
Chemical Composition of Essential Oils from Eight Tunisian Eucalyptus Species and Their Antifungal and Herbicidal Activities
 , , , , , ,
, , , , , ,  ,
,  , and
, and
Abstract
:1. Introduction
2. Results
2.1. Yields and Chemical Composition
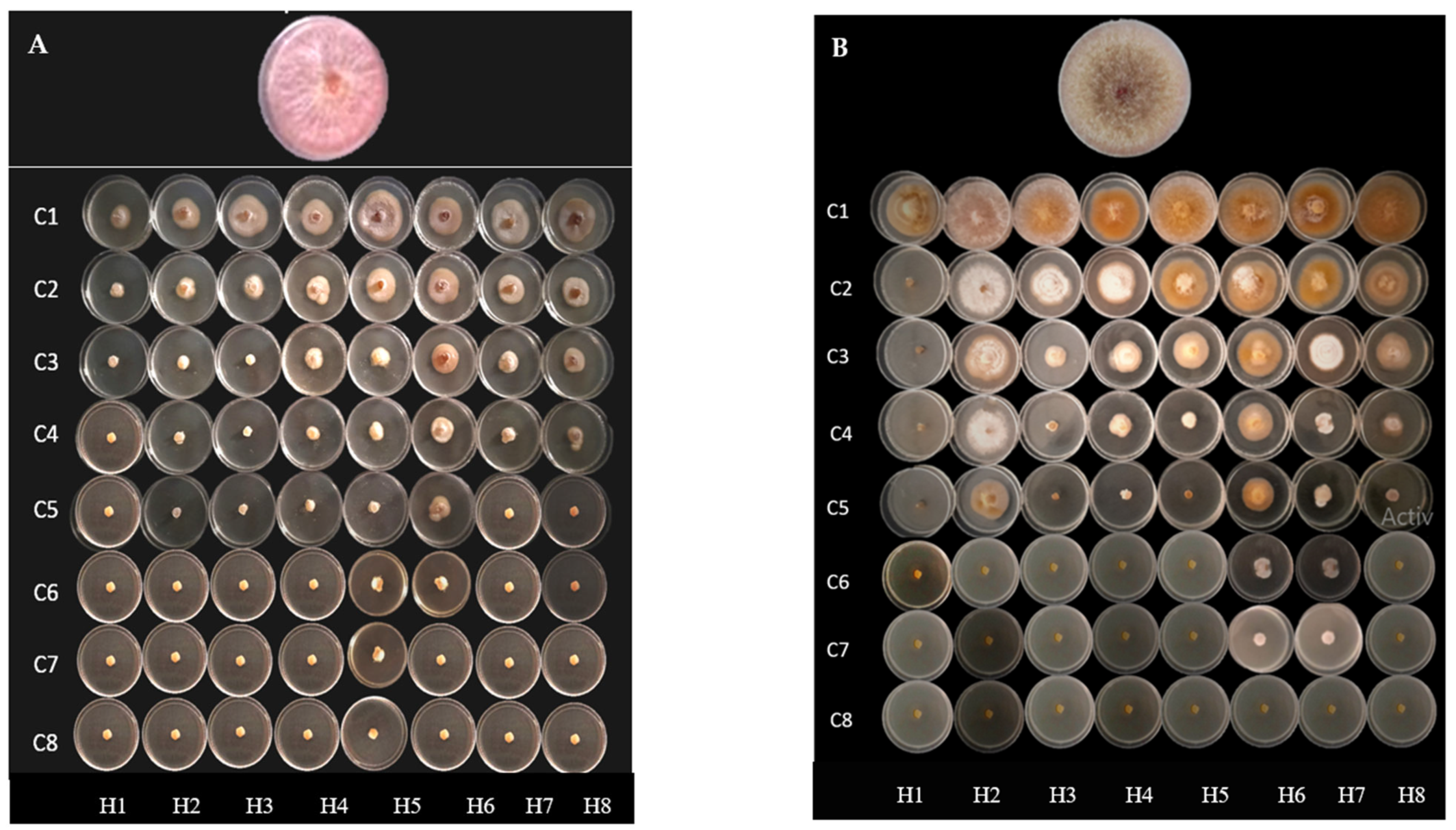
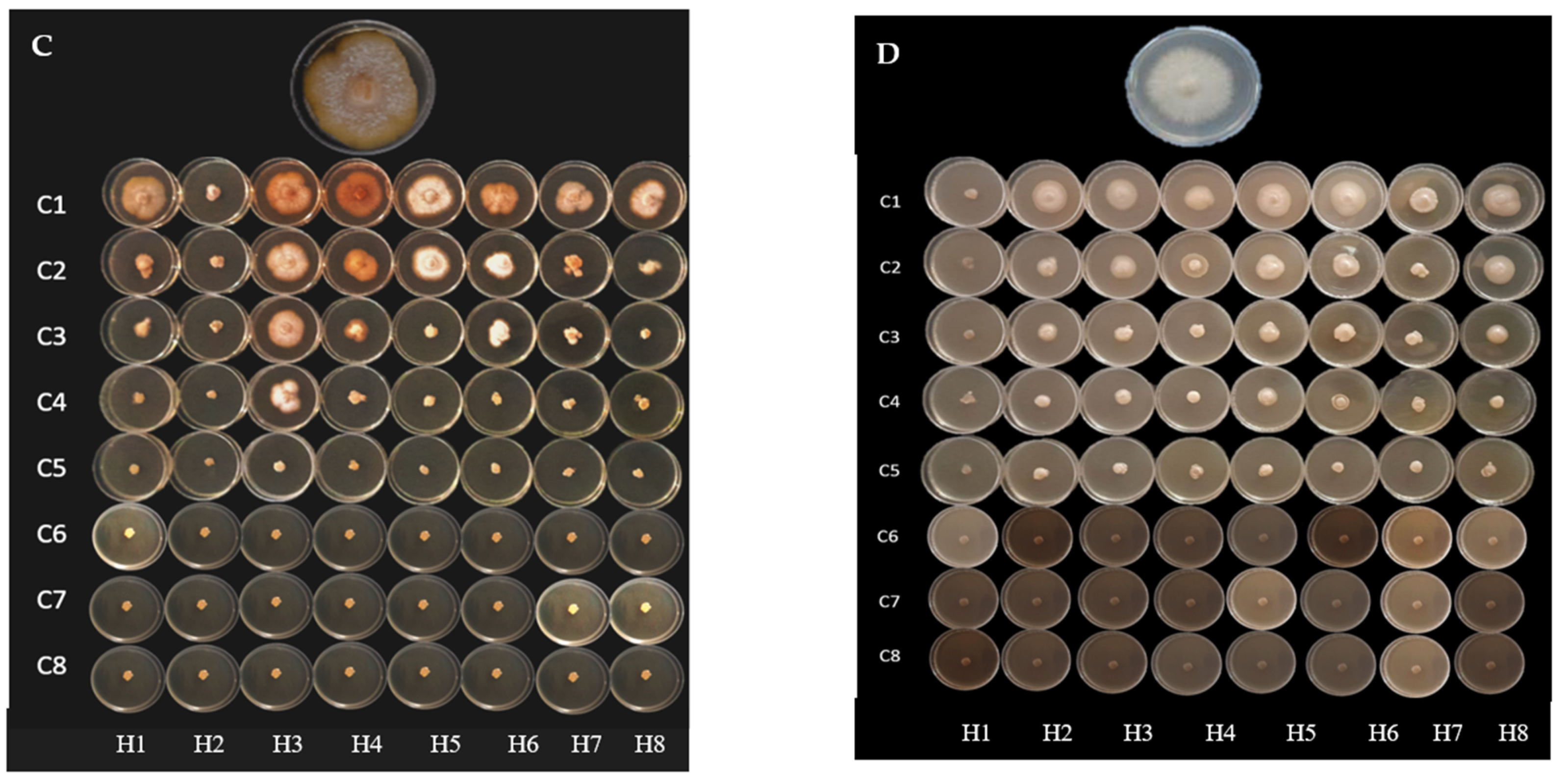
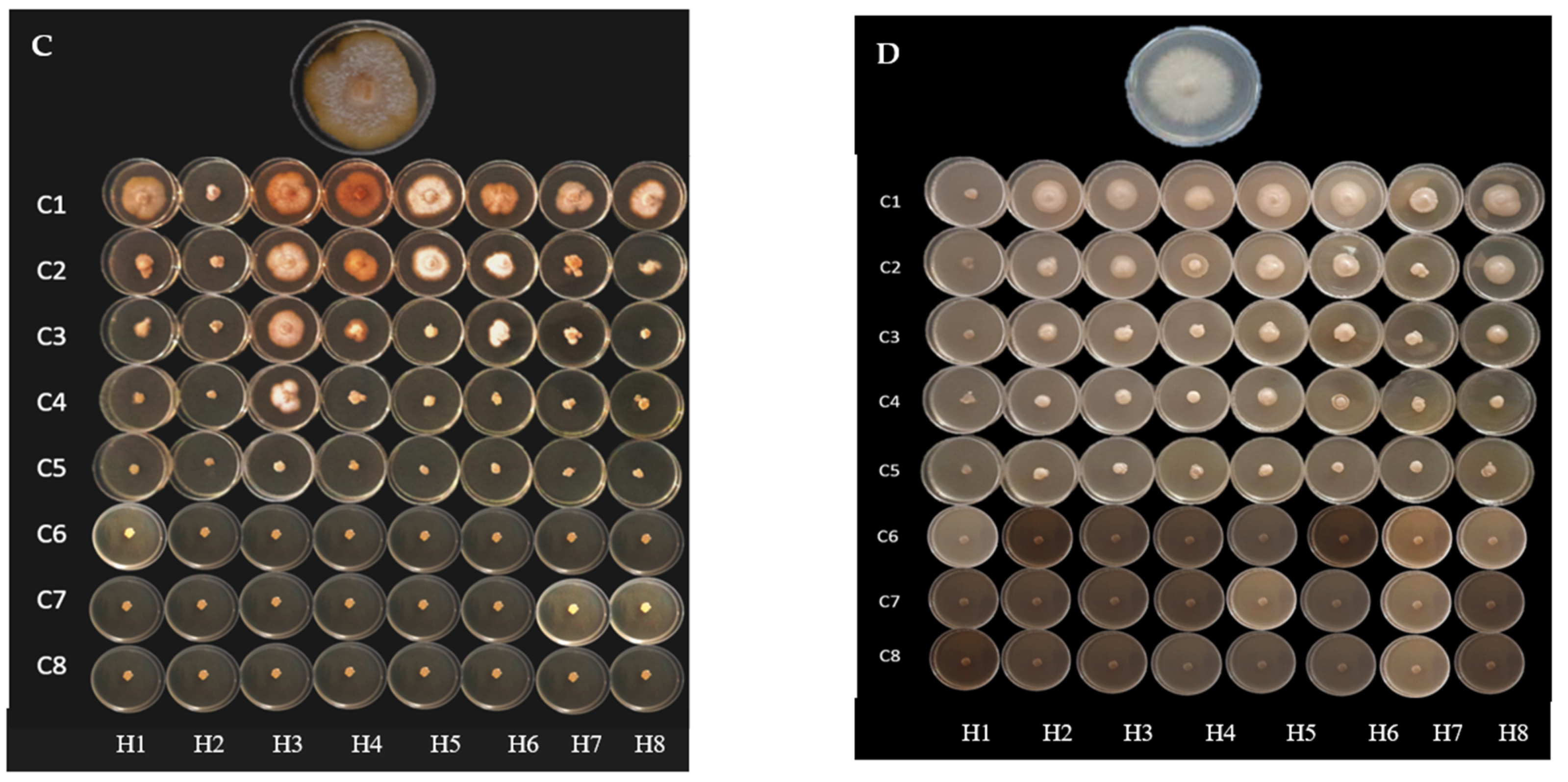
2.2. Antifungal Activity
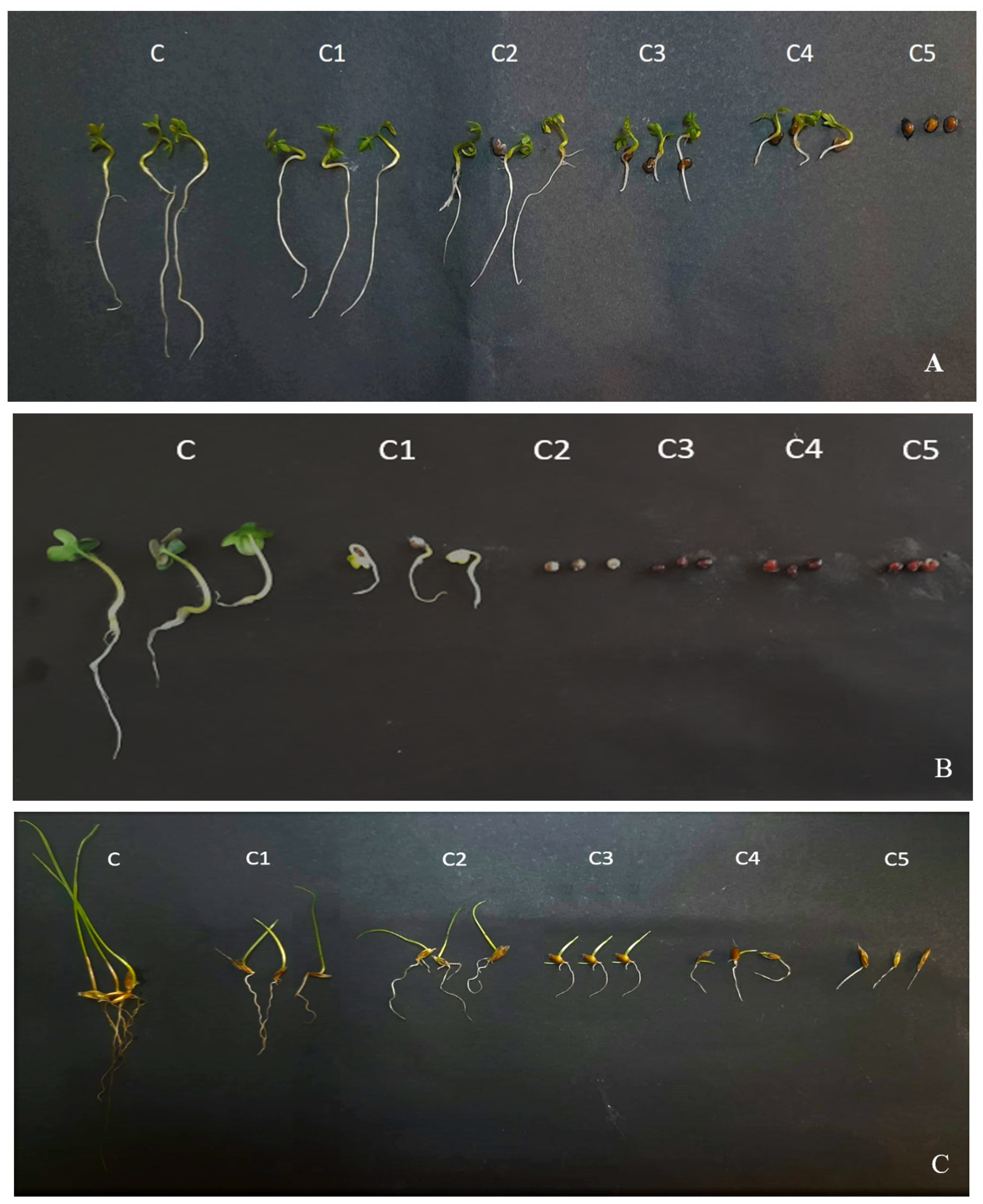
2.3. Herbicidal Effects of the EOs on Germination and Seedling Growth of Weeds and Cultivated Crops
3. Discussion
4. Materials and Methods
4.1. Plant Material
4.2. Essential Oils Extraction
4.3. Analysis of the EOs
4.4. Antifungal Activity
4.5. Effect of Eucalyptus EOs of Seed Germination and Seedling Growth
4.6. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Aktar, W.; Sengupta, D.; Chowdhury, A. Impact of Pesticides Use in Agriculture: Their Benefits and Hazards. Interdiscip. Toxicol. 2009, 2, 1–12. [Google Scholar] [CrossRef]
- Mabrouk, Y.; Mejri, S.; Belhadj, O. Biochemical mechanisms of induced resistance by rhizobial lipopolysaccharide in pea against crenate broomrape. Braz. J. Bot. 2016, 39, 107–114. [Google Scholar] [CrossRef]
- Hemissi, I.; Mabrouk, Y.; Mejri, S.; Saidi, M.; Sifi, B. Enhanced Defence Responses of Chickpea Plants Against Rhizoctonia solani by Pre-Inoculation with Rhizobia. J. Phytopathol. 2013, 161, 412–418. [Google Scholar] [CrossRef]
- Fletcher, J.; Bender, C.; Budowle, B.; Cobb, W.T.; Gold, S.E.; Ishimaru, C.A.; Tolin, S.A. Plant Pathogen Forensics: Capabilities, Needs, and Recommendations. Microbiol. Mol. Biol. Rev. 2006, 70, 450–471. [Google Scholar] [CrossRef] [PubMed]
- Mabrouk, Y.; Zourgui, L.; Sifi, S.; Belhadj, O. The potential of Rhizobium strains for biological control of Orobanche crenata. Biologia 2007, 62, 139–143. [Google Scholar] [CrossRef]
- Latiri, K.; Lhomme, J.P.; Annabi, M.; Setter, T.L. Wheat Production in Tunisia: Progress, Inter-Annual Variability and Relation to Rainfall. Eur. J. Agron. 2010, 33, 33–42. [Google Scholar] [CrossRef]
- Capinera, J.L.; Abrol, D.P. (Eds.) Integrated Pest Management: Current Concepts and Ecological Perspective. Fla. Entomol. 2014, 97, 1272. [Google Scholar] [CrossRef]
- Bouabidi, W.; Hanana, M.; Gargouri, S.; Amri, I.; Fezzani, T.; Ksontini, M.; Jamoussi, B.; Hamrouni, L. Chemical composition, phytotoxic and antifungal properties of Ruta chalepensis L. essential oils. Nat. Prod. Res. 2015, 29, 864–868. [Google Scholar] [CrossRef]
- Amri, I.; Mancini, E.; De Martino, L.; Hamrouni, L.; Hanana, M.; Jamoussi, B.; Gargouri, S.; Scognamiglio, M.; De Feo, V. Chemical Composition and Biological Activities of Tunisian Cupressus arizonica Greene Essential Oils. Chem. Biodivers. 2014, 11, 150–160. [Google Scholar] [CrossRef]
- Isman, M.B.; Miresmailli, S.; Machial, C. Commercial Opportunities for Pesticides Based on Plant Essential Oils in Agriculture, Industry, and Consumer Products. Phytochem. Rev. 2011, 10, 197–204. [Google Scholar] [CrossRef]
- Khammassi, M.; Khedhri, H.; Mighri, H.; Souihi, M.; Kochti, O.; Emine, S.; Amri, I.; Bassem, J.; Mabrouk, Y. Phytochemical Screening of Essential Oils and Methanol Extract Constituents of Wild Foeniculum vulgare Mill.: A Potential Natural Source for Bioactive Molecules. Chem. Afr. 2023, 6, 1227–1240. [Google Scholar] [CrossRef]
- Amri, I.; De Martino, L.; Marandino, A.; Hamrouni, L.; Hanana, M.; Scandolera, E.; De Feo, V.; Mancini, E. Chemical composition and biological activities of the essential oil from Artemisia herba-alba growing wild in Tunisia. Nat. Prod. Commun. 2013, 8, 407–410. [Google Scholar] [CrossRef] [PubMed]
- Amri, I.; Kouki, H.; Mabrouk, Y.; Hanana, M.; Jamoussi, B.; Hamrouni, L. Essential Oils of Tunisian Pinus radiata D. Don, Chemical Composition and Study of Their Herbicidal Activity. Vietnam J. Chem. 2021, 59, 247–252. [Google Scholar] [CrossRef]
- Tyagi, A.K.; Malik, A. Antimicrobial Potential and Chemical Composition of Eucalyptus globulus Oil in Liquid and Vapour Phase against Food Spoilage Microorganisms. Food Chem. 2011, 126, 228–235. [Google Scholar] [CrossRef]
- Limam, H.; Ben Jemaa, M.; Tammar, S.; Ksibi, N.; Khammassi, S.; Jallouli, S.; Msaada, K. Variation in chemical profile of leaves essential oils from thirteen Tunisian Eucalyptus species and evaluation of their antioxidant and antibacterial properties. Ind. Crop. Prod. 2020, 158, 112964. [Google Scholar] [CrossRef]
- Águas, A.; Ferreira, A.; Maia, P.; Fernandes, P.M.; Roxo, L.; Keizer, J.; Moreira, F. Natural Establishment of Eucalyptus globulus Labill. in Burnt Stands in Portugal. Forests 2014, 323, 47–56. [Google Scholar] [CrossRef]
- Dhakad, A.K.; Pandey, V.V.; Beg, S.; Rawat, J.M.; Singh, A. Biological, Medicinal and Toxicological Significance of Eucalyptus Leaf Essential Oil: A Review. J. Sci. Food Agric. 2017, 98, 833–848. [Google Scholar] [CrossRef]
- Ben Ghnaya, A.; Hamrouni, L.; Amri, I.; Ahoues, H.; Hanana, M.; Romane, A. Study of allelopathic effects of Eucalyptus erythrocorys L. crude extracts against germination and seedling growth of weeds and wheat. Nat. Prod. Res. 2015, 30, 2058–2064. [Google Scholar] [CrossRef]
- Polito, F.; Kouki, H.; Khedhri, S.; Hamrouni, L.; Mabrouk, Y.; Amri, I.; Nazzaro, F.; Fratianni, F.; De Feo, V. Chemical Composition and Phytotoxic and Antibiofilm Activity of the Essential Oils of Eucalyptus bicostata, E. gigantea, E. intertexta, E. obliqua, E. pauciflora, and E. tereticornis. Plants 2022, 11, 3017. [Google Scholar] [CrossRef]
- Silva, E.R.; Igartuburu, J.M.; Overbeck, G.E.; Soares, G.L.G.; Macías, F.A. Bioherbicide Potential of Eucalyptus saligna Leaf Litter Essential Oil. Chem. Biodivers. 2020, 17, e2000407. [Google Scholar] [CrossRef]
- Khedhri, S.; Polito, F.; Caputo, L.; Manna, F.; Khammassi, M.; Hamrouni, L.; Amri, I.; Nazzaro, F.; De Feo, V.; Fratianni, F. Chemical Composition, Phytotoxic and Antibiofilm Activity of Seven Eucalyptus Species from Tunisia. Molecules 2022, 27, 8227. [Google Scholar] [CrossRef]
- Amri, I.; Khammassi, M.; Gargouri, S.; Hanana, M.; Jamoussi, B.; Hamrouni, L.; Mabrouk, Y. Tunisian pine essential oils: Chemical composition, herbicidal and antifungal properties. J. Essent. Oil-Bear. Plants 2022, 25, 430–443. [Google Scholar] [CrossRef]
- Foudil-Cherif, Y.; Meklati, B.Y.; Verzera, A.; Mondello, L.; Dugo, G. Chemical Examination of Essential Oils from the Leaves of Nine Eucalyptus Species Growing in Algeria. J. Essent. Oil Res. 2000, 12, 186–191. [Google Scholar] [CrossRef]
- Farah, A.; Fechtal, M.; Chaouch, A. Effet de l’hybridation interspécifique sur la teneur et la composition chimique des huiles essentielles d’Eucalyptus cultivés au Maroc. Biotechnol. Agron. Soc. Environ. 2002, 6, 163–169. [Google Scholar]
- Ameur, E.; Sarra, M.; Yosra, D.; Mariem, K.; Nabil, A.; Lynen, F.; Larbi, K.M. Chemical Composition of Essential Oils of Eight Tunisian Eucalyptus Species and Their Antibacterial Activity against Strains Responsible for Otitis. BMC Complement. Med. Ther. 2021, 21, 209. [Google Scholar] [CrossRef]
- Elaissi, A.; Medini, H.; Marzouki, H.; Larbi Khouja, M.; Lynene, F.; Chemli, R.; Harzallah-Skhiri, F. Variation in Volatile Leaf Oils of Twelve Eucalyptus Species Harvested from Hajeb Layoun Arboreta (Tunisia). Chem. Biodivers. 2010, 7, 705–716. [Google Scholar] [CrossRef] [PubMed]
- Kouki, H.; Polito, F.; De Martino, L.; Mabrouk, Y.; Hamrouni, L.; Amri, I.; Fratianni, F.; De Feo, V.; Nazzaro, F. Chemistry and Bioactivities of Six Tunisian Eucalyptus Species. Pharmaceuticals 2022, 15, 1265. [Google Scholar] [CrossRef]
- Sadeghi, H.; Jamalpoor, S.; Shirzadi, M.H. Variability in Essential Oil of Teucrium polium L. of Different Latitudinal Populations. Ind. Crop. Prod. 2014, 54, 130–134. [Google Scholar] [CrossRef]
- Li, A.; Wu, H.; Feng, Y.; Deng, S.; Hou, A.; Che, F.; Liu, Y.; Geng, Q.; Ni, H.; Wei, Y. A Strategy of Rapidly Screening out Herbicidal Chemicals from Eucalyptus Essential Oils. Pest Manag. Sci. 2019, 76, 917–927. [Google Scholar] [CrossRef]
- Fouad, R.; Bousta, D.; El Ouali Lalami, A.; Fouad, O.; Ismail, A.; Jamoussi, B.; Greche, H. Chemical Composition and Herbicidal Effects of Essential Oils of Cymbopogon citratus (DC) Stapf, Eucalyptus cladocalyx, Origanum vulgare L. and Artemisia absinthium L. Cultivated in Morocco. J. Essent. Oil-Bear. Plants 2015, 18, 112–123. [Google Scholar] [CrossRef]
- Ben Hassine, D.; Ben Ismail, H.; Chokri, J.; Khouja, M.L.; Abderrabba, M. Chemical composition of some Tunisian Eucalyptus essential oils as obtained by hydrodistillation using Clevenger type apparatus. Biosci. Biotech. Res. Asia 2010, 7, 647–656. [Google Scholar]
- Elaissi, A.; Medini, H.; Simmonds, M.; Lynen, F.; Farhat, F.; Chemli, R.; Harzallah-Skhiri, F.; Khouja, M.L. Variation in Volatile Leaf Oils of Seven Eucalyptus Species Harvested from Zerniza Arboreta (Tunisia). Chem. Biodivers. 2011, 8, 362–372. [Google Scholar] [CrossRef]
- Ud-Daula, A.; Demirci, F.; Salim, K.A.; Demirci, B.; Lim, L.B.L.; Bber, K.H.; Ahmad, N. Chemical composition, antioxidant and antimicrobial activities of essential oils from leaves, aerial stems, basal stems, and rhizomes of Etlingera fimbriobracteata (K.Schum.) R.M.Sm. Ind. Crop. Prod. 2016, 84, 189–198. [Google Scholar] [CrossRef]
- Su, Y.C.; Ho, C.L.; Eugene, I.C.; Chang, T. Antifungal activities and chemical compositions of essential oils from leaves of four Eucalyptus. Taiwan J. For. Sci. 2006, 21, 49–61. [Google Scholar]
- Amri, I.; Khammassi, M.; Ben Ayed, R.; Khedhri, S.; Mansour, M.B.; Kochti, O.; Pieracci, Y.; Flamini, G.; Mabrouk, Y.; Gargouri, S.; et al. Essential Oils and Biological Activities of Eucalyptus falcata, E. sideroxylon and E. citriodora Growing in Tunisia. Plants 2023, 12, 816. [Google Scholar] [CrossRef] [PubMed]
- Kouki, H.; Amri, I.; Souihi, M.; Pieracci, Y.; Trabelsi, I.; Hamrouni, L.; Flamini, G.; Hirsch, A.; Mabrouk, Y. Chemical composition, antioxidant, herbicidal, and antifungal activities of leaf essential oils from three Tunisian Eucalyptus species. J. Plant Dis. Prot. 2023, in press. [Google Scholar] [CrossRef]
- Ivanov, M.; Kannan, A.; Stojković, D.S.; Glamočlija, J.; Calhelha, R.C.; Ferreira, I.C.F.R.; Sanglard, D.; Soković, M. Camphor and Eucalyptol-Anticandidal Spectrum, Antivirulence Effect, Efflux Pumps Interference and Cytotoxicity. Int. J. Mol. Sci. 2021, 22, 483. [Google Scholar] [CrossRef]
- Kim, E.; Park, I.-K. Fumigant Antifungal Activity of Myrtaceae Essential Oils and Constituents from Leptospermum petersonii against Three Aspergillus Species. Molecules 2012, 17, 10459–10469. [Google Scholar] [CrossRef] [PubMed]
- Cox, S.; Mann, C.; Markham, J.; Bell, H.; Gustafson, J.; Warmington, J.; Wyllie, S. The mode of antimicrobial action of the essential oil of Melaleuca alternifolia (tea tree oil). J. Appl. Microbiol. 2000, 88, 170–175. [Google Scholar] [CrossRef] [PubMed]
- Lima, I.O.; de Araújo, G.; Oliveira, R.; Lima, E.d.O.; de Souza, E.L.; Farias, N.P.; de Fátima Navarro, D. Inhibitory effect of some phytochemicals in the growth of yeasts potentially causing opportunistic infections. Rev. Bras. Ciências Farm. 2005, 41, 199–203. [Google Scholar] [CrossRef]
- Tangarife-Castaño, V.; Correa-Royero, J.; Zapata-Londoño, B.; Durán, C.; Stanshenko, E.; Mesa-Arango, A.C. Anti-Candida albicans activity, cytotoxicity and interaction with antifungal drugs of essential oils and extracts from aromatic and medicinal plants. Infectio 2011, 15, 160–167. [Google Scholar] [CrossRef]
- Kordali, S.; Kotan, R.; Cakir, A. Screening of antifungal activities of 21 oxygenated monoterpenes in-vitro as plant disease control agents. Allelopath J. 2007, 19, 373–392. [Google Scholar]
- Zhao, L.; Yany, X.; Li, X.; Mu, W.; Liu, F. Antifungal, insecticidal and herbicidal properties of volatile components from Paenibacillus polymyxa strain BMP-11. Agric. Sci. China 2011, 10, 728–736. [Google Scholar] [CrossRef]
- García, R.; Alves, E.S.S.; Santos, M.P.; Viégas Aquije, G.M.F.; Fernandes, A.A.R.; Dos Santos, R.B.; Ventura, J.A.; Fernandes, P.M.B. Antimicrobial activity and potential use of monoterpenes as tropical fruits preservatives. Braz. J. Microbiol. 2008, 39, 163–168. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.-M.; Kwon, H.; Kim, K.; Lee, S.-E. Antifungal and Antiaflatoxigenic Activities of 1,8-Cineole and t-Cinnamaldehyde on Aspergillus flavus. Appl. Sci. 2018, 8, 1655. [Google Scholar] [CrossRef]
- Yu, D.; Wang, J.; Shao, X.; Xu, F.; Wang, H. Antifungal modes of action of tea tree oil and its two characteristic components against Botrytis cinerea. J. Appl. Microbiol. 2015, 119, 1253–1262. [Google Scholar] [CrossRef] [PubMed]
- Cristani, M.; D’Arrigo, M.; Mandalari, G.; Castelli, F.; Sarpietro, M.G.; Micieli, D.; Venuti, V.; Bisignano, G.; Saija, A.; Trombetta, D. Interaction of four monoterpenes contained in essential oils with model membranes: Implications for their antibacterial activity. J. Agric. Food Chem. 2007, 55, 6300–6308. [Google Scholar] [CrossRef]
- Khammassi, M.; Polito, F.; Amri, I.; Khedhri, S.; Hamrouni, L.; Nazzaro, F.; Fratianni, F.; De Feo, V. Chemical Composition and Phytotoxic, Antibacterial and Antibiofilm Activity of the Essential Oils of Eucalyptus occidentalis, E. striaticalyx and E. stricklandii. Molecules 2022, 27, 5820. [Google Scholar] [CrossRef]
- Batish, D.R.; Singh, H.P.; Nidhi, S.; Shalinder, K.; Kohli, R.K. Chemical composition and inhibitory activity of essential oil from decaying leaves of Eucalyptus citriodora. Z. Naturforsch. 2006, 61, 52–56. [Google Scholar] [CrossRef]
- Singh, H.P.; Batish, D.R.; Setia, N.; Kohli, R.K. Herbicidal activity of volatile oils from Eucalyptus citriodora against Parthenium hysterophorus. Ann. Appl. Biol. 2005, 146, 189–194. [Google Scholar] [CrossRef]
- Zeng, R.S.; Li, P.W. Allelopathic effect of Eucalytus exserta and E. urophylla. J. South China Agric. Univ. 1997, 18, 6–10. [Google Scholar]
- Zhang, J.; An, M.; Wu, H.; Liu, D.L.; Stanton, R. Chemical Composition of Essential Oils of Four Eucalyptus Species and Their Phytotoxicity on Silver Leaf Nightshade (Solanum elaeagnifolium Cav) in Australia. Plant Growth Regul. 2012, 68, 231–237. [Google Scholar] [CrossRef]
- Singh, H.P.; Batish, R.D.; Kaur, S.; Arora, K.; Kohli, K.R. α-Pinene inhibits growth and induces oxidative stress in roots. Ann. Bot. 2006, 98, 1261–1269. [Google Scholar] [CrossRef] [PubMed]
- Romagni, J.G.; Allen, S.N.; Dayan, F.E. Allelopathic Effects of Volatile Cineoles on Two Weedy Plant Species. J. Chem. Ecol. 2000, 26, 303–313. [Google Scholar] [CrossRef]
- De Martino, L.; Mancini, E.; Almeida, L.F.R.; De Feo, V. The antigerminative activity of twenty-seven monoterpenes. Molecules 2010, 15, 6630–6637. [Google Scholar] [CrossRef] [PubMed]
- Nishida, N.; Tamotsu, S.; Nagata, N.; Saito, C.; Sakai, A. Allelopathic effects of volatile monoterpenoids produced by Salvia leucophylla: Inhibition of cell proliferation and DNA synthesis in the root apical meristem of Brassica campestris seedlings. J. Chem. Ecol. 2005, 31, 1187–1203. [Google Scholar] [CrossRef]
- Mahdavikia, F.; Saharkhiz, M.J.; Karami, A. Defensive response of radish seedlings to the oxidative stress arising from phenolic compounds in the extract of peppermint (Mentha x piperita L). Sci. Hortic. 2017, 214, 133–140. [Google Scholar] [CrossRef]
- Fagodia, S.K.; Singh, H.P.; Batish, D.R.; Kohli, R.K. Phytotoxicity and cytotoxicity of Citrus aurantiifolia essential oil and its major constituents: Limonene and citral. Ind. Crop. Prod. 2017, 108, 708–715. [Google Scholar] [CrossRef]
- Adams, R.P. Identification of Essential Oil Components by Gas Chromatography/Mass Spectrometry; Allured Publishing Co.: Carol Stream, IL, USA, 2007. [Google Scholar]
- Cakir, A.; Kordali, S.; Kilic, H.; Kaya, E. Antifungal properties of essential oil and crude extracts of Hypericum linarioides Bosse. Biochem. Syst. Ecol. 2005, 33, 245–256. [Google Scholar] [CrossRef]
- Tworkoski, T. Herbicide effects of essential oils. Weed Sci. 2002, 50, 425–431. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
| Compounds | RT (min) | Class | A | B | C | D | E | F | G | H | Identification |
|---|---|---|---|---|---|---|---|---|---|---|---|
| α-pinene | 6.930 | MH | 6.69 | 5.6 | 3.04 | 2.5 | 9.19 | 0.8 | 14.49 | 9.48 | MS, LRI, IN |
| Camphene | 7.214 | MH | - | 0.94 | 0.63 | 1.1 | - | - | 0.21 | - | MS, LRI |
| β-pinene | 7.884 | MH | 8.85 | - | - | - | - | - | 0.59 | - | MS, LRI, IN |
| α-phellandrene | 8.552 | MH | - | - | 0.56 | 0.61 | - | - | 1.5 | 0.46 | MS, LRI, IN |
| p-cymene | 8.713 | MH | - | 44.79 | 15.27 | - | 20.88 | 1.9 | - | MS, LRI, IN | |
| Eucalyptol | 9.193 | OM | 44.68 | 60.5 | 6.36 | 20.36 | 66.67 | 3.22 | 26.3 | 55.93 | MS, LRI, IN |
| γ-terpinene | 9.862 | MH | - | - | 0.2 | - | - | 4.65 | 0.58 | - | MS, LRI, IN |
| cis-linalool oxide | 10.190 | OM | - | - | 0.35 | 0.82 | - | 0.56 | 0.22 | - | MS, LRI |
| Verbenol | 10.580 | OM | 0.37 | 0.38 | 0.56 | 0.57 | - | 0.47 | 0.37 | 0.23 | MS, LRI |
| Isopentyl valerate | 10.792 | NT | 0.34 | 1.22 | 0.23 | 0.42 | 0.32 | 0.28 | 2.18 | 0.87 | MS, LRI |
| Fenchol | 10.987 | OM | 0.35 | 2.97 | 1.91 | 3.72 | 0.4 | 0.89 | 0.6 | 0.2 | MS, LRI |
| α-campholenal | 11.230 | OM | 1.26 | 0.23 | - | 0.45 | - | - | 0.59 | - | MS, LRI |
| trans-pinocarveol | 11.878 | OM | 6.63 | 7.71 | 7.39 | 4.35 | 8.91 | 0.47 | 4.42 | 17.54 | MS, LRI |
| Verbenol | 12.284 | OM | 1.57 | - | - | 0.2 | - | - | 0.7 | - | MS, LRI, IN |
| Pinocarvone | 12.381 | OM | 0.2 | 6.72 | 3.42 | 1.64 | 1.88 | - | 2.51 | - | MS, LRI |
| Isoborneol | 12.478 | OM | 3.34 | 6.23 | 4.81 | 10.54 | 0.59 | 0.68 | 1.33 | 5.53 | MS, LRI |
| Terpinen-4-ol | 12.700 | OM | 1.1 | 0.34 | 1.08 | 0.65 | 0.97 | 4.03 | 0.97 | - | MS, LRI, IN |
| α-terpineol | 12.915 | OM | 8.6 | 3.41 | 6.46 | 7.44 | 1.68 | 17.72 | 3.85 | 1.02 | MS, LRI, IN |
| Myrtenol | 13.159 | OM | 0.86 | 0.34 | 0.43 | 0.24 | - | - | 0.35 | 0.43 | MS, LRI, IN |
| trans-pulegol | 13.277 | OM | - | - | 0.24 | - | - | 1.21 | 0.48 | - | MS, LRI, IN |
| Verbenone | 13.393 | OM | 1.56 | - | - | - | - | - | 0.2 | - | MS, LRI |
| cis-carveol | 13.624 | OM | 0.78 | 0.26 | 0.37 | 0.42 | - | 0.67 | 0.29 | - | MS, LRI |
| Verbenol | 13.823 | OM | 0.73 | 0.88 | 2.51 | 1.14 | 0.33 | 3.13 | 0.39 | 0.67 | MS, LRI |
| Cumaldehyde | 14.053 | OM | 2.06 | 0.26 | 0.31 | - | 0.2 | 1.12 | 0.35 | 0.32 | MS, LRI |
| Piperetone | 14.398 | OM | 0.29 | - | 0.29 | - | - | 0.7 | 1.61 | - | MS, LRI |
| Phellandral | 15.176 | OM | 0.32 | - | - | - | - | - | - | - | MS, LRI |
| p-cymen-7-ol | 15.248 | OM | 0.48 | - | - | 0.28 | - | - | - | - | MS, LRI |
| Thymol | 15.316 | PP | - | - | 1.21 | - | - | 2.8 | - | - | MS, LRI, IN |
| Carvacrol | 15.419 | PP | 0.37 | - | - | 0.4 | - | 1.07 | 1.02 | - | MS, LRI, IN |
| α-terpinyl acetate | 16.882 | OM | - | - | - | 0.34 | - | 27.44 | 0.33 | - | MS, LRI |
| Linalyl isobutyrate | 16.950 | OM | - | - | - | 0.88 | - | - | 0.21 | - | MS, LRI |
| Aromandendrene | 18.281 | SH | - | - | 0.38 | 7.2 | - | - | 2.37 | - | MS, LRI |
| Isoamyl phenylacetate | 19.098 | PP | 0.22 | 0.33 | - | 0.21 | - | 1.14 | 1.11 | 0.4 | MS, LRI |
| Spathulenol | 20.188 | OS | 1.35 | 0.3 | 3.1 | 4.1 | 1.69 | 0.36 | - | 0.88 | MS, LRI, IN |
| Caryophyllene oxide | 20.925 | OS | - | - | - | - | - | - | 2.01 | - | MS, LRI |
| Viridiflorol | 21.113 | OS | 3.44 | - | 2.04 | 2.63 | 1.45 | 0.32 | 8.13 | 0.53 | MS, LRI |
| Ledol | 21.225 | OS | 0.77 | - | 2.39 | 3.83 | 0.81 | - | 4.31 | 0.36 | MS, LRI |
| Guaiol | 21.687 | OS | - | - | - | 3.13 | - | - | 2.46 | 0.22 | MS, LRI |
| β-eudesmol | 21.758 | OS | - | - | 1.05 | 0.28 | 0.78 | - | 2.04 | 1.92 | MS, LRI |
| α-eudesmol | 22.278 | OS | - | - | 0.69 | 0.29 | 0.63 | 0.23 | 5.58 | - | MS, LRI |
| Eicosane | 31.482 | NT | - | - | 0.27 | - | 0.47 | 1.47 | - | 0.21 | MS, LRI |
| Total identification | 97.21 | 98.62 | 97.07 | 96.01 | 96.97 | 96.31 | 96.55 | 97.2 | |||
| * Yield (w/w %) | 0.2 b | 0.56 c | 0.81 e | 0.64 d | 1 f | 0.59 c,d | 0.12 a | 1.32 g | |||
| Monoterpene hydrocarbons | (MH)% | 15.54 | 6.54 | 49.22 | 19.48 | 9.19 | 26.33 | 19.27 | 9.94 | ||
| Oxygentated monoterpenes | (OS)% | 75.18 | 90.23 | 36.49 | 54.04 | 81.63 | 62.31 | 46.07 | 81.87 | ||
| Sesquiterpene hydrocarbons | (SH)% | - | - | 0.38 | 7.2 | - | - | 2.37 | - | ||
| Oxygentated sesquiterpenes | (OS)% | 5.56 | 0.3 | 9.27 | 14.26 | 5.36 | 0.91 | 24.53 | 3.91 | ||
| Phenylpropanoids | (PP)% | 0.59 | 0.33 | 1.21 | 0.61 | 0 | 5.01 | 2.13 | 0.4 | ||
| Non-terpene derivatives | (NT)% | 0.34 | 1.22 | 0.5 | 0.42 | 0.79 | 1.75 | 2.18 | 1.08 |
| Eucalyptus Species | Doses (μL/mL) | F. oxysporum Solani | F. culmorum | F. oxysporum f. sp. matthioli | F. redolens |
|---|---|---|---|---|---|
| E. cladocalyx | 0 | 0 ± 0 A,a | 0 ± 0 A,a | 0 ± 0 A,a | 0 ± 0 A,a |
| 1 | 78.84 ± 0.64 B,b | 58.88 ± 0.64 B,c | 85.55 ± 0.32 B,b | 100 ± 0 B,a | |
| 2 | 90 ± 0 C,c | 100 ± 0 C,a | 94.44 ± 0 C,b | 100 ± 0 B,a | |
| 3 | 100 ± 0 D,a | 100 ± 0 C,a | 100 ± 0 D,a | 100 ± 0 B,a | |
| 4 | 100 ± 0 D,a | 100 ± 0 C,a | 100 ± 0 D,a | 100 ± 0 B,a | |
| 5 | 100 ± 0 D,a | 100 ± 0 C,a | 100 ± 0 D,a | 100 ± 0 B,a | |
| 6 | 100 ± 0 D,a | 100 ± 0 C,a | 100 ± 0 D,a | 100 ± 0 B,a | |
| 7 | 100 ± 0 D,a | 100 ± 0 C,a | 100 ± 0 D,a | 100 ± 0 B,a | |
| 8 | 100 ± 0 D,a | 100 ± 0 C,a | 100 ± 0 D,a | 100 ± 0 B,a | |
| E. microcorys | 0 | 0 ± 0 A,a | 0 ± 0 A,a | 0 ± 0 A,a | 0 ± 0 A,a |
| 1 | 74.44 ± 0 B,a | 44.44 ± 0 B,c | 76.66 ± 0.32 B,a | 67.18 ± 0.25 B,b | |
| 2 | 92.22 ± 0.64 C,a | 63.32 ± 0.32 C,c | 81.10 ± 0.32 C,b | 81.53 ± 0 C,b | |
| 3 | 93.33 ± 0.02 D,a | 74.44 ± 0.64 E,c | 94.44 ± 0 D,a | 84.61 ± 0.01 D,b | |
| 4 | 100 ± 0 E,a | 88.88 ± 0 F,c | 96.66 ± 0 E,b | 90.25 ± 0.51 E,d | |
| 5 | 100 ± 0 E,a | 94.44 ± 0 G,b | 97.77 ± 0 F,b | 95.38 ± 0 F,b | |
| 6 | 100 ± 0 E,a | 100 ± 0 G,a | 100 ± 0 G,a | 100 ± 0 G,a | |
| 7 | 100 ± 0 E,a | 100 ± 0 G,a | 100 ± 0 G,a | 100 ± 0 G,a | |
| 8 | 100 ± 0 E,a | 100 ± 0 G,a | 100 ± 0 G,a | 100 ± 0 G,a | |
| E. resinifera | 0 | 0 ± 0 A,a | 0 ± 0 A,a | 0 ± 0 A,a | 0 ± 0 A,a |
| 1 | 70 ± 1.92 B,d | 44.44 ± 0.64 B,c | 59.99 ± 0.32 B,f | 65.38 ± 0.44 B,f | |
| 2 | 84.44 ± 0.25 C,a | 68.88 ± 0.32 C,c | 83.33 ± 0 C,a | 78.72 ± 0.25 C,b | |
| 3 | 87.77 ± 0.64 D,b | 88.88 ± 0.67 D,b | 97.77 ± 0.57 D,a | 86.15 ± 0 D,b | |
| 4 | 100 ± 0 E,a | 91.11 ± 0 E,b | 100 ± 0 E,a | 89.23 ± 0.01 E,e | |
| 5 | 100 ± 0 E,a | 100 ± 0 F,a | 100 ± 0 E,a | 100 ± 0 F,a | |
| 6 | 100 ± 0 E,a | 100 ± 0 F,a | 100 ± 0 E,a | 100 ± 0 F,a | |
| 7 | 100 ± 0 E,a | 100 ± 0 F,a | 100 ± 0 E,a | 100 ± 0 F,a | |
| 8 | 100 ± 0 E,a | 100 ± 0 F,a | 100 ± 0 E,a | 100 ± 0 F,a | |
| E. saligna | 0 | 0 ± 0 A,a | 0 ± 0 A,a | 0 ± 0 A,a | 0 ± 0 A,a |
| 1 | 64.44 ± 0 B,d | 59.66 ± 0.11 B,a | 64.44 ± 0 B,d | 75.38 ± 0.04 B,c | |
| 2 | 71.11 ± 0 C,b | 72.22 ± 0.28 C,b | 81.55 ± 0.19 C,a | 78.46 ± 0.02 C,b | |
| 3 | 92.22 ± 0.75 D,e | 77.77 ± 0.51 D,c | 83.33 ± 0.32 D,f | 90.76 ± 0 D,b | |
| 4 | 97.77 ± 0 E,b | 81.11 ± 0 E,e | 96.66 ± 0.19 E,b | 92.3 ± 0 E,b | |
| 5 | 100 ± 0 F,a | 100 ± 0 F,a | 100 ± 0 F,a | 100 ± 0 F,a | |
| 6 | 100 ± 0 F,a | 100 ± 0 F,a | 100 ± 0 F,a | 100 ± 0 F,a | |
| 7 | 100 ± 0 F,a | 100 ± 0 F,a | 100 ± 0 F,a | 100 ± 0 F,a | |
| 8 | 100 ± 0 F,a | 100 ± 0 F,a | 100 ± 0 F,a | 100 ± 0 F,a | |
| E. angulosa | 0 | 0 ± 0 A,a | 0 ± 0 A,a | 0 ± 0 A,a | 0 ± 0 A,a |
| 1 | 66.66 ± 0.57 B,d,e | 44.44 ± 0.25 B,c | 58.88 ± 0.32 B,g | 66.15 ± 0.0 B,f | |
| 2 | 79.99 ± 0.64 C,e | 61.11 ± 0 C,a | 80 ± 0.57 C,e | 76.92 ± 0 C,d | |
| 3 | 83.33 ± 0 D,b | 73.33 ± 0 D,c | 87.77 ± 0 D,a | 83.07 ± 0.04 D,b | |
| 4 | 91.11 ± 0.64 E,a | 83.33 ± 1.15 E,b | 90 ± 1.15 E,a | 86.61 ± 0 E,b | |
| 5 | 100 ± 0 F,a | 100 ± 0 F,a | 94.44 ± 0.06 F,c | 91.79 ± 0.51 F,c | |
| 6 | 100 ± 0 F,a | 100 ± 0 F,a | 95.55 ± 0 F,b | 100 ± 0 G,a | |
| 7 | 100 ± 0 F,a | 100 ± 0 F,a | 97.77 ± 0 G,b | 100 ± 0 G,a | |
| 8 | 100 ± 0 F,a | 100 ± 0 F,a | 98.88 ± 0 G,b | 100 ± 0 G,a | |
| E. diversicolor | 0 | 0 ± 0 A,a | 0 ± 0 A,a | 0 ± 0 A,a | 0 ± 0 A,a |
| 1 | 66.66 ± 0.64 B,d,e | 44.44 ± 0.64 B,c | 64.44 ± 0.02 B,d | 63.84 ± 0.44 B,g | |
| 2 | 71.11 ± 0.64 C,f | 56.66 ± 0.64 C,g | 75.55 ± 0.02 C,f | 75.38 ± 0.01 C,e | |
| 3 | 74.44 ± 0 D,h | 72.22 ± 0 D,e | 81.11 ± 0.32 D,g | 84.61 ± 0 D,e | |
| 4 | 79.25 ± 0.98 E,d | 77.77 ± 0.44 E,a | 87.77 ± 0.57 E,e | 88.46 ± 0.02 E,f | |
| 5 | 95.55 ± 0 F,b | 81.11 ± 0 F,d | 90 ± 0 F,c | 100 ± 0 F,a | |
| 6 | 100 ± 0 G,a | 83.33 ± 0 G,c | 96.66 ± 0 G,b | 100 ± 0 F,a | |
| 7 | 100 ± 0 G,a | 91.11 ± 0.64 H,c | 100 ± 0 H,a | 100 ± 0 F,a | |
| 8 | 100 ± 0 G,a | 100 ± 0 I,a | 100 ± 0 H,a | 100 ± 0 F,a | |
| E. ovata | 0 | 0 ± 0 A,a | 0 ± 0 A,a | 0 ± 0 A,a | 0 ± 0 A,a |
| 1 | 92.22 ± 0.64 B,a | 47.7 ± 0 B,d | 68.88 ± 0.32 B,c | 76.40 ± 0.51 B,b | |
| 2 | 95.55 ± 0 C,a | 58.88 ± 0 C,d | 80 ± 0 C,c | 82.04 ± 0.25 C,b | |
| 3 | 97.77 ± 0 D,a | 72.22 ± 0.32 D,c | 88.88 ± 0 D,b | 89.23 ± 0 D,b | |
| 4 | 98.88 ± 0 E,ab | 83.33 ± 0 E,c | 96.33 ± 0.11 E,b | 92.3 ± 1.73 E,b | |
| 5 | 100 ± 0 F,a | 88.88 ± 0 F,c | 100 ± 0 F,a | 100 ± 0 F,a | |
| 6 | 100 ± 0 F,a | 91.11 ± 0 G,b | 100 ± 0 F,a | 100 ± 0 F,a | |
| 7 | 100 ± 0 F,a | 94.44 ± 0 H,b | 100 ± 0 F,a | 100 ± 0 F,a | |
| 8 | 100 ± 0 F,a | 100 ± 0 I,a | 100 ± 0 F,a | 100 ± 0 F,a | |
| E. sargentii | 0 | 0 ± 0 A,a | 0 ± 0 A,a | 0 ± 0 A,a | 0 ± 0 A,a |
| 1 | 66.66 ± 0.64 B,d,e | 44.44 ± 0 B,c | 61.11 ± 0.32 B,e | 70.76 ± 0.01 B,d | |
| 2 | 88.88 ± 0.57 C,c | 63.33 ± 1.15 C,d | 78.88 ± 0.32 C,e | 72.3 ± 0.44 C,f | |
| 3 | 94.44 ± 0 D,c | 74.44 ± 0.32 D,d | 87.77 ± 0.32 D,e | 75.12 ± 0.25 D,g | |
| 4 | 100 ± 0 E,a | 88.88 ± 0 E,d | 94.44 ± 0 E,b | 91.53 ± 0.01 E,c | |
| 5 | 100 ± 0 E,a | 94.44 ± 0.57 F,b | 100 ± 0 F,a | 100 ± 0 F,a | |
| 6 | 100 ± 0 E,a | 100 ± 0 G,a | 100 ± 0 F,a | 100 ± 0 F,a | |
| 7 | 100 ± 0 E,a | 100 ± 0 G,a | 100 ± 0 F,a | 100 ± 0 F,a | |
| 8 | 100 ± 0 E,a | 100 ± 0 G,a | 100 ± 0 F,a | 100 ± 0 F,a |
| F. oxysporum Solani | F. culmorum | F. oxysporum f. matthioli | F. redolens | |||||
|---|---|---|---|---|---|---|---|---|
| Eucalyptus species | MIC (μL/mL) | MFC (μL/mL) | MIC (μL/mL) | MFC (μL/mL) | MIC (μL/mL) | MFC (μL/mL) | MIC (μL/mL) | MFC (μL/mL) |
| E. cladocalyx | 3 | 3 | 2 | 3 | 3 | 3 | 1 | 3 |
| E. microcorys | 4 | >8 | 6 | 8 | 6 | >8 | 6 | 7 |
| E. resinifera | 4 | >8 | 5 | 8 | 4 | 8 | 5 | 8 |
| E. saligna | 5 | >8 | 5 | 7 | 5 | >8 | 5 | 8 |
| E. angulosa | 5 | >8 | 5 | 7 | 8 | 6 | 6 | 7 |
| E. diversicolor | 6 | >8 | 8 | 8 | 7 | >8 | 5 | >8 |
| E. ovata | 5 | >8 | 8 | >8 | 5 | >8 | 5 | >8 |
| E. sargentii | 4 | >8 | 6 | 6 | 5 | 6 | 5 | 6 |
| Percentage Inhibition of Radial Growth | |||||||||
|---|---|---|---|---|---|---|---|---|---|
| Fosetyl-Al Doses (mg/mL) | 0 | 0.25 | 0.5 | 1 | 1.5 | 2 | 3 | MIC (mg/mL) | MFC (mg/mL) |
| Fungal strains | |||||||||
| F. oxysporumsolani | 0 ± 0 A | 56.64 ± 1.37 B | 90.02 ± 0.86 C | 100 ± 0 D | 100 ± 0 D | 100 ± 0 D | 100 ± 0 D | 1 | >3 |
| F. culmorum | 0 ± 0 A | 42.05 ± 0.64 B | 82.24 ± 0.43 C | 94.86 ± 0.48 D | 100 ± 0 E | 100 ± 0 E | 100 ± 0 E | 1.5 | 3 |
| F. oxysporumf. sp. matthioli | 0 ± 0 A | 37.78 ± 1.11 B | 55.91 ± 0.68 C | 91.59 ± 0.75 D | 100 ± 0 E | 100 ± 0 E | 100 ± 0 E | 1.5 | >3 |
| F. redolens | 0 ± 0 A | 39.02 ± 1.40 B | 53.66 ± 0 C | 95.12 ± 0 D | 96.26 ± 0.08 D,E | 97.56 ± 0 E | 100 ± 0 F | 3 | >3 |
| Tested Plants | Doses (μL/mL) | E. cladocalyx | E. microcorys | E. resinifera | E. saligna | E. angulosa | E. diversicolor | E. ovata | E. sargentii | Glyphosate |
|---|---|---|---|---|---|---|---|---|---|---|
| Trifolium campestre Schreb. | 0 | 100 ± 0 A,a | 100 ± 0 A,a | 100 ± 0 A,a | 100 ± 0 A,a | 100 ± 0 A,a | 100 ± 0 A,a | 100 ± 0 A,a | 100 ± 0 A,a | 100 ± 0 A,a |
| 1 | 66.7 ± 3.3 B,b | 83.3 ± 3.3 B,a | 83.3 ± 3.3 B,a | 70 ± 0 B,b | 90 ± 0 B,a | 86.7 ± 3.3 B,a | 90 ± 0 B,a | 90 ± 0 B,a | 83.3 ± 3.3 B,a | |
| 2 | 33.3 ± 3.3 C,d | 56.7 ± 3.3 C,c | 70 ± 0 C,b | 33.3 ± 3.3 C,d | 80 ± 0 C,a,b | 76.7 ± 3.3 C,a,b | 80 ± 0 C,a,b | 83.3 ± 3.3 C,a | 73.3 ± 3.3 C,a,b | |
| 3 | 0 ± 0 D,f | 10 ± 0 D,e | 50 ± 0 D,c | 16.7 ± 3.3 D,d | 60 ± 0 D,b | 70 ± 0 C,a | 70 ± 0 D,a | 70 ± 0 D,a | 70 ± 0 C,a | |
| 4 | 0 ± 0 D,c | 0 ± 0 E,c | 0 ± 0 E,c | 0 ± 0 E,c | 0 ± 0 E,c | 53.3 ± 3.3 D,b | 0 ± 0 E,c | 60 ± 0 E,a | 56.7 ± 3.3 D,a,b | |
| 5 | 0 ± 0 D,d | 0 ± 0 E,d | 0 ± 0 E,d | 0 ± 0 E,d | 0 ± 0 E,d | 26.7 ± 3.3 E,c | 0 ± 0 E,d | 43.3 ± 3.3 F,b | 50 ± 0 D,a | |
| Lolium rigidum Gaudich. | 0 | 76.7 ± 3.3 A,a | 76.7 ± 3.3 A,a | 76.7 ± 3.3 A,a | 76.7 ± 3.3 A,a | 76.7 ± 3.3 A,a | 76.7 ± 3.3 A,a | 76.7 ± 3.3 A,a | 76.7 ± 3.3 A,a | 76.7 ± 3.3 A,a |
| 1 | 63.3 ± 3.3 B,a | 63.3 ± 3.3 B,a | 36.7 ± 3.3 B,c | 50 ± 0 B,b | 53.3 ± 3.3 B,a,b | 56.7 ± 3.3 B,a,b | 50 ± 0 B,b | 60 ± 0 B,a,b | 0 ± 0 B,d | |
| 2 | 53.3 ± 3.3 C,a | 36.7 ± 3.3 C,c,d | 30 ± 0 B,C,d | 0 ± 0 C,f | 16.7 ± 3.3 C,e | 40 ± 0 Cb,c | 30 ± 0 C,d | 40 ± 0 C,b,c | 0 ± 0 B,f | |
| 3 | 0 ± 0 D,e | 0 ± 0 D,e | 23.3 ± 3.3 C,D,c | 0 ± 0 C,e | 0 ± 0 D,e | 30 ± 0 D,b | 10 ± 0 D,d | 36.7 ± 3.3 C,a | 0 ± 0 B,e | |
| 4 | 0 ± 0 D,c | 0 ± 0 D,c | 16.7 ± 3.3 D,b | 0 ± 0 C,c | 0 ± 0 D,c | 0 ± 0 E,c | 0 ± 0 E,c | 30 ± 0 D,a | 0 ± 0 B,c | |
| 5 | 0 ± 0 D,a | 0 ± 0 D,a | 0 ± 0 E,a | 0 ± 0 C,a | 0 ± 0 D,a | 0 ± 0 E,a | 0 ± 0 E,a | 0 ± 0 E,a | 0 ± 0 B,a | |
| Sinapis arvensis L. | 0 | 93.3 ± 3.3 A,a | 93.3 ± 3.3 A,a | 93.3 ± 3.3 A,a | 93.3 ± 3.3 A,a | 93.3 ± 3.3 A,a | 93.3 ± 3.3 A,a | 93.3 ± 3.3 A,a | 93.3 ± 3.3 A,a | 93.3 ± 3.3 A,a |
| 1 | 0 ± 0 B,f | 0 ± 0 B,f | 33.3 ± 3.3 B,d | 30 ± 0 B,d | 50 ± 0 B,b | 40 ± 0 B,c | 0 ± 0 B,f | 63.3 ± 3.3 B,d | 50 ± 0 B,b | |
| 2 | 0 ± 0 B,e | 0 ± 0 B,e | 16.7 ± 3.3 C,c,d | 0 ± 0 C,e | 10 ± 0 C,d | 20 ± 0 C,b | 0 ± 0 B,e | 0 ± 0 C,e | 50 ± 0 B,a | |
| 3 | 0 ± 0 B,b | 0 ± 0 B,b | 0 ± 0 D,b | 0 ± 0 C,b | 0 ± 0 D,b | 0 ± 0 D,b | 0 ± 0 B,b | 0 ± 0 C,b | 30 ± 0 C,a | |
| 4 | 0 ± 0 B,b | 0 ± 0 B,b | 0 ± 0 D,b | 0 ± 0 C,b | 0 ± 0 D,b | 0 ± 0 D,b | 0 ± 0 B,b | 0 ± 0 C,b | 30 ± 0 C,a | |
| 5 | 0 ± 0 B,a | 0 ± 0 B,a | 0 ± 0 D,a | 0 ± 0 C,a | 0 ± 0 D,a | 0 ± 0 D,a | 0 ± 0 B,a | 0 ± 0 C,a | 20 ± 0 D,a | |
| Lepidium sativum L. | 0 | 100 ± 0 A,a | 100 ± 0 A,a | 100 ± 0 A,a | 100 ± 0 A,a | 100 ± 0 A,a | 100 ± 0 A,a | 100 ± 0 A,a | 100 ± 0 A,a | 100 ± 0 A,a |
| 1 | 30 ± 0 B,e | 90 ± 0 B,a,b | 90 ± 0 B,a,b | 70 ± 0 A,d | 93.3 ± 3.3 B,a | 83.3 ± 3.3 B,b,c | 93.3 ± 3.3 A,a | 80 ± 0 B,c | 90 ± 0 B,a,b | |
| 2 | 16.7 ± 3.3 C,f | 86.7 ± 3.3 B,a | 80 ± 0 C,a,b | 30 ± 0 A,e | 80 ± 0 C,a,b | 70 ± 0 C,d | 73.3 ± 3.3 B,b,c | 60 ± 0 C,d | 76.7 ± 3.3 C,b,c | |
| 3 | 0 ± 0 D,f | 66.7 ± 3.3 C,a,b | 73.3 ± 3.3 D,a | 10 ± 0 A,e | 73.3 ± 3.3 D,a | 26.7 ± 3.3 D,d | 60 ± 0 C,b | 50 ± 0 D,c | 60 ± 0 D,b | |
| 4 | 0 ± 0 D,d | 46.7 ± 3.3 D,b | 60 ± 0 E,a | 0 ± 0 A,d | 20 ± 0 E,a | 0 ± 0 E,d | 16.7 ± 3.3 D,c | 46.7 ± 3.3 D,b | 53.3 ± 3.3 D,b | |
| 5 | 0 ± 0 D,c | 33.3 ± 3.3 E,b | 0 ± 0 F,c | 0 ± 0 A,c | 0 ± 0 E,c | 0 ± 0 E,c | 0 ± 0 E,c | 30 ± 0 E,b | 43.3 ± 3.3 E,a | |
| Raphanus sativus L. | 0 | 83.3 ± 3.3 A,a | 83.3 ± 3.3 A,a | 83.3 ± 3.3 A,a | 83.3 ± 3.3 A,a | 83.3 ± 3.3 A,a | 83.3 ± 3.3 A,a | 83.3 ± 3.3 A,a | 83.3 ± 3.3 A,a | 83.3 ± 3.3 A,a |
| 1 | 0 ± 0 B,f | 30 ± 0 B,d | 36.7 ± 3.3 B,c,d | 30 ± 0 B,d | 26.7 ± 3.3 B,d | 50 ± 0 B,b | 20 ± 0 B,e | 40 ± 0 B,c | 60 ± 0 B,a | |
| 2 | 0 ± 0 B,f | 23.3 ± 3.3 C,c,d | 30 ± 0 B,b,c | 20 ± 0 C,d | 20 ± 0 C,d | 33.3 ± 3.3 C,b | 10 ± 0 C,e | 33.3 ± 3.3 B,b | 60 ± 0 B,a | |
| 3 | 0 ± 0 B,e | 0 ± 0 D,e | 16.7 ± 3.3 C,c | 0 ± 0 D,e | 10 ± 0 D,d | 20 ± 0 D,b,c | 0 ± 0 D,e | 23.3 ± 3.3 C,b | 60 ± 0 B,a | |
| 4 | 0 ± 0 B,b | 0 ± 0 D,b | 10 ± 0 C,b | 0 ± 0 D,b | 0 ± 0 E,b | 0 ± 0 E,b | 0 ± 0 D,b | 20 ± 0 C,b | 60 ± 0 B,a | |
| 5 | 0 ± 0 B,c | 0 ± 0 D,c | 0 ± 0 D,c | 0 ± 0 D,c | 0 ± 0 E,c | 0 ± 0 E,c | 0 ± 0 D,c | 13.3 ± 3.3 C,b | 50 ± 0 C,a | |
| Triticum durum Desf. | 0 | 90 ± 0 A,a | 90 ± 0 A,a | 90 ± 0 A,a | 90 ± 0 A,a | 90 ± 0 A,a | 90 ± 0 A,a | 90 ± 0 A,a | 90 ± 0 A,a | 90 ± 0 A,a |
| 1 | 60 ± 0 B,c | 76.7 ± 3.3 B,b | 60 ± 0 B,c | 33.3 ± 3.3 B,d | 80 ± 0 B,a,b | 30 ± 0 A,d | 20 ± 0 B,e | 60 ± 0 B,c | 60 ± 0 B,c | |
| 2 | 53.3 ± 3.3 C,c | 50 ± 0 C,c | 40 ± 0 C,d | 0 ± 0 C,f | 70 ± 0 C,b | 10 ± 0 A,e | 10 ± 0 C,e | 50 ± 0 C,c | 53.3 ± 3.3 C,c | |
| 3 | 50 ± 0 C,b | 40 ± 0 D,c | 30 ± 0 D,d | 0 ± 0 C,e | 56.7 ± 3.3 D,a | 0 ± 0 A,e | 0 ± 0 D,e | 33.3 ± 3.3 D,d | 50 ± 0 C,b | |
| 4 | 43.3 ± 3.3 D,a | 33.3 ± 3.3 E,b | 10 ± 0 E,d | 0 ± 0 C,e | 0 ± 0 E,e | 0 ± 0 A,e | 0 ± 0 D,e | 23.3 ± 3.3 E,c | 43.3 ± 3.3 D,a | |
| 5 | 40 ± 0 D,a | 0 ± 0 F,b | 0 ± 0 F,b | 0 ± 0 C,b | 0 ± 0 E,b | 0 ± 0 A,b | 0 ± 0 D,b | 0 ± 0 F,b | 40 ± 0 D,a |
| Tested Plants | Doses (μL/mL) | E. cladocalyx | E. microcorys | E. resinifera | E. saligna | E. angulosa | E. diversicolor | E. ovata | E. sargentii | Glyphosate |
|---|---|---|---|---|---|---|---|---|---|---|
| Trifolium campestre Schreb. | 0 | 5.17 ± 0.04 A,a | 5.17 ± 0.04 A,a | 5.17 ± 0.04 A,a | 5.17 ± 0.04 A,a | 5.17 ± 0.04 A,a | 5.17 ± 0.04 A,a | 5.17 ± 0.04 A,a | 5.17 ± 0.04 A,a | 5.17 ± 0.04 A,a |
| 1 | 1.37 ± 0.02 B,f | 1.45 ± 0.01 B,f | 3.73 ± 0.04 B,b | 2.25 ± 0.08 B,d | 2.86 ± 0.02 B,c | 4.72 ± 0.09 B,a | 2.87 ± 0.07 B,c | 3.61 ± 0.03 B,b | 0.64 ± 0.01 B,g | |
| 2 | 0.83 ± 0.02 C,g | 0.25 ± 0.06 C,j | 2.86 ± 0.04 C,b | 0.42 ± 0.02 C,i | 1.50 ± 0.02 C,f | 3.26 ± 0.05 C,a | 1.93 ± 0 C,d | 2.09 ± 0.02 C,c | 0.59 ± 0.04 B,h | |
| 3 | 0 ± 0 D,h | 0.17 ± 0.03 C,g | 0.70 ± 0.03 D,e | 0.2 ± 0 D,g | 1.02 ± 0 D,d | 2.31 ± 0.03 D,a | 0.64 ± 0.01 D,e,f | 1.51 ± 0.05 D,c | 0.6 ± 0.01 B,f | |
| 4 | 0 ± 0 D,e | 0 ± 0 D,e | 0 ± 0 E,e | 0 ± 0 E,e | 0 ± 0 E,e | 1.58 ± 0.03 E,a | 0 ± 0 E,e | 0.89 ± 0.04 E,c | 0.47 ± 0.01 C,d | |
| 5 | 0 ± 0 D,d | 0 ± 0 D,d | 0 ± 0 E,d | 0 ± 0 E,d | 0 ± 0 E,d | 0.26 ± 0.02 F,c | 0 ± 0 E,d | 0.29 ± 0.01 F,c | 0.41 ± 0.01 C,b | |
| Lolium rigidum Gaudich. | 0 | 4.66 ± 0.05 A,a | 4.66 ± 0.05 A,a | 4.66 ± 0.05 A,a | 4.66 ± 0.05 A,a | 4.66 ± 0.05 A,a | 4.66 ± 0.05 A,a | 4.66 ± 0.05 A,a | 4.66 ± 0.05 A,a | 4.66 ± 0.05 A,a |
| 1 | 2.87 ± 0.09 B,c | 2.48 ± 0.10 B,d | 1.86 ± 0.02 B,e | 1.67 ± 0.05 B,e | 2.63 ± 0.05 B,d | 3.98 ± 0.09 B,a | 3.38 ± 0.02 B,b | 4.16 ± 0.06 B,a | 0 ± 0 B,f | |
| 2 | 1.07 ± 0.04 C,d,e | 2.32 ± 0.06 B,a | 0.71 ± 0.02 C,f | 0 ± 0 C,g | 0.87 ± 0.04 C,e,f | 1.17 ± 0.01 C,d | 1.96 ± 0.03 C,b | 1.43 ± 0.08 C,c | 0 ± 0 B,g | |
| 3 | 0 ± 0 D,d | 0 ± 0 C,d | 0.51 ± 0.02 D,c | 0 ± 0 C,d | 0 ± 0 D,d | 0.66 ± 0.02 D,b | 0.53 ± 0.03 D,c | 0.55 ± 0.06 D,c | 0 ± 0 B,d | |
| 4 | 0 ± 0 D,d | 0 ± 0 C,d | 0.23 ± 0.03 E,c | 0 ± 0 C,d | 0 ± 0 D,d | 0 ± 0 E,d | 0 ± 0 E,d | 0.3 ± 0.05 E,b | 0 ± 0 B,d | |
| 5 | 0 ± 0 D,b | 0 ± 0 C,b | 0 ± 0 F,b | 0 ± 0 C,b | 0 ± 0 D,b | 0 ± 0 E,b | 0 ± 0 E,b | 0 ± 0 F,b | 0 ± 0 B,b | |
| Sinapis arvensis L. | 0 | 3.74 ± 0.08 A,a | 3.74 ± 0.08 A,a | 3.74 ± 0.08 A,a | 3.74 ± 0.08 A,a | 3.74 ± 0.08 A,a | 3.74 ± 0.08 A,a | 3.74 ± 0.08 A,a | 3.74 ± 0.08 A,a | 3.74 ± 0.08 A,a |
| 1 | 0 ± 0 B,g | 0 ± 0 B,g | 2.66 ± 0.08 B,b | 1.70 ± 0.05 B,c | 3.48 ± 0.20 A,a | 1.25 ± 0.03 B,d | 0 ± 0 B,g | 1.74 ± 0.06 B,c | 0.5 ± 0 B,f | |
| 2 | 0 ± 0 B,d | 0 ± 0 B,d | 1.06 ± 0.07 C,b | 0 ± 0 C,d | 1.73 ± 0.14 B,a | 0.65 ± 0.05 C,c | 0 ± 0 B,d | 0 ± 0 C,d | 0.5 ± 0 B,c | |
| 3 | 0 ± 0 B,b | 0 ± 0 B,b | 0.48 ± 0.06 D,a | 0 ± 0 C,b | 0 ± 0 C,b | 0 ± 0 D,b | 0 ± 0 B,b | 0 ± 0 C,b | 0.5 ± 0 B,a | |
| 4 | 0 ± 0 B,b | 0 ± 0 B,b | 0 ± 0 E,b | 0 ± 0 C,b | 0 ± 0 C,b | 0 ± 0 D,b | 0 ± 0 B,b | 0 ± 0 C,b | 0.5 ± 0 B,a | |
| 5 | 0 ± 0 B,b | 0 ± 0 B,b | 0 ± 0 E,b | 0 ± 0 C,b | 0 ± 0 C,b | 0 ± 0 D,b | 0 ± 0 B,b | 0 ± 0 C,b | 0.5 ± 0 B,a | |
| Lepidium sativum L. | 0 | 6.52 ± 0.05 A,a | 6.52 ± 0.05 A,a | 6.52 ± 0.05 A,a | 6.52 ± 0.05 A,a | 6.52 ± 0.05 A,a | 6.52 ± 0.05 A,a | 6.52 ± 0.05 A,a | 6.52 ± 0.05 A,a | 6.52 ± 0.05 A,a |
| 1 | 2.30 ± 0.12 B,f | 2.98 ± 0.06 B,e | 4.24 ± 0.03 B,c | 1.07 ± 0.02 B,h | 4.44 ± 0.06 B,b | 3.40 ± 0.07 B,d | 5.58 ± 0.10 B,a | 2.07 ± 0.02 B,g | 0.72 ± 0.02 B,i | |
| 2 | 0.63 ± 0.04 C,f | 0.79 ± 0.01 C,f | 3.60 ± 0.06 C,b | 0.34 ± 0.01 C,g | 3.64 ± 0.03 C,b | 2.74 ± 0.06 C,d | 2.89 ± 0.05 C,c | 1.61 ± 0.03 C,e | 0.66 ± 0.02 B,f | |
| 3 | 0 ± 0 D,i | 0.34 ± 0.01 D,g | 2.70 ± 0.12 D,c | 0.20 ± 0.00 D,h | 3.18 ± 0.01 D,a | 0.54 ± 0.04 D,f | 1.75 ± 0.07 D,d | 1.41 ± 0.02 D,e | 0.58 ± 0.01 C,f | |
| 4 | 0 ± 0 D,f | 0.22 ± 0.01 E,e | 1.43 ± 0.07 E,a | 0 ± 0 E,f | 0 ± 0 E,f | 0 ± 0 E,f | 0.85 ± 0.03 E,c | 0.96 ± 0.02 E,b | 0.42 ± 0.01 D,d | |
| 5 | 0 ± 0 D,e | 0.12 ± 0.01 F,d | 0 ± 0 F,e | 0 ± 0 E,e | 0 ± 0 E,e | 0 ± 0 E,e | 0 ± 0 F,e | 0.75 ± 0.02 F,b | 0.35 ± 0.04 D,c | |
| Raphanus sativus L. | 0 | 8.67 ± 0.11 A,a | 8.67 ± 0.11 A,a | 8.67 ± 0.11 A,a | 8.67 ± 0.11 A,a | 8.67 ± 0.11 A,a | 8.67 ± 0.11 A,a | 8.67 ± 0.11 A,a | 8.67 ± 0.11 A,a | 8.67 ± 0.11 A,a |
| 1 | 0 ± 0 B,i | 4.43 ± 0.04 B,b | 0 ± 0 B,i | 1.07 ± 0.03 B,g | 5.50 ± 0 B,a | 1.43 ± 0.04 B,f | 3.58 ± 0.08 B,d | 1.89 ± 0.05 B,e | 0.5 ± 0 B,h | |
| 2 | 0 ± 0 B,g | 2.75 ± 0.06 C,a | 0 ± 0 B,g | 0.20 ± 0.00 C,f | 2.60 ± 0.08 C,b | 1.12 ± 0.04 C,c | 0.50 ± 0 C,e | 0.73 ± 0.03 C,d | 0.5 ± 0 B,e | |
| 3 | 0 ± 0 B,c | 0 ± 0 D,c | 0 ± 0 B,c | 0 ± 0 D,c | 1.03 ± 0.03 D,a | 0.47 ± 0.04 D,b | 0 ± 0 D,c | 0.52 ± 0.02 D,b | 0.5 ± 0 B,b | |
| 4 | 0 ± 0 B,c | 0 ± 0 D,c | 0 ± 0 B,c | 0 ± 0 D,c | 0 ± 0 E,c | 0 ± 0 E,c | 0 ± 0 D,c | 0.28 ± 0.02 E,b | 0.5 ± 0 B,a | |
| 5 | 0 ± 0 B,c | 0 ± 0 D,c | 0 ± 0 B,c | 0 ± 0 D,c | 0 ± 0 E,c | 0 ± 0 E,c | 0 ± 0 D,c | 0.13 ± 0.02 E,b | 0.5 ± 0 B,a | |
| Triticum durum Desf. | 0 | 15.42 ± 0.04 A,a | 15.42 ± 0.04 A,a | 15.42 ± 0.04 A,a | 15.42 ± 0.04 A,a | 15.42 ± 0.04 A,a | 15.42 ± 0.04 A,a | 15.42 ± 0.04 A,a | 15.42 ± 0.04 A,a | 15.42 ± 0.04 A,a |
| 1 | 0.50 ± 0 B,g | 9.80 ± 0.15 B,b | 10.89 ± 0.08 B,a | 7.83 ± 0.60 B,c | 6.71 ± 0.17 B,d | 3.29 ± 0.01 B,f | 5.63 ± 0.07 B,e | 11.37 ± 0.16 B,a | 0.35 ± 0 B,g | |
| 2 | 0 ± 0 C,h | 5.63 ± 0.12 C,c | 7.07 ± 0.02 C,a | 4.83 ± 0.17 Cd,e | 4.70 ± 0.10 C,e | 1.00 ± 0 C,g | 2.90 ± 0.06 C,f | 5.01 ± 0.04 C,d | 0.20 ± 0 C,h | |
| 3 | 0 ± 0 C,g | 2.98 ± 0.05 D,c | 5.58 ± 0.06 D,a | 2 ± 0 D,e | 2.30 ± 0.08 D,d | 0 ± 0 D,g | 0 ± 0 D,g | 3.86 ± 0.05 D,b | 0.20 ± 0 C,f | |
| 4 | 0 ± 0 C,f | 1.87 ± 0.02 E,b | 2.00 ± 0 E,a | 0 ± 0 E,f | 0 ± 0 E,f | 0 ± 0 D,f | 0 ± 0 D,f | 0.88 ± 0.01 E,d | 0.20 ± 0 C,e | |
| 5 | 0 ± 0 C,b | 0 ± 0 F,b | 0 ± 0 F,b | 0 ± 0 E,b | 0 ± 0 E,b | 0 ± 0 D,b | 0 ± 0 D,b | 0 ± 0 F,b | 0.20 ± 0 C,a |
| Tested Plants | Doses (μL/mL) | Eucalyptus cladocalyx | Eucalyptus microcorys | Eucalyptus resinifera | Eucalyptus saligna | Eucalyptus angulosa | Eucalyptus diversicolor | Eucalyptus ovata | Eucalyptus sargentii | Glyphosate |
|---|---|---|---|---|---|---|---|---|---|---|
| Trifolium campestre Schreb. | 0 | 1.97 ± 0.01 A,a | 1.97 ± 0.01 A,a | 1.97 ± 0.01 A,a | 1.97 ± 0.01 A,a | 1.97 ± 0.01 A,a | 1.97 ± 0.01 A,a | 1.97 ± 0.01 A,a | 1.97 ± 0.01 A,a | 1.97 ± 0.01 A,a |
| 1 | 1.01 ± 0.01 B,c | 0.54 ± 0.02 B,g | 1.37 ± 0.01 B,a | 0.70 ± 0.01 B,f | 1.02 ± 0.01 B,c | 0.88 ± 0.02 B,d | 0.82 ± 0.01 B,e | 0.78 ± 0.02 B,e | 0.23 ± 0.01 B,h | |
| 2 | 0.72 ± 0.03 C,b | 0.22 ± 0 C,f | 1.06 ± 0.03 C,a | 0.50 ± 0 C,e | 0.65 ± 0.02 C,c | 0.60 ± 0.02 C,c,d | 0.54 ± 0.01 C,d,e | 0.55 ± 0.01 C,d,e | 0.18 ± 0.01 C,f | |
| 3 | 0 ± 0 D,f | 0.10 ± 0 D,e | 0.45 ± 0.01 D,b | 0.32 ± 0.02 D,c | 0.37 ± 0.00 D,c | 0.49 ± 0 D,b | 0.31 ± 0.01 D,c | 0.47 ± 0.01 D,b | 0.18 ± 0.01 C,d | |
| 4 | 0 ± 0 D,d | 0 ± 0 E,d | 0 ± 0 E,d | 0 ± 0 E,d | 0 ± 0 E,d | 0.34 ± 0.02 E,b | 0 ± 0 E,d | 0.35 ± 0.01 E,b | 0.15 ± 0.01 C,c | |
| 5 | 0 ± 0 D,c | 0 ± 0 E,c | 0 ± 0 E,c | 0 ± 0 E,b,c | 0 ± 0 E,c | 0.19 ± 0.03 F,b | 0 ± 0 E,c | 0.16 ± 0.02 F,c | 0.15 ± 0.01 C,b | |
| Lolium rigidum Gaudich. | 0 | 4.04 ± 0 A,a | 4.04 ± 0 A,a | 4.04 ± 0 A,a | 4.04 ± 0 A,a | 4.04 ± 0 A,a | 4.04 ± 0 A,a | 4.04 ± 0 A,a | 4.04 ± 0 A,a | 4.04 ± 0 A,a |
| 1 | 2.60 ± 0.09 B,c | 2.21 ± 0.03 B,d | 1.65 ± 0.02 B,e | 1.39 ± 0.08 B,f | 2.47 ± 0.02 B,c,d | 3.54 ± 0.12 B,a | 3.14 ± 0.02 B,b | 3.28 ± 0.10 B,b | 0 ± 0 B,g | |
| 2 | 1.18 ± 0.05 C,e | 1.50 ± 0.03 C,d | 0.47 ± 0.01 C,g | 0 ± 0 C,h | 0.73 ± 0.06 C,f | 2.39 ± 0.08 C,a | 2.08 ± 0.01 C,b | 2.14 ± 0.03 C,b | 0 ± 0 B,h | |
| 3 | 0 ± 0 D,e | 0 ± 0 D,e | 0.29 ± 0.01 D,d | 0 ± 0 C,e | 0 ± 0 D,e | 0.72 ± 0.06 D,c | 0.93 ± 0.03 D,b | 1.10 ± 0.09 D,a | 0 ± 0 B,g | |
| 4 | 0 ± 0 D,b | 0 ± 0 D,b | 0.12 ± 0.02 E,b | 0 ± 0 C,b | 0 ± 0 D,b | 0 ± 0 E,b | 0 ± 0 E,b | 0.57 ± 0.10 E,a | 0 ± 0 B,b | |
| 5 | 0 ± 0 D,b | 0 ± 0 D,b | 0 ± 0 F,b | 0 ± 0 C,b | 0 ± 0 D,b | 0 ± 0 E,b | 0 ± 0 E,b | 0 ± 0 F,b | 0 ± 0 B,b | |
| Sinapis arvensis L. | 0 | 2.77 ± 0.03 A,a | 2.77 ± 0.03 A,a | 2.77 ± 0.03 A,a | 2.77 ± 0.03 A,a | 2.77 ± 0.03 A,a | 2.77 ± 0.03 A,a | 2.77 ± 0.03 A,a | 2.77 ± 0.03 A,a | 2.77 ± 0.03 A,a |
| 1 | 0 ± 0 B,g | 0 ± 0 B,g | 0.92 ± 0.04 B,d | 0.59 ± 0.01 B,e | 1.80 ± 0.02 B,c | 1.89 ± 0.05 B,b | 0 ± 0 B,g | 2.17 ± 0.03 B,a | 0.50 ± 0 B,f | |
| 2 | 0 ± 0 B,d | 0 ± 0 B,d | 0.55 ± 0.03 C,b | 0 ± 0 C,d | 0.87 ± 0.03 C,a | 0.08 ± 0.02 C,d | 0 ± 0 B,d | 0 ± 0 C,d | 0.50 ± 0 B,b | |
| 3 | 0 ± 0 B,b | 0 ± 0 B,b | 0 ± 0 D,b | 0 ± 0 C,b | 0 ± 0 D,b | 0 ± 0 C,b | 0 ± 0 B,b | 0 ± 0 C,b | 0.50 ± 0 B,a | |
| 4 | 0 ± 0 B,b | 0 ± 0 B,b | 0 ± 0 D,b | 0 ± 0 C,b | 0 ± 0 D,b | 0 ± 0 C,b | 0 ± 0 B,b | 0 ± 0 C,b | 0.50 ± 0 B,a | |
| 5 | 0 ± 0 B,b | 0 ± 0 B,b | 0 ± 0 D,b | 0 ± 0 C,b | 0 ± 0 D,b | 0 ± 0 C,b | 0 ± 0 B,b | 0 ± 0 C,b | 0.50 ± 0 B,a | |
| Lepidium sativum L. | 0 | 1.42 ± 0.02 A,a | 1.42 ± 0.02 A,a | 1.42 ± 0.02 A,a | 1.42 ± 0.02 A,a | 1.42 ± 0.02 A,a | 1.42 ± 0.02 A,a | 1.42 ± 0.02 A,a | 1.42 ± 0.02 A,a | 1.42 ± 0.02 A,a |
| 1 | 0.96 ± 0.03 B,a | 0.77 ± 0.04 B,b | 0.74 ± 0.07 B,b | 0.54 ± 0 B,c | 0.68 ± 0.06 B,b | 0.96 ± 0.01 B,a | 0.87 ± 0.01 B,a | 0.90 ± 0.01 B,a | 0.31 ± 0.01 B,d | |
| 2 | 0.67 ± 0.02 C,b | 0.64 ± 0.02 C,b | 0.67 ± 0.04 B,b | 0.31 ± 0.01 C,d | 0.52 ± 0.01 C,c | 0.78 ± 0.03 C,a | 0.51 ± 0.01 C,c | 0.75 ± 0.02 C,a | 0.25 ± 0 C,d | |
| 3 | 0 ± 0 D,e | 0.55 ± 0.02 D,a | 0.55 ± 0.02 C,a | 0.20 ± 0 D,d | 0 ± 0 D,e | 0.49 ± 0.02 D,b | 0.37 ± 0.02 D,c | 0.47 ± 0.01 D,b | 0.21 ± 0.01 D,d | |
| 4 | 0 ± 0 D,d | 0.49 ± 0.01 D,a | 0.52 ± 0.04 C,a | 0 ± 0 E,d | 0 ± 0 D,d | 0 ± 0 E,d | 0.17 ± 0.03 E,c | 0.35 ± 0.02 E,b | 0.15 ± 0 E,c | |
| 5 | 0 ± 0 D,d | 0 ± 0 E,d | 0 ± 0 D,d | 0 ± 0 E,d | 0 ± 0 D,d | 0 ± 0 E,d | 0 ± 0 F,d | 0.25 ± 0.02 F,b | 0.10 ± 0 F,c | |
| Raphanus sativus L. | 0 | 2.56 ± 0.04 A,a | 2.56 ± 0.04 A,a | 2.56 ± 0.04 A,a | 2.56 ± 0.04 A,a | 2.56 ± 0.04 A,a | 2.56 ± 0.04 A,a | 2.56 ± 0.04 A,a | 2.56 ± 0.04 A,a | 2.56 ± 0.04 A,a |
| 1 | 0 ± 0 B,g | 2.43 ± 0.04 B,a | 0 ± 0 B,g | 0.49 ± 0.01 B,f | 1.33 ± 0.03 B,d | 1.19 ± 0.02 B,e | 1.53 ± 0.02 B,c | 2.08 ± 0.04 B,b | 0.50 ± 0 B,f | |
| 2 | 0 ± 0 B,h | 2.09 ± 0.05 C,a | 0 ± 0 B,h | 0.17 ± 0.02 C,g | 0.93 ± 0.02 C,d | 0.62 ± 0.02 C,e | 0.97 ± 0.03 C,d | 1.88 ± 0.05 C,b | 0.50 ± 0 B,f | |
| 3 | 0 ± 0 B,d | 0 ± 0 D,d | 0 ± 0 B,d | 0 ± 0 D,d | 0.37 ± 0.03 D,c | 0.45 ± 0.03 D,b | 0 ± 0 D,d | 1.36 ± 0.05 D,a | 0.50 ± 0 B,b | |
| 4 | 0 ± 0 B,c | 0 ± 0 D,c | 0 ± 0 B,c | 0 ± 0 D,c | 0 ± 0 E,c | 0 ± 0 E,c | 0 ± 0 D,c | 0.23 ± 0.02 E,b | 0.50 ± 0 B,a | |
| 5 | 0 ± 0 B,b | 0 ± 0 D,b | 0 ± 0 B,b | 0 ± 0 D,b | 0 ± 0 E,b | 0 ± 0 E,b | 0 ± 0 D,b | 0 ± 0 F,b | 0.50 ± 0 B,b | |
| Triticum durum Desf. | 0 | 11.47 ± 0.02 A,a | 11.47 ± 0.02 A,a | 11.47 ± 0.02 A,a | 11.47 ± 0.02 A,a | 11.47 ± 0.02 A,a | 11.47 ± 0.02 A,a | 11.47 ± 0.02 A,a | 11.47 ± 0.02 A,a | 11.47 ± 0.02 A,a |
| 1 | 0.50 ± 0 B,h | 7.21 ± 0.22 B,d | 8.86 ± 0.04 B,c | 10.13 ± 0.19 B,a,b | 5.44 ± 0.07 B,f | 9.84 ± 0.10 B,b | 5.90 ± 0.06 B,e | 10.5 ± 0.25 B,a | 0.44 ± 0.01 B,h | |
| 2 | 0 ± 0 C,i | 2.84 ± 0.03 C,g | 5.36 ± 0.03 C,b | 4.90 ± 0.06 C,c | 3.48 ± 0.04 C,e | 7.10 ± 0.06 C,a | 3.97 ± 0.09 C,d | 7.05 ± 0.04 C,a | 0.20 ± 0 C,h | |
| 3 | 0 ± 0 C,g | 0.53 ± 0 D,e | 3.51 ± 0.04 D,b | 0.87 ± 0.09 D,d | 0.62 ± 0.04 D,e | 0 ± 0 D,g | 0 ± 0 D,g | 3.99 ± 0.03 D,a | 0.20 ± 0 C,f | |
| 4 | 0 ± 0 C,f | 0.52 ± 0.04 D,c | 1.50 ± 0 E,a | 0 ± 0 E,f | 0 ± 0 E,f | 0 ± 0 D,f | 0 ± 0 D,f | 0.60 ± 0.03 E,b | 0.20 ± 0 C,e | |
| 5 | 0 ± 0 C,b | 0 ± 0 E,b | 0 ± 0 F,b | 0 ± 0 E,b | 0 ± 0 E,b | 0 ± 0 D,b | 0 ± 0 D,b | 0 ± 0 F,b | 0.20 ± 0 C,a |
| Species | Preserved Specimen Code | Date of Harvest | Harvest Site | Bioclimatic Stage |
|---|---|---|---|---|
| E. ovata Labill. | EOV211 | December 2021 | Arboretum of Zerniza (Region of Sejnene) | Humid lower biocli-matic |
| E. resinifera Sm. | ERE212 | |||
| E. angulosa Schauer | EAN213 | |||
| E. microcorys F.Muell. | EMI214 | |||
| E. diversicolor F.Muell. | EDI227 | September 2022 | Arboretum of Zerniza (Region of Sejnene) | Humid lower bioclimatic |
| E. saligna Sm. | ESA222 | |||
| E. cladocalyx F.Muell. | ECL224 | |||
| E. sargentii Maiden | ESAR218 | January 2021 | Arboretum of Hanya (Governorate of Sousse) | Semiarid bioclimate |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ayed, A.; Polito, F.; Mighri, H.; Souihi, M.; Caputo, L.; Hamrouni, L.; Amri, I.; Nazzaro, F.; De Feo, V.; Hirsch, A.M.; et al. Chemical Composition of Essential Oils from Eight Tunisian Eucalyptus Species and Their Antifungal and Herbicidal Activities. Plants 2023, 12, 3068. https://doi.org/10.3390/plants12173068
Ayed A, Polito F, Mighri H, Souihi M, Caputo L, Hamrouni L, Amri I, Nazzaro F, De Feo V, Hirsch AM, et al. Chemical Composition of Essential Oils from Eight Tunisian Eucalyptus Species and Their Antifungal and Herbicidal Activities. Plants. 2023; 12(17):3068. https://doi.org/10.3390/plants12173068
Chicago/Turabian StyleAyed, Amira, Flavio Polito, Hedi Mighri, Mouna Souihi, Lucia Caputo, Lamia Hamrouni, Ismail Amri, Filomena Nazzaro, Vincenzo De Feo, Ann M. Hirsch, and et al. 2023. "Chemical Composition of Essential Oils from Eight Tunisian Eucalyptus Species and Their Antifungal and Herbicidal Activities" Plants 12, no. 17: 3068. https://doi.org/10.3390/plants12173068
APA StyleAyed, A., Polito, F., Mighri, H., Souihi, M., Caputo, L., Hamrouni, L., Amri, I., Nazzaro, F., De Feo, V., Hirsch, A. M., & Mabrouk, Y. (2023). Chemical Composition of Essential Oils from Eight Tunisian Eucalyptus Species and Their Antifungal and Herbicidal Activities. Plants, 12(17), 3068. https://doi.org/10.3390/plants12173068