandez, Y.D.; Carrillo-Salazar, J.A. Pitahaya (Hylocereus spp.): A short review. Commun. Sci. 2012, 3, 220–237. [Google Scholar]
- Fullerton, R.A.; Lee, W. Dragon Fruit—From Production to Market; Food and Fertilizer Technology Center (FFTC): Taiwan, 2021; ISBN 9789869936637. [Google Scholar]
- Dios, H.C.; Martinez, R.C.; Canché, H.J.C. Caracterización De La Producción De Pitahaya (Hylocereus spp.) en la zona maya de Quintana Roo, México. Agroecologia 2014, 9, 123–132. [Google Scholar]
- Merten, S. A review of Hylocereus production in the United States. J. Prof. Assoc. Cactus Dev. 2003, 5, 98–105. [Google Scholar]
- Mizrahi, Y.; Nerd, A.; Sitrit, Y. New Fruits for Arid Climates. In Trends in New Crops and New Uses; ASHS Press: Alexandria, VA, USA, 2002; pp. 378–384. [Google Scholar]
- Tel-Zur, N. Breeding an underutilized fruit crop: A long-term program for Hylocereus. Hortic. Res. 2022, 9, uhac078. [Google Scholar] [CrossRef]
- Trivellini, A.; Lucchesini, M.; Ferrante, A.; Massa, D.; Orlando, M.; Incrocci, L.; Mensuali-Sodi, A. Pitaya, an Attractive Alternative Crop for Mediterranean Region. Agronomy 2020, 10, 1065. [Google Scholar] [CrossRef]
- Cacioppo, O. Fico d’India e Pitaya; L’Informatore Agrario: Verona, Italy, 1991. [Google Scholar]
- Cacioppo, O. Risultati Sperimentali di Anni di Coltivazione Della Pitaya e Avocado in Provincia di Latina. In Proceedings of the Incontro Nazionale Sulla Frutticoltura Alternativa, Milazzo, Italy, 22–23 October 2016; pp. 70–79. [Google Scholar]
- Kokkini, M. Exotic Dragon Fruit Grown 30 km from the Center of Athens. Available online: https://www.lifo.gr/tropos-zois/gefsi/exotika-dragon-fruit-poy-kalliergoyntai-30hlm-apo-kentro-tis-athinas (accessed on 31 August 2023).
- Anonymous. Pitaya Fruits for Better Life. Available online: https://pitaya.gr/ (accessed on 31 August 2023).
- Becerra, J.A.; Lacerda, V.R. Cultivo da pitaia na Espanha, Importância da Pitaia Orgânica e Futuro da comercialização da Pitaia no Mundo. In I Circuito Internacional de Pitaia: Tendências e Projeções Latino-Americanas Para a Cultura da Pitaia; Lacerda, V.R., Almeida, S., Eds.; FEPAF: Botucatu, Brazil, 2022; Volume 1, pp. 33–35. [Google Scholar]
- Duarte, A.; Trindade, A.R. Situação atual do cultivo da pitaia em Portugal. In I Circuito Internacional de Pitaia: Tendências e Projeções Latino-Americanas Para a Cultura da Pitaia; Lacerda, V.R., Almeida, S., Eds.; FEPAF: Botucatu, Brazil, 2022; Volume 1, pp. 87–99. [Google Scholar]
- Trindade, A.; Reis, A.; Sabbo, L.; Trindade, D.; Paiva, P.; Duarte, A. Pitaia: Perspetivas e Dificuldades de Uma “Nova” Cultura. Agrotec 2019, 30, 32–34. [Google Scholar]
- Korotkova, N.; Aquino, D.; Arias, S.; Eggli, U.; Franck, A.; Gómez-Hinostrosa, C.; Guerrero, P.C.; Hernández, H.M.; Kohlbecker, A.; Köhler, M.; et al. Cactaceae at Caryophyllales.org—A dynamic online species-level taxonomic backbone for the family. Willdenowia 2021, 51, 251–270. [Google Scholar] [CrossRef]
- Rojas-Sandoval, J.; Praciak, A. Hylocereus undatus (dragon fruit). CABI Compend. 2022. [Google Scholar] [CrossRef]
- Mizrahi, Y. Do we need new crops for arid regions? A review of fruit species domestication in Israel. Agronomy 2020, 10, 1995. [Google Scholar] [CrossRef]
- García-Cruz, L.; Valle-Guadarrama, S.; Guerra-Ramírez, D.; Martínez-Damián, M.T.; Zuleta-Prada, H. Cultivation, quality attributes, postharvest behavior, bioactive compounds, and uses of Stenocereus: A review. Sci. Hortic. 2022, 304, 111336. [Google Scholar] [CrossRef]
- Sitrit, Y.; Golan, E.; Bar, E.; Lewinsohn, E. Fruit quality evaluation of two new cactus crops for arid zones: Cereus peruvianus and Cereus jamacaru. Isr. J. Plant Sci. 2012, 60, 335–343. [Google Scholar]
- Bravo-Hollis, H.; Sánchez-Mejorada, H. Las cactáceas de México; UNAM: Mexico City, México, 1991; Volume 3. [Google Scholar]
- Quiroz-González, B.; García-Mateos, R.; Corrales-García, J.J.E.; Colinas-León, M.T. Pitaya (Stenocereus spp.): An under-utilized fruit. J. Prof. Assoc. Cactus Dev. 2018, 20, 82–100. [Google Scholar] [CrossRef]
- Hunt, D. Notes on Selenicereus (A. Berger) Britton & Rose and Aporocactus Lemaire (Cactaceae-Hylocereinae). Bradleya 1989, 7, 89–96. [Google Scholar] [CrossRef]
- Hunt, D.; Taylor, N. Notes on miscellaneous genera of Cactaceae. Bradleya 1991, 9, 81–92. [Google Scholar] [CrossRef]
- Casas, A.; Caballero, J.; Valiente-Banuet, A.; Soriano, J.A.; Dávila, P. Morphological variation and the process of domestication of Stenocereus stellatus (Cactaceae) in Central Mexico. Am. J. Bot. 1999, 86, 522–533. [Google Scholar] [CrossRef]
- Grimaldo-Juárez, O.; Terrazas, T.; García-Velásquez, A.; Cruz-Villagas, M.; Ponce-Medina, J.F. Morphometric Analysis of 21 Pitahaya (Hylocereus undatus) Genotypes. J. Prof. Assoc. Cactus Dev. 2007, 9, 99–117. [Google Scholar]
- Nascimento, D.D.; Lopes, A.P.M.; Ferreira, R.J.; Carvalho, V.R.; Bombonato, L.T.; Diniz, D.B.; Silva, J.A.A.; Wilcker, S.R.S.; Soares, P.L.M. First Report of Meloidogyne javanica Infecting Hylocereus megalanthus in Brazil. Plant Dis. 2020, 104, 2526. [Google Scholar] [CrossRef]
- Guzman-Piedrabita, O.A.; Perez, L.; Patiño, A. Reconocimiento de nematodos fitoparásitos pitahaya amarilla (Selenicereus megalanthus HAW.). Boletín Científico Mus. Hist. Nat. 2012, 16, 149–161. [Google Scholar]
- Madadi, E.; Mazloum-Ravasan, S.; Yu, J.S.; Ha, J.W.; Hamishehkar, H.; Kim, K.H. Therapeutic application of betalains: A review. Plants 2020, 9, 1219. [Google Scholar] [CrossRef] [PubMed]
- Mori, C.V.; Patel, A.R.; Parmar, V.K.; Patel, G.S. Dragon fruit (Kamalam): An excellent exotic fruit crop of India. Pharma Innov. J. 2023, 12, 115–123. [Google Scholar] [CrossRef]
- Santos, D.N.; Pio, L.A.S.; Faleiro, F.G. Pitaya: Uma Alternativa Frutífera, 1st ed.; EMBRAPA: Brasília, Brazil, 2022; Volume 1, ISBN 978-65-991179-1-6. [Google Scholar]
- Faleiro, F.G.; Junqueira, N.T.V. Melhoramento genético das pitaias na Embrapa e parceiros. In III Encontro Nacional dos Produtores de Pitaia; Lacerda, V.R., Ed.; FEPAF: Botucatu, Brazil, 2022; Volume 3, pp. 1–184. ISBN 9786589571100. [Google Scholar]
- Gasic, K.; Preece, J.E.; Karp, D. (Eds.) Register of New Fruit and Nut Cultivars List 48. HortScience 2016, 51, 620–652. [Google Scholar] [CrossRef]
- Karp, D.; Gasic, K. Register of New Fruit and Nut Cultivars List 51. HortScience 2022, 57, 1174–1233. [Google Scholar] [CrossRef]
- Ben-Asher, J.; Nobel, P.S.; Yossov, E.; Mizrahi, Y. Net CO2 uptake rates for Hylocereus undatus and Selenicereus megalanthus under field conditions: Drought influence and a novel method for analyzing temperature dependence. Photosynthetica 2006, 44, 181–186. [Google Scholar] [CrossRef]
- Barbeau, G. La pitahaya rouge, un nouveau fruit exotique. Fruits Paris. 1990, 45, 141–147. [Google Scholar]
- Thomson, P. Pitahaya: A Promosing New Crop for Southern California, 2nd ed.; Bonsall Publications: Bonsall, CA, USA, 2002. [Google Scholar]
- Nerd, A.; Neumann, P.M. Phloem Water Transport Maintains Stem Growth in a Drought-stressed Crop Cactus (Hylocereus undatus). J. Am. Soc. Hortic. Sci. 2004, 129, 486–490. [Google Scholar] [CrossRef]
- Yuqing, H.; Weiyuan, Y.; Ling, M.; Guangping, X.; Zhongfeng, Z.; Danjuan, Z.; Chengxin, H.; Daxing, G. Physiological Effect on Hylocereus undulatus and Hylocereus undatus Under Simulated Karst Soil Water Deficiency. J. Resour. Ecol. 2015, 6, 269–275. [Google Scholar] [CrossRef]
- Lima, C.A. Caracterização, Propagação e Melhoramento Genético de Pitaya Comercial e Nativa do Cerrado. Ph.D. Thesis, Universidade de Brasillia, Brasíllia, Brazil, 2013. [Google Scholar]
- Qu, Y.; Nong, Q.; Jian, S.; Lu, H.; Zhang, M.; Xia, K. An AP2/ERF Gene, HuERF1, from Pitaya (Hylocereus undatus) Positively Regulates Salt Tolerance. Int. J. Mol. Sci. 2020, 21, 4586. [Google Scholar] [CrossRef]
- Sousa, G.G.d.; Sousa, S.B.; Pereira, A.C.d.S.; Marques, V.B.; Silva, M.L.G.d.; Lopes, J.d.S. Effect of saline water and shading on dragon fruit (‘pitaya’) seedling growth. Rev. Bras. Eng. Agrícola e Ambient. 2021, 25, 547–552. [Google Scholar] [CrossRef]
- Gunasena, H.P.M.; Pushpakumara, D.K.N.G.; Kariayawasam, M. Dragon fruit Hylocereus undatus (Haw.) Britton and Rose. In Underutilized Fruit Trees in Sri Lanka; World Agroforestry (ICRAF): New Delhi, India, 2007; pp. 110–141. [Google Scholar]
- Raveh, E.; Nerd, A.; Mizrahi, Y. Responses of two hemiepiphytic fruit crop cacti to different degrees of shade. Sci. Hortic. 1998, 73, 151–164. [Google Scholar] [CrossRef]
- Almeida, E.I.B.; De Medeiros Corrêa, M.C.; Mesquita, R.O.; Queiroz, R.F.; Cajazeira, J.P.; Amorim, F.F.V.R. Growth and gas exchanges of red pitaya under different shading conditions. Rev. Bras. Cienc. Agrar. 2018, 13, 1–8. [Google Scholar] [CrossRef]
- de Oliveira, M.M.T.; Albano-Machado, F.G.; Penha, D.M.; Pinho, M.M.; Natale, W.; de Miranda, M.R.A.; Moura, C.F.H.; Alves, R.E.; de Medeiros Corrêa, M.C. Shade improves growth, photosynthetic performance, production and postharvest quality in red pitahaya (Hylocereus costaricensis). Sci. Hortic. 2021, 286, 110217. [Google Scholar] [CrossRef]
- Trindade, A.R.; Trindade, D.; Marques, N.; Neto, L.; Duarte, A. Manual Técnico: A Cultura da Pitaia; Grupo Operacional Fruta Dragão: Validar a Capacidade Produtiva da Pitaia Vermelha: Lisbon, Portugal, 2022; ISBN 978-989-8319-55-5. [Google Scholar]
- Nerd, A.; Mizrahi, Y. Reproductive biology of cactus fruit crops. Hortic. Rev. (Am. Soc. Hortic. Sci.) 1997, 18, 321–346. [Google Scholar]
- Trindade, A.R. Floração e Frutificação da Pitaia (Hylocereus undatus). Master’s Thesis, Universidade do Algarve, Faro, Portugal, 2022. [Google Scholar]
- Weiss, J.; Nerd, A.; Mizrahi, Y. Flowering behavior and pollination requirements in climbing cacti with fruit crop potential. HortScience 1994, 29, 1487–1492. [Google Scholar] [CrossRef]
- Cruz, M.C.M.; Martins, R.S. Pitaia No Brasil, Nova Opção de Cultivo, 1st ed.; Epagri: Florianópolis, Brazil, 2022; Volume 1, ISBN 978-65-990745-9-2. [Google Scholar]
- Muniz, J.P.d.O.; Bomfim, I.G.A.; Corrêa, M.C.d.M.; Freitas, B.M. Floral biology, pollination requirements and behavior of floral visitors in two species of pitaya. Rev. Cienc. Agron. 2019, 50, 640–649. [Google Scholar] [CrossRef]
- Silva, D.; Villa, F.; Silva, L. Biologia Floral de Pitaia. In III Encontro Nacional dos Produtores de Pitaia; Lacerda, V., Ed.; FEPAF: Botucatu, Brazil, 2022; Volume 1, pp. 10–21. [Google Scholar]
- Freitas, B.M.; Muniz, J.P.d.O.; Bomfim, I.G.A. Polinização complementar por abelhas para maximizar a produção e qualidade de frutos da pitaia. In III Encontro Nacional dos Produtores de Pitaia; Lacerda, V.R., Ed.; FEPAF: Botucatu, Brazil, 2022; Volume 3, pp. 22–41. ISBN 9786589571100. [Google Scholar]
- Taiz, L.; Zeiger, E.; Moller, I.M.; Murphy, A. Fisiologia e Desenvolvimento Vegetal, 6th ed.; Artmed Editora: Porto Alegre, Brazil, 2017; Volume 6, ISBN 978-85-8271-366-2. [Google Scholar]
- Magalhães, D.S.; Ramos, J.D.; Pio, L.A.S.; Vilas Boas, E.V.d.B.; Pasqual, M.; Rodrigues, F.A.; Rufini, J.C.M.; Santos, V.A. Dos Physical and physicochemical modifications of white-fleshed pitaya throughout its development. Sci. Hortic. 2019, 243, 537–543. [Google Scholar] [CrossRef]
- Jiang, Y.; Liao, Y.; Lin, T.; Lee, C. The Photoperiod-regulated Bud Formation of Red Pitaya (Hylocereus sp.). HortScience 2012, 47, 1063–1067. [Google Scholar] [CrossRef]
- Mizrahi, Y. Thirty-one Years of Research and Development in the Vine Cacti Pitaya in Israel. Improv. Pitaya Prod. Mark. 2015, 1–18. [Google Scholar]
- Silva, A.; Martins, A.; Cavallari, L. Qualidade de frutos de pitaya em função da época de polinização, da fonte de pólen e da coloração da cobertura. Rev. Bras. Frutic. 2012, 33, 1162–1168. [Google Scholar] [CrossRef]
- Jacobs, D. Pitaya, Hylocereus Undatus (Haw)—A Potential New Crop for Australia; Wageningen Agricultural University: Wageningen, The Netherlands, 1998. [Google Scholar]
- Rainho, A.; Alves, P.; Amorim, F.; Marques, J.T. Atlas dos Morcegos de Portugal Continental; ICNF (Portugal): Lisbon, Portugal, 2013; ISBN 978-972-775-226-3. [Google Scholar]
- Rainho, A. Summer foraging habitats of bats in a Mediterranean region of the Iberian Peninsula. Acta Chiropterol. 2007, 9, 171–181. [Google Scholar] [CrossRef]
- Horácek, I. Remarks on the causality of population decline in european bats. Myotis 1984, 21–22, 138–147. [Google Scholar]
- Bellec, F.L.; Vallant, F.; Imbert, E. Pitahaya (Hylocereus spp.): A new fruit crop, a market with a future. Fruits 2006, 61, 3–7. [Google Scholar] [CrossRef]
- Lichtenzveig, J.; Abbo, S.; Nerd, A.; Tel-Zur, N.; Mizrahi, Y. Cytology and mating systems in the climbing cacti Hylocereus and Selenicereus. Am. J. Bot. 2000, 87, 1058–1065. [Google Scholar] [CrossRef]
- Tel-Zur, N.; Mizrahi, Y.; Cisneros, A.; Mouyal, J.; Schneider, B.; Doyle, J.J. Phenotypic and genomic characterization of vine cactus collection (Cactaceae). Genet. Resour. Crop Evol. 2011, 58, 1075–1085. [Google Scholar] [CrossRef]
- Dani, D.; Rokhmah, D.N. A review of the role of pollination on the yield of cocoa plant. Kultivasi 2022, 21, 249–255. [Google Scholar] [CrossRef]
- Denney, J.O. Xenia Includes Metaxenia. HortScience 1992, 27, 722–728. [Google Scholar] [CrossRef]
- Deng, L.; Wang, T.; Hu, J.; Yang, X.; Yao, Y.; Jin, Z.; Huang, Z.; Sun, G.; Xiong, B.; Liao, L.; et al. Effects of Pollen Sources on Fruit Set and Fruit Characteristics of ‘Fengtangli’ Plum (Prunus salicina Lindl.) Based on Microscopic and Transcriptomic Analysis. Int. J. Mol. Sci. 2022, 23, 12959. [Google Scholar] [CrossRef]
- Shahsavar, A.R.; Shahhosseini, A. The metaxenia effects of different pollen grains on secondary metabolites enzymes and sugars of ‘Piarom’ date palm fruit. Sci. Rep. 2022, 12, 10058. [Google Scholar] [CrossRef]
- Zhang, H.; Liu, C.; Yao, J.-L.; Deng, C.H.; Chen, S.; Chen, J.; Wang, Z.; Yu, Q.; Cheng, Y.; Xu, J. Citrus mangshanensis Pollen Confers a Xenia Effect on Linalool Oxide Accumulation in Pummelo Fruit by Enhancing the Expression of a Cytochrome P450 78A7 Gene CitLO 1. J. Agric. Food Chem. 2019, 67, 9468–9476. [Google Scholar] [CrossRef]
- Alinezhad Jahromi, H.; Zarei, A.; Mohammadkhani, A. Analysis the effects of pollen grain sources on the fruits set and their characteristics of ‘Clementine’ mandarin using microscopic and molecular approaches. Sci. Hortic. 2019, 249, 347–354. [Google Scholar] [CrossRef]
- Zhang, M.; Wang, Z.; Mao, Y.; Hu, Y.; Yang, L.; Wang, Y.; Zhang, L.; Shen, X. Effects of quince pollen pollination on fruit qualities and phenolic substance contents of apples. Sci. Hortic. 2019, 256, 108628. [Google Scholar] [CrossRef]
- Doi, K.; Inoue, R.; Iwasaki, N. Seed weight mediates effects of pollen on berry weight, ripening, and anthocyanin content in highbush blueberry. Sci. Hortic. 2021, 288, 110313. [Google Scholar] [CrossRef]
- Pagliaccia, D.; Vidalakis, G.; Douhan, G.W.; Lobo, R.; Tanizaki, G. Genetic characterization of pitahaya accessions based on amplified fragment length polymorphism analysis. HortScience 2015, 50, 332–336. [Google Scholar] [CrossRef]
- da Silva, A.D.C.C.; Sabião, R.R.; Chiamolera, F.M.; Segantini, D.M.; Martins, A.B.G. Morphological Traits as Tool to Verify Genetic Variability of Interspecific Dragon Fruit Hybrids. Rev. Bras. Frutic. 2017, 39, e-168. [Google Scholar] [CrossRef]
- Wu, Q.; Gao, H.; You, Z.; Zhang, Z.; Zhu, H.; He, M.; He, J.; Duan, X.; Jiang, Y.; Yun, Z. Multiple Metabolomics Comparatively Investigated the Pulp Breakdown of Four Dragon Fruit Cultivars during Postharvest Storage. Food Res. Int. 2023, 164, 112410. [Google Scholar] [CrossRef]
- Lima, C.A.d.; Faleiro, F.G.; Junqueira, N.T.V. Diversidade genética intra e interespecífica de pitaya com base nas características físico-químicas de frutos. Rev. Bras. Frutic. 2014, 35, 1066–1072. [Google Scholar] [CrossRef]
- Mizrahi, Y.; Mouyal, J.; Nerd, A.; Sitrit, Y. Metaxenia in the vine cacti Hylocereus polyrhizus and Selenicereus spp. Ann. Bot. 2004, 93, 469–472. [Google Scholar] [CrossRef]
- Chen, C.; Li, F.; Xie, F.; Chen, J.; Hua, Q.; Chen, J.; Wu, Z.; Zhang, Z.; Zhang, R.; Zhao, J.; et al. Pitaya Genome and Multiomics Database (PGMD): A Comprehensive and Integrative Resource of Selenicereus undatus. Genes 2022, 13, 745. [Google Scholar] [CrossRef]
- Nobel, P.S.; De La Barrera, E. CO2 uptake by the cultivated hemiepiphytic cactus, Hylocereus undatus. Ann. Appl. Biol. 2004, 144, 1–8. [Google Scholar] [CrossRef]
- Nobel, P.S. A Nutrient Index Quantifying Productivity of Agaves and Cacti. J. Appl. Ecol. 1989, 26, 635. [Google Scholar] [CrossRef]
- Nobel, P.S. Parenchyma-chlorenchyma water movement during drought for the hemiepiphytic cactus Hylocereus undatus. Ann. Bot. 2006, 97, 469–474. [Google Scholar] [CrossRef] [PubMed]
- Weiss, I.; Raveh, E.; Mizrahi, Y. Effects of CO2—Enrichment and fertilization regimes on net CO2 uptake and growth of Hylocereus undatus. J. Am. Soc. Hortic. Sci. 2009, 134, 364–371. [Google Scholar] [CrossRef]
- Weiss, I.; Mizrahi, Y.; Raveh, E. Effect of elevated CO2 on vegetative and reprodutive growth characteristics of the CAM plants Hylocereus undatus and Selenicereus megalanthus. Sci. Hortic. 2010, 123, 531–536. [Google Scholar] [CrossRef]
- Munekata, P.E.S.; Pateiro, M.; Domínguez, R.; Nieto, G.; Kumar, M.; Dhama, K.; Lorenzo, J.M. Bioactive Compounds from Fruits as Preservatives. Foods 2023, 12, 343. [Google Scholar] [CrossRef] [PubMed]
- Azeredo, H.M.C. Betalains: Properties, sources, applications, and stability—A review. Int. J. Food Sci. Technol. 2009, 44, 2365–2376. [Google Scholar] [CrossRef]
- Ismail, O.M.; Abdel-Aziz, M.S.; Ghareeb, M.A.; Hassan, R.Y.A. Exploring the biological activities of the Hylocereus polyrhizus extract. J. Innov. Pharm. Biol. Sci. 2017, 4, 1–6. [Google Scholar]
- Song, H.; Zheng, Z.; Wu, J.; Lai, J.; Chu, Q.; Zheng, X. White pitaya (Hylocereus undatus) juice attenuates insulin resistance and hepatic steatosis in diet-induced obese mice. PLoS ONE 2016, 11, e0149670. [Google Scholar] [CrossRef]
- Nishikito, D.F.; Borges, A.C.A.; Laurindo, L.F.; Otoboni, A.M.M.B.; Direito, R.; Goulart, R.d.A.; Nicolau, C.C.T.; Fiorini, A.M.R.; Sinatora, R.V.; Barbalho, S.M. Anti-Inflammatory, Antioxidant, and Other Health Effects of Dragon Fruit and Potential Delivery Systems for Its Bioactive Compounds. Pharmaceutics 2023, 15, 159. [Google Scholar] [CrossRef]
- Poolsup, N.; Suksomboon, N.; Paw, N.J. Effect of dragon fruit on glycemic control in prediabetes and type 2 diabetes: A systematic review and meta-analysis. PLoS ONE 2017, 12, e0184577. [Google Scholar] [CrossRef] [PubMed]
- Cheah, L.; Eid, A.; Aziz, A.; Ariffin, F.; Elmahjoubi, A.; Elmarzugi, N. Phytochemical Properties and Health Benefits of Hylocereus undatus. Nanomed. Nanotechnol. 2016, 1, 1–10. [Google Scholar]
- Adnan, L.; Osman, A.; Abdul Hamid, A. Antioxidant activity of different extracts of red pitaya (Hylocereus polyrhizus) seed. Int. J. Food Prop. 2011, 14, 1171–1181. [Google Scholar] [CrossRef]
- Liaotrakoon, W.; De Clercq, N.; Van Hoed, V.; Dewettinck, K. Dragon Fruit (Hylocereus spp.) Seed Oils: Their Characterization and Stability Under Storage Conditions. J. Am. Oil Chem. Soc. 2013, 90, 207–215. [Google Scholar] [CrossRef]
- Liaotrakoon, W.; de Clercq, N.; van Hoed, V.; van de Walle, D.; Lewille, B.; Dewettinck, K. Impact of Thermal Treatment on Physicochemical, Antioxidative and Rheological Properties of White-Flesh and Red-Flesh Dragon Fruit (Hylocereus spp.) Purees. Food Bioprocess Technol. 2013, 6, 416–430. [Google Scholar] [CrossRef]
- Michelle, C.J.; Joice, V.C.O.; Maria, R.C.G.N. Nutritional pharmacological and toxicological characteristics of pitaya (Hylocereus undatus): A review of the literature. Afr. J. Pharm. Pharmacol. 2017, 11, 300–304. [Google Scholar] [CrossRef]
- Shahid, M.; Shahid-Ul-Islam; Mohammad, F. Recent advancements in natural dye applications: A review. J. Clean. Prod. 2013, 53, 310–331. [Google Scholar] [CrossRef]
- Mello, F.R.d.; Bernardo, C.; Dias, C.O.; Gonzaga, L.; Amante, E.R.; Fett, R.; Candido, L.M.B. Antioxidant properties, quantification and stability of betalains from pitaya (Hylocereus undatus) peel. Cienc. Rural. 2014, 45, 323–328. [Google Scholar] [CrossRef]
- Nurmahani, M.M.; Osman, A.; Hamid, A.; Ghazali, M.; Dek, P. Antibacterial activity of Hylocereus polyrhizus and Hylocereus undatus peel extract. Int. Food Res. J. 2012, 19, 77–84. [Google Scholar]
- Sousa, E.M.P. Extração, Análise de Estabilidade, Higroscopicidade e Reologia do Corante de Pitaya (Hylocereus costaricencis). Ph.D. Thesis, Universidade Federal do Pará, Belém, Pará, Brazil, 2015. [Google Scholar]
- Roriz, C.L.; Heleno, S.A.; Alves, M.J.; Oliveira, M.B.P.P.; Pinela, J.; Dias, M.I.; Calhelha, R.C.; Morales, P.; Ferreira, I.C.F.R.; Barros, L. Red Pitaya (Hylocereus costaricensis) Peel as a Source of Valuable Molecules: Extraction Optimization to Recover Natural Colouring Agents. Food Chem. 2022, 372, 131344. [Google Scholar] [CrossRef]
- Esquivel, P.; Araya Quesada, Y. Características del fruto de la pitahaya (Hylocereus sp.) y su potencial de uso en la industria alimentaria. Rev. Venez. Cienc. Y Tecnol. Aliment. 2012, 3, 113–129. [Google Scholar]
- Herbach, K.M.; Stintzing, F.C.; Carle, R. Betalain Stability and Degradation? Structural and Chromatic Aspects. J. Food Sci. 2006, 71, R41–R50. [Google Scholar] [CrossRef]
- Herbach, K.M.; Maier, C.; Stintzing, F.C.; Carle, R. Effects of processing and storage on juice colour and betacyanin stability of purple pitaya (Hylocereus polyrhizus) juice. Eur. Food Res. Technol. 2007, 224, 649–658. [Google Scholar] [CrossRef]
- Mizrahi, Y. Vine-cacti pitayas: The new crops of the world. Rev. Bras. Frutic. 2014, 36, 124–138. [Google Scholar] [CrossRef]
- Pollnow, G.E. Pitaia, da propagação à colheita: Uma revisão. Agropecuária Catarin. 2018, 31, 73–78. [Google Scholar]
- Nunes, E.N.; Sousa, A.S.B.d.; Lucena, C.M.d.; Silva, S.d.M.; Lucena, R.F.P.d.; Alves, C.A.B.; Alves, R.E. Pitaia (Hylocereus sp.): Uma revisão para o Brasil. Gaia Sci. 2014, 8, 90–98. [Google Scholar]
- Perween, T.; Mandal, K.; Hasan, M. Dragon fruit: An exotic super future fruit of India. J. Pharmacogn. Phytochem. 2018, 7, 1022–1026. [Google Scholar]
- Le, N.L. Functional compounds in dragon fruit peels and their potential health benefits: A review. Int. J. Food Sci. Technol. 2022, 57, 2571–2580. [Google Scholar] [CrossRef]
- Mercado-Silva, E.M. Pitaya—Hylocereus Undatus (Haw); Elsevier Inc.: Amsterdam, The Netherlands, 2018; ISBN 9780128031384. [Google Scholar]
- Jiang, H.; Zhang, W.; Li, X.; Shu, C.; Jiang, W.; Cao, J. Nutrition, phytochemical profile, bioactivities and applications in food industry of pitaya (Hylocereus spp.) peels: A comprehensive review. Trends Food Sci. Technol. 2021, 116, 199–217. [Google Scholar] [CrossRef]
- Huang, Y.; Brennan, M.A.; Kasapis, S.; Richardson, S.J.; Brennan, C.S. Maturation Process, Nutritional Profile, Bioactivities and Utilisation in Food Products of Red Pitaya Fruits: A Review. Foods 2021, 10, 2862. [Google Scholar] [CrossRef]
- Jalgaonkar, K.; Mahawar, M.K.; Bibwe, B.; Kannaujia, P. Postharvest Profile, Processing and Waste Utilization of Dragon Fruit (Hylocereus Spp.): A Review. Food Rev. Int. 2022, 38, 733–759. [Google Scholar] [CrossRef]
- Harlev, E.; Nevo, E.; Solowey, E.; Bishayee, A. Cancer preventive and curative attributes of plants of the Cactaceae family: A review. Planta Med. 2013, 79, 713–722. [Google Scholar] [CrossRef]
- Ibrahim, S.R.M.; Mohamed, G.A.; Khedr, A.I.M.; Zayed, M.F.; El-Kholy, A.A.-E.S. Genus Hylocereus: Beneficial phytochemicals, nutritional importance, and biological relevance—A review. J. Food Biochem. 2018, 42, e12491. [Google Scholar] [CrossRef]
- Hossain, F.; Numan, S.M.; Akhtar, S. Cultivation, Nutritional Value and Health Benefits of Dragon Fruit (Hylocereus Spp.): A Review. Int. J. Hortic. Sci. Technol. 2021, 9, 259–269. [Google Scholar] [CrossRef]
- Li, X.; Liu, Y.; Zheng, Y.; Wang, P.; Zhang, Y. The Effect of Vitamin D Supplementation on Glycemic Control in Type 2 Diabetes Patients: A Systematic Review and Meta-Analysis. Nutrients 2018, 10, 375. [Google Scholar] [CrossRef]
- Lopes, M.R.d.S.; Fonseca, N.M.C.d.J.; Nascimento, V.A.d.; Guimarães, R.d.C.A. Study of the species and technological applica ons of pitaya: A review. Multitemas 2022, 67, 167–187. [Google Scholar] [CrossRef]
- Ramírez-Rodríguez, Y.; Martínez-Tagüeña, N.; Trujillo, J.; Martínez-Huélamo, M.; Pedraza-Chaverri, J.; Ramírez, V. Ethnobotanical, nutritional and medicinal properties of Mexican drylands Cactaceae Fruits: Recent findings and research opportunities. Food Chem. 2020, 312, 126073. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species and Cultivars | Peel Color | Pulp Color | Fruit Size (g) | TSS * (°Brix) | Fertility ** |
|---|---|---|---|---|---|
| S. undatus | |||||
| ‘Common White’ | Red | White | 500–1000 | 15 | SI |
| ‘Vietnamese White’ | Red | White | 500–1000 | 15 | SF |
| ‘Golden’ | Yellow | White | 200–800 | 15 | PSC |
| S. monacanthus | |||||
| ‘Tesoro’ | Red | Red | 400–800 | 18 | SC |
| S. costaricensis | |||||
| ‘Costa Rica’ | Red | Red | 400–800 | 18 | SC |
| S. megalanthus | |||||
| ‘Palora’ | Yellow | White | 200–500 | 22 | SC |
| ‘Colombian yellow’ | Yellow | White | 200–500 | 22 | SC |
| Topic | Amount | References |
|---|---|---|
| Agricultural sciences | 6 | [6,68,109,110,111,112] |
| Food chemistry and health | 16 | [68,90,91,94,100,113,114,115,116,117,118,119,120,121,122,123] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Trindade, A.R.; Paiva, P.; Lacerda, V.; Marques, N.; Neto, L.; Duarte, A. Pitaya as a New Alternative Crop for Iberian Peninsula: Biology and Edaphoclimatic Requirements. Plants 2023, 12, 3212. https://doi.org/10.3390/plants12183212
Trindade AR, Paiva P, Lacerda V, Marques N, Neto L, Duarte A. Pitaya as a New Alternative Crop for Iberian Peninsula: Biology and Edaphoclimatic Requirements. Plants. 2023; 12(18):3212. https://doi.org/10.3390/plants12183212
Chicago/Turabian StyleTrindade, Ana Rita, Paulo Paiva, Vander Lacerda, Natália Marques, Luís Neto, and Amílcar Duarte. 2023. "Pitaya as a New Alternative Crop for Iberian Peninsula: Biology and Edaphoclimatic Requirements" Plants 12, no. 18: 3212. https://doi.org/10.3390/plants12183212
APA StyleTrindade, A. R., Paiva, P., Lacerda, V., Marques, N., Neto, L., & Duarte, A. (2023). Pitaya as a New Alternative Crop for Iberian Peninsula: Biology and Edaphoclimatic Requirements. Plants, 12(18), 3212. https://doi.org/10.3390/plants12183212