

Exogenous Sorbitol Application Confers Drought Tolerance to Maize Seedlings through Up-Regulating Antioxidant System and Endogenous Sorbitol Biosynthesis

 ,
,
Abstract
1. Introduction
2. Results
2.1. Exogenous Sorbitol Application Alleviates Growth Inhibition Effect Caused by Drought Stress
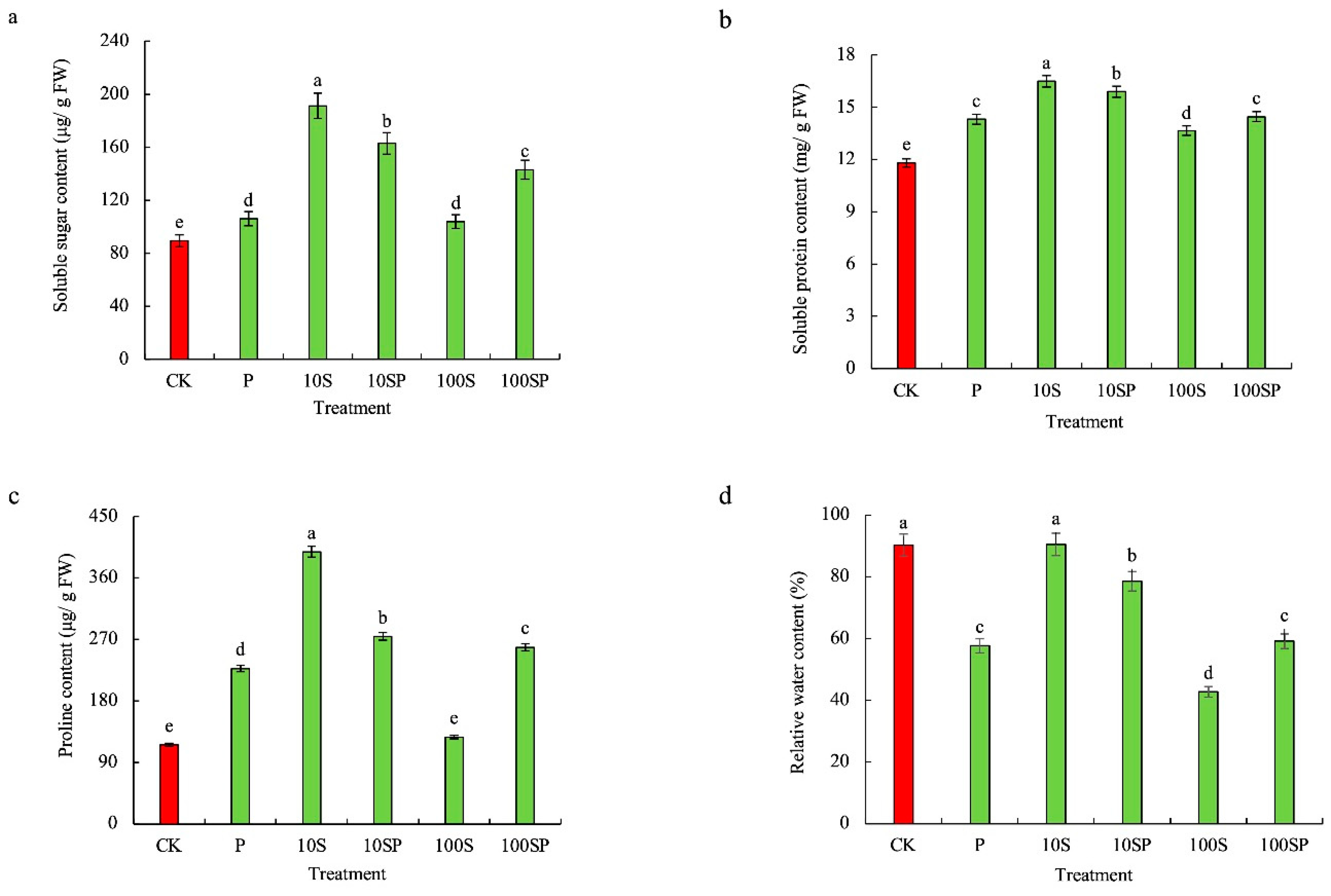
2.2. Effects of Exogenous Sorbitol Application on Chlorophyll Accumulation, Osmolyte Content and Relative Water Content of Maize Seedlings under Drought Stress
2.3. Effects of Exogenous Sorbitol Application on Lipid Peroxidation and Membrane Permeability of Maize Seedlings under Drought Stress
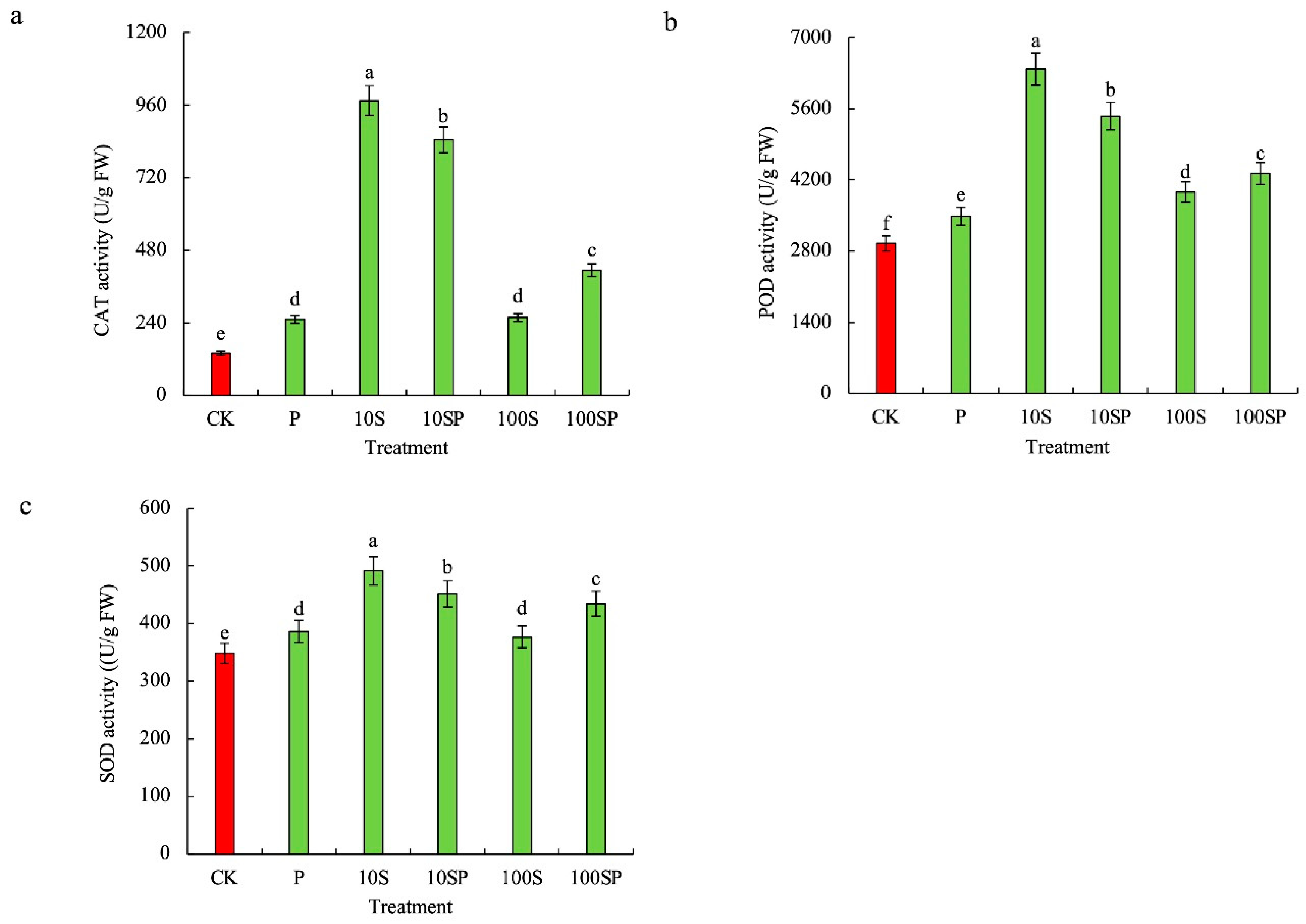
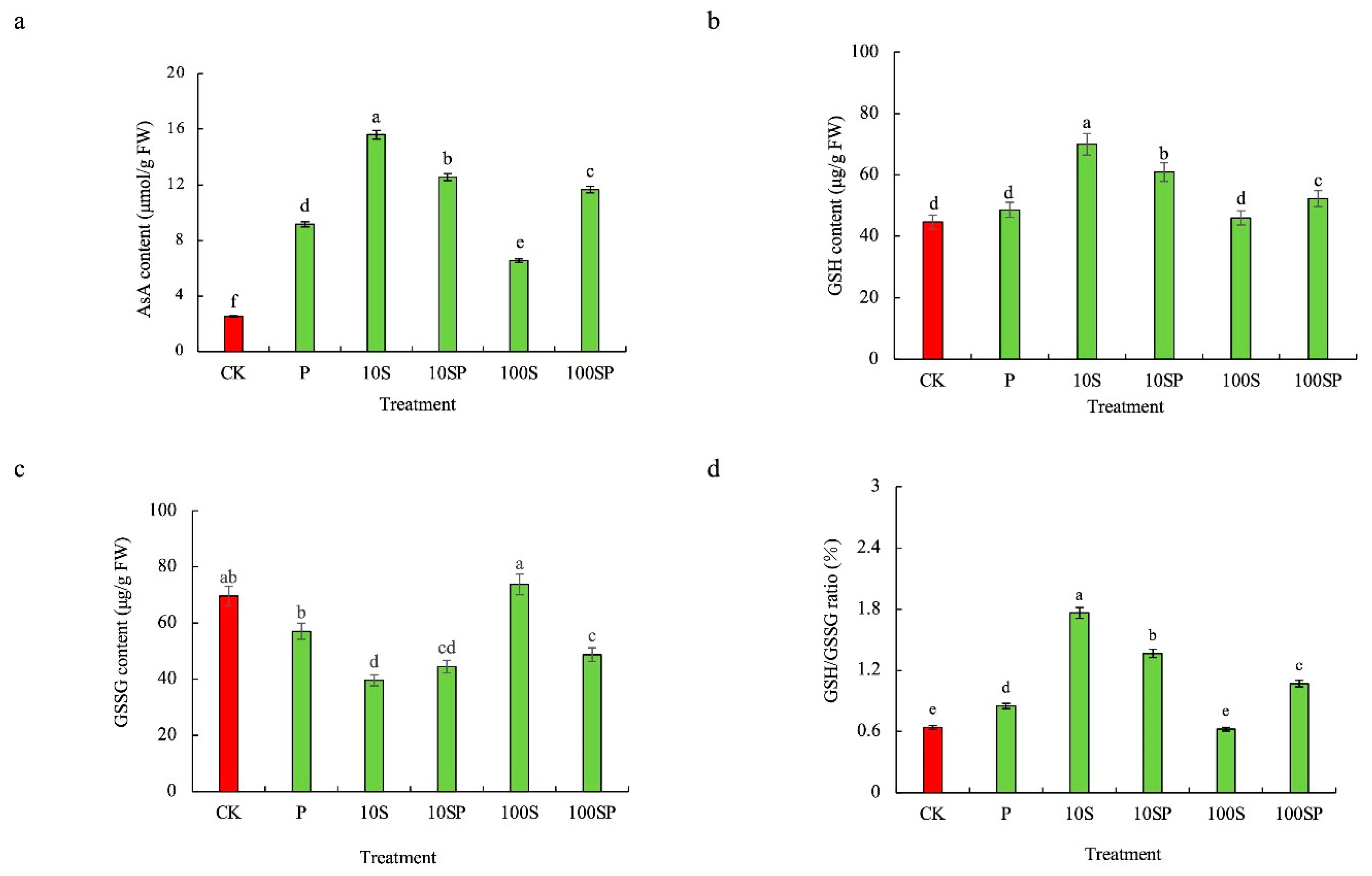
2.4. Effects of Exogenous Sorbitol Application on Antioxidant Systems of Maize Seedlings under Drought Stress
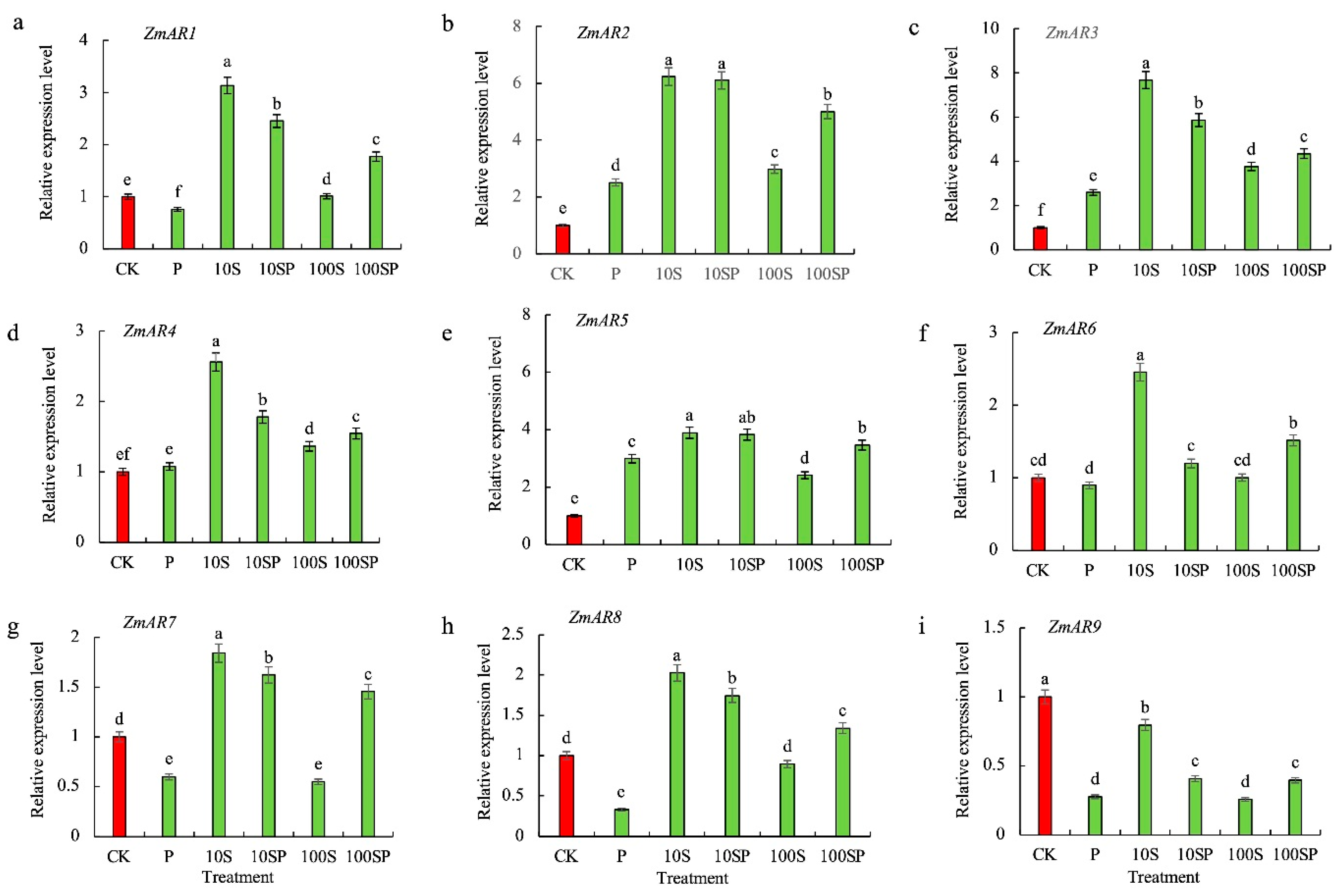
2.5. Effects of Exogenous Sorbitol Application on Sorbitol Content and Aldose Reductase Activities under Drought Stress
2.6. Effects of Exogenous Sorbitol on Transcript Levels of Aldose Reductase (AR)-Related Genes in Maize Seedlings under Drought Stress
3. Discussion
4. Methods
4.1. Plant Materials and Drought Treatment
4.2. Measurements of Growth Parameters, Relative Water Content and Sampling
4.3. Assays of Physiological Indicators
4.4. qRT-PCR Analysis of Aldose Reductase (AR)-Related Genes
4.5. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Sharma, J.K.; Sihmar, M.; Santal, A.R.; Singh, N.P. Impact assessment of major abiotic stresses on the proteome profiling of some important crop plants: A current update. Biotechnol. Genet. Eng. Rev. 2019, 35, 126–160. [Google Scholar] [CrossRef]
- Jin, X.; Liu, T.; Xu, J.; Gao, Z.; Hu, X. Exogenous GABA enhances muskmelon tolerance to salinity-alkalinity stress by regulating redox balance and chlorophyll biosynthesis. BMC Plant Biol. 2019, 19, 48. [Google Scholar] [CrossRef]
- Theerakulpisut, P.; Gunnula, W. Exogenous sorbitol and trehalose mitigated salt stress damage in salt-sensitive but not salt-tolerant rice seedlings. Asian. J. Crop Sci. 2012, 4, 165–170. [Google Scholar] [CrossRef]
- Khazaei, Z.; Estaji, A. Effect of foliar application of ascorbic acid on sweet pepper (Capsicum annuum) plants under drought stress. Acta Physiol. Plant. 2020, 42, 661–666. [Google Scholar] [CrossRef]
- Rezaei, Z.; Sarmast, M.K.; Atashi, S. 6-Benzylaminopurine (6-BA) ameliorates drought stress response in tall fescue via the influencing of biochemicals and strigolactone-signaling genes. Plant Physiol. Biochem. 2020, 155, 877–887. [Google Scholar] [CrossRef]
- Tayyab, N.; Naz, R.; Yasmin, H.; Nosheen, A.; Keyani, R.; Sajjad, M.; Hassan, M.N.; Roberts, T.H. Combined seed and foliar pre-treatments with exogenous methyl jasmonate and salicylic acid mitigate drought-induced stress in maize. PLoS ONE 2020, 15, e0232269. [Google Scholar] [CrossRef]
- Awan, S.A.; Khan, I.; Rizwan, M.; Zhang, X.; Brestic, M.; Khan, A.; El-Sheikh, M.A.; Alyemeni, M.N.; Ali, S.; Huang, L. Exogenous abscisic acid and jasmonic acid restrain polyethylene glycol-induced drought by improving the growth and antioxidative enzyme activities in pearl millet. Physiol. Plant. 2021, 172, 809–819. [Google Scholar] [CrossRef] [PubMed]
- Wani, S.H.; Singh, N.B.; Haribhushan, A.; Mir, J.I. Compatible solute engineering in plants for abiotic stress tolerance-role of glycine betaine. Curr. Genom. 2013, 14, 157–165. [Google Scholar] [CrossRef]
- Benitez, L.C.; Vighi, I.L.; Auler, P.A.; Amaral, M.N.; Moraes, G.P.; Rodrigues, G.S.; Maia, L.C.; Júnior, A.M.; Braga, E.J. Correlation of proline content and gene expression involved in the metabolism of this amino acid under abiotic stress. Acta Physiol. Plant. 2016, 38, 267. [Google Scholar] [CrossRef]
- Bieleski, R.L. Sugar alcohols, Plant carbohydrates I. Intracellular carbohydrates. In Encyclopedia of Plant Physiology; Loewus, F.A., Tanner, W., Eds.; Springer: Berlin/Heidelberg, Germany; New York, NY, USA, 1982; Volume 13A, pp. 158–192. [Google Scholar]
- Hsu, S.H.; Kao, C.H. Differential effect of sorbitol and polyethylene glycol on antioxidant enzymes in rice leaves. J. Plant Growth Regul. 2003, 39, 83–90. [Google Scholar] [CrossRef]
- Pommerrenig, B.; Papini-Terzi, F.S.; Sauer, N. Differential regulation of sorbitol and sucrose loading into the phloem of Plantago major in response to salt stress. Plant Physiol. 2007, 144, 1029. [Google Scholar] [CrossRef] [PubMed]
- Jain, M.; Tiwary, S.; Gadre, R. Sorbitol-induced changes in various growth and biochemici parameters in maize. Plant Soil Environ. 2010, 64, 263–267. [Google Scholar] [CrossRef]
- Rizwan, S.; Aftab, F. Morphological and biochemical responses of Jatropha curcas under osmotic stress. Int. J. Agric. Biol. 2018, 20, 1929–1936. [Google Scholar]
- Tari, I.; Kiss, G.; Deér, A.; Csiszár, J.; Erdei, L.; Gallé, A.; Gémes, K.; Horváth, F.; Poór, P.; Szepesi, A.; et al. Salicylic acid increased aldose reductase activity and sorbitol accumulation in tomato plants under salt stress. Biol. Plant. 2010, 54, 677–683. [Google Scholar] [CrossRef]
- Ho, L.H.; Rode, R.; Siegel, M.; Reinhardt, F.; Neuhaus, H.E.; Yvin, J.C.; Pluchon, S.; Hosseini, S.A.; Pommerrenig, B. Potassium application boosts photosynthesis and sorbitol biosynthesis and accelerates cold acclimation of common plantain (Plantago major L.). Plants 2020, 9, 1259. [Google Scholar] [CrossRef] [PubMed]
- Gao, M.; Tao, R.; Miura, K.; Dandekar, A.M.; Sugiura, A. Transformation of Japanese persimmon (Diospyros kaki Thunb.) with apple cDNA encoding NADP-dependent sorbitol-6-phosphate dehydrogenase. Plant Sci. 2001, 160, 837–845. [Google Scholar] [CrossRef] [PubMed]
- Deguchi, M.; Koshita, Y.; Gao, M.; Tao, R.; Tetsumura, T.; Yamaki, S.; Kanayama, Y. Engineered sorbitol accumulation induces dwarfism in Japanese persimmon. J. Plant Physiol. 2004, 161, 1177–1184. [Google Scholar] [CrossRef]
- Jez, J.M.; Flynn, T.G.; Penning, T.M. A new nomenclature for the aldo-keto reductase superfamily. Biochem. Pharmacol. 1997, 54, 639–647. [Google Scholar] [CrossRef]
- de Sousa, S.M.; Paniago, M.G.; Arruda, P.; Yunes, J.A. Sugar levels modulate sorbitol dehydrogenase expression in maize. Plant Mol. Biol. 2008, 68, 203–213. [Google Scholar] [CrossRef]
- Sengupta, D.; Naik, D.; Reddy, A.R. Plant aldo-keto reductases (AKRs) as multi-tasking soldiers involved in diverse plant metabolic processes and stress defense: A structure-function update. J. Plant Physiol. 2015, 179, 40–55. [Google Scholar] [CrossRef]
- Yang, X.Y.; Zhu, K.L.; Guo, X.M.; Pei, Y.H.; Zhao, M.A.; Song, X.Y.; Li, Y.B.; Liu, S.T.; Li, J. Constitutive expression of aldose reductase 1 from Zea mays exacerbates salt and drought sensitivity of transgenic Escherichia coli and Arabidopsis. Plant Physiol. Biochem. 2020, 156, 436–444. [Google Scholar] [CrossRef] [PubMed]
- Hussain, H.A.; Men, S.; Hussain, S.; Chen, Y.; Ali, S.; Zhang, S.; Zhang, K.; Li, Y.; Xu, Q.; Liao, C.; et al. Interactive effects of drought and heat stresses on morpho-physiological attributes, yield, nutrient uptake and oxidative status in maize hybrids. Sci. Rep. 2019, 9, 3890. [Google Scholar] [CrossRef] [PubMed]
- Yasin, M.; Ahmad, A.; Khaliq, T.; Habib-Ur-Rahman, M.; Niaz, S.; Gaiser, T.; Ghafoor, I.; Hassan, H.S.U.; Qasim, M.; Hoogenboom, G. Climate change impact uncertainty assessment and adaptations for sustainable maize production using multi-crop and climate models. Environ. Sci. Pollut. Res. Int. 2022, 29, 18967–18988. [Google Scholar] [CrossRef]
- Ling, M.H.; Han, H.B.; Hu, X.Y.; Xia, Q.Y.; Guo, X.M. Drought characteristics and causes during summer maize growth period on Huang-Huai-Hai Plain based on daily scale SPEI. Agric. Water Manag. 2023, 280, 108198. [Google Scholar] [CrossRef]
- Rubinstein, B. Regulation of H+ excretion: Effects of osmotic shock. Plant Physiol. 1982, 69, 939–944. [Google Scholar] [CrossRef]
- Abu-Romman, S. Responses of cucumber callus to sorbitol-induced osmotic stress. J. Genet. Eng. Biotechnol. 2010, 8, 45–50. [Google Scholar]
- Meher; Shivakrishna, P.; Ashokreddy, K.; Rao, D.M. Effect of PEG-6000 imposed drought stress on RNA content, relative water content (RWC), and chlorophyll content in peanut leaves and roots. Saudi J. Biol. Sci. 2018, 25, 285–289. [Google Scholar] [CrossRef]
- Ahmed, H.G.M.; Sajjad, M.; Li, M.; Azmat, M.A.; Rizwan, M.; Maqsood, R.H.; Khan, S.H. Selection criteria for drought-tolerant bread wheat genotypes at seedling stage. Sustainability 2019, 11, 2584. [Google Scholar] [CrossRef]
- Sattar, A.; Sher, A.; Ijaz, M.; Ul-Allah, S.; Rizwan, M.S.; Hussain, M.; Jabran, K.; Cheema, M.A. Terminal drought and heat stress alter physiological and biochemical attributes in flag leaf of bread wheat. PLoS ONE 2020, 15, e0232974. [Google Scholar] [CrossRef]
- Miller, G.; Suzuki, N.; Ciftci-Yilmaz, S.; Mittler, R. Reactive oxygen species homeostasis and signaling during drought and salinity stresses. Plant Cell Environ. 2010, 33, 453–467. [Google Scholar] [CrossRef]
- Singh, M.; Kumar, J.; Singh, S.; Singh, V.P.; Prasad, S.M. Roles of osmoprotectants in improving salinity and drought tolerance in plants: A review. Rev. Environ. Sci. Biotechnol. 2015, 14, 407–426. [Google Scholar] [CrossRef]
- Ashraf, M. Biotechnological approach of improving plant salt tolerance using antioxidants as markers. Biotechnol. Adv. 2009, 27, 84–93. [Google Scholar] [CrossRef] [PubMed]
- Gugliuzza, G.; Talluto, G.; Martinelli, F.; Farina, V.; Lo Bianco, R. Water deficit affects the growth and leaf metabolite composition of young loquat plants. Plants 2020, 19, 274. [Google Scholar] [CrossRef] [PubMed]
- Lo Bianco, R.; Rieger, M.; Sung, S. Effect of drought on sorbitol and sucrose metabolism in sinks and sources of peach. Physiol. Plant. 2000, 108, 71–78. [Google Scholar] [CrossRef]
- Arnon, D.I. Copper enzymes in isolated chloroplasts: Polyphenoloxidase in Beta vulgaris. Plant Physiol. 1949, 24, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Yusuf, C.S.; Chand, R.; Mishra, V.K.; Joshi, A.K. The association between leaf malondialdehyde and lignin content and resistance to spot blotch in wheat. J. Phytopathol. 2016, 164, 896–903. [Google Scholar] [CrossRef]
- Aminian, M.; Nabatchian, F.; Vaisi-Raygani, A.; Torabi, M. Mechanism of Coomassie Brilliant Blue G-250 binding to cetyltrimethylammonium bromide: An interference with the Bradford assay. Anal. Biochem. 2013, 434, 287–291. [Google Scholar] [CrossRef]
- Hackmann, T.J.; Keyser, B.L.; Firkins, J.L. Evaluation of methods to detect changes in reserve carbohydrate for mixed rumen microbes. J. Microbiol. Meth. 2013, 93, 284–291. [Google Scholar] [CrossRef]
- Martinez-Gutierrez, R.; Mora-Herrera, M.E.; Lopez-Delgado, H.A. Exogenous H2O2 in phytoplasma-infected potato plants promotes antioxidant activity and tuber production under drought conditions. Am. J. Potato Res. 2012, 89, 53–62. [Google Scholar] [CrossRef]
- Usluoglu, A.; Arabaci, G. The effect of acid dyes on antioxidant enzymes from cress (Lepidum sativum). Curr. Opin. Biotechnol. 2011, 22, 140. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment | Chlorophyll a (mg g−1 FW) | Chlorophyll b (mg g−1 FW) | Chlorophyll (a + b) (mg g−1 FW) |
|---|---|---|---|
| CK | 0.98 ± 0.04 b | 0.26 ± 0.02 b | 1.24 ± 0.03 b |
| P | 0.83 ± 0.05 c | 0.23 ± 0.01 b | 1.06 ± 0.05 c |
| 10S | 1.37 ± 0.05 a | 0.37 ± 0.02 a | 1.74 ± 0.06 a |
| 10SP | 0.90 ± 0.03 b | 0.24 ± 0.01 b | 1.14 ± 0.05 bc |
| 100S | 0.67 ± 0.03 d | 0.19 ± 0.02 c | 0.86 ± 0.03 d |
| 100SP | 0.91 ± 0.04 b | 0.27 ± 0.01 b | 1.18 ± 0.04 bc |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, J.; Zhao, M.; Liu, L.; Guo, X.; Pei, Y.; Wang, C.; Song, X. Exogenous Sorbitol Application Confers Drought Tolerance to Maize Seedlings through Up-Regulating Antioxidant System and Endogenous Sorbitol Biosynthesis. Plants 2023, 12, 2456. https://doi.org/10.3390/plants12132456
Li J, Zhao M, Liu L, Guo X, Pei Y, Wang C, Song X. Exogenous Sorbitol Application Confers Drought Tolerance to Maize Seedlings through Up-Regulating Antioxidant System and Endogenous Sorbitol Biosynthesis. Plants. 2023; 12(13):2456. https://doi.org/10.3390/plants12132456
Chicago/Turabian StyleLi, Jun, Meiai Zhao, Ligong Liu, Xinmei Guo, Yuhe Pei, Chunxiao Wang, and Xiyun Song. 2023. "Exogenous Sorbitol Application Confers Drought Tolerance to Maize Seedlings through Up-Regulating Antioxidant System and Endogenous Sorbitol Biosynthesis" Plants 12, no. 13: 2456. https://doi.org/10.3390/plants12132456
APA StyleLi, J., Zhao, M., Liu, L., Guo, X., Pei, Y., Wang, C., & Song, X. (2023). Exogenous Sorbitol Application Confers Drought Tolerance to Maize Seedlings through Up-Regulating Antioxidant System and Endogenous Sorbitol Biosynthesis. Plants, 12(13), 2456. https://doi.org/10.3390/plants12132456