
Genetic Regulation of Mitosis–Meiosis Fate Decision in Plants: Is Callose an Oversighted Polysaccharide in These Processes?
Abstract
1. Introduction
2. Meiotic Cell-Fate Decision in Flowering Plants
3. Meiotic Cell Cycle Control in Flowering Plants
4. Callose: A Hallmark for Meiosis Initiation in Flowering Plants
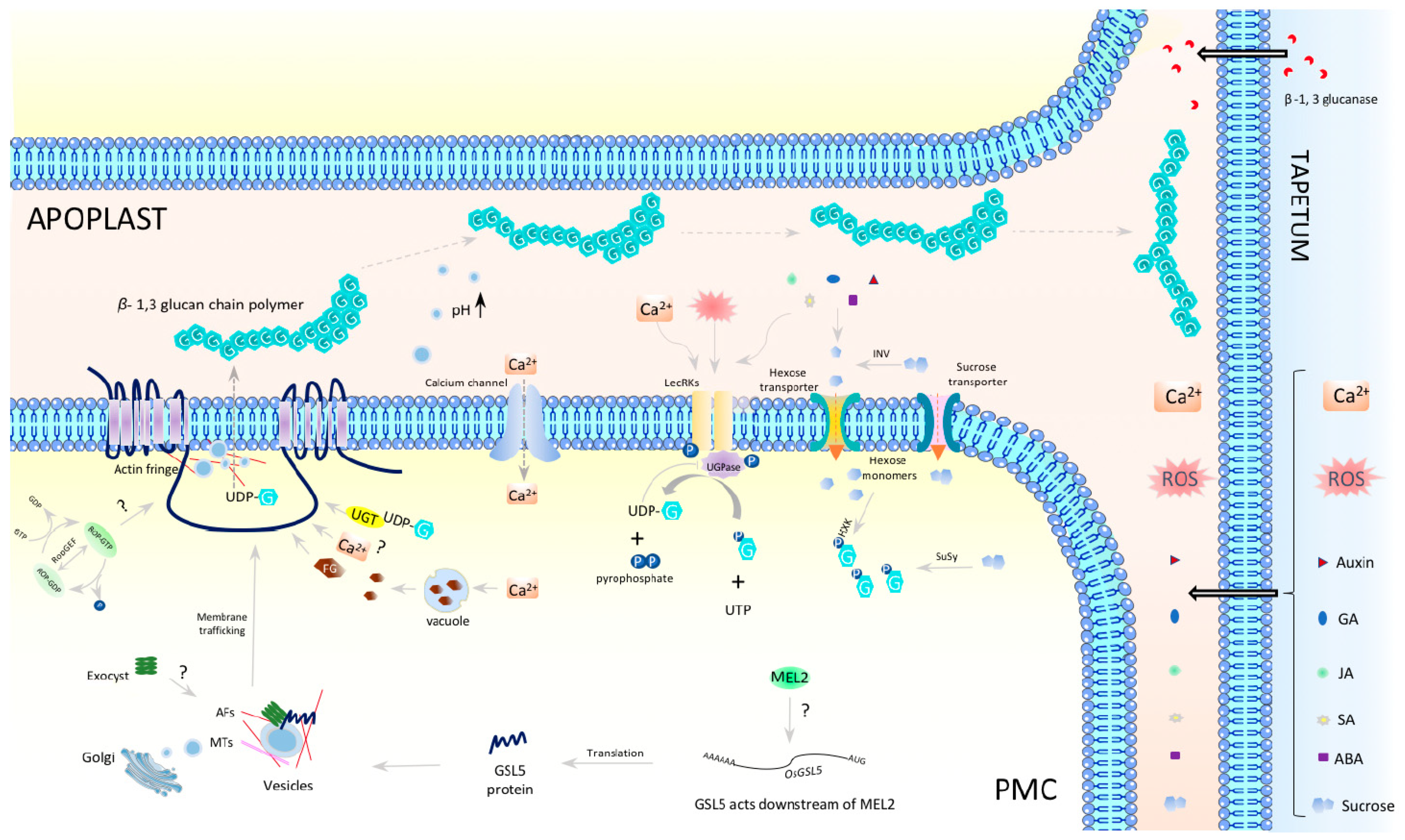
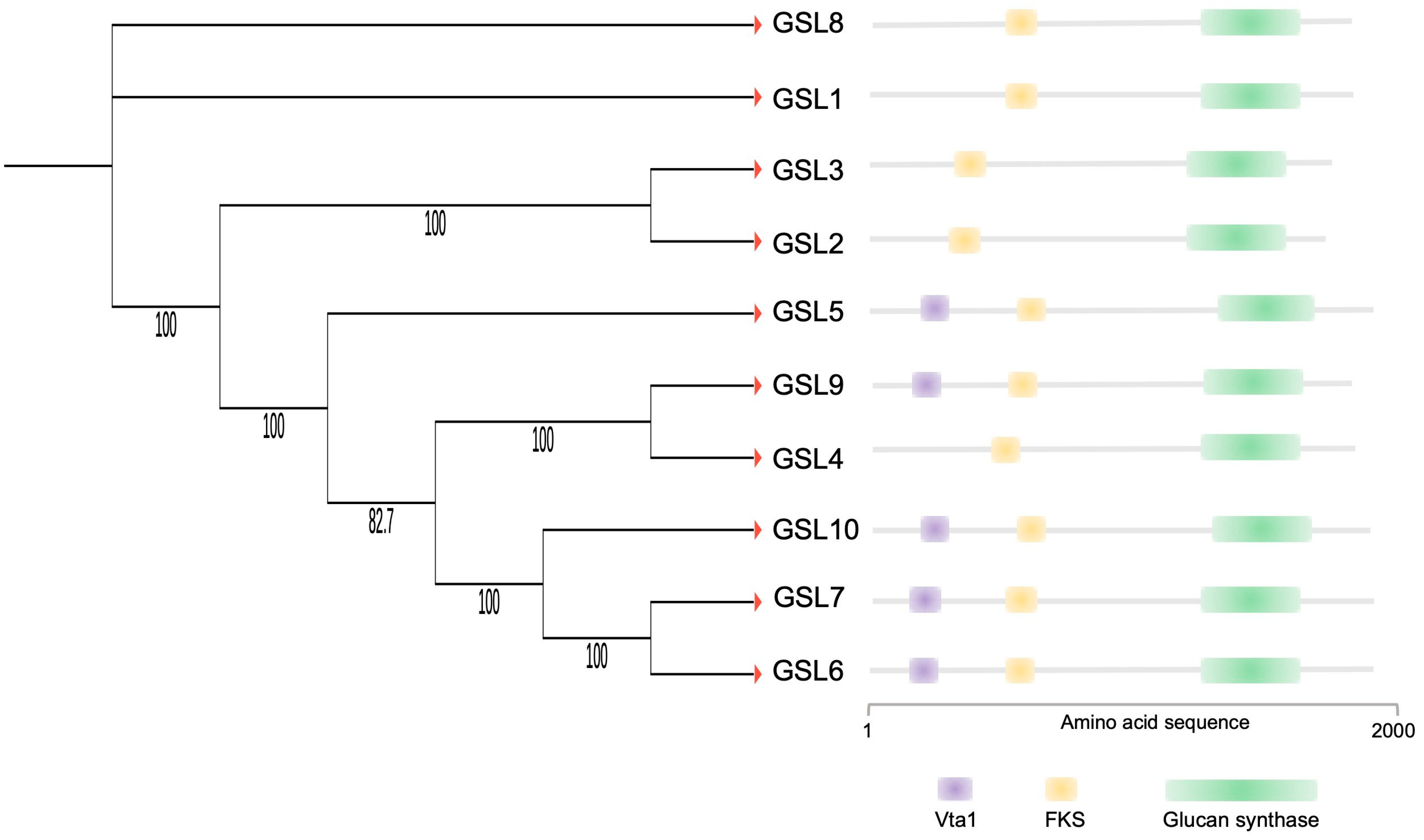
5. OsGSL5, a Central Player in Callose Biosynthesis in Anthers at Premeiosis and Meiosis
6. Fate of Meiosis and Pollen Formation in Plants Lacking Meiotic Callose
7. Future Directions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Watanabe, N.; Broome, M.; Hunter, T. Regulation of the human WEE1Hu CDK tyrosine 15-kinase during the cell cycle. EMBO J. 1995, 14, 1878–1891. [Google Scholar] [CrossRef] [PubMed]
- Murakami, H.; Keeney, S. Temporospatial Coordination of Meiotic DNA Replication and Recombination via DDK Recruitment to Replisomes. Cell 2014, 158, 861–873. [Google Scholar] [CrossRef] [PubMed]
- Higgins, J.D.; Perry, R.M.; Barakate, A.; Ramsay, L.; Waugh, R.; Halpin, C.; Armstrong, S.J.; Franklin, C.H. Spatiotemporal Asymmetry of the Meiotic Program Underlies the Predominantly Distal Distribution of Meiotic Crossovers in Barley. Plant Cell. 2012, 24, 4096–4109. [Google Scholar] [CrossRef]
- Pratto, F.; Brick, K.; Cheng, G.; Lam, K.G.; Cloutier, J.M.; Dahiya, D.; Wellard, S.R.; Jordan, P.W.; Camerini-Otero, R.D. Meiotic recombination mirrors patterns of germline replication in mice and humans. Cell 2021, 184, 4251–4267.e20. [Google Scholar] [CrossRef]
- Herskowitz, I. Life cycle of the budding yeast Saccharomyces cerevisiae. Microbiol. Rev. 1988, 52, 536–553. [Google Scholar] [CrossRef]
- Willer, M.; Hoffmann, L.; Styrkársdóttir, U.; Egel, R.; Davey, J.; Nielsen, O. Two-step activation of meiosis by the mat1 locus in Schizosaccharomyces pombe. Mol. Cell. Biol. 1995, 15, 4964–4970. [Google Scholar] [CrossRef]
- Kassir, Y.; Granot, D.; Simchen, G. IME1, a positive regulator gene of meiosis in S. cerevisiae. Cell 1988, 52, 853–862. [Google Scholar] [CrossRef]
- Smith, H.E.; Su, S.S.; Neigeborn, L.; Driscoll, S.E.; Mitchell, A.P. Role of IME1 expression in regulation of meiosis in Saccharomyces cerevisiae. Mol. Cell. Biol. 1990, 10, 6103–6113. [Google Scholar] [CrossRef]
- Bähler, J.; Schuchert, P.; Grimm, C.; Kohli, J. Synchronized meiosis and recombination in fission yeast: Observations with pat1-114 diploid cells. Curr. Genet. 1991, 19, 445–451. [Google Scholar] [CrossRef]
- Iino, Y.; Yamamoto, M. Negative control for the initiation of meiosis in Schizosaccharomyces pombe. Proc. Natl. Acad. Sci. USA 1985, 82, 2447–2451. [Google Scholar] [CrossRef]
- Goodenough, U.; Lin, H.; Lee, J.H. Sex determination in Chlamydomonas. Semin. Cell Dev. Biol. 2007, 18, 350–361. [Google Scholar] [CrossRef]
- Endo, T.; Mikedis, M.M.; Nicholls, P.K.; Page, D.C.; Rooij, D.G. Retinoic Acid and Germ Cell Development in the Ovary and Testis. Biomolecules 2019, 9, 775. [Google Scholar] [CrossRef] [PubMed]
- Koubova, J.; Hu, Y.C.; Bhattacharyya, T.; Soh, Y.Q.; Gill, M.E.; Goodheart, M.L.; Hogarth, C.A.; Griswold, M.D.; Page, D.C. Retinoic acid activates two pathways required for meiosis in mice. PLoS Genet. 2014, 10, 1004541. [Google Scholar] [CrossRef] [PubMed]
- Griswold, M.D.; Hogarth, C.A.; Bowles, J.; Koopman, P. Initiating meiosis: The case for retinoic acid. Biol. Reprod. 2012, 86, 35. [Google Scholar] [CrossRef]
- Childs, A.J.; Cowan, G.; Kinnell, H.L.; Anderson, R.A.; Saunders, P.T. Retinoic Acid signalling and the control of meiotic entry in the human fetal gonad. PLoS ONE 2011, 6, e20249. [Google Scholar] [CrossRef] [PubMed]
- Le Bouffant, R.; Guerquin, M.J.; Duquenne, C.; Frydman, N.; Coffigny, H.; Rouiller-Fabre, V.; Frydman, R.; Habert, R.; Livera, G. Meiosis initiation in the human ovary requires intrinsic retinoic acid synthesis. Hum. Reprod. 2010, 25, 2579–2590. [Google Scholar] [CrossRef] [PubMed]
- Yamaki, S.; Nagato, Y.; Kurata, N.; Nonomura, K. Ovule is a lateral organ finally differentiated from the terminating floral meristem in rice. Dev. Biol. 2011, 351, 208–216. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.; Wilson, Z.A. Stamen specification and anther development in rice. Chin. Sci. Bull. 2009, 54, 2342–2353. [Google Scholar] [CrossRef]
- Goldberg, R.B.; Beals, T.P.; Sanders, P.M. Anther development: Basic principles and practical applications. Plant Cell 1993, 5, 1217–1229. [Google Scholar] [CrossRef]
- Ali, Z.; Raza, Q.; Atif, R.M.; Aslam, U.; Ajmal, M.; Chung, G. Genetic and Molecular Control of Floral Organ Identity in Cereals. Int. J. Mol. Sci. 2019, 20, 2743. [Google Scholar] [CrossRef]
- Kuo, P.; Ines, O.; Lambing, C. Rewiring Meiosis for Crop Improvement. Front. Plant Sci. 2021, 12, 708948. [Google Scholar] [CrossRef] [PubMed]
- Nie, H.; Cheng, C.; Kong, J.; Li, H.; Hua, J. Plant non-coding RNAs function in pollen development and male sterility. Front. Plant Sci. 2023, 14, 1109941. [Google Scholar] [CrossRef]
- Sablowski, R. Control of patterning, growth, and differentiation by floral organ identity genes. J. Exp. Bot. 2015, 66, 1065–1073. [Google Scholar] [CrossRef] [PubMed]
- Twell, D. Male gametogenesis and germline specification in flowering plants. Sex Plant Reprod. 2011, 24, 149–160. [Google Scholar] [CrossRef] [PubMed]
- Wei, S.; Ma, L. Comprehensive Insight into Tapetum-Mediated Pollen Development in Arabidopsis thaliana. Cells 2023, 12, 247. [Google Scholar] [CrossRef]
- Matsoukas, I.G. Attainment of reproductive competence, phase transition, and quantification of juvenility in mutant genetic screens. Front. Plant Sci. 2014, 5, 32. [Google Scholar] [CrossRef]
- Bradley, D.; Carpenter, R.; Sommer, H.; Hartley, N.; Coen, E. Complementary floral homeotic phenotypes result from opposite orientations of a transposon at the plena locus of Antirrhinum. Cell 1993, 72, 85–95. [Google Scholar] [CrossRef]
- Dreni, L.; Pilatone, A.; Yun, D.; Erreni, S.; Pajoro, A.; Caporali, E.; Zhang, D.; Kater, M.M. Functional analysis of all AGAMOUS subfamily members in rice reveals their roles in reproductive organ identity determination and meristem determinacy. Plant Cell 2011, 23, 2850–2863. [Google Scholar] [CrossRef]
- Mizukami, Y.; Ma, H. Determination of Arabidopsis floral meristem identity by AGAMOUS. Plant Cell 1997, 9, 393–408. [Google Scholar] [CrossRef]
- Pelayo, M.A.; Yamaguchi, N.; Ito, T. One factor, many systems: The floral homeotic protein AGAMOUS and its epigenetic regulatory mechanisms. Curr. Opin. Plant Biol. 2021, 61, 102009. [Google Scholar] [CrossRef]
- Kelliher, T.; Walbot, V. Hypoxia triggers meiotic fate acquisition in maize. Science 2012, 337, 345–348. [Google Scholar] [CrossRef]
- Nonomura, K.; Morohoshi, A.; Nakano, M.; Eiguchi, M.; Miyao, A.; Hirochika, H.; Kurata, N. A germ cell specific gene of the ARGONAUTE family is essential for the progression of premeiotic mitosis and meiosis during sporogenesis in rice. Plant Cell 2007, 19, 2583–2594. [Google Scholar] [CrossRef] [PubMed]
- Komiya, R.; Ohyanagi, H.; Niihama, M.; Watanabe, T.; Nakano, M.; Kurata, N.; Nonomura, K. Rice germline-specific Argonaute MEL1 protein binds to phasiRNAs generated from more than 700 lincRNAs. Plant J. 2014, 78, 385–397. [Google Scholar] [CrossRef] [PubMed]
- Jiang, P.; Lian, B.; Liu, C.; Fu, Z.; Shen, Y.; Cheng, Z.; Qi, Y. 21-nt phasiRNAs direct target mRNA cleavage in rice male germ cells. Nature Commun. 2020, 11, 5191. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.C.; Lei, M.Q.; Zhou, Y.F.; Yang, Y.W.; Lian, J.P.; Yu, Y.; Feng, Y.Z.; Zhou, K.R.; He, R.R.; He, H.; et al. Reproductive phasiRNAs regulate reprogramming of gene expression and meiotic progression in rice. Nat. Commun. 2020, 11, 6031. [Google Scholar] [CrossRef]
- Hong, L.; Tang, D.; Zhu, K.; Wang, K.; Li, M.; Cheng, Z. Somatic and reproductive cell development in rice anther is regulated by a putative glutaredoxin. Plant Cell 2012, 24, 577–588. [Google Scholar] [CrossRef]
- Yang, H.; Li, Y.; Cao, Y.; Shi, W.; Xie, E.; Mu, N.; Du, G.; Shen, Y.; Tang, D.; Cheng, Z. Nitrogen nutrition contributes to plant fertility by affecting meiosis initiation. Nat. Commun. 2022, 13, 485. [Google Scholar] [CrossRef]
- Yang, W.C.; Ye, D.; Xu, J.; Sundaresan, V. The SPOROCYTELESS gene of Arabidopsis is required for initiation of sporogenesis and encodes a novel nuclear protein. Genes Dev. 1999, 13, 2108–2117. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Y.; Wang, D.; Ye, S.; Chen, W.; Li, G.; Xu, Z.; Bai, S.; Zhao, F. Auxin guides germ-cell specification in Arabidopsis anthers. Proc. Natl. Acad. Sci. USA 2021, 118, e2101492118. [Google Scholar] [CrossRef]
- Ren, L.; Tang, D.; Zhao, T.; Zhang, F.; Liu, C.; Xue, Z.; Shi, W.; Du, G.; Shen, Y.; Li, Y.; et al. OsSPL regulates meiotic fate acquisition in rice. New Phytol. 2018, 218, 789–803. [Google Scholar] [CrossRef] [PubMed]
- Li, L.C.; Qin, G.J.; Tsuge, T.; Hou, X.H.; Ding, M.Y.; Aoyama, T.; Oka, A.; Chen, Z.; Gu, H.; Zhao, Y.; et al. SPOROCYTELESS modulates YUCCA expression to regulate the development of lateral organs in Arabidopsis. New Phytol. 2008, 179, 751–764. [Google Scholar] [CrossRef]
- Yamaguchi, N.; Huang, J.; Tatsumi, Y.; Abe, M.; Sugano, S.S.; Kojima, M.; Takebayashi, Y.; Kiba, T.; Yokoyama, R.; Nishitani, K.; et al. Chromatin-mediated feed-forward auxin biosynthesis in floral meristem determinacy. Nat Commun. 2018, 9, 5290. [Google Scholar] [CrossRef] [PubMed]
- Sakata, T.; Oshino, T.; Miura, S.; Tomabechi, M.; Tsunaga, Y.; Higashitani, N.; Miyazawa, Y.; Takahashi, H.; Watanabe, M.; Higashitani, A. Auxins reverse plant male sterility caused by high temperatures. Proc. Natl. Acad. Sci. USA 2010, 107, 8569–8574. [Google Scholar] [CrossRef] [PubMed]
- Ross, K.J.; Fransz, P.; Jones, G.H. A light microscopic atlas of meiosis in Arabidopsis thaliana. Chromosome Res. Int. J. Mol. Supramol Evol. Asp. Chromosome Biol. 1996, 4, 507–516. [Google Scholar] [CrossRef] [PubMed]
- Whelan, E.D.P. Discontinuities in the Callose Wall, Intermeiocyte Connections, and Cytomixis in Angiosperm Meiocytes’. Can. J. Bot. 1974, 52, 1219–1224. [Google Scholar] [CrossRef]
- Magnard, J.L.; Yang, M.; Chen, Y.C.; Leary, M.; McCormick, S. The Arabidopsis gene tardy asynchronous meiosis is required for the normal pace and synchrony of cell division during male meiosis. Plant Physiol. 2001, 127, 1157–1166. [Google Scholar] [CrossRef] [PubMed]
- Lew, D.J.; Kornbluth, S. Regulatory roles of cyclin dependent kinase phosphorylation in cell cycle control. Curr. Opin. Cell Biol. 1996, 8, 795–804. [Google Scholar] [CrossRef]
- Nilsson, I.; Hoffmann, I. Cell cycle regulation by the Cdc25 phosphatase family. Prog. Cell Cycle Res. 2000, 4, 107–114. [Google Scholar] [CrossRef]
- Nigg, E.A. Mitotic kinases as regulators of cell division and its checkpoints. Nat. Rev. Mol. Cell Biol. 2001, 2, 21–32. [Google Scholar] [CrossRef]
- Ohi, R.; Gould, K.L. Regulating the onset of mitosis. Curr Opin Cell Biol. 1999, 11, 267–273. [Google Scholar] [CrossRef]
- Komaki, S.; Sugimoto, K. Control of the plant cell cycle by developmental and environmental cues. Plant Cell Physiol. 2012, 53, 953–964. [Google Scholar] [CrossRef]
- Shimotohno, A.; Aki, S.S.; Takahashi, N.; Umeda, M. Regulation of the Plant Cell Cycle in Response to Hormones and the Environment. Annu. Rev. Plant Biol. 2021, 72, 273–296. [Google Scholar] [CrossRef] [PubMed]
- d’Erfurth, I.; Cromer, L.; Jolivet, S.; Girard, C.; Horlow, C.; Sun, Y.; To, J.P.; Berchowitz, L.E.; Copenhaver, G.P.; Mercier, R. The cyclin-A CYCA1;2/TAM is required for the meiosis I to meiosis II transition and cooperates with OSD1 for the prophase to first meiotic division transition. PLoS Genet. 2010, 6, e1000989. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Magnard, J.L.; McCormick, S.; Yang, M. Progression through meiosis I and meiosis II in Arabidopsis anthers is regulated by an A-type cyclin predominately expressed in prophase I. Plant Physiol. 2004, 136, 4127–4135. [Google Scholar] [CrossRef][Green Version]
- Azumi, Y.; Liu, D.; Zhao, D.; Li, W.; Wang, G.; Hu, Y.; Ma, H. Homolog interaction during meiotic prophase I in Arabidopsis requires the SOLO DANCERS gene encoding a novel cyclin-like protein. EMBO J. 2002, 21, 3081–3095. [Google Scholar] [CrossRef]
- Zhao, X.; Bramsiepe, J.; Van Durme, M.; Komaki, S.; Prusicki, M.A.; Maruyama, D.; Forner, J.; Medzihradszky, A.; Wijnker, E.; Harashima, H.; et al. RETINOBLASTOMA RELATED1 mediates germline entry in Arabidopsis. Science 2017, 356, eaaf6532. [Google Scholar] [CrossRef]
- Golubovskaya, I.; Grebennikova, Z.K.; Avalkina, N.A.; Sheridan, W.F. The role of the ameiotic1 gene in the initiation of meiosis and in subsequent meiotic events in maize. Genetics 1993, 135, 1151–1166. [Google Scholar] [CrossRef]
- Pawlowski, W.P.; Wang, C.J.; Golubovskaya, I.N.; Szymaniak, J.M.; Shi, L.; Hamant, O.; Zhu, T.; Harper, L.; Sheridan, W.F.; Cande, W.Z. Maize AMEIOTIC1 is essential for multiple early meiotic processes and likely required for the initiation of meiosis. Proc. Natl. Acad. Sci. USA 2009, 106, 3603–3608. [Google Scholar] [CrossRef]
- Mercier, R.; Armstrong, S.J.; Horlow, C.; Jackson, N.P.; Makaroff, C.A.; Vezon, D.; Pelletier, G.; Jones, G.H.; Franklin, F.C. The meiotic protein SWI1 is required for axial element formation and recombination initiation in Arabidopsis. Development 2003, 130, 3309–3318. [Google Scholar] [CrossRef]
- Mercier, R.; Vezon, D.; Bullier, E.; Motamayor, J.C.; Sellier, A.; Lefèvre, F.; Pelletier, G.; Horlow, C. SWITCH1 (SWI1): A novel protein required for the establishment of sister chromatid cohesion and for bivalent formation at meiosis. Genes Dev. 2001, 15, 1859–1871. [Google Scholar] [CrossRef]
- Ravi, M.; Marimuthu, M.P.; Siddiqi, I. Gamete formation without meiosis in Arabidopsis. Nature 2008, 451, 1121–1124. [Google Scholar] [CrossRef]
- Che, L.; Tang, D.; Wang, K.; Wang, M.; Zhu, K.; Yu, H.; Gu, M.; Cheng, Z. OsAM1 is required for leptotene-zygotene transition in rice. Cell Res. 2011, 21, 654–665. [Google Scholar] [CrossRef]
- Nan, G.L.; Ronceret, A.; Wang, R.; Fernandes, J.F.T.; Zacheus Cande, W.; Walbot, V. Global transcriptome analysis of two ameiotic1 alleles in maize anthers: Defining steps in meiotic entry and progression through prophase I. BMC Plant Biol. 2011, 11, 120. [Google Scholar] [CrossRef]
- Reddy, T.V.; Kaur, J.; Agashe, B.; Sundaresan, V.; Siddiqi, I. The DUET gene is necessary for chromosome organization and progression during male meiosis in Arabidopsis and encodes a PHD finger protein. Development 2003, 130, 5975–5987. [Google Scholar] [CrossRef]
- Li, J.; Dukowic-Schulze, S.; Lindquist, I.E.; Farmer, A.D.; Kelly, B.; Li, T.; Smith, A.G.; Retzel, E.F.; Mudge, J.; Chen, C. The plant-specific protein FEHLSTART controls male meiotic entry, initializing meiotic synchronization in Arabidopsis. Plant J. 2015, 84, 659–671. [Google Scholar] [CrossRef]
- Kapoor, S.; Takatsuji, H. Silencing of an anther-specific zinc-finger gene, MEZ1, causes aberrant meiosis and pollen abortion in petunia. Plant Mol. Biol. 2006, 61, 415–430. [Google Scholar] [CrossRef] [PubMed]
- Nonomura, K.; Eiguchi, M.; Nakano, M.; Takashima, K.; Komeda, N.; Fukuchi, S.; Miyazaki, S.; Miyao, A.; Hirochika, H.; Kurata, N. A novel RNA-recognition-motif protein is required for premeiotic G1/S-phase transition in rice (Oryza sativa L.). PLoS Genet. 2011, 7, e1001265. [Google Scholar] [CrossRef]
- Zhao, T.; Ren, L.; Chen, X.; Yu, H.; Liu, C.; Shen, Y.; Shi, W.; Tang, D.; Du, G.; Li, Y.; et al. The OsRR24/LEPTO1 Type-B Response Regulator is Essential for the Organization of Leptotene Chromosomes in Rice Meiosis. Plant Cell 2018, 30, 3024–3037. [Google Scholar] [CrossRef]
- Prymakowska-Bosak, M.; Przewłoka, M.R.; Slusarczyk, J.; Kuraś, M.; Lichota, J.; Kiliańczyk, B.; Jerzmanowski, A. Linker histones play a role in male meiosis and the development of pollen grains in tobacco. Plant Cell 1999, 11, 2317–2329. [Google Scholar] [CrossRef]
- Jackson, N.; Sanchez-Moran, E.; Buckling, E.; Armstrong, S.J.; Jones, G.H.; Franklin, F.C. Reduced meiotic crossovers and delayed prophase I progression in AtMLH3-deficient Arabidopsis. EMBO J. 2006, 25, 1315–1323. [Google Scholar] [CrossRef]
- Byzova, M.V.; Franken, J.; Aarts, M.G.; de Almeida-Engler, J.; Engler, G.; Mariani, C.; Van Lookeren Campagne, M.M.; Angenent, G.C. Arabidopsis STERILE APETALA, a multifunctional gene regulating inflorescence, flower, and ovule development. Genes Dev. 1999, 13, 1002–1014. [Google Scholar] [CrossRef]
- Shi, X.; Sun, X.; Zhang, Z.; Feng, D.; Zhang, Q.; Han, L.; Wu, J.; Lu, T. GLUCAN SYNTHASE-LIKE 5 (GSL5) plays an essential role in male fertility by regulating callose metabolism during microsporogenesis in rice. Plant Cell Physiol. 2015, 56, 497–509. [Google Scholar] [CrossRef]
- Somashekar, H.; Mimura, M.; Tsuda, K.; Nonomura, K.I. Rice GLUCAN SYNTHASE-LIKE5 promotes anther callose deposition to maintain meiosis initiation and progression. Plant Physiol. 2023, 191, 400–413. [Google Scholar] [CrossRef]
- Matsuo, Y.; Arimura, S.; Tsutsumi, N. Distribution of cellulosic wall in the anthers of Arabidopsis during microsporogenesis. Plant Cell Rep. 2013, 32, 1743–1750. [Google Scholar] [CrossRef] [PubMed]
- Heslop-Harrison, J. Cell Walls, Cell Membranes and Protoplasmic Connections during Meiosis and Pollen Development. In Pollen Physiology and Fertilisation; Linskens, H.F., Ed.; North Holland Publishers: Amsterdam, The Netherlands, 1964. [Google Scholar]
- Heslop-Harrison, J.; Mackenzie, A. Autoradiography of soluble [2-14-C]thymidine derivatives during meiosis and microsporogenesis in Lilium anthers. J. Cell Sci. 1967, 2, 387–400. [Google Scholar] [CrossRef] [PubMed]
- Rodkiewicz, B. Callose in cell walls during megasporogenesis in angiosperms. Planta 1970, 93, 39–47. [Google Scholar] [CrossRef] [PubMed]
- Waterkeyn, L.; Bienfait, A. On a Possible Function of the Callosic Special Wall in Ipomoea purpurea (L.). Roth. Grana 1970, 10, 13–20. [Google Scholar] [CrossRef]
- Rodriguez-García, M.I.; Majewska-Sawka, A. Is the special callose wall of microsporocytes an impermeable barrier? J. Exp. Bot. 1992, 43, 1659–1663. [Google Scholar] [CrossRef]
- Yim, K.O.; Bradford, K.J. Callose deposition is responsible for apoplastic semipermeability of the endosperm envelope of muskmelon seeds1. Plant Physiol. 1998, 118, 83–90. [Google Scholar] [CrossRef]
- Godwin, H. The origin of exine. New Phytol. 1968, 67, 667. [Google Scholar] [CrossRef]
- Halac, I.N.; Hatte, C. Female Gametophytic Competence in Relation to Polarisation Phenomena during Megagametogenesis and Development of the Embryo Sac in the Genus Oenothera. In Gamete Competition in Plants and Animals; Muscaly, D.L., Ed.; North-Holland Publishing: Amsterdam, The Netherlands, 1975. [Google Scholar]
- Unal, M.; Vardar, F.; Ayturk, Z. Callose in Plant Sexual Reproduction. In Current Progress in Biological Research; InTech: London, UK, 2013. [Google Scholar] [CrossRef]
- Wan, L.; Zha, W.; Cheng, X.; Liu, C.; Lv, L.; Liu, C.; Wang, Z.; Du, B.; Chen, R.; Zhu, L.; et al. A rice β-1,3-glucanase gene Osg1 is required for callose degradation in pollen development. Planta 2011, 233, 309–323. [Google Scholar] [CrossRef] [PubMed]
- Abramova, L.I.; Avalkina, N.A.; Golubeva, E.A.; Golubovskava, I.N. Synthesis and Deposition of Callose in Anthers and Ovules of Meiotic Mutants of Maize (Zea mays). Russ. J. Plant Physiol. 2003, 50, 324–329. [Google Scholar] [CrossRef]
- Musiał, K.; Kościńska-Pająk, M.; Antolec, R.; Joachimiak, A.J. Deposition of callose in young ovules of two Taraxacum species varying in the mode of reproduction. Protoplasma 2015, 252, 135–144. [Google Scholar] [CrossRef] [PubMed]
- Chen, R.; Zhao, X.; Shao, Z.; Wei, Z.; Wang, Y.; Zhu, L.; Zhao, J.; Sun, M.; He, R.; He, G. Rice UDP-glucose pyrophosphorylase1 is essential for pollen callose deposition and its cosuppression results in a new type of thermosensitive genic male sterility. Plant Cell 2007, 19, 847–861. [Google Scholar] [CrossRef]
- Khan, R.M.; Yu, P.; Sun, L.; Abbas, A.; Shah, L.; Xiang, X.; Wang, D.; Sohail, A.; Zhang, Y.; Liu, Q.; et al. DCET1 Controls Male Sterility Through Callose Regulation, Exine Formation, and Tapetal Programmed Cell Death in Rice. Front. Genet. 2021, 12, 790789. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.J.; Nan, G.L.; Kelliher, T.; Timofejeva, L.; Vernoud, V.; Golubovskaya, I.N.; Harper, L.; Egger, R.; Walbot, V.; Cande, W.Z. Maize multiple archesporial cells 1 (mac1), an ortholog of rice TDL1A, modulates cell proliferation and identity in early anther development. Development 2012, 139, 2594–2603. [Google Scholar] [CrossRef]
- Cai, G.; Faleri, C.; Del Casino, C.; Emons, A.M.; Cresti, M. Distribution of callose synthase, cellulose synthase, and sucrose synthase in tobacco pollen tube is controlled in dissimilar ways by actin filaments and microtubules. Plant Physiol. 2011, 155, 1169–1190. [Google Scholar] [CrossRef]
- Parton, R.M.; Fischer-Parton, S.; Watahiki, M.K.; Trewavas, A.J. Dynamics of the apical vesicle accumulation and the rate of growth are related in individual pollen tubes. J. Cell Sci. 2001, 114, 2685–2695. [Google Scholar] [CrossRef]
- Ketelaar, T.; Galway, M.E.; Mulder, B.M.; Emons, A.M.C. Rates of exocytosis and endocytosis in Arabidopsis root hairs and pollen tubes. J. Microsc. 2008, 231, 265–273. [Google Scholar] [CrossRef]
- Bove, J.; Vaillancourt, B.; Kroeger, J.; Hepler, P.K.; Wiseman, P.W.; Geitmann, A. Magnitude and direction of vesicle dynamics in growing pollen tubes using spatiotemporal image correlation spectroscopy and fluorescence recovery after photobleaching. Plant Physiol. 2008, 147, 1646–1658. [Google Scholar] [CrossRef]
- Vidali, L.; Hepler, P.K. Actin and pollen tube growth. Protoplasma 2001, 215, 64–76. [Google Scholar] [CrossRef]
- Xu, Y.; Huang, S. Control of the Actin Cytoskeleton Within Apical and Subapical Regions of Pollen Tubes. Front. Cell Dev. Biol. 2020, 8, 614821. [Google Scholar] [CrossRef]
- Ge, L.L.; Xie, C.; Wei, D.M.; Qiu, Y.; Tian, H.Q. Calcium Distribution during Anther Development of Tobacco. Int. J. Plant Dev. Biol. 2007, 1, 287–293. [Google Scholar]
- Kulich, I.; Vojtíková, Z.; Sabol, P.; Ortmannová, J.; Neděla, V.; Tihlaříková, E.; Žárský, V. Exocyst Subunit EXO70H4 Has a Specific Role in Callose Synthase Secretion and Silica Accumulation. Plant Physiol. 2018, 176, 2040–2051. [Google Scholar] [CrossRef]
- Clément, C.; Audran, J.C. Anther wall layers control pollen sugar nutrition in Lilium. Protoplasma 1995, 187, 172–181. [Google Scholar] [CrossRef]
- Dorion, S.; Lalonde, S.; Saini, H.S. Induction of Male Sterility in Wheat by Meiotic-Stage Water Deficit Is Preceded by a Decline in Invertase Activity and Changes in Carbohydrate Metabolism in Anthers. Plant Physiol. 1996, 111, 137–145. [Google Scholar] [CrossRef]
- Weber, H.; Borisjuk, L.; Heim, U.; Buchner, P.; Wobus, U. Seed-coat associated invertases of fava bean control both unloading and storage functions: Cloning of cDNAs and cell type-specific expression. Plant Cell 1995, 7, 1835–1846. [Google Scholar] [PubMed]
- Amor, Y.; Haigler, C.H.; Johnson, S.; Wainscott, M.; Delmer, D.P. A membrane-associated form of sucrose synthase and its potential role in synthesis of cellulose and callose in plants. Proc. Natl. Acad. Sci. USA 1995, 92, 9353–9357. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, N.; Shimizu, M.; Suzuki, H. Characterization of hexose kinases from camellia and lily pollen grains. Physiol. Plant 1991, 81, 215–220. [Google Scholar] [CrossRef]
- Wang, B.; Fang, R.; Zhang, J.; Han, J.; Chen, F.; He, F.; Liu, Y.G.; Chen, L. Rice LecRK5 phosphorylates a UGPase to regulate callose biosynthesis during pollen development. J. Exp. Bot. 2020, 71, 4033–4041. [Google Scholar] [CrossRef]
- Hong, Z.; Zhang, Z.; Olson, J.M.; Verma, D.P. A novel UDP-glucose transferase is part of the callose synthase complex and interacts with phragmoplastin at the forming cell plate. Plant Cell 2001, 13, 769–779. [Google Scholar] [CrossRef]
- Eggert, D.; Naumann, M.; Reimer, R. Nanoscale glucan polymer network causes pathogen resistance. Sci. Rep. 2014, 4, 4159. [Google Scholar] [CrossRef] [PubMed]
- Harada, T.; Misaki, A.; Saito, H. Curdlan: A bacterial gel-forming beta-1,3-glucan. Arch. Biochem. Biophys. 1968, 124, 292–298. [Google Scholar] [CrossRef]
- Saitô, H.; Ohki, T.; Sasaki, T. A 13C nuclear magnetic resonance study of gel-forming (1 goes to 3)-beta-d-glucans. Evidence of the presence of single-helical conformation in a resilient gel of a curdlan-type polysaccharide 13140 from Alcaligenes faecalis var. myxogenes IFO 13140. Biochemistry 1977, 16, 908–914. [Google Scholar] [CrossRef]
- Abou-Saleh, R.H.; Hernandez-Gomez, M.C.; Amsbury, S. Interactions between callose and cellulose revealed through the analysis of biopolymer mixtures. Nat. Commun. 2018, 9, 4538. [Google Scholar] [CrossRef] [PubMed]
- An, Q.; Hückelhoven, R.; Kogel, K.H.; Bel, A.J. Multivesicular bodies participate in a cell wall-associated defence response in barley leaves attacked by the pathogenic powdery mildew fungus. Cell Microbiol. 2006, 8, 1009–1019. [Google Scholar] [CrossRef] [PubMed]
- Voigt, C.A. Callose-mediated resistance to pathogenic intruders in plant defense-related papillae. Front. Plant Sci. 2014, 5, 168. [Google Scholar] [CrossRef] [PubMed]
- Sevilem, I.; Miyashima, S.; Helariutta, Y. Cell-to-cell communication via plasmodesmata in vascular plants. Cell Adhes. Migr. 2013, 7, 27–32. [Google Scholar] [CrossRef] [PubMed]
- Mamun, E.; Cantrill, L.; Overall, R.; Sutton, B. Cellular organisation and differentiation of organelles in pre-meiotic rice anthers. Cell Biol. Int. 2005, 29, 792–802. [Google Scholar] [CrossRef] [PubMed]
- Mursalimov, S.R.; Baiborodin, S.I.; Sidorchuk, Y.V.; Shumny, V.K.; Deineko, E.V. Characteristics of the cytomictic channel formation in Nicotiana tabacum L. pollen mother cells. Cytol. Genet. 2010, 44, 14–18. [Google Scholar] [CrossRef]
- Mursalimov, S.R.; Sidorchuk, Y.V.; Deineko, E.V. New insights into cytomixis: Specific cellular features and prevalence in higher plants. Planta 2013, 238, 415–423. [Google Scholar] [CrossRef]
- Li, W.; Yang, J.; Pan, Y.F.; Guo, G.Q.; Zheng, G.C. Chromosome localization of genes that control synchronous development of pollen mother cells in wheat. Caryologia 2003, 56, 275–279. [Google Scholar] [CrossRef]
- Li, W.; Godzik, A. Cd-hit: A fast program for clustering and comparing large sets of protein or nucleotide sequences. Bioinformatics 2006, 22, 1658–1659. [Google Scholar] [CrossRef] [PubMed]
- Katoh, K.; Standley, D.M. MAFFT multiple sequence alignment software version 7: Improvements in performance and usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef]
- Capella-Gutiérrez, S.; Silla-Martínez, J.M.; Gabaldón, T. trimAl: A tool for automated alignment trimming in large-scale phylogenetic analyses. Bioinformatics 2009, 25, 1972–1973. [Google Scholar] [CrossRef]
- Nguyen, L.T.; Schmidt, H.A.; von Haeseler, A.; Minh, B.Q. IQ-TREE: A fast and effective stochastic algorithm for estimating maximum-likelihood phylogenies. Mol. Biol. Evol. 2015, 32, 268–274. [Google Scholar] [CrossRef]
- Periasamy, K.; Amalathas, J. Absence of Callose and Tetrad in the Microsporogenesis of Pandanus Odoratissimus with Well-Formed Pollen Exine. Ann. Bot. 1991, 67, 29–33. Available online: https://www.jstor.org/stable/42758399 (accessed on 20 January 2023). [CrossRef]
- Vijayaraghavan, M.R.; Shukla, A.K. Absence of Callose Around the Microspore Tetrad and Poorly Developed Exine in Pergularia Daemia. Ann. Bot. 1977, 41, 923–926. [Google Scholar] [CrossRef]
- Ford, J.H. Ultrastructural and Chemical Studies of Pollen Wall in Epacridaceae. In Sporopollenin; Brooks, J., Grant, P.R., Muir, M.P., van Gijzel, P., Shaw, G., Eds.; Academic Press: London, UK, 1971; pp. 130–173. [Google Scholar]
- Naumova, T.; Den Nijs, A.P.M.; Willemse, M.T.M. Quantitative analysis of aposporous parthenogenesis in Poa pratensis genotypes. Acta Bot. Neerl. 1993, 42, 299–312. [Google Scholar] [CrossRef]
- Carman, J.G.; Crane, C.F.; Riera-Lizarazu, O. Comparative histology of cell walls during meiotic and apomeiotic megasporogenesis in two hexaploid Australasian elymus species. Crop Sci. 1991, 31, 1527–1532. [Google Scholar] [CrossRef]
- Leblanc, O.; Peel, M.D.; Carman, J.G.; Savidan, Y. Megasporogenesis and Megagametogenesis in Several Tripsacum Species (Poaceae). Am. J. Bot. 1995, 82, 57–63. [Google Scholar] [CrossRef]
- Mimura, M.; Ono, S.; Nonomura, K.I. Rice MEL2 Regulates the Timing of Meiotic Transition as a Component of Cytoplasmic RNA Granules. bioRxiv 2021. [Google Scholar] [CrossRef]
- Wang, B.; Andargie, M.; Fang, R. The function and biosynthesis of callose in high plants. Heliyon 2022, 8, 09248. [Google Scholar] [CrossRef]
- Hörandl, E.; Hadacek, F. The oxidative damage initiation hypothesis for meiosis. Plant Reprod. 2013, 26, 351–367. [Google Scholar] [CrossRef] [PubMed]
- Sankaranarayanan, S.; Ju, Y.; Kessler, S.A. Reactive Oxygen Species as Mediators of Gametophyte Development and Double Fertilization in Flowering Plants. Front. Plant Sci. 2020, 11, 1199. [Google Scholar] [CrossRef] [PubMed]
- Yu, J.; Zhang, D. Molecular Control of Redox Homoeostasis in Specifying the Cell Identity of Tapetal and Microsporocyte Cells in Rice. Rice 2019, 12, 42. [Google Scholar] [CrossRef] [PubMed]
- Chaubal, R.; Anderson, J.R.; Trimnell, M.R.; Fox, T.W.; Albertsen, M.C.; Bedinger, P. The transformation of anthers in the msca1 mutant of maize. Planta 2003, 216, 778–788. [Google Scholar] [CrossRef]
- Xing, S.; Zachgo, S. ROXY1 and ROXY2, two Arabidopsis glutaredoxin genes, are required for anther development. Plant J. 2008, 53, 790–801. [Google Scholar] [CrossRef]
- Jung, J.Y.; Ahn, J.H.; Schachtman, D.P. CC-type glutaredoxins mediate plant response and signaling under nitrate starvation in Arabidopsis. BMC Plant Biol. 2018, 18, 281. [Google Scholar] [CrossRef]
- Jaeger-Braet, J.; Krause, L.; Buchholz, A.; Schnittger, A. Heat stress reveals a specialized variant of the pachytene checkpoint in meiosis of Arabidopsis thaliana. Plant Cell 2022, 34, 433–454. [Google Scholar] [CrossRef]
- Zhou, Q.; Cheng, X.; Kong, B.; Zhao, Y.; Li, Z.; Sang, Y.; Wu, J.; Zhang, P. Heat shock-induced failure of meiosis I to meiosis II transition leads to 2n pollen formation in a woody plant. Plant Physiol. 2022, 189, 2110–2127. [Google Scholar] [CrossRef]
- Satake, T.; Hayase, H. Male sterility caused by cooling treatment at the young microspore stage in rice plants. Proc. Crop Sci. Soc. Jpn. 1974, 43, 36–39. [Google Scholar] [CrossRef][Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Plant | Protein Type | Meiosis Progression | Expression Site/Stage | Refs. |
|---|---|---|---|---|---|
| MIL1 | Rice | Plant-specific CC-type glutaredoxin | Sporogenous cells fail to enter into meiosis | Sporogenous cells and inner parietal cells | [36] |
| AM1 | Maize | Coiled-Coiled domain protein | Failure in mitosis-meiosis switch and prophase I progression in male and female | Male meiocytes. Localize on chromatin and pericentromeres | [57,58] |
| SWI/DYAD | Arabidopsis | WINGS-APART LIKE (WAPL) inhibitor | Fail to switch female meiosis. Male meiosis is initiated but abnormal | Male and female meiocytes at G1/S phase and prophase I | [59,60] |
| MEL2 | Rice | RNA binding protein | Asynchronous meiosis initiation and germ cells arrest at leptotene/zygotene | Male and female meiocytes at G1/S/G2 and phase | [67] |
| OsRR24/LEPTO1 | Rice | Type-B response regulator | Leptotene/Zygotene transition | Early meiosis PMCs | [68] |
| SDS | Arabidopsis | Plant specific cyclin | Reduced frequency of pachytene I stage. Defects in synapses and sister chromatid separation | Male and female meiocytes | [55] |
| TAM/CYCA1;2 | Arabidopsis | Cyclin A | Delayed pachytene I and longer meiosis II. Asynchrony from diplotene to tetrad | Male meiocyte at pachytene I stage | [46,53,54] |
| MLH3 | Arabidopsis | DNA repair protein | Prophase I prolonged for ~25 h | Expressed in buds. Localize to chromosome axes | [70] |
| DUET/MMD1 | Arabidopsis | PHD finger transcription factor | Prolonged, Metaphase I | Sporogenous and tapetum cells at premeiosis and meiosis stage | [64] |
| SAP | Arabidopsis | Putative transcription factor | Female germ cells fail to transit into meiosis II | Floral meristem, influorescence and young ovules | [71] |
| FST | Arabidopsis | bHLH transcription factor | Early entry and asynchrony in meiosis | Male meiocytes nuclear localized | [55] |
| GSL5 | Rice | Callose synthase | Early entry and defects in meiosis progression | Male meiocytes at premeiosis, meiosis and pollen stage | [72,73] |
| MEZ1 | Petunia | Putative transcription factor | Abnormal meiosis progression | Premeiotic anther stage | [66] |
| Histone H1A, H1B | Tobacco | Linker histone | Asynchronous meiosis II in male germ cells. Effect on female meiosis is unknown | Universal expression | [69] |
| Plant | Family | Callose Deficit Stage/Organ | Meiosis Fate | Refs. |
|---|---|---|---|---|
| Lilium candidum | Liliaceae | In apomeiosis at MMC side walls and cross walls | Altered meiosis | [77] |
| Lilium regale | Liliaceae | In apomeiosis at MMC side walls and cross walls | Altered meiosis | [77] |
| Tulipa spp. | Liliaceae | In apomeiosis at MMC side walls and cross walls | Altered meiosis | [77] |
| Pandanus odoratissimus | Pandanaceae | Throughout male meiosis in anthers | Monads instead of tetrads. Microspores are normal | [120] |
| Pergularia daemia | Apocynaceae | Meiosis, tetrad and microspore stage in anthers | Defects in exine wall formation on pollen grain surface | [121] |
| Styphelia spp. | Epacridaceae | Tetrad (Very weak callose deposition is seen) in anthers | Fragmentaed exine wall formation | [122] |
| Poa nemoralis | Poaceae/Graminaeae | During apomeiosis in MMC micropylar region | Apospory due to abnormal meiosis | [123] |
| Elymus restisetus | Poaceae/Graminaeae | During apomeiosis around MMC | Death of some 2n megaspores/binucleate female gametophyte | [124] |
| Tripsacum dactyloides | Poaceae/Graminaeae | During apomeiosis around Megasporocytes | Diplospory due to failure of meiosis | [125] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Somashekar, H.; Nonomura, K.-I. Genetic Regulation of Mitosis–Meiosis Fate Decision in Plants: Is Callose an Oversighted Polysaccharide in These Processes? Plants 2023, 12, 1936. https://doi.org/10.3390/plants12101936
Somashekar H, Nonomura K-I. Genetic Regulation of Mitosis–Meiosis Fate Decision in Plants: Is Callose an Oversighted Polysaccharide in These Processes? Plants. 2023; 12(10):1936. https://doi.org/10.3390/plants12101936
Chicago/Turabian StyleSomashekar, Harsha, and Ken-Ichi Nonomura. 2023. "Genetic Regulation of Mitosis–Meiosis Fate Decision in Plants: Is Callose an Oversighted Polysaccharide in These Processes?" Plants 12, no. 10: 1936. https://doi.org/10.3390/plants12101936
APA StyleSomashekar, H., & Nonomura, K.-I. (2023). Genetic Regulation of Mitosis–Meiosis Fate Decision in Plants: Is Callose an Oversighted Polysaccharide in These Processes? Plants, 12(10), 1936. https://doi.org/10.3390/plants12101936