

Colored LED Lights: Use One Color Alone or with Others for Growth in Hedyotis corymbosa In Vitro?

 ,
,  , , , , ,
, , , , ,  and
and
Abstract
1. Introduction
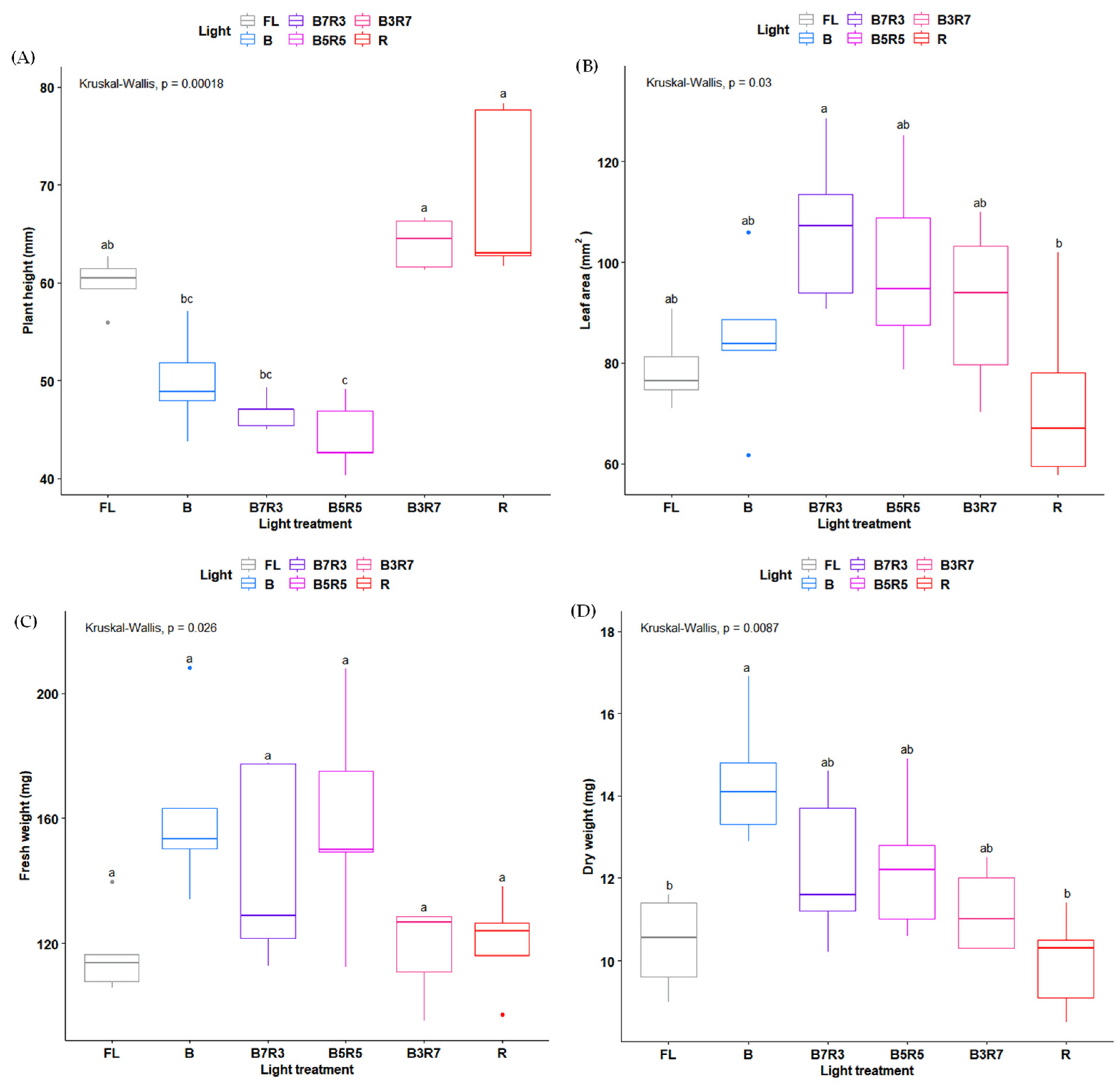
2. Results
3. Discussion
4. Materials and Methods
4.1. Plant Material and Growth Conditions
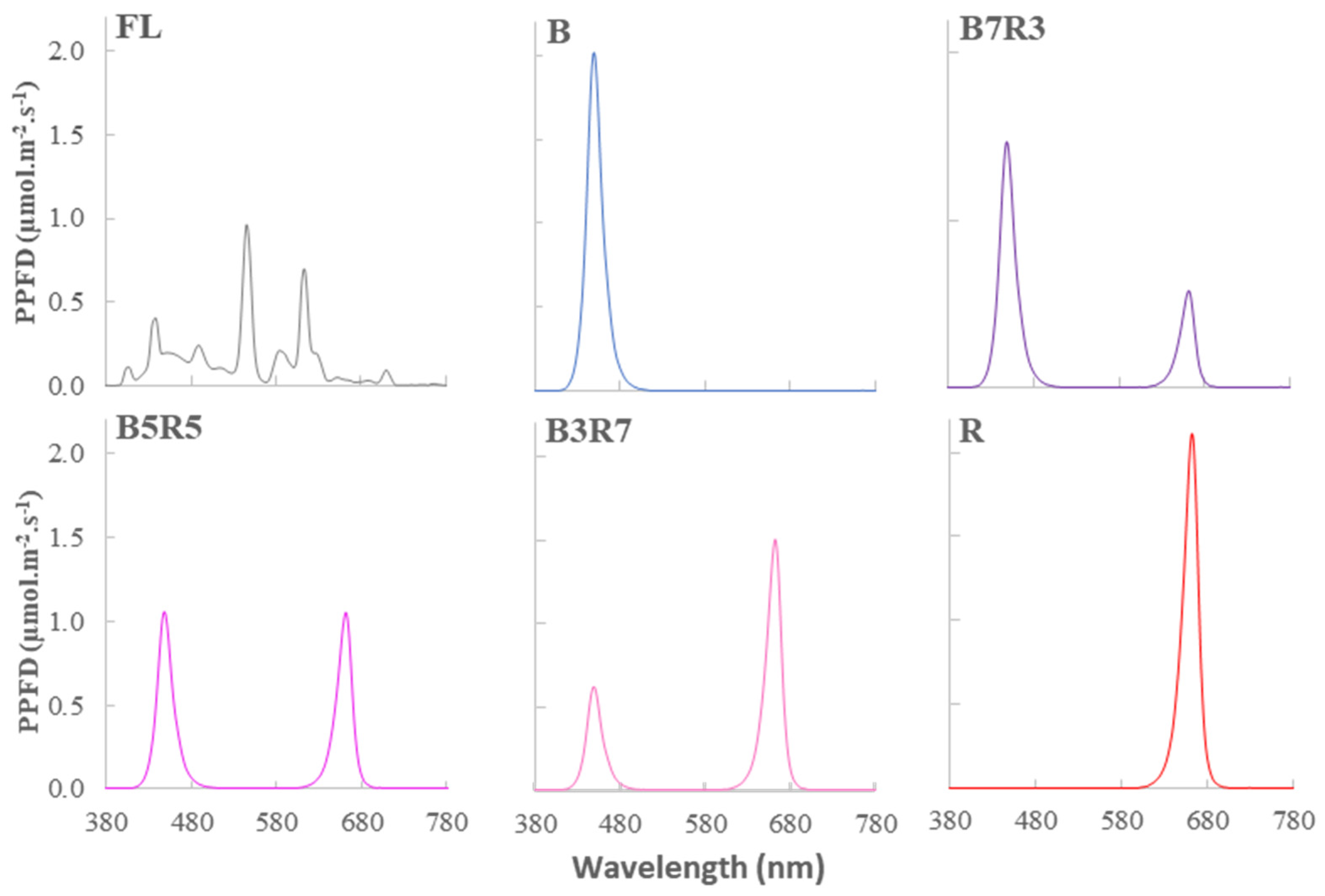
4.2. Light Treatment
- White fluorescent lamp (FL);
- 100% blue (B);
- 70% blue and 30% red (B7R3);
- 50% blue and 50% red (B5R5);
- 30% blue and 70% red (B3R7);
- 100% red (R).
4.3. Measurements
4.4. Determination of the Triterpenoid Content
4.5. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kozai, T.; Niu, G.; Takagaki, M. Plant Factory: An Indoor Vertical Farming System for Efficient Quality Food Production; Academic Press: Cambridge, MA, USA, 2015; ISBN 9780128017753. [Google Scholar]
- Dueck, T.; van Ieperen, W.; Taulavuori, K. Light Perception, Signalling and Plant Responses to Spectral Quality and Photoperiod in Natural and Horticultural Environments. Environ. Exp. Bot. 2016, 121, 1–3. [Google Scholar] [CrossRef]
- Lee, M.J.; Son, K.H.; Oh, M.M. Increase in Biomass and Bioactive Compounds in Lettuce under Various Ratios of Red to Far-Red LED Light Supplemented with Blue LED Light. Hortic. Environ. Biotechnol. 2016, 57, 139–147. [Google Scholar] [CrossRef]
- Dłużniewska, J.; Klimek-Kopyra, A.; Czech, T.; Dobrowolski, J.W.; Dacewicz, E. The Use of Coherent Laser Stimulation of Seeds and a Fungal Inoculum to Increase the Productivity and Health of Soybean Plants. Agronomy 2021, 11, 1923. [Google Scholar] [CrossRef]
- Jung, W.S.; Chung, I.M.; Hwang, M.H.; Kim, S.H.; Yu, C.Y.; Ghimire, B.K. Application of Light-Emitting Diodes for Improving the Nutritional Quality and Bioactive Compound Levels of Some Crops and Medicinal Plants. Molecules 2021, 26, 1477. [Google Scholar] [CrossRef] [PubMed]
- Viršile, A.; Brazaityte, A.; Sirtautas, R.; Duchovskis, P. Light Spectral Effects on Phenolic Compounds in Perilla frutescens Leaves as Related to the Leaf Age, Color and Duration of Exposure. Acta Hortic. 2017, 1170, 981–988. [Google Scholar] [CrossRef]
- Hasan, M.M.; Bashir, T.; Ghosh, R.; Lee, S.K.; Bae, H. An Overview of LEDs’ Effects on the Production of Bioactive Compounds and Crop Quality. Molecules 2017, 22, 1420. [Google Scholar] [CrossRef] [PubMed]
- Darko, E.; Heydarizadeh, P.; Schoefs, B.; Sabzalian, M.R. Photosynthesis under Artificial Light: The Shift in Primary and Secondary Metabolism. Philos. Trans. R. Soc. B 2014, 369, 20130243. [Google Scholar] [CrossRef]
- McCree, K.J. The Action Spectrum, Absorptance and Quantum Yield of Photosynthesis in Crop Plants. Agric. Meteorol. 1971, 9, 191–216. [Google Scholar] [CrossRef]
- Rabinowitch, E.I. ; Govindjee The Role of Chlorophyll in Photosynthesis. Sci. Am. 1965, 213, 74–83. [Google Scholar] [CrossRef]
- Hu, J.; Dai, X.; Sun, G. Morphological and Physiological Responses of Morus alba Seedlings under Different Light Qualities. Not. Bot. Horti Agrobot. Cluj-Napoca 2016, 44, 382–392. [Google Scholar] [CrossRef]
- Kong, Y.; Nemali, K. Blue and Far-Red Light Affect Area and Number of Individual Leaves to Influence Vegetative Growth and Pigment Synthesis in Lettuce. Front. Plant Sci. 2021, 12, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Van Iersel, M.W. Photosynthetic Physiology of Blue, Green, and Red Light: Light Intensity Effects and Underlying Mechanisms. Front. Plant Sci. 2021, 12, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Meng, X.; Wang, Z.; He, S.; Shi, L.; Song, Y.; Lou, X.; He, D. LED-Supplied Red and Blue Light Alters the Growth, Antioxidant Status, and Photochemical Potential of in Vitro-Grown Gerbera jamesonii Plantlets. Hortic. Sci. Technol. 2019, 37, 473–489. [Google Scholar] [CrossRef]
- Miao, Y.-x.; Wang, X.-z.; Gao, L.-h.; Chen, Q.-y.; Qu, M. Blue Light Is More Essential than Red Light for Maintaining the Activities of Photosystem II and I and Photosynthetic Electron Transport Capacity in Cucumber Leaves. J. Integr. Agric. 2016, 15, 87–100. [Google Scholar] [CrossRef]
- Hogewoning, S.W.; Trouwborst, G.; Maljaars, H.; Poorter, H.; van Ieperen, W.; Harbinson, J. Blue Light Dose-Responses of Leaf Photosynthesis, Morphology, and Chemical Composition of Cucumis sativus Grown under Different Combinations of Red and Blue Light. J. Exp. Bot. 2010, 61, 3107–3117. [Google Scholar] [CrossRef]
- Ouzounis, T.; Fretté, X.; Ottosen, C.O.; Rosenqvist, E. Spectral Effects of LEDs on Chlorophyll Fluorescence and Pigmentation in Phalaenopsis “Vivien” and “Purple star”. Physiol. Plant. 2015, 154, 314–327. [Google Scholar] [CrossRef]
- Cioć, M.; Szewczyk, A.; Żupnik, M.; Kalisz, A.; Pawłowska, B. LED Lighting Affects Plant Growth, Morphogenesis and Phytochemical Contents of Myrtus communis L. in Vitro. Plant Cell. Tissue Organ. Cult. 2018, 132, 433–447. [Google Scholar] [CrossRef]
- Ramírez-Mosqueda, M.A.; Iglesias-Andreu, L.G.; Luna-Sánchez, I.J. Light Quality Affects Growth and Development of in Vitro Plantlet of Vanilla planifolia Jacks. S. Afr. J. Bot. 2017, 109, 288–293. [Google Scholar] [CrossRef]
- Lewis, W.H. Oldenlandia corymbosa (Rubiaceae). Grana Palynol. 1964, 5, 330–341. [Google Scholar] [CrossRef]
- Gupta, R.K.; Singh, R.K.; Swain, S.R.; Hussain, T.; Rao, C.V. Anti-Hepatotoxic Potential of Hedyotis corymbosa against D-Galactosamine-Induced Hepatopathy in Experimental Rodents. Asian Pac. J. Trop. Biomed. 2012, 2, 1542–1547. [Google Scholar] [CrossRef]
- Andriyani, R.; Risdian, C.; Udin, Z. Cytotoxicity Assay From Fractions of Hedyotis corymbosa Extract Against Breast Cancer Cell Line T47D. Indones. J. Cancer Chemoprevention 2011, 2, 182–186. [Google Scholar] [CrossRef][Green Version]
- Sultana, T.; Rashid, M.A.; Ali, M.A.; Mahmood, S.F. Hepatoprotective and Antibacterial Activity of Ursolic Acid Extracted from Hedyotis corymbosa (L.). Bangladesh J. Sci. Ind. Res. 2010, 45, 27–34. [Google Scholar] [CrossRef]
- Furtado, N.A.J.C.; Pirson, L.; Edelberg, H.; Miranda, L.M.; Loira-Pastoriza, C.; Preat, V.; Larondelle, Y.; André, C.M. Pentacyclic Triterpene Bioavailability: An Overview of in Vitro and in Vivo Studies. Molecules 2017, 22, 400. [Google Scholar] [CrossRef] [PubMed]
- Mlala, S.; Oyedeji, A.O.; Gondwe, M.; Oyedeji, O.O. Ursolic Acid and Its Derivatives as Bioactive Agents. Molecules 2019, 24, 2751. [Google Scholar] [CrossRef] [PubMed]
- Laszczyk, M.N. Pentacyclic Triterpenes of the Lupane, Oleanane and Ursane Group as Tools in Cancer Therapy. Planta Med. 2009, 75, 1549–1560. [Google Scholar] [CrossRef]
- Le, A.T.; Phan, N.H.; Do, T.K. Effect of Blue Light on the Photosynthesis and Flavonoid Accumulation in Leaves of Hedyotis corymbosa (L.) Lam. Plant Sci. Today 2021, 8, 955–962. [Google Scholar] [CrossRef]
- Le, A.T.; Yu, J.K.; Han, G.D.; Do, T.K.; Chung, Y.S. Potential Use of Colored LED Lights to Increase the Production of Bioactive Metabolites Hedyotis corymbosa (L.) Lam. Plants 2022, 11, 225. [Google Scholar] [CrossRef]
- Le, A.T.; Hoang, T.T.T.; Phan, N.H. Development of Shoots of Hedyotis corymbosa (L.) Lam. in Vitro Culture. Sci. Technol. Dev. J. 2015, 5, 75–84. [Google Scholar] [CrossRef]
- Zheng, L.; Van Labeke, M.C. Long-Term Effects of Red- and Blue-Light Emitting Diodes on Leaf Anatomy and Photosynthetic Efficiency of Three Ornamental Pot Plants. Front. Plant Sci. 2017, 8, 917. [Google Scholar] [CrossRef]
- Miao, Y.; Chen, Q.; Qu, M.; Gao, L.; Hou, L. Blue Light Alleviates ‘Red Light Syndrome’ by Regulating Chloroplast Ultrastructure, Photosynthetic Traits and Nutrient Accumulation in Cucumber Plants. Sci. Hortic. 2019, 257, 108680. [Google Scholar] [CrossRef]
- Xu, Y.; Liang, Y.; Yang, M. Effects of Composite LED Light on Root Growth and Antioxidant Capacity of Cunninghamia lanceolata Tissue Culture Seedlings. Sci. Rep. 2019, 9, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Zhou, L.; Wu, S.; Liu, L.; Huang, M.; Lin, S.; Ding, G. Effects of LED Light on Acacia melanoxylon Bud Proliferation in Vitro and Root Growth Ex Vitro. Open Life Sci. 2019, 14, 349–357. [Google Scholar] [CrossRef] [PubMed]
- Yuan, Z.; Cao, Q.; Zhang, K.; Ata-Ul-Karim, S.T.; Tan, Y.; Zhu, Y.; Cao, W.; Liu, X. Optimal Leaf Positions for SPAD Meter Measurement in Rice. Front. Plant Sci. 2016, 7, 719. [Google Scholar] [CrossRef] [PubMed]
- Parry, C.; Blonquist, J.M.; Bugbee, B. In Situ Measurement of Leaf Chlorophyll Concentration: Analysis of the Optical/Absolute Relationship. Plant Cell Environ. 2014, 37, 2508–2520. [Google Scholar] [CrossRef] [PubMed]
- Chen, M.; Blankenship, R.E. Expanding the Solar Spectrum Used by Photosynthesis. Trends Plant Sci. 2011, 16, 427–431. [Google Scholar] [CrossRef]
- Sakuraba, Y.; Yokono, M.; Akimoto, S.; Tanaka, R.; Tanaka, A. Deregulated Chlorophyll b Synthesis Reduces the Energy Transfer Rate between Photosynthetic Pigments and Induces Photodamage in Arabidopsis thaliana. Plant Cell Physiol. 2010, 51, 1055–1065. [Google Scholar] [CrossRef]
- Bukhov, N.G. Dynamic Light Regulation of Photosynthesis (a Review). Russ. J. Plant Physiol. 2004, 51, 742–753. [Google Scholar] [CrossRef]
- Baker, N.R. Chlorophyll Fluorescence: A Probe of Photosynthesis in Vivo. Annu. Rev. Plant Biol. 2008, 59, 89–113. [Google Scholar] [CrossRef]
- Kalaji, H.M.; Schansker, G.; Ladle, R.J.; Goltsev, V.; Bosa, K.; Allakhverdiev, S.I.; Brestic, M.; Bussotti, F.; Calatayud, A.; Dąbrowski, P.; et al. Frequently Asked Questions about in Vivo Chlorophyll Fluorescence: Practical Issues. Photosynth. Res. 2014, 122, 121–158. [Google Scholar] [CrossRef]
- Wang, Z.-H.; Wu, X.-S.; Chang, X.-P.; Li, R.-Z.; Jing, R.-L. Chlorophyll Content and Chlorophyll Fluorescence Kinetics Parameters of Flag Leaf and Their Gray Relational Grade with Yield in Wheat. Acta Agron. Sin. 2010, 36, 217–227. [Google Scholar] [CrossRef]
- Rochaix, J.D. Regulation of Photosynthetic Electron Transport. Biochim. Biophys. Acta—Bioenerg. 2011, 1807, 375–383. [Google Scholar] [CrossRef] [PubMed]
- Yin, J.; Ma, H.; Gong, Y.; Xiao, J.; Jiang, L.; Zhan, Y.; Li, C.; Ren, C.; Yang, Y. Effect of MeJA and Light on the Accumulation of Betulin and Oleanolic Acid in the Saplings of White Birch (Betula platyphylla Suk.). Am. J. Plant Sci. 2013, 4, 7–15. [Google Scholar] [CrossRef]
- Aminfar, Z.; Rabiei, B.; Tohidfar, M.; Mirjalili, M.H. Identification of Key Genes Involved in the Biosynthesis of Triterpenic Acids in the Mint Family. Sci. Rep. 2019, 9, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Nawae, W.; Yoocha, T.; Narong, N.; Paemanee, A.; Ketngamkum, Y.; Romyanon, K.; Toojinda, T.; Tangphatsornruang, S.; Pootakham, W. Transcriptome Sequencing Revealed the Influence of Blue Light on the Expression Levels of Light-Stress Response Genes in Centella asiatica. PLoS ONE 2021, 16, e0260468. [Google Scholar] [CrossRef] [PubMed]
- Murashige, T.; Skoog, F. A Revised Medium for Rapid Growth and Bioassay with Tobacco Tissue Cultures. Physiol. Plant 1962, 15, 473–497. [Google Scholar] [CrossRef]
- Henselová, M.; Regecová, M.; Sováková, A. Isolation of Chloroplasts in the Karwinskia Species and Determination of Their Photochemical Activity under in Vitro Conditions. Plant Soil Environ. 2004, 50, 149–156. [Google Scholar] [CrossRef]
- Li, J.; Zu, Y.G.; Fu, Y.J.; Yang, Y.C.; Li, S.M.; Li, Z.N.; Wink, M. Optimization of Microwave-Assisted Extraction of Triterpene Saponins from Defatted Residue of Yellow Horn (Xanthoceras sorbifolia Bunge.) Kernel and Evaluation of Its Antioxidant Activity. Innov. Food Sci. Emerg. Technol. 2010, 11, 637–643. [Google Scholar] [CrossRef]
- Cai, C.; Ma, J.; Han, C.; Jin, Y.; Zhao, G.; He, X. Extraction and Antioxidant Activity of Total Triterpenoids in the Mycelium of a Medicinal Fungus, Sanghuangporus sanghuang. Sci. Rep. 2019, 9, 7418. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Light Source | Fv/Fm | Y(II) | Y(NPQ) | Y(NO) | qP | qN | ETR (μmol Electron·m−2·s−1) |
|---|---|---|---|---|---|---|---|
| FL | 0.749 a | 0.666 a | 0. 037 a | 0.312 b | 0.801 bc | 0.311 a | 12.179 ab |
| B | 0.753 a | 0.623 a | 0.075 a | 0.302 b | 0.875 ab | 0.273 a | 13.259 a |
| B7R3 | 0.739 ab | 0.663 a | 0.043 a | 0.289 b | 0.957 a | 0.154 a | 14.159 a |
| B5R5 | 0.737 ab | 0.662 a | 0.028 a | 0.309 b | 0.924 ab | 0.147 a | 13.902 a |
| B3R7 | 0.681 b | 0.510 b | 0.074 a | 0.360 a | 0.768 bc | 0.271 a | 9.849 b |
| R | 0.694 b | 0.420 b | 0.095 a | 0.396 a | 0.734 c | 0.267 a | 8.978 b |
| Light Source | SPAD Value | Hill Reaction Activity (nmol DCIP.Million of Chloroplast−1·min−1) |
|---|---|---|
| FL | 26.2 a | 0.087 a |
| B | 23.9 b | 0.085 ab |
| R | 19.7 c | 0.076 b |
| Light Source | Total Triterpenoid (mg UAE/g DW) |
|---|---|
| FL | 27.03 a |
| B | 26.33 a |
| R | 18.41 b |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Le, A.T.; Choi, I.-L.; Han, G.-D.; Kang, H.-M.; Jung, D.H.; Park, W.-P.; Yildiz, M.; Do, T.K.; Chung, Y.S. Colored LED Lights: Use One Color Alone or with Others for Growth in Hedyotis corymbosa In Vitro? Plants 2023, 12, 93. https://doi.org/10.3390/plants12010093
Le AT, Choi I-L, Han G-D, Kang H-M, Jung DH, Park W-P, Yildiz M, Do TK, Chung YS. Colored LED Lights: Use One Color Alone or with Others for Growth in Hedyotis corymbosa In Vitro? Plants. 2023; 12(1):93. https://doi.org/10.3390/plants12010093
Chicago/Turabian StyleLe, Anh Tuan, In-Lee Choi, Gyung-Deok Han, Ho-Min Kang, Dae Ho Jung, Won-Pyo Park, Mehtap Yildiz, Thuong Kiet Do, and Yong Suk Chung. 2023. "Colored LED Lights: Use One Color Alone or with Others for Growth in Hedyotis corymbosa In Vitro?" Plants 12, no. 1: 93. https://doi.org/10.3390/plants12010093
APA StyleLe, A. T., Choi, I.-L., Han, G.-D., Kang, H.-M., Jung, D. H., Park, W.-P., Yildiz, M., Do, T. K., & Chung, Y. S. (2023). Colored LED Lights: Use One Color Alone or with Others for Growth in Hedyotis corymbosa In Vitro? Plants, 12(1), 93. https://doi.org/10.3390/plants12010093