
Carbon Storage in Different Compartments in Eucalyptus Stands and Native Cerrado Vegetation

 ,
,  , , ,
, , ,  , , and
, , and
Abstract

1. Introduction
2. Materials and Methods
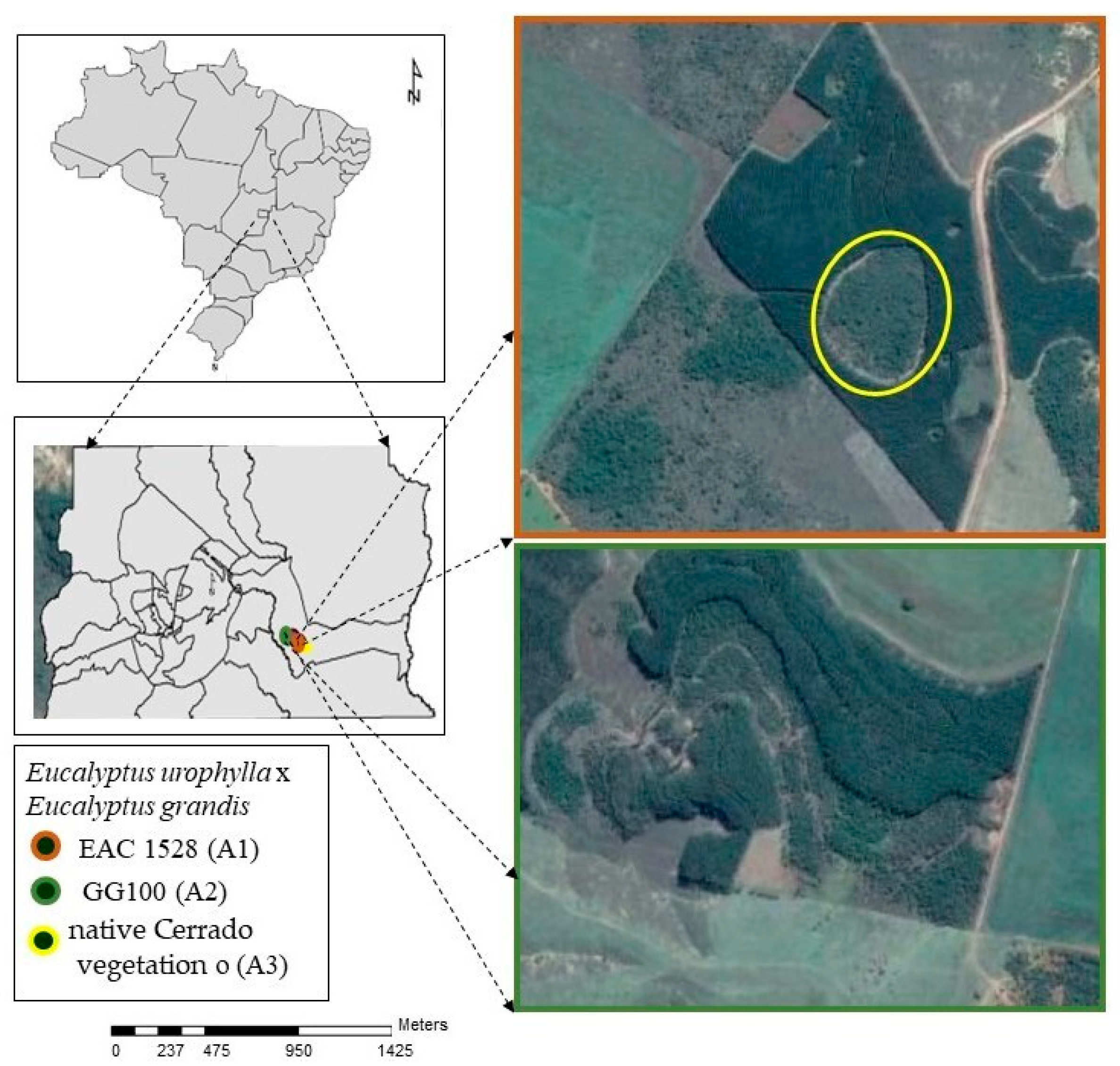
2.1. Study Area
2.2. Data Collection and Analysis
2.2.1. Aboveground Biomass of Native Cerrado Vegetation
2.2.2. Aboveground Biomass in Stands
2.2.3. Root Biomass
2.2.4. Soil Collection and Analysis
2.2.5. Measurements of CH4 and N2O Fluxes
2.2.6. Global Warming Potential and Equivalent Carbon
2.3. Statistical Analyses
3. Results and Discussion
3.1. Aboveground and Belowground Tree Biomass and Carbon Pools
3.1.1. C Stock in the Cerrado Vegetation
3.1.2. Carbon Stock of Eucalyptus Stands
3.2. Soil Carbon
3.3. Carbon Equivalent (C eq) and Global Warming Potential
3.4. Carbon Storage
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- IPCC. Climate Change Impacts, Adaptation, and Vulnerability; Contribution of Working Group II to the Sixth Assessment Report of the Intergovernmental Panel on Climate Change; Pörtner, H.-O., Roberts, D.C., Tignor, M., Poloczanska, E.S., Mintenbeck, K., Alegría, A., Craig, M., Langsdorf, S., Löschke, S., Möller, V., et al., Eds.; Cambridge University Press: Cambridge, UK; New York, NY, USA, 2022; 3056p. [Google Scholar] [CrossRef]
- Yang, Y.H.; Shi, Y.; Sun, W.J.; Chang, J.F.; Zhu, J.X.; Chen, L.Y.; Wang, X.; Guo, Y.P.; Zhang, H.T.; Yu, L.F.; et al. Terrestrial carbon sinks in China and around the world and their contribution to carbon neutrality. Sci. China Life Sci. 2022, 65, 861–895. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.C.; Wang, C.K.; Bond-Lamberty, B. Quantifying and reducing the differences in forest CO2-fluxes estimated by eddy covariance, biometric and chamber methods: A global synthesis. Agric. For. Meteorol. 2017, 247, 93–103. [Google Scholar] [CrossRef]
- van der Werf, G.R.; Randerson, J.T.; Giglio, L.; Collatz, G.J.; Mu, M.; Kasibhatla, P.S.; Morton, D.C.; DeFries, R.S.; Jin, Y.; van Leeuwen, T.T. Global fire emissions and the contribution of deforestation, savanna, forest, agricultural, and peat fires (1997–2009). Atmos. Chem. Phys. 2010, 10, 16153–16230. [Google Scholar] [CrossRef]
- Liu, Y.Y.; Van Dijk, A.I.J.M.; De Jeu, R.A.M.; Canadell, J.G.; Mccabe, M.F.; Evans, J.P.; Wang, G. Recent reversal in loss of global terrestrial biomass. Nat. Clim. Chang. 2015, 5, 470–474. [Google Scholar] [CrossRef]
- Xu, L.; Saatchi, S.S.; Yang, Y.; Yu, Y.; Pongratz, J.; Bloom, A.A.; Bowman, K.; Worden, J.; Liu, J.; Yin, Y.; et al. Changes in global terrestrial live biomass over the 21st century. Sci. Adv. 2021, 7, eabe9829. [Google Scholar] [CrossRef]
- Garrett, R.D.; Grabs, J.; Cammelli, F.; Gollnow, F.; Levy, S.A. Should payments for environmental services be used to implement zero-deforestation supply chain policies? The case of soy in the Brazilian Cerrado. World Dev. 2022, 152, 105814. [Google Scholar] [CrossRef]
- Castaldi, S.; Bertolini, T.; Valente, A.; Chiti, T.; Valentini, R. Nitrous oxide emissions from soil of an African rain forest in Ghana. Biogeosciences 2013, 10, 4179–4187. [Google Scholar] [CrossRef]
- Martins, C.S.C.; Nazaries, L.; Macdonald, C.A.; Anderson, I.C.; Singh, B.K. Water availability and abundance of microbial groups are key determinants of greenhouse gas fluxes in a dryland forest ecosystem. Soil Biol. Biochem. 2015, 86, 5–16. [Google Scholar] [CrossRef]
- De Oliveira, A.D.; Ribeiro, F.P.; Ferreira, E.A.B.; Malaquias, J.V.; Gatto, A.; Zuim, D.R.; Pinheiro, L.D.A.; Pulrolnik, K.; Soares, J.P.G.; De Carvalho, A.M. CH4 and N2O fluxes from planted forests and native Cerrado ecosystems in Brazil. Sci. Agric. 2021, 78, e20180355. [Google Scholar] [CrossRef]
- UNFCCC. Nationally Determined Contributions (NDCs): The Paris Agreement and NDCs. 2015. Available online: https://unfccc.int/sites/default/files/english_paris_agreement.pdf (accessed on 20 June 2023).
- Brasil. Contribuição Nacionalmente Determinada para Consecução do Objetivo da Convenção-Quadro das Nações Unidas sobre Mudança do Clima. 2015. Brasília, DF. Available online: https://unfccc.int/sites/default/files/NDC/2022-06/Updated%20-%20First%20NDC%20-%20%20FINAL%20-%20PDF.pdf (accessed on 20 June 2023).
- IPCC. Guidelines for National Greenhouse Gas Inventories, Prepared by the National Greenhouse Gas Inventories Program. Eggleston. H.S., Buendia, L., Miwa, K., Ngara, T., Tanabe, K., Eds.; IGES: Japan; v 5, Chapter 6. 2006. Available online: https://www.ipcc-nggip.iges.or.jp/public/2006gl/ (accessed on 10 August 2022).
- Strassburg, B.B.N.; Brooks, T.; Feltran-Barbieri, R.; Iribarrem, A.; Crouzeilles, R.; Loyola, R.; Latawiec, A.E.; Filho, F.J.B.O.; Scaramuzza, C.A.D.M.; Scarano, F.R.; et al. Moment of truth for the Cerrado hotspot. Nat. Ecol. Evol. 2017, 1, 99. [Google Scholar] [CrossRef]
- MapBiomas. The MapBiomas Project Provides Annual Maps of Land Cover and Land Use, as Well as Maps with Changes over Time for the Entire Period between 1985–2021. 2022. Available online: https://mapbiomas-br-site.s3.amazonaws.com/Infograficos/MBI-Infografico-7.0-BR-cerrado-rev2.jpg (accessed on 15 April 2023).
- Schüler, J.; Bustamante, M.M.C. Spatial planning for restoration in Cerrado: Balancing the trade-offs between conservation and agriculture. J. Appl. Ecol. 2022, 59, 2616–2626. [Google Scholar] [CrossRef]
- Brasil. Estimativas Anuais de Emissões de Efeito Estufa no Brasil. 6ª edição. 2022. Available online: https://www.gov.br/mcti/pt-br/centrais-de-conteudo/publicacoes-mcti/estimativas-anuais-de-emissoes-de-gases-de-efeito-estufa-no-brasil/estimativas-anuais-de-emissoes-de-gases-de-efeito-estufa-no-brasil.pdf (accessed on 20 June 2023).
- INPE. Incrementos de Desmatamento Acumulado—Cerrado—Estados. 2022. Available online: http://terrabrasilis.dpi.inpe.br/app/dashboard/deforestation/biomes/cerrado/increments (accessed on 23 June 2023).
- de Carvalho, A.M.; de Jesus, D.R.; de Sousa, T.R.; Ramos, M.L.G.; de Figueiredo, C.C.; de Oliveira, A.D.; Marchão, R.L.; Ribeiro, F.; Dantas, R.; de Araújo Barbosa Borges, L. Soil Carbon Stocks and Greenhouse Gas Mitigation of Agriculture in the Brazilian Cerrado—A Review. Plants 2023, 12, 2449. [Google Scholar] [CrossRef] [PubMed]
- Guerino, R.M.G.; Morais, I.L.; Santos, A.B.S.; Campos, R.M. Expansão e impactos socioambientais da cultura de Eucalyptus spp. (Myrtaceae) no Brasil: Um panorama da literature. Res. Soc. Dev. 2022, 11, 3. [Google Scholar] [CrossRef]
- Zinn, Y.L.; Resck, D.V.; da Silva, J.E. Soil Organic Carbon as Affected by Afforestation with Eucalyptus and Pinus in the Cerrado Region of Brazil. For. Ecol. Manag. 2002, 166, 285–294. [Google Scholar] [CrossRef]
- Pulrolnik, K.; de Barros, N.F.; Silva, I.R.; Novais, R.F.; Brandani, C.B. Estoques de carbono e nitrogênio em frações lábeis e estáveis da matéria orgânica de solos sob eucalipto, pastagem e Cerrado no vale do Jequitinhonha—MG. Rev. Bras. Ciência Solo 2009, 33, 1125–1136. [Google Scholar] [CrossRef]
- Rossi, C.Q.; Pereira, M.G.; Giácomo, S.G.; Betta, M.; Polidoro, J.C. Organic Fractions and Soil Organic Carbon Management Index in an Oxisol under Soybean Cultivation in the Cerrado Region of Goiás, Brazil. Rev. Bras. Cienc. Agrar. 2012, 7, 233–241. [Google Scholar] [CrossRef]
- Leite, L.F.C.; de Arruda, F.P.; Costa, C.D.N.; Ferreira, J.D.S.; Holanda Neto, M.R. Chemical Quality of Soil and Dynamics of Carbon under Monoculture and Intercroping of Acrocomia Palm and Pasture. Rev. Bras. Eng. Agrícola Ambient. 2013, 17, 1257–1263. [Google Scholar] [CrossRef]
- Gatto, A.; Bussinger, A.P.; Ribeiro, F.C.; Azevedo, G.B.; Bueno, M.C.; Monteiro, M.M.; Souza, P.F. Ciclagem e balanço de nutrientes no sistema solo-planta em um plantio de Eucalyptus sp. no Distrito Federal. Rev. Bras.Ciência Solo 2014, 38, 879–887. [Google Scholar] [CrossRef]
- Rodrigues, G.G.; Zanatta, J.A.; Rachwal, M.F.G.; Higa, R.C.V.; Lavoranti, O.; Silva, L.D. Methane and nitrous oxide fluxes from clonal eucalyptus forests in different spacing arrangements. For. Int. J. For. Res. 2023, 96, 1–12. [Google Scholar] [CrossRef]
- Indústria Brasileira de Árvores—IBÁ. Relatório Ibá 2022, 96p. Available online: https://www.iba.org/datafiles/publicacoes/relatorios/relatorio-anual-iba2022-compactado.pdf (accessed on 10 February 2023).
- Morais, V.A.; de Mello, J.M.; de Mello, C.R.; Silva, C.A.; Scolforo, J.R.S. Spatial distribution of the litter carbon stock in the Cerrado biome in Minas Gerais state, Brazil. Ciência Agrotecnologia 2017, 41, 580–589. [Google Scholar] [CrossRef]
- Morais, V.A.; Santos, C.A.; Mello, J.M.; Dadid, H.C.; Araújo, E.J.G.; Scolforo, J.R.S. Spatial and vertical distribution of litter and belowground carbon in a Brazilian Cerrado vegetation. CERNE 2017, 23, 43–52. [Google Scholar] [CrossRef]
- Qi, Y.; Wei, W.; Chen, C.; Chen, L. Plant root-shoot biomass allocation over diverse biomes: A global synthesis. Glob. Ecol. Conserv. 2019, 18, e00606. [Google Scholar] [CrossRef]
- Felfili, J.M.; Carvalho, F.A.; Haidar, R.F. Manual Para o Monitoramento de Parcelas Permanentes nos Biomas Cerrado e Pantanal; Universidade de Brasília: Brasília, Brazil, 2005; 51p. [Google Scholar]
- Rezende, A.V.; Vale, A.T.; Sanquetta, C.R.; Figueiredo Filho, A.; Felfili, J.M. Comparação de modelos matemáticos para estimativa do volume, biomassa e estoque de carbono da vegetação lenhosa de um cerrado sensu stricto em Brasília, DF. Sci. For. 2006, 71, 65–76. Available online: https://www.ipef.br/publicacoes/scientia/nr71/cap07.pdf (accessed on 14 June 2022).
- Teixeira, P.C.; Donagemma, G.K.; Fontana, A.; Teixeira, W.G. Manual de Métodos de Análise de Solo; Embrapa Solos: Brasília, Brazil, 2017; 574p. [Google Scholar]
- Veldkamp, E. Organic Carbon Turnover in Three Tropical Soils under Pasture after Deforestation. Soil Sci. Soc. Am. J. 1994, 58, 175–180. [Google Scholar] [CrossRef]
- Alves, B.J.R.; Smith, K.A.; Flores, R.A.; Cardoso, A.S.; Oliveira, W.R.D.; Jantalia, C.P.; Urquiaga, S.; Boddey, R.M. Selection of the most suitable sampling time for static chambers for the estimation of daily mean N2O flux from soils. Soil Biol. Biochem. 2012, 46, 129–136. [Google Scholar] [CrossRef]
- Bayer, C.; Gomes, J.; Zanatta, J.A.; Vieira, F.C.B.; Piccolo, M.C.; Dieckow, J.; Six, J. Soil nitrous oxide emissions as affected by long-term tillage, cropping systems and nitrogen fertilization in Southern Brazil. Soil Tillage Res. 2015, 146, 213–222. [Google Scholar] [CrossRef]
- Myhre, G.; Shindell, D.; Bréon, F.-M.; Collins, W.; Fuglestvedt, J.; Huang, J.; Koch, D.; Lamarque, J.-F.; Lee, D.; Mendoza, B.; et al. Anthropogenic and natural radiative forcing. In Climate Change: The Physical Science Basis. Contribution of Working Group I to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change; Cambridge University Press: Cambridge, UK; New York, NY, USA, 2013; pp. 659–740. [Google Scholar] [CrossRef]
- Miranda, S.C.; Bustamante, M.; Palácio, M.; Hagen, S.; Keller, M.; Ferreira, L.G. Regional Variations in Biomass Distribution in Brazilian Savanna Woodland. Biotropica 2014, 46, 125–138. [Google Scholar] [CrossRef]
- Silveira, E.M.D.O.; Terra, M.D.C.N.S.; ter Steege, H.; Maeda, E.E.; Júnior, F.W.A.; Scolforo, J.R.S. Carbon-diversity hotspots and their owners in Brazilian southeastern Savanna, Atlantic Forest and Semi-Arid Woodland domains. For. Ecol. Manag. 2019, 452, 117575. [Google Scholar] [CrossRef]
- Ribeiro, S.C.; Jacovine, L.A.G.; Torres, C.M.M.E.; Souza, A.L. Influence of Interspecific Variation on Tree Carbon Stock of a Brazilian Cerrado. Rev. Árvore 2017, 41, e410506. [Google Scholar] [CrossRef][Green Version]
- Santana, R.C.; de Barros, N.F.; Novais, R.F.; Leite, H.G.; Comerford, N.B. Alocação de nutrientes em plantios de eucalipto no Brasil. R. Bras. Ci. Solo 2008, 32, 2723–2733. [Google Scholar] [CrossRef]
- Gatto, A.; Barros, N.F.; Novais, R.F.; Silva, I.R.; Leite, H.G.; Villani, E.M.A. Carbon stock in the biomass of eucalyptus crops in central-east region of the state of Minas Gerais—Brazil. Rev. Árvore 2011, 35, 895–905. [Google Scholar] [CrossRef]
- Zhou, X.; Yuanguang, W.; Uromi, G.G.; Hua, Z.; Hongguang, Z.; Xiao, Q.L.; Yeming, V.; Li, Y.Y.S.; Xueman, H. Optimal rotation length for carbon sequestration in Eucalyptus plantations in subtropical China. New For. 2017, 48, 609–627. [Google Scholar] [CrossRef]
- Kumar, P.; Mishra, A.K.; Chaudhari, S.K.; Sharma, D.K.; Rai, A.K.; Kailash, S.; Rai, P.; Singh, R. Carbon Sequestration and Soil Carbon Build-Up Under Eucalyptus Plantation in Semi-Arid Regions of North-West India. J. Sustain. For. 2020, 40, 319–331. [Google Scholar] [CrossRef]
- Viera, M.; Rodríguez-Soalleiro, R. A Complete Assessment of Carbon Stocks in Above and Belowground Biomass Components of a Hybrid Eucalyptus Plantation in Southern Brazil. Forest 2019, 10, 536. [Google Scholar] [CrossRef]
- Nagar, B.; Rawat, S.; Rathiesh, P.; Sekar, I.I. Impact of initial spacing on growth and yield of Eucalyptus camaldulensis in arid region of India. World Appl. Sci. J. 2015, 33, 1362–1368. [Google Scholar]
- Sausen, T.L.; Schaefer, G.F.P.; Tomazi, M.; Santos, L.S.; Bayer, C.; Rosa, L.M.G. Clay content drives carbon stocks in soils under a plantation of Eucalyptus saligna Labill. in southern Brazil. Acta Bot. Bras. 2014, 28, 266–273. [Google Scholar] [CrossRef]
- Cook, L.R.; Binkley, D.; Stape, J.L. Eucalyptus plantation effects on soil carbon after 20 years and three rotations in Brazil. For. Ecol. Manag. 2016, 359, 92–98. [Google Scholar] [CrossRef]
- Ribeiro, F.P.; Gatto, A.; Oliveira, A.D.; Pulrolnik, K.; Ferreira, E.A.B.; Carvalho, A.M.; Bussinguer, A.P.; Muller, A.G.; Moraes-Neto, S.P. Litter Dynamics in Eucalyptus and Native Forest in the Brazilian Cerrado. J. Agric. Sci. 2018, 10, 29–43. [Google Scholar] [CrossRef]
- Ribeiro, F.P.; Oliveira, A.D.; Bussinguer, A.P.; Rodrigues, M.I.; Cardoso, M.S.S.; Lustosa-Junior, I.; Valadão, M.B.X.; Gatto, A. How long does it take to decompose all litter in Brazilian savanna forest? Cerne 2022, 28, e102819. [Google Scholar] [CrossRef]
- Zinn, Y.L.; Lal, R.; Resck, D.V.S. Eucalypt plantation effects on organic carbon and aggregation of three different-textured soils in Brazil. Soil Res. 2011, 49, 614–624. [Google Scholar] [CrossRef]
- Valadão, M.B.X.; Carneiro, K.M.S.; Inkotte, J.; Ribeiro, F.P.; Miguel, E.P.; Gatto, A. Litterfall, litter layer and leaf decomposition in Eucalyptus stands on Cerrado soils. Sci. For. 2019, 42, 256–264. [Google Scholar] [CrossRef]
- Lira-Martins, D.; Luciano Nascimento, D.; Abrahão, A.; Costa, P.B.; D’Angioli, A.M.; Valézio, E.; Rowland, L.; Oliveira, R.S. Soil properties and geomorphic processes influence vegetation composition, structure, and function in the Cerrado Domain. Plant Soil 2022, 476, 549–588. [Google Scholar] [CrossRef]
- Gomes, S.S.; Gonçalves, J.L.M.; Rocha, J.H.T.; Menegale, M.L.C. Tolerance of Eucalyptus and Pinus seedlings to exchangeable Aluminium. Sci. Agric. 2019, 76, 494–500. [Google Scholar] [CrossRef]
- Cabreira, W.V.; Pereira, M.G.; Balieiro, F.C.; Matos, E.S.; Rodrigues, R.A.R.; Santos, R.N. Mixed-species plantations of Eucalyptus and Acacia mangium increase labile carbon and phosphorus levels in Ferralsol soils in the Cerrado biome, Brazil. South. For. 2020, 82, 342–351. [Google Scholar] [CrossRef]
- McMahon, D.E.; Vergütz, L.; Valadares, S.V.; Silva, I.R.; Jackson, R.B. Soil nutrient stocks are maintained over multiple rotations in Brazilian Eucalyptus plantations. For. Ecol. Manag. 2019, 448, 364–375. [Google Scholar] [CrossRef]
- Dionizio, E.A.; Pimenta, F.M.; Lima, L.B.; Costa, M.H. Carbon stocks and dynamics of different land uses on the Cerrado agricultural frontier. PLoS ONE 2020, 15, e0241637. [Google Scholar] [CrossRef] [PubMed]
- Pinheiro, F.M.; Nair, P.K.R.; Nair, V.D.; Tonucci, R.G.; Venturin, R.P. Soil Carbon Stock and Stability under Eucalyptus-Based Silvopasture and Other Land-Use Systems in the Cerrado Biodiversity Hotspot. J. Environ. Manag. 2021, 299, 113676. [Google Scholar] [CrossRef]
- Ferreira, E.A.B.; Bustamante, M.M.D.C.; Resck, D.V.S.; De Figueiredo, C.C.; Pinto, A.D.S.; Malaquias, J.V. Carbon Stocks in Compartments of Soil Organic Matter 31 Years after Substitution of Native Cerrado Vegetation by Agroecosystems. Rev. Bras. Cienc Solo 2016, 40, e0150059. [Google Scholar] [CrossRef]
- Martins, M.R.; Jantalaia, C.P.; Polidoro, J.C.; Batista, J.N.; Alves, B.J.R.; Boddey, R.M.; Urquiaga, S. Nitrous oxide and ammonia emission from N fertilization of maize crop under no-till in Cerrado soil. Soil Tillage Res. 2015, 151, 75–81. [Google Scholar] [CrossRef]
- Santos, I.L.; Oliveira, A.D.; Figueiredo, C.C.; Malaquias, J.V.; Santos, J.D.G., Jr.; Ferreira, E.A.B.; Sa, M.A.C.; Carvalho, A.M. Soil N2O emissions from long-term agroecosystems: Interactive effects of rainfall seasonality and crop rotation in the Brazilian Cerrado. Agric. Ecosyst. Environ. 2016, 233, 111–120. [Google Scholar] [CrossRef]
- de Carvalho, A.M.; de Oliveira, W.R.D.; Ramos, M.L.G.; Coser, T.R.; de Oliveira, A.D.; Pulrolnik, K.; Souza, K.W.; Vilela, L.; Marchão, R.L. Soil N2O fluxes in integrated production systems, continuous pasture and Cerrado. Nutr. Cycl. Agroecosyst. 2017, 108, 69–83. [Google Scholar] [CrossRef]
- Sato, J.H.; de Carvalho, A.M.; de Figueiredo, C.C.; Coser, T.R.; de Sousa, T.R.; Vilela, L.; Marchão, R.L. Nitrous oxide fluxes in a Brazilian clayey oxisol after 24 years of integrated crop-livestock management. Nutr. Cycl. Agroecosyst. 2017, 108, 55–68. [Google Scholar] [CrossRef]
- Livesley, S.J.; Kiese, R.; Miehle, P.; Weston, C.J.; Butterbach-Bahl, K.; Arndt, S.K. Soil-atmosphere exchange of greenhouse gases in a Eucalyptus marginata woodland, a clover-grass pasture, and Pinus radiata and Eucalyptus globulus plantations. Glob. Chang. Biol. 2009, 15, 425–440. [Google Scholar] [CrossRef]
- Grover, S.P.P.; Livesley, S.J.; Hutley, L.B.; Jamali, H.; Fest, B.; Beringer, J.; Butterbach-Bahl, K.; Arndt, S.K. Land use change and the impact on greenhouse gas exchange in north Australian savanna soils. Biogeosciences 2012, 9, 423–437. [Google Scholar] [CrossRef]
- Luo, G.J.; Kiese, B.; Wolf, B.; Butterbach-Bahl, K. Effects of soil temperature and moisture on methane uptake and nitrous oxide emissions across three different ecosystem types. Biogeosciences 2013, 10, 3205–3219. [Google Scholar] [CrossRef]
- Carvalho, J.L.N.; Raucci, G.S.; Frazao, L.A.; Cerri, C.E.P.; Bernoux, M.; Cerri, C.C. Crop-pasture rotation: A strategy to reduce soil greenhouse gas emissions in the Brazilian Cerrado. Agric. Ecosyst. Environ. 2014, 183, 167–175. [Google Scholar] [CrossRef]
- Bustamante, M.M.C.; Nardoto, G.B.; Pinto, A.S.; Resende, J.C.F.; Takahashi, F.S.C.; Vieira, L.C.G. Potential impacts of climate change on biogeochemical functioning of Cerrado Ecosystems. Braz. J. Biol. 2012, 72, 655–671. [Google Scholar] [CrossRef]
- Zanoni, M.M.V.; Zanatta, J.A.; Dieckow, J.; Kan, A.; Reissmann, C.B. Emissão de metano por decomposição de resíduo florestal inundado. Rev. Bras. Eng. Agríc. Ambiente. 2015, 19, 173–179. [Google Scholar] [CrossRef][Green Version]
- Snyder, C.S.; Bruulsema, T.W.; Jensen, T.L.; Fixen, P.E. Review of greenhouse gas emissions from crop production systems and fertilizer management effects. Agric. Ecosyst. Environ. 2009, 133, 247–266. [Google Scholar] [CrossRef]
- Singh, B.K.; Bardgett, R.D.; Smith, P.; Reay, D.S. Microorganisms and climate change: Terrestrial feedbacks and mitigation options. Nat. Rev. Microbiol. 2010, 9, 79–98. [Google Scholar] [CrossRef]
- Godoi, S.G.; Neufeld, A.D.H.; Ibarr, M.A.; Ferreto, D.O.C.; Bayer, C.; Lorentz, L.H.; Vieira, F.C.B. The conversion of grassland to acacia forest as an effective option for net reduction in greenhouse gas emissions. J. Environ. Manag. 2016, 169, 91–102. [Google Scholar] [CrossRef] [PubMed]
- Miranda, E.; Carmo, J.; Couto, E.; Camargo, P. Long-Term Changes in Soil Carbon Stocks in the Brazilian Cerrado Under Commercial Soybean. Land Degrad. Devel. 2015, 27, 1586–1594. [Google Scholar] [CrossRef]
- Laclau, J.P.; Silva, E.A.; Lambais, G.R.; Bernoux, M.; Maire, G.; Stape, J.L.; Bouillet, J.P.; Gonçalves, J.L.M.; Jourdan, C.; Nouvellon, Y. Dynamics of soil exploration by fine roots down to a depth of 10 m throughout the entire rotation in Eucalyptus grandis plantations. Front. Plant Sci. 2013, 4, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Lima, M.; Vicente, L.C.; Gama-Rodrigues, E.F.; Gama-Rodrigues, A.C.; Lisbôa, F.M.; Aleixo, S. Carbon functional groups of leaf litter in cacao and rubber agroforestry systems in southern Bahia, Brazil. Agroforest Syst. 2023, 97, 249–260. [Google Scholar] [CrossRef]
- Valadão, M.B.X.; Carneiro, K.M.S.; Ribeiro, F.P.; Inkotte, J.; Rodrigues, M.I.; Mendes, T.R.S.; Vieira, D.A.; Matias, R.A.M.; Lima, M.B.O.; Miguel, E.P.; et al. Modeling Biomass and Nutrients in a Eucalyptus Stand in the Cerrado. Forests 2020, 11, 1097. [Google Scholar] [CrossRef]
- Silva, S.C.; Valadão, M.B.X.; Inkotte, J.; Ribeiro, F.P.; Miguel, E.P.; Gatto, A. Legal provision on the use of exotic species in forest restoration: Eucalyptus leaf decomposition in different Brazilian savannah formations. Aust. J. Crop Sci. 2023, 14, 14–19. [Google Scholar]



{kind=link}
{kind=link}
{kind=link}
| Areas | Depth (cm) | Al3+ | Ca | Mg (1) | pH H2O | H + Al | OM (2) | Mn | K | P | Zn | Fe | B | Density |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| ---- cmolc dm−3--- | mg dm−3 | dag kg−1 | ------------------ mg dm−3 ---------------------- | g cm3 | ||||||||||
| A1 | 0–5 | 2.2 | 0.2 | 0.6 | 5.1 | 9.5 | 3.8 | 12.2 | 230.7 | 1.8 | 0.6 | 142.3 | 0.5 | 1.14 |
| A1 | 5–10 | 2.3 | 0.1 | 0.3 | 5.1 | 8.6 | 3.2 | 9.1 | 185.3 | 1.3 | 0.4 | 141.0 | 0.5 | 1.24 |
| A1 | 10–20 | 2.3 | 0.0 | 0.2 | 5.1 | 8.0 | 2.9 | 6.2 | 158.7 | 0.8 | 0.3 | 122.0 | 0.3 | 1.26 |
| A1 | 20–30 | 2.1 | 0.1 | 0.2 | 5.0 | 7.2 | 2.6 | 3.8 | 109.3 | 0.5 | 0.2 | 59.7 | 0.3 | 1.21 |
| A1 | 30–40 | 2.1 | 0.0 | 0.1 | 5.1 | 7.1 | 2.3 | 4.9 | 112.7 | 0.2 | 0.3 | 82.3 | 0.3 | 1.19 |
| A1 | 40–60 | 1.9 | 0.0 | 0.1 | 5.1 | 6.2 | 1.9 | 4.2 | 93.3 | 0.1 | 0.2 | 63.0 | 0.3 | 1.16 |
| A1 | 60–80 | 1.6 | 0.0 | 0.1 | 5.1 | 5.2 | 1.1 | 3.3 | 83.3 | 0.0 | 0.2 | 55.0 | 0.2 | 1.09 |
| A1 | 80–100 | 1.4 | 0.0 | 0.1 | 5.1 | 4.8 | 1.2 | 7.3 | 76.7 | 0.0 | 0.2 | 38.7 | 0.2 | 1.08 |
| A2 | 0–5 | 2.0 | 0.8 | 0.6 | 5.0 | 8.2 | 3.3 | 12.4 | 157.3 | 1.5 | 0.3 | 102.0 | 0.5 | 1.2 |
| A2 | 5–10 | 2.1 | 0.7 | 0.5 | 4.9 | 8.3 | 2.9 | 10.7 | 138.0 | 1.4 | 0.3 | 101.7 | 0.5 | 1.2 |
| A2 | 10–20 | 2.3 | 0.5 | 0.4 | 4.8 | 8.0 | 2.5 | 6.4 | 111.3 | 1.2 | 0.3 | 61.3 | 0.5 | 1.2 |
| A2 | 20–30 | 2.4 | 0.1 | 0.2 | 4.9 | 8.1 | 2.2 | 6.3 | 121.3 | 1.0 | 0.3 | 106.0 | 0.5 | 1.1 |
| A2 | 30–40 | 2.3 | 0.1 | 0.1 | 4.8 | 7.2 | 2.0 | 3.3 | 90.0 | 0.8 | 2.0 | 94.3 | 0.3 | 1.1 |
| A2 | 40–60 | 2.1 | 0.1 | 0.1 | 5.0 | 6.4 | 1.7 | 3.1 | 80.0 | 0.4 | 0.2 | 63.3 | 0.2 | 1.1 |
| A2 | 60–80 | 1.8 | 0.0 | 0.1 | 5.1 | 5.5 | 1.2 | 3.2 | 74.7 | 0.2 | 0.1 | 45.0 | 0.2 | 1.0 |
| A2 | 80–100 | 1.5 | 0.1 | 0.1 | 5.3 | 4.6 | 1.1 | 2.8 | 70.7 | 0.2 | 0.1 | 38.7 | 0.2 | 1.0 |
| A3 | 0–5 | 0.6 | 2.5 | 0.9 | 5.4 | 7.0 | 3.0 | 12.7 | 347.3 | 5.3 | 0.6 | 68.0 | 1.2 | 1.11 |
| A3 | 5–10 | 1.0 | 1.7 | 0.6 | 5.3 | 7.1 | 2.6 | 9.4 | 134.7 | 3.5 | 1.3 | 85.3 | 0.9 | 1.17 |
| A3 | 10–20 | 1.5 | 0.7 | 0.3 | 5.0 | 6.8 | 2.3 | 4.7 | 148.0 | 2.1 | 0.4 | 87.0 | 0.8 | 1.15 |
| A3 | 20–30 | 1.8 | 0.5 | 0.2 | 4.9 | 6.3 | 2.0 | 3.1 | 75.3 | 1.1 | 0.2 | 80.0 | 0.9 | 1.22 |
| A3 | 30–40 | 1.5 | 0.6 | 0.2 | 5.0 | 5.6 | 1.6 | 3.0 | 62.7 | 0.7 | 0.3 | 71.0 | 1.1 | 1.12 |
| A3 | 40–60 | 1.2 | 0.3 | 0.1 | 5.1 | 5.0 | 1.2 | 2.1 | 47.3 | 0.4 | 0.2 | 51.3 | 1.6 | 1.03 |
| A3 | 60–80 | 0.8 | 0.2 | 0.1 | 5.2 | 4.1 | 0.9 | 1.8 | 44.0 | 0.4 | 0.1 | 42.7 | 0.9 | 1.06 |
| A3 | 80–100 | 0.7 | 0.2 | 0.1 | 4.9 | 4.0 | 0.8 | 1.4 | 43.3 | 0.4 | 0.3 | 43.0 | 0.6 | 1.07 |
| Species | Basal Area | Volume | Biomass | Carbon |
|---|---|---|---|---|
| m2 ha−1 | m3 | kg ha−1 | kg ha−1 | |
| Qualea grandiflora | 2.59 | 6.42 | 5758.24 | 2879.12 |
| Miconia pohliana | 3.12 | 7.08 | 5655.73 | 2827.87 |
| Qualea parviflora | 1.05 | 3.29 | 3701.77 | 1850.89 |
| Xylopia aromatica | 1.41 | 3.52 | 3233.56 | 1616.78 |
| Emmotum nitens | 0.70 | 2.13 | 2342.85 | 1171.43 |
| Curatella americana | 0.62 | 1.57 | 1434.87 | 717.43 |
| Amaioua guianensis | 0.51 | 1.41 | 1411.61 | 705.80 |
| Miconia albicans | 0.83 | 1.73 | 1268.01 | 634.00 |
| Terminalia agentea | 0.42 | 1.19 | 1241.02 | 620.51 |
| Astronium fraxinifolium | 0.48 | 1.25 | 1180.94 | 590.47 |
| Eriotheca pubescens | 0.33 | 1.01 | 1098.38 | 549.19 |
| Xylopia brasiliense | 0.37 | 0.89 | 791.30 | 395.65 |
| Alibertia edulis | 0.42 | 0.94 | 777.53 | 388.76 |
| Kielmeyera coriacea | 0.54 | 1.10 | 753.74 | 376.87 |
| Rudgea viburnoides | 0.30 | 0.69 | 578.97 | 289.49 |
| Simarouba versicolor | 0.19 | 0.53 | 541.49 | 270.74 |
| Schefflera macrocarpa | 0.18 | 0.48 | 491.72 | 245.86 |
| Eugenia dysenterica | 0.20 | 0.46 | 378.78 | 189.39 |
| Ouratea grandiflora | 0.14 | 0.35 | 298.20 | 149.10 |
| Virola sebifera | 0.09 | 0.25 | 264.26 | 132.13 |
| Other species in total | 2.41 | 5.37 | 4223.35 | 2111.67 |
| Dead trees | 4.07 | 8.93 | 6749.48 | 3374.74 |
| Vine | 0.03 | 0.06 | 25.79 | 12.89 |
| Total | 21.02 | 50.65 | 44,201.59 | 22,100.78 |
| Compartment | A1 | A2 | ||||
|---|---|---|---|---|---|---|
| Biomass | Carbon | Biomass | Carbon | |||
| ---Mg ha−1 --- | (%) | ---Mg ha−1--- | (%) | |||
| Leaves | 5.3 | 2.4 | 3.76 | 2.3 | 1.0 | 1.21 |
| Branches | 5.3 | 2.4 | 3.76 | 3.4 | 1.5 | 1.79 |
| Wood | 114.8 | 50.3 | 81.36 | 167.8 | 72.5 | 88.46 |
| Bark | 15.6 | 7.1 | 11.06 | 16.2 | 6.8 | 8.54 |
| Aboveground total | 141.1 | 62.1 | 100 | 189.7 | 81.7 | 100 |
| N2O | |||
|---|---|---|---|
| kg ha−1 yr−1 | GWP100 1 | kg CO2 eq ha−1 yr −1 | |
| A1 | 0.43 (±0.23) a | 298 | 127.82 (±77) a |
| A2 | 0.85 (±0.45) a | 298 | 271.30 (±116) a |
| A3 | 0.33 (±0.20) a | 298 | 98.15 (±59) a |
| CH4 | |||
| A1 | −1.85 (±1.36) a | 34 | −63.09 (±46) a |
| A2 | −0.98 (±0.91) a | 34 | −33.31 (±31) a |
| A3 | −0.63 (±0.53) a | 34 | −21.53 (±18) a |
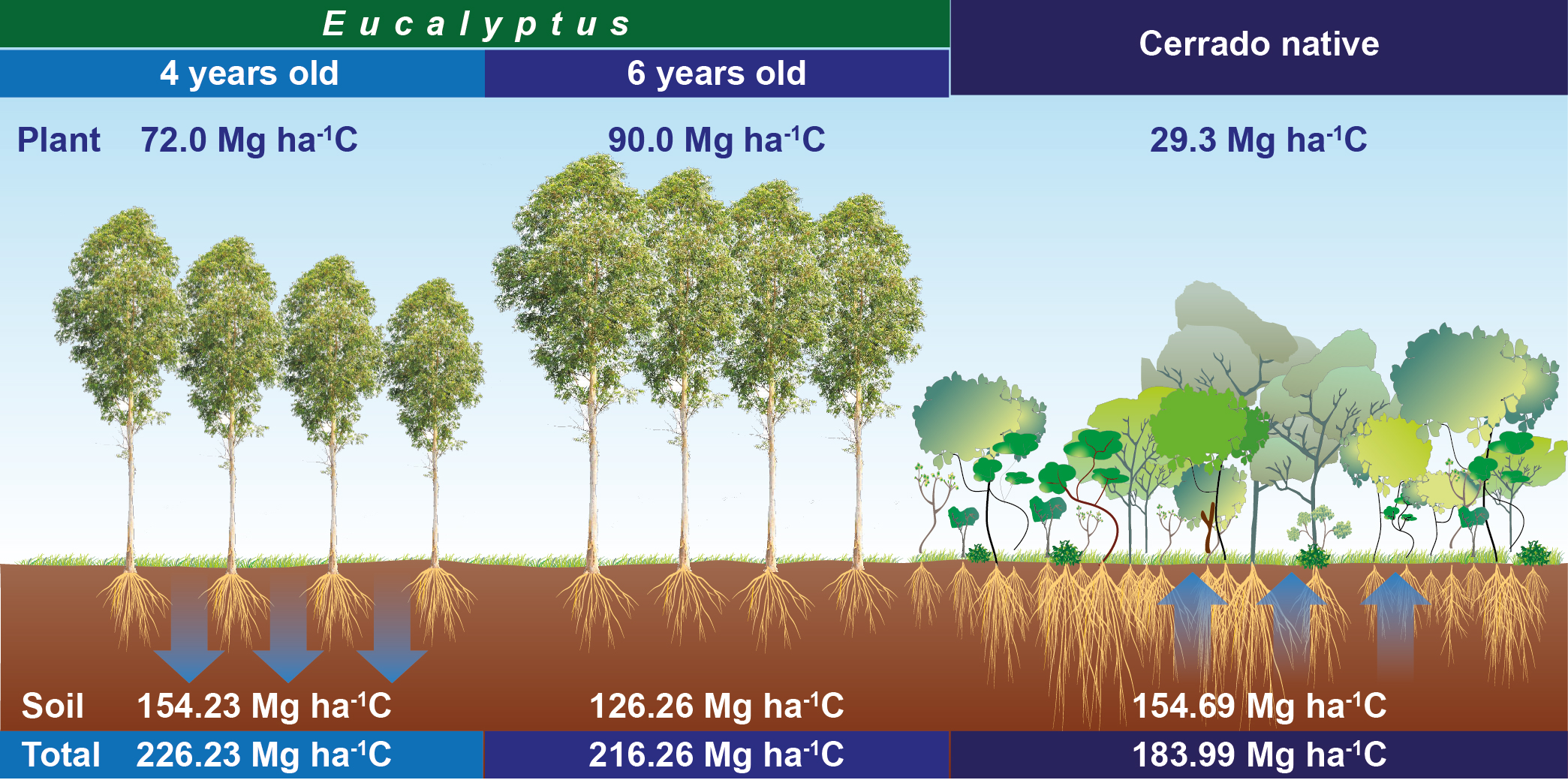
| Compartment | A1 | A2 | A3 |
|---|---|---|---|
| --- Mg ha−1 --- | |||
| Aerial part | (62.1 ± 3.6) b | (81.7 ± 22.2) a | (22.1 ± 0.02) c |
| Roots 0–60 cm | (4.9 ± 2.1) a | (1.9 ± 0.84) b | (3.1 ± 0.95) ab |
| Litter | (5.0 ± 0.23) ab | (6.4 ± 0.2) a | (4.1 ± 0.28) b |
| Plant total | (72 ± 6.0) ab | (90 ± 23.3) a | (29.3 ± 1.3) c |
| Soil (0–100 cm) | (154.23 ± 15.64) a | (126.26 ± 13.35) b | (154.69 ± 15.05) a |
| Soil/plant total | (226.23 ± 21.6) a | (216.26 ± 36.7) b | (183.99 ± 16.3) c |
| Stored CO2eq | (828.94 ± 79.3) a | (792.41 ± 134.3) b | (674.17 ± 59.8) c |
| Emission CO2eq | (0.06 ± 0.08) a | (0.24 ± 0.11) a | (0.07 ± 0.04) a |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ribeiro, F.P.; Gatto, A.; Oliveira, A.D.d.; Pulrolnik, K.; Valadão, M.B.X.; Araújo, J.B.C.N.; Carvalho, A.M.d.; Ferreira, E.A.B. Carbon Storage in Different Compartments in Eucalyptus Stands and Native Cerrado Vegetation. Plants 2023, 12, 2751. https://doi.org/10.3390/plants12142751
Ribeiro FP, Gatto A, Oliveira ADd, Pulrolnik K, Valadão MBX, Araújo JBCN, Carvalho AMd, Ferreira EAB. Carbon Storage in Different Compartments in Eucalyptus Stands and Native Cerrado Vegetation. Plants. 2023; 12(14):2751. https://doi.org/10.3390/plants12142751
Chicago/Turabian StyleRibeiro, Fabiana Piontekowski, Alcides Gatto, Alexsandra Duarte de Oliveira, Karina Pulrolnik, Marco Bruno Xavier Valadão, Juliana Baldan Costa Neves Araújo, Arminda Moreira de Carvalho, and Eloisa Aparecida Belleza Ferreira. 2023. "Carbon Storage in Different Compartments in Eucalyptus Stands and Native Cerrado Vegetation" Plants 12, no. 14: 2751. https://doi.org/10.3390/plants12142751
APA StyleRibeiro, F. P., Gatto, A., Oliveira, A. D. d., Pulrolnik, K., Valadão, M. B. X., Araújo, J. B. C. N., Carvalho, A. M. d., & Ferreira, E. A. B. (2023). Carbon Storage in Different Compartments in Eucalyptus Stands and Native Cerrado Vegetation. Plants, 12(14), 2751. https://doi.org/10.3390/plants12142751