
Phytoextraction and Migration Patterns of Cadmium in Contaminated Soils by Pennisetum hybridum



Abstract
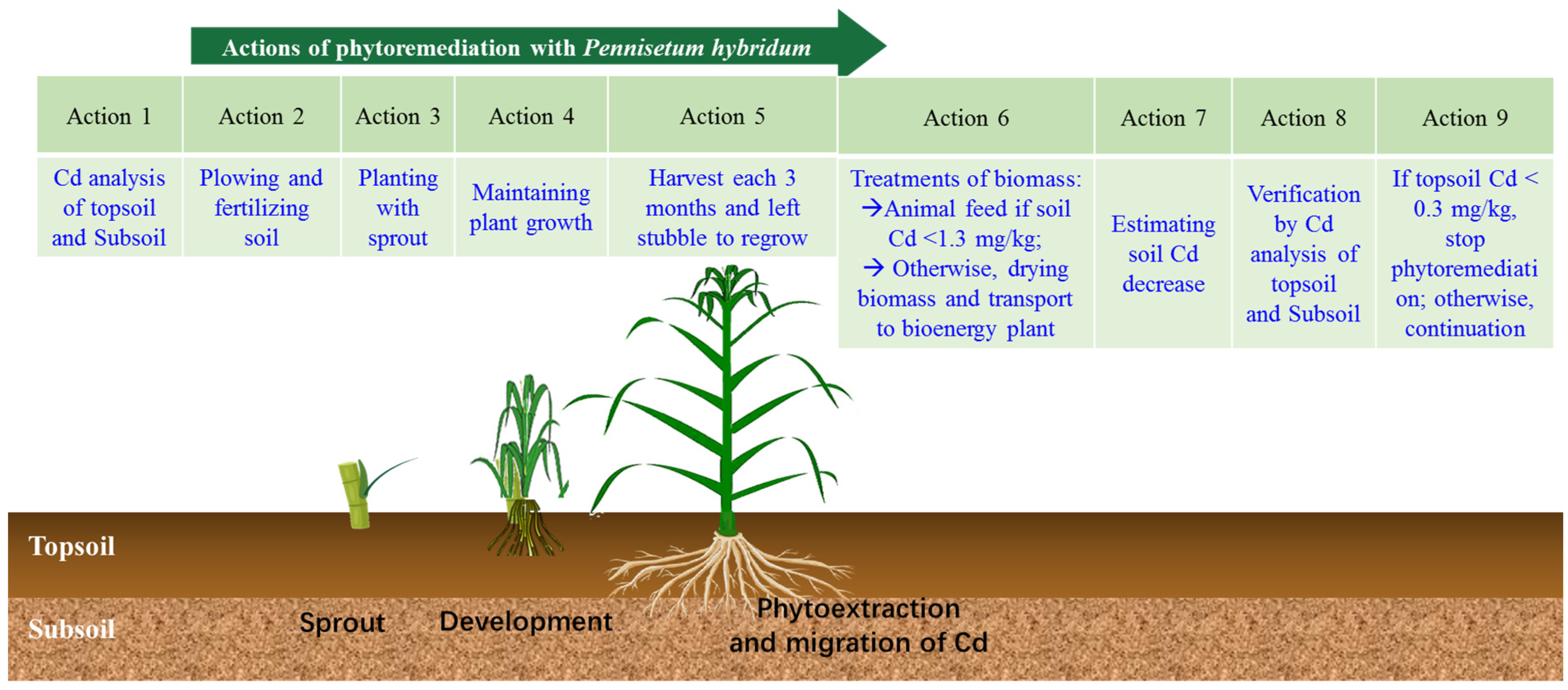
1. Introduction
2. Results
2.1. Plant Growth of P. hybridum
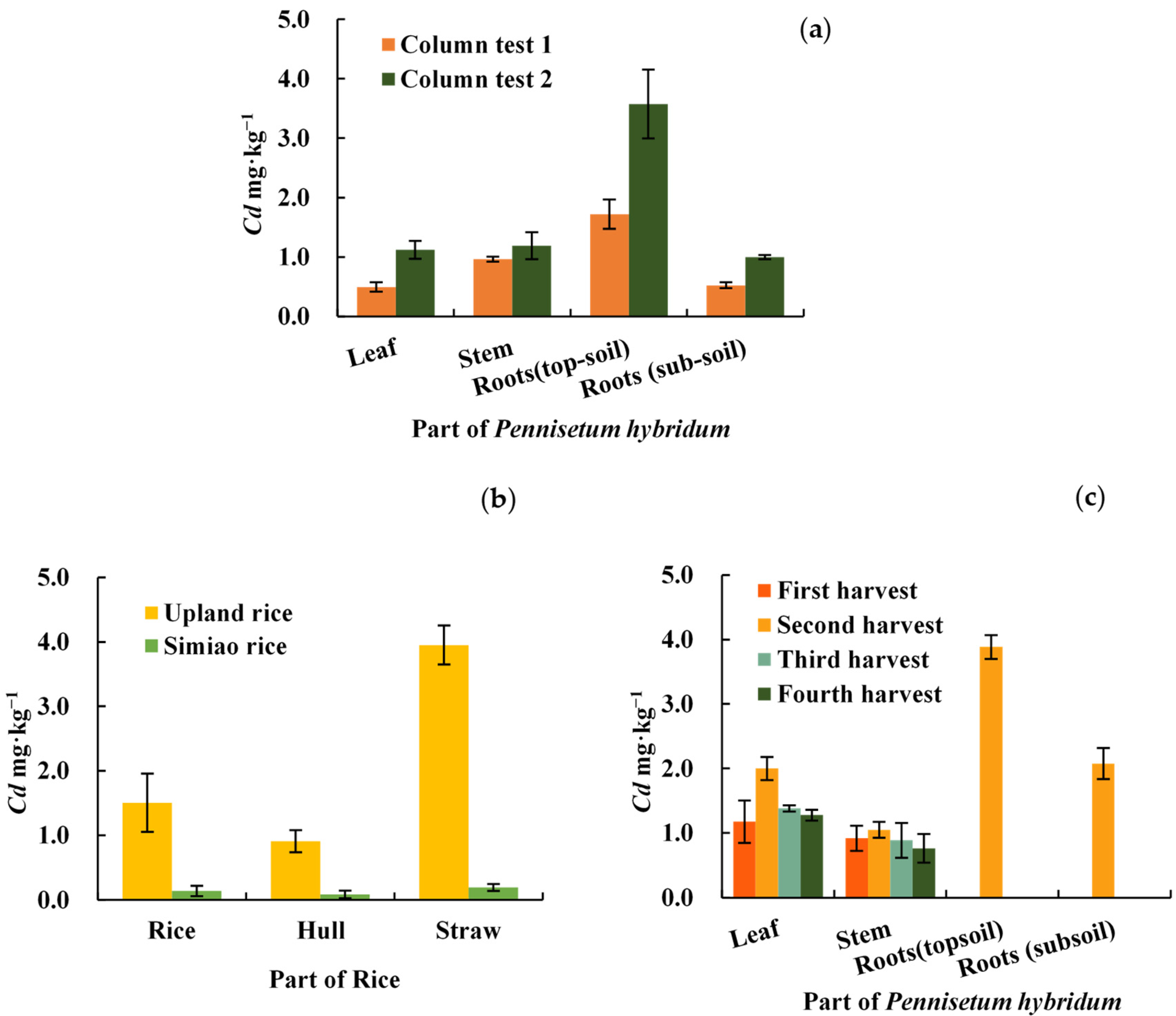
2.2. Cd Distribution in Various Parts of P. hybridum
2.3. Cd Uptake and Accumulation Patterns of P. hybridum
2.3.1. Bioconcentration and Translocation Factors of Cd in Various Parts of P. hybridum
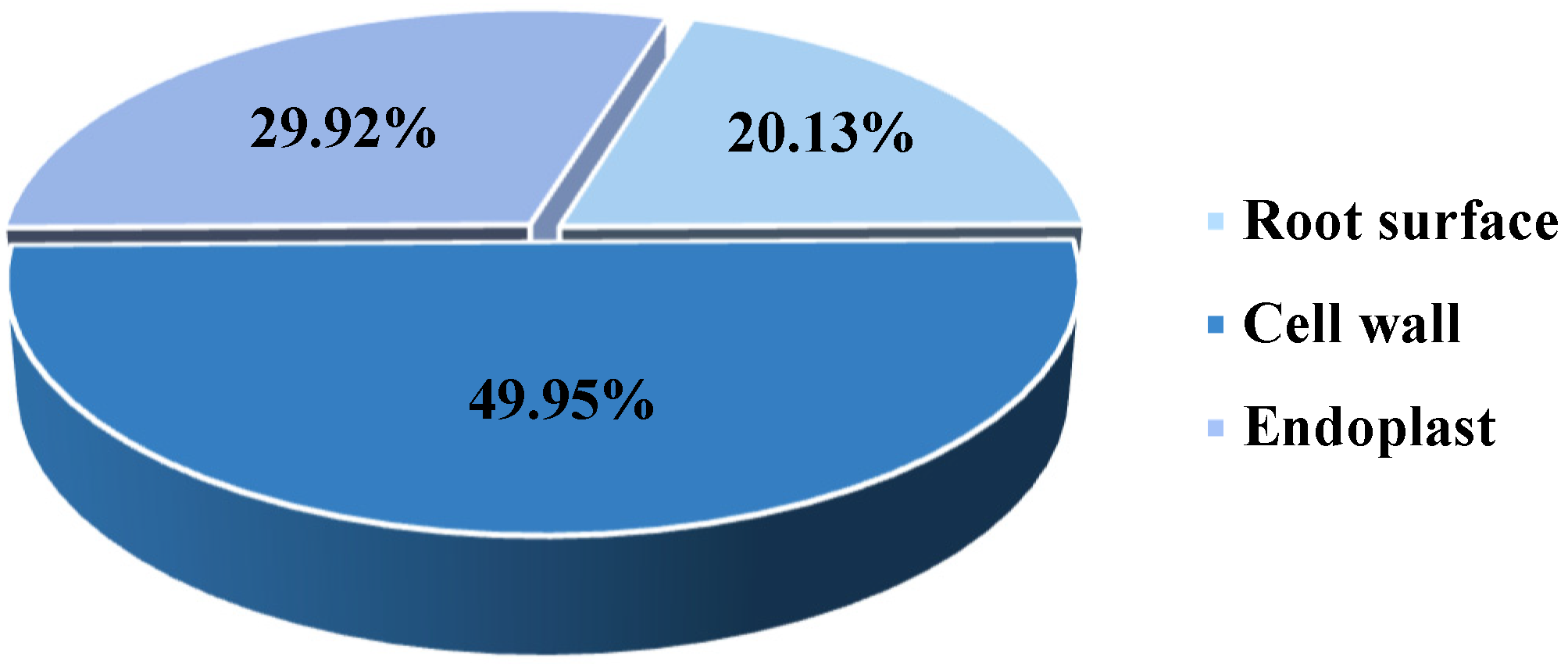
2.3.2. Location of Cd Storage in P. hybridum Roots
2.4. Phytoextraction Capacity of P. hybridum for Cd
2.5. Variation of Soil pH and Cd Concentration
2.5.1. Variation of Soil pH
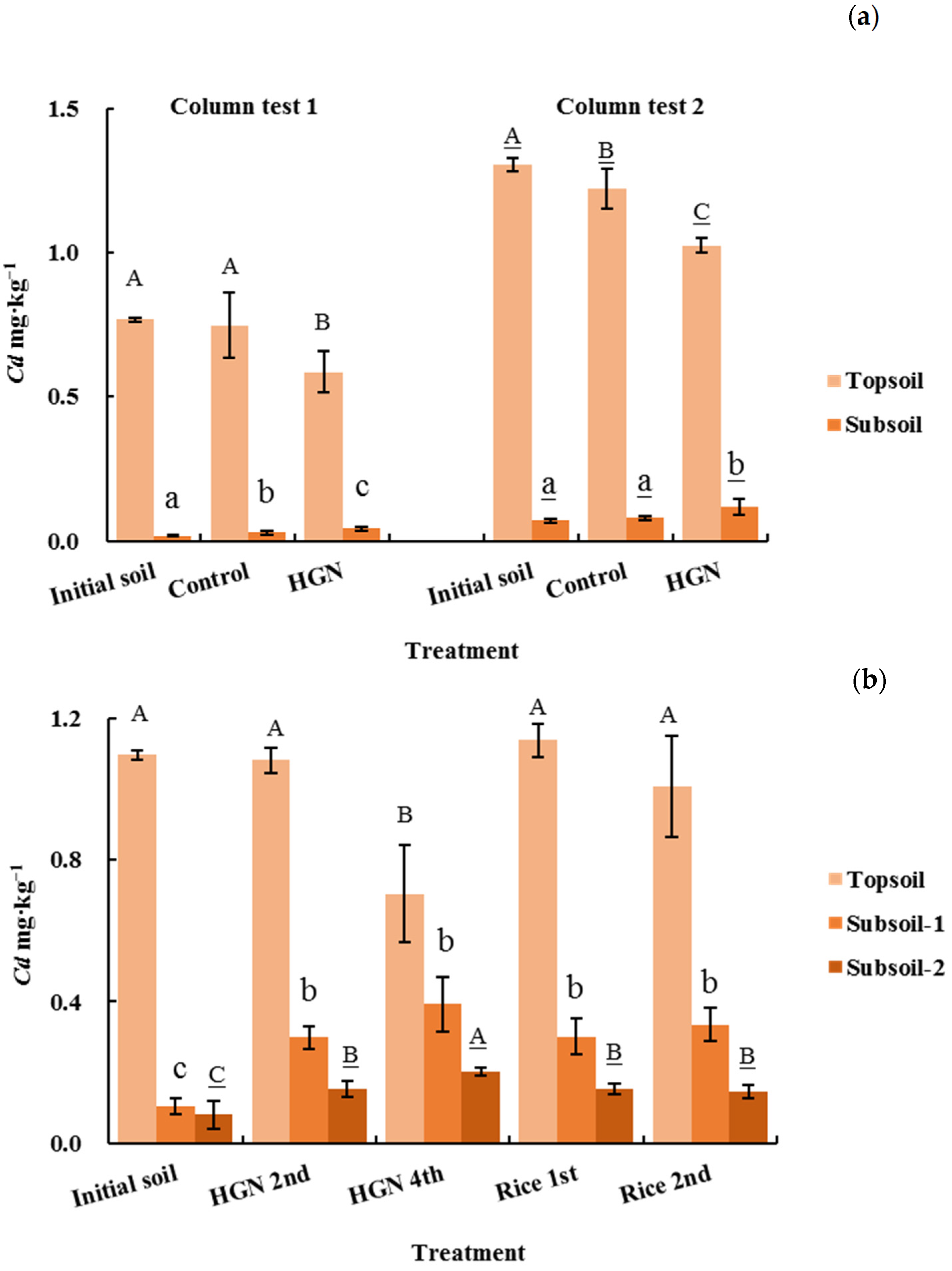
2.5.2. Variation of Soil Cd Concentration
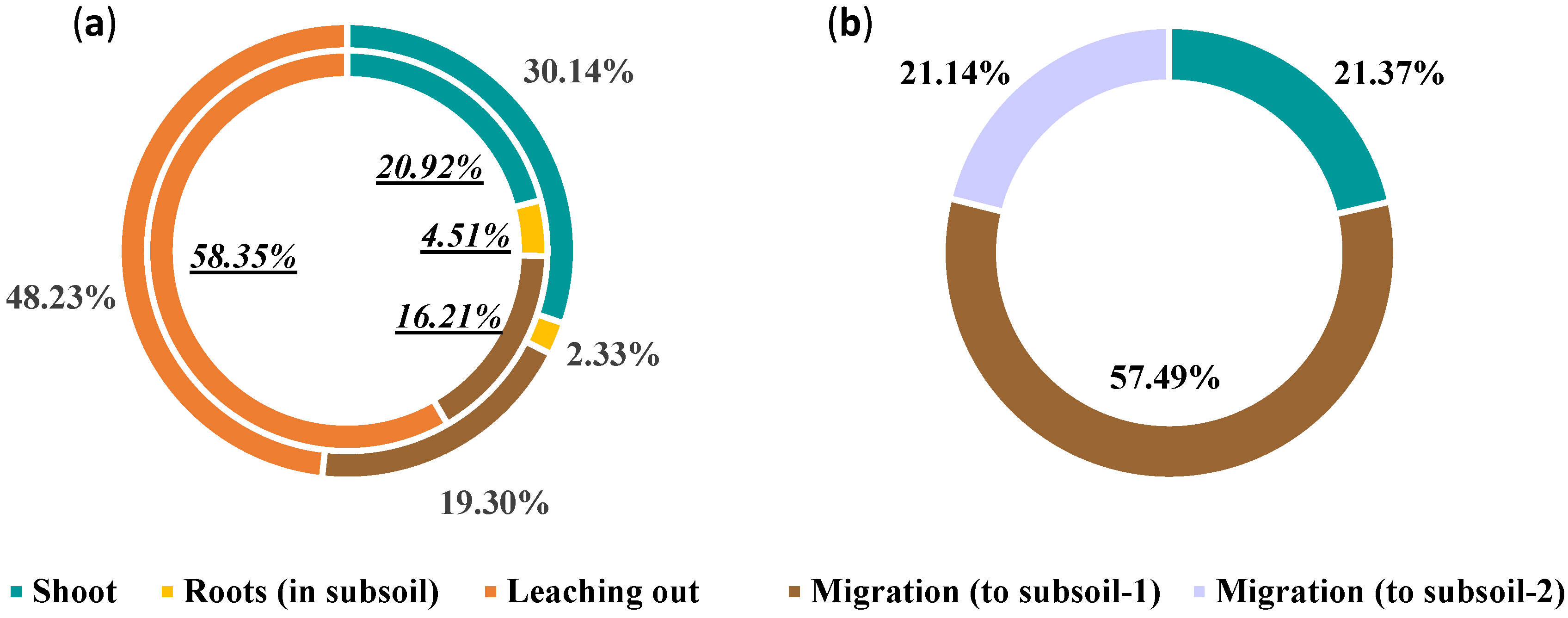
2.6. Soil Cd Migration and Removal Pathways
3. Discussion
3.1. Advantages of P. hybridum for Phytoremediation of Cd-Contaminated Soils
3.2. Forms of Cd Stored in Roots of P. hybridum
4. Materials and Methods
4.1. Experimental Materials
4.2. Experimental Design
4.2.1. Column Test
4.2.2. Lysimeter Test (12 July 2021–13 July 2022)
4.3. Sample Preparation and Analysis
4.4. Data Analysis
- Bioconcentration factor (BCF) = Cd concentration in a plant part/initial soil Cd total concentration;
- Translocation factor (TF) = Cd concentration in an aerial part/root Cd concentration;
- Extraction efficiency = Cd extraction amount by plants/initial total Cd content in soil;
- Removal rate = decrease in soil Cd content after planting/initial total Cd content in soil.
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kumar, A.; Sharma, T.; Mulla, S.I.; Kamyab, H.; Pant, D.; Sharma, S. Let’s Protect Our Earth: Environmental Challenges and Implications. In Microbes and Enzymes in Soil Health and Bioremediation (Microorganisms for Sustainability, 16); Springer: Singapore, 2019; pp. 1–10. [Google Scholar]
- Zhang, X.; Zhong, T.; Liu, L.; Ouyang, X. Impact of soil heavy metal pollution on food safety in China. PLoS ONE 2015, 10, e0135182. [Google Scholar] [CrossRef] [PubMed]
- Xue, W.; Zhang, X.; Zhang, C.; Wang, C.; Huang, Y.; Liu, Z. Mitigating the toxicity of reactive oxygen species induced by cadmium via restoring citrate valve and improving the stability of enzyme structure in rice. Chemosphere 2023, 327, 138511. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Tang, Z.; Wang, P.; Zhao, F.J. Geographical variations of cadmium and arsenic concentrations and arsenic speciation in Chinese rice. Env. Pollut. 2018, 238, 482–490. [Google Scholar] [CrossRef]
- Rizwan, M.; Ali, S.; Zia Ur Rehman, M.; Rinklebe, J.; Tsang, D.C.W.; Bashir, A.; Maqbool, A.; Tack, F.M.G.; Ok, Y.S. Cadmium phytoremediation potential of Brassica crop species: A review. Sci. Total Environ. 2018, 631–632, 1175–1191. [Google Scholar] [CrossRef]
- Hei, L.; Lee, C.C.; Wang, H.; Lin, X.Y.; Chen, X.H.; Wu, Q.T. Using a high biomass plant Pennisetum hydridum to phyto-treat fresh municipal sewage sludge. Bioresour. Technol. 2016, 217, 252–256. [Google Scholar] [CrossRef]
- Tan, F.; He, L.; Zhu, Q.; Wang, Y.; Chen, C.; He, M. Pennisetum hydridum: A potential energy crop with multiple functions and the current status in China. BioEnergy Res. 2021, 15, 850–862. [Google Scholar] [CrossRef]
- Wiangkham, N.; Prapagdee, B. Potential of Napier grass with cadmium-resistant bacterial inoculation on cadmium phytoremediation and its possibility to use as biomass fuel. Chemosphere 2018, 201, 511–518. [Google Scholar] [CrossRef]
- Hu, L.; Wang, R.; Liu, X.; Xu, B.; Xie, T.; Li, Y.; Wang, M.; Wang, G.; Chen, Y. Cadmium phytoextraction potential of king grass (Pennisetum sinese Roxb.) and responses of rhizosphere bacterial communities to a cadmium pollution gradient. Environ. Sci. Pollut. Res. Int. 2018, 25, 21671–21681. [Google Scholar] [CrossRef]
- He, L.; Zhu, Q.; Wang, Y.; Chen, C.; He, M.; Tan, F. Irrigating digestate to improve cadmium phytoremediation potential of Pennisetum hybridum. Chemosphere 2021, 279, 130592. [Google Scholar] [CrossRef]
- Zhang, X.F.; Wu, P.; Feng, J.F.; Guo, Y.H.; Gao, B. Effects of intercropping on Cd, Pb, and Zn accumulation using hyperaccumulators and energy plant. J. Agro-Environ. Sci. 2021, 40, 1481–1491. [Google Scholar]
- Ahmad, Z.; Khan, S.M.; Page, S.E.; Balzter, H.; Ullah, A.; Ali, S.; Jehangir, S.; Ejaz, U.; Afza, R.; Razzaq, A.; et al. Environmental sustainability and resilience in a polluted ecosystem via phytoremediation of heavy metals and plant physiological adaptations. J. Clean. Prod. 2023, 385, 135733. [Google Scholar] [CrossRef]
- Chen, Y.H.; Hu, L.; Liu, X.G.; Deng, Y.W.; Liu, M.J.; Xu, B.; Wang, M.K.; Wang, G. Influences of king grass (Pennisetum sinese Roxb)-enhanced approaches for phytoextraction and microbial communities in multi-metal contaminated soil. Geoderma 2017, 307, 253–266. [Google Scholar] [CrossRef]
- Zhou, Z.; Guo, Y.; Hu, L.; He, L.; Xu, B.; Huang, Z.; Wang, G.; Chen, Y. Potential use of king grass (Pennisetum purpureum Schumach. × Pennisetum glaucum (L.) R.Br.) for phytoextraction of cadmium from fields. Env. Sci. Pollut. Res. Int. 2020, 27, 35249–35260. [Google Scholar] [CrossRef] [PubMed]
- Yong, Y.; Wang, W.; Jiang, R.f.; Li, H.f. Comparison of phytoextraction efficiency of Cd with the hyperaccumulator Thlaspi caerulescens and three high biomass species. Acta Ecol. Sin. 2009, 29, 2732–2737. [Google Scholar]
- Shen, S.L.; Li, H.S.; Xia, B.C.; Yang, C.L. A Field experiment on phytoextraction of heavy metals from highly contaminated soil using big biomass plants of Sauropus androgynus and Manihot sp. J. Agro-Environ. Sci. 2013, 32, 572–578. [Google Scholar]
- Ma, C.J.; Ming, H.; Lin, C.H.; Naidu, R.; Bolan, N. Phytoextraction of heavy metal from tailing waste using Napier grass. Catena 2016, 136, 74–83. [Google Scholar] [CrossRef]
- Xie, H.; Zhao, X.M.; Xie, Z.; Wu, K.Q.; Li, X.L.; Yang, R.G.; Peng, B.; Yu, M.H.; He, J.H. Phytoremediation efficiency of Pennisetum hydridum for acid- and cadmium-polluted soil and its safe utilization. J. Agro-Environ. Sci. 2016, 35, 478–484. [Google Scholar]
- Liao, Y.; Fang, H.; Zhang, H.; Yu, Z.; Liu, Z.; Ma, X. Energy analysis and environmental impacts of Hybrid Giant Napier (Pennisetum Hydridum) direct-fired power generation in South China. IOP Conf. Ser. Mater. Sci. Eng. 2017, 199, 012094. [Google Scholar] [CrossRef]
- Ma, C.J.; Liu, F.G.; Lin, C.H. Study on improving the degraded soil with Napier grass (Pennisetum Hydridum). J. Shaoguan Univ. ·Nat. Sci. 2012, 33, 44–47. [Google Scholar]
- Sheoran, V.; Sheoran, A.S.; Poonia, P. Factors affecting phytoextraction: A Review. Pedosphere 2016, 26, 148–166. [Google Scholar] [CrossRef]
- Song, Y.; Jin, L.; Wang, X. Cadmium absorption and transportation pathways in plants. Int. J. Phytoremediat. 2017, 19, 133–141. [Google Scholar] [CrossRef] [PubMed]
- Gallego, S.M.; Pena, L.B.; Barcia, R.A.; Azpilicueta, C.E.; Lannone, M.F.; Rosales, E.P.; Zawoznik, M.S.; Groppa, M.D.; Benavides, M.P. Unravelling cadmium toxicity and tolerance in plants: Insight into regulatory mechanisms. Environ. Exp. Bot. 2012, 83, 33–46. [Google Scholar]
- Sun, X.X.; Zhu, J.; Tao, R.P.; Xiong, H.X.; Xu, Y.Q. Effect of exogenous iron on soil Cd accumulation of rice. J. Yangzhou Univ. (Nat. Sci. Ed.) 2022, 25, 74–78. [Google Scholar]
- Zhao, J.; Xia, B.; Meng, Y.; Yang, Z.; Pan, L.; Zhou, M.; Zhang, X. Transcriptome analysis to shed light on the molecular mechanisms of early responses to Cadmium in roots and leaves of king grass (Pennisetum americanum × P. purpureum). Int. J. Mol. Sci. 2019, 20, 2532. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.F.; Tian, C.; Gao, B. Heavy metal tolerance and phytoremediation potential of energy crop, king grass. Chin. J. Environ. Eng. 2017, 11, 3204–3213. [Google Scholar]
- Huang, R.; Xin, J.; Tian, R. Growth changes and characteristics of cadmium accumulation and distribution in Pistia stratiotes under cadmium stress. J. Agro-Environ. Sci. 2022, 41, 2033–2042. [Google Scholar]
- Hart, J.J.; Di Tomaso, J.M.; Linscott, D.L.; Kochian, L.V. Characterization of the transport and cellular compartmentation of paraquat in roots of intact maize seedlings. Pestic. Biochem. Physiol. 1992, 43, 212–222. [Google Scholar] [CrossRef]
- Sun, Y.; Ye, H.; Wei, Z.; Kong, X.; Wu, Q. Root cell walls and phytochelatins in low-cadmium cultivar of Brassica parachinensis. Pedosphere 2020, 30, 426–432. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Root Length (cm) | Height (cm) | Biomass (DW, g·plant−1) | ||||
|---|---|---|---|---|---|---|
| Root (in Topsoil) | Root (in Subsoil-1) | Stem | Leaf | |||
| Test 1 | 57.67 ± 2.08 | 174.00 ± 4.58 | 16.84 ± 1.09 | 9.60 ± 1.11 | 44.36 ± 2.52 | 42.27 ± 7.10 |
| Test 2 | 51.67 ± 4.51 | 190.00 ± 8.66 | 17.91 ± 1.94 | 11.59 ± 0.90 | 95.51 ± 2.25 | 25.96 ± 3.00 |
| Height (cm) | Biomass (DW, g·plant −1) | Yield (DW, 104 kg·ha−1) | |||
|---|---|---|---|---|---|
| Pennisetum hybridum | Stem | Leaf | Stem | Leaf | |
| First harvest | 312.50 ± 5.45 | 856.95 ± 92.15 | 332.86 ± 36.98 | 3.430 | 1.332 |
| Second harvest | 267.08 ± 12.14 | 919.83 ± 102.2 | 555.17 ± 62.00 | 3.681 | 2.222 |
| Third harvest | 259.17 ± 15.28 | 625.15 ± 47.32 | 478.89 ± 36.37 | 2.502 | 1.917 |
| Fourth harvest | 265.00 ± 10.00 | 721.52 ± 59.47 | 653.39 ± 54.68 | 2.888 | 2.615 |
| Rice | Straw | Grain | Straw | Grain | |
| Upland rice | 130.00 ± 6.00 | 31.73 ± 1.69 | 43.29 ± 1.95 | 0.5080 | 0.6930 |
| Simiao rice | 120.00 ± 5.50 | 22.96 ± 3.97 | 15.03 ± 1.48 | 0.3675 | 0.2406 |
| BCF | TF | |||||
|---|---|---|---|---|---|---|
| Root (in Topsoil) | Root (in Subsoil-1) | Stem | Leaf | Stem | Leaf | |
| Column test 1 | 2.24 | 0.68 | 1.25 | 0.65 | 0.56 | 0.29 |
| Column test 2 | 2.74 | 0.76 | 0.91 | 0.86 | 0.33 | 0.31 |
| Lysimeter test | 3.54 | 1.89 | 0.95 | 1.82 | 0.27 | 0.51 |
| Extraction Amount (mg·plant−1) | Extraction Efficiency (%) | ||||||
|---|---|---|---|---|---|---|---|
| Root (in Topsoil) | Root (in Subsoil-1) | Stem | Leaf | Total | Roots | Shoots | |
| Test 1 | 0.0567 ± 0.0044 | 0.0138 ± 0.0021 | 0.0428 ± 0.0043 | 0.0212 ± 0.0060 | 0.1345 | 4.60 | 4.17 |
| Test 2 | 0.0642 ± 0.0086 | 0.0116 ± 0.0008 | 0.1136 ± 0.0022 | 0.0363 ± 0.0034 | 0.2256 | 2.90 | 5.74 |
| Extraction Amount (mg·m–2) | Extraction Efficiency (%) | ||||
|---|---|---|---|---|---|
| Pennisetum hybridum | Stem | Leaf | Each Harvest | Annual | |
| First harvest | 3.153 ± 0.085 | 1.564 ± 0.042 | 4.717 | 23.40 | 8.53 |
| Second harvest | 3.850 ± 0.107 | 4.438 ± 0.123 | 8.288 | ||
| Third harvest | 2.216 ± 0.041 | 2.641 ± 0.049 | 4.857 | ||
| Fourth harvest | 2.207 ± 0.046 | 3.334 ± 0.069 | 5.541 | ||
| Rice | Straw | Grain | |||
| Upland rice | 2.006 ± 0.107 | 0.913 ± 0.041 | 2.919 | 3.02 | 1.10 |
| Simiao rice | 0.071 ± 0.012 | 0.029 ± 0.003 | 0.100 | ||
| Test 1 | Test 2 | |||||
|---|---|---|---|---|---|---|
| Initial Value | Control | P. hybridum | Initial Value | Control | P. hybridum | |
| Topsoil | 5.57 ± 0.03 A | 5.20 ± 0.27 B | 4.87 ± 0.05 C | 5.27 ± 0.01 a | 5.14 ± 0.01 b | 4.81 ± 0.01 b |
| Subsoil-1 | 4.47 ± 0.03 A | 5.20 ± 0.28 A | 3.76 ± 0.12 B | 6.27 ± 0.04 a | 6.42 ± 0.02 a | 5.86 ± 0.19 b |
| Initial Value | Pennisetum hybridum | Rice | |||
|---|---|---|---|---|---|
| Second Harvest | Fourth Harvest | Upland Rice | Simiao Rice | ||
| Topsoil | 5.58 ± 0.11 B | 5.06 ± 0.14 C | 4.37 ± 0.23 D | 5.47 ± 0.10 B | 5.95 ± 0.31 A |
| Subsoil-1 | 5.59 ± 0.25 A | 5.10 ± 0.03 C | 4.76 ± 0.17 D | 5.45 ± 0.03 AB | 5.30 ± 0.14 BC |
| Cd Concentration (mg·kg−1) | Cd Removal Rate (%) | ||
|---|---|---|---|
| Initial | After Planting | ||
| Column test 1 | 0.7676 ± 0.0077 | 0.5863 ± 0.0429 | 23.62 |
| Column test 2 | 1.3058 ± 0.0232 | 1.0250 ± 0.0265 | 21.50 |
| Lysimeter test | 1.0970 ± 0.0123 | 0.7041 ± 0.1385 | 35.81 |
| Shoots | Roots (in Topsoil) | Roots (in Subsoil) | Topsoil Reduction | Subsoil Increase | Leaching Out | |
|---|---|---|---|---|---|---|
| Planted test 1 | 0.0640 | 0.0567 | 0.0138 | 0.3626 | 0.0496 | 0.1785 |
| Planted test 2 | 0.1499 | 0.0642 | 0.0116 | 0.5616 | 0.0960 | 0.2399 |
| No plant test 1 | - | - | - | 0.0418 | 0.0209 | 0.0209 |
| No plant test 2 | - | - | - | 0.1677 | 0.0207 | 0.1470 |
| Column Test 1 | Column Test 2 | Lysimeter Test | ||||
|---|---|---|---|---|---|---|
| Topsoil | Subsoil | Topsoil | Subsoil | Topsoil | Subsoil | |
| pH | 5.57 ± 0.09 | 4.47 ± 0.04 | 5.27 ± 0.01 | 6.27 ± 0.04 | 4.95 ± 0.05 | 4.86 ± 0.11 |
| OM g·kg−1 | 34.42 ± 0.08 | 5.60 ± 0.02 | 39.94 ± 0.17 | 10.51 ± 0.30 | 39.53 ± 4.43 | 16.10 ± 2.46 |
| TN g·kg−1 | 1.96 ± 0.10 | 0.29 ± 0.02 | 1.27 ± 0.12 | 0.62 ± 0.06 | 2.24 ± 0.03 | 0.89 ± 0.11 |
| TP g·kg−1 | 0.75 ± 0.08 | 0.26 ± 0.02 | 0.44 ± 0.02 | 0.16 ± 0.02 | 0.47 ± 0.04 | 0.24 ± 0.02 |
| TK g·kg−1 | 7.59 ± 0.19 | 3.29 ± 0.62 | 9.56 ± 0.15 | 8.34 ± 0.30 | 10.43 ± 0.13 | 12.63 ± 0.20 |
| Cd mg·kg−1 | 0.7676 ± 0.0077 | 0.0181 ± 0.0019 | 1.3058 ± 0.0232 | 0.0698 ± 0.0078 | 1.0236 ± 0.0294 | 0.1472 ± 0.0298 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, C.; Wei, Z.; Hu, K.; Wu, Q.-T. Phytoextraction and Migration Patterns of Cadmium in Contaminated Soils by Pennisetum hybridum. Plants 2023, 12, 2321. https://doi.org/10.3390/plants12122321
Chen C, Wei Z, Hu K, Wu Q-T. Phytoextraction and Migration Patterns of Cadmium in Contaminated Soils by Pennisetum hybridum. Plants. 2023; 12(12):2321. https://doi.org/10.3390/plants12122321
Chicago/Turabian StyleChen, Canming, Zebin Wei, Kuangzheng Hu, and Qi-Tang Wu. 2023. "Phytoextraction and Migration Patterns of Cadmium in Contaminated Soils by Pennisetum hybridum" Plants 12, no. 12: 2321. https://doi.org/10.3390/plants12122321
APA StyleChen, C., Wei, Z., Hu, K., & Wu, Q.-T. (2023). Phytoextraction and Migration Patterns of Cadmium in Contaminated Soils by Pennisetum hybridum. Plants, 12(12), 2321. https://doi.org/10.3390/plants12122321