

Comparative Analysis of the Genetic Diversity of Chilean Cultivated Potato Based on a Molecular Study of Authentic Herbarium Specimens and Present-Day Gene Bank Accessions

 , , , and
, , , and
Abstract
1. Introduction
1.1. Historical Concepts of the Origin of Chilean Cultivated Potato
1.2. Molecular Studies of the Origin and Genetic Diversity of Chilean Cultivated Potato
2. Results
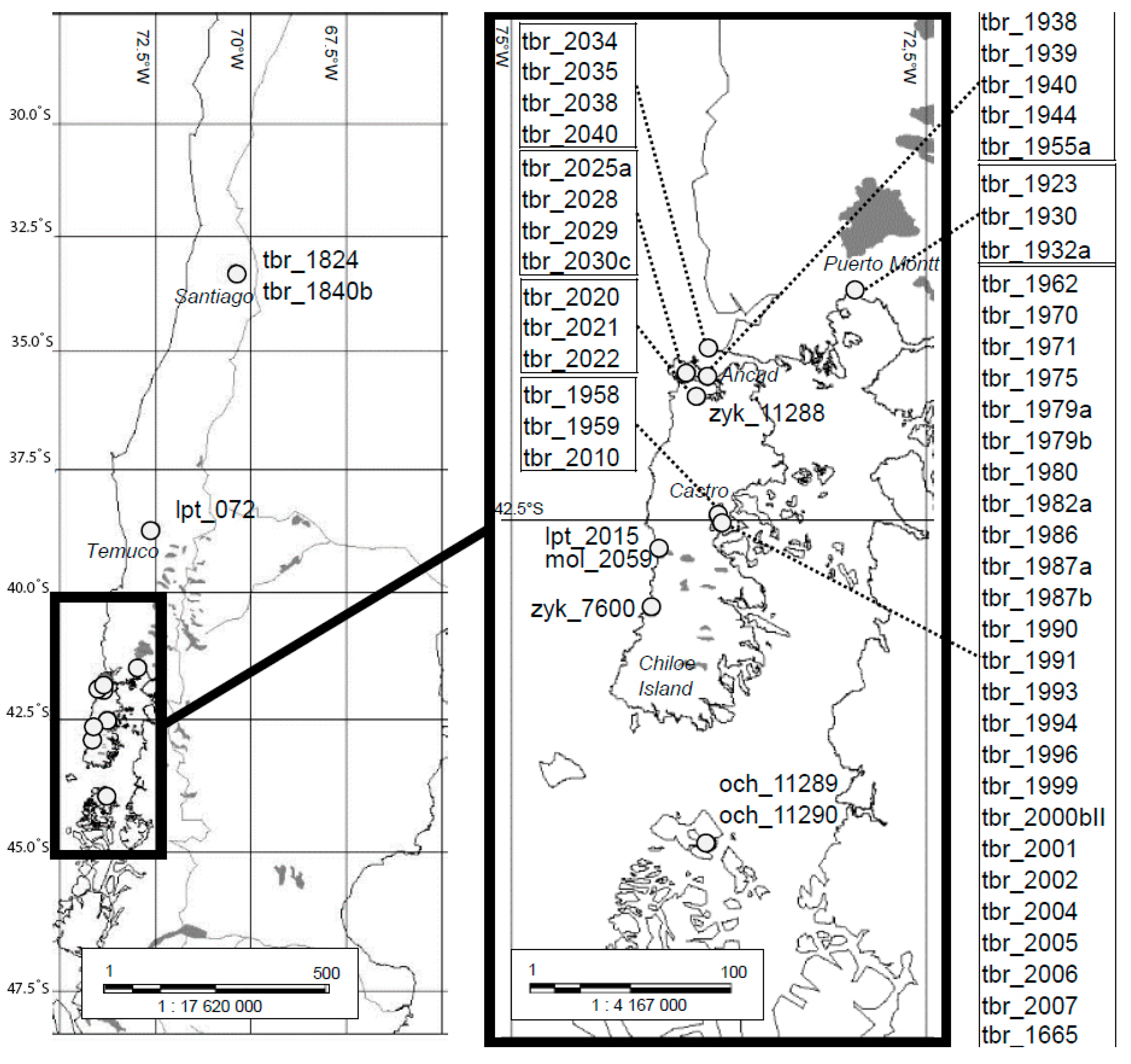
2.1. Geographic Origin of Authentic Herbarium Specimens
2.2. Chlorotypes of Herbarium Specimens
2.2.1. Plastid DNA Types of Herbarium Specimens
2.2.2. Plastid Microsatellite Haplotypes of Herbarium Specimens
2.3. Chlorotypes of the Living Chilean Landrace Accessions from the VIR Field Potato Collection
2.4. Mitotypes and Cytoplasm Types of the Living Chilean Landrace Accessions from the VIR Field Potato Collection
2.5. Molecular Screening with Markers of the R1, R3a, and Rysto Genes of Living Chilean Landrace Accessions from the VIR Field Gene Bank
3. Discussion
4. Materials and Methods
4.1. Sampling of Chilean Herbarium Specimens
4.2. Sampling of Living Chilean Accessions from the VIR Field Gene Bank
4.3. Geographical Distribution of Chilean Herbarium Specimens
4.4. DNA Isolation
4.5. Determination of Organelle DNA Type in Chilean Specimens
4.6. Plastid Microsatellite Analysis
4.7. Detection of the Diagnostic Markers of the Resistance R-Genes from Mexican Polyploid Species
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Hijmans, R.J.; Gavrilenko, T.; Stephenson, S.; Bamberg, J.; Salas, A.; Spooner, D.M. Geographic and environmental range expansion through polyploidy in wild potatoes (Solanum section Petota). Glob. Ecol. Biogeogr. 2007, 16, 485–495. [Google Scholar] [CrossRef]
- Spooner, D.M.; Ghislain, M.; Simon, R.; Jansky, S.H.; Gavrilenko, T. Systematics, Diversity, Genetics, and Evolution of Wild and Cultivated Potatoes. Bot. Rev. 2014, 80, 283–383. [Google Scholar] [CrossRef]
- Hawkes, J.G. The potato: Evolution, Biodiversity and Genetic Resources; Belhaven Press: Oxford, UK, 1990. [Google Scholar]
- Spooner, D.M.; Núñez, J.; Trujillo, G.M.; Del Rosario, H.; Guzmán, F.; Ghislain, M. Extensive simple sequence repeat genotyping of potato landraces supports a major reevaluation of their gene pool structure and classification. Proc. Natl. Acad. Sci. USA 2007, 104, 19398–19403. [Google Scholar] [CrossRef]
- Ovchinnikova, A.; Krylova, E.; Gavrilenko, T.; Smekalova, T.; Zhuk, M.; Knapp, S.; Spooner, D. Taxonomy of cultivated potatoes (Solanum section Petota: Solanaceae). Bot. J. Linn. Soc. 2011, 165, 107–155. [Google Scholar] [CrossRef]
- Spooner, D.M.; Gavrilenko, T.; Jansky, S.H.; Ovchinnikova, A.; Krylova, E.; Knapp, S.; Simon, R. Ecogeography of ploidy variation in cultivated potato (Solanum sect. Petota). Am. J. Bot. 2010, 97, 2049–2060. [Google Scholar] [CrossRef] [PubMed]
- Juzepczuk, S.V.; Bukasov, S.M. Proc. All-Union Congress Genet. Breed. Seed Prod. 1929, 3, 593–611.
- Bukasov, S.M. The potatoes of South America and their breeding possibilities. Proc. Appl. Bot. Gen. Breed 1933, 58, 192. [Google Scholar]
- Vavilov, N.I. Theoretical Basis of Plant Breeding; State Agicultural Oubling House: Leningrad, Russia, 1935. [Google Scholar]
- Vavilov, N.I. The great agricultural cultures of pre-Columbian America and their relationships. News State Geogr. Soc. 1939, 71, 1487–1515. [Google Scholar]
- Juzepczuk, S.V. New species of the genus Solanum L. in the group Tuberarium Dun. Bull. Acad. Sci. USSR 1937, 2, 295–331. [Google Scholar]
- Hawkes, J.G. Taxonomic studies on the tuber–bearing Solanum. I. Solanum tuberosum and the tetraploid species complex. Proc. Linn. Soc. Lond. 1956, 166, 97–144. [Google Scholar] [CrossRef]
- Chukhina, I.G.; Krylova, E.A.; Ovchinnikova, A.B.; Gavrilenko, T.A. Herbarium specimens of wild Chilean potato species belonging to the section Petota Dumort. Genus Solanum L. in the VIR Herbarium. Proc. Appl. Bot. Gen. Breed 2016, 177, 110–121. [Google Scholar] [CrossRef]
- Ugent, D.; Dillehay, T.; Ramirez, C. Potato remains from a late Pleistocene settlement in southcentral Chile. Econ. Bot. 1987, 41, 17–27. [Google Scholar] [CrossRef]
- Salaman, R.N. The early European potato: Its character and place of origin. J. Linn. Soc. Lond. Bot. 1946, 53, 1–27. [Google Scholar] [CrossRef]
- Salaman, R.N.; Hawkes, J.G. The character of the early European potato. Proc. Linn. Soc. Lond. 1949, 161, 71–84. [Google Scholar] [CrossRef]
- Correll, D.S. The potato and its wild relatives. In Contributions of the Texas Research Foundation, Botanical Studies; Texas Research Foundation: Renner, TX, USA, 1962. [Google Scholar]
- Simmonds, N.W. Studies of the Tetraploid Potatoes. J. Linn. Soc. Lond. Bot. 1964, 59, 43–56. [Google Scholar] [CrossRef]
- Grun, P. The evolution of cultivated potatoes. Econ. Bot. 1990, 44, 39–55. [Google Scholar] [CrossRef]
- Grun, P. Cytoplasmic sterilities that separate the Group Tuberosum cultivated potato from its putative tetraploid ancestor. Evolution 1973, 27, 633–643. [Google Scholar] [CrossRef]
- Grun, P.; Ochoa, C.; Capage, D. Evolution of cytoplasmic factors in tetraploid cultivated potatoes (Solanaceae). Am. J. Bot. 1977, 64, 412–420. [Google Scholar] [CrossRef]
- Spooner, D.M.; McLean, K.; Ramsay, G.; Waugh, R.; Bryan, G.J. A Single domestication for potato based on multilocus amplified fragment length polymorphism genotyping. Proc. Natl. Acad. Sci. USA 2005, 102, 14694–14699. [Google Scholar] [CrossRef]
- Li, Y.; Colleoni, C.; Zhang, J.; Liang, Q.; Hu, Y.; Ruess, H.; Simon, R.; Liu, Y.; Liu, H.; Yu, G.; et al. Genomic Analyses Yield Markers for Identifying Agronomically Important Genes in Potato. Mol. Plant 2018, 11, 473–484. [Google Scholar] [CrossRef]
- Tang, D.; Jia, Y.; Zhang, J.; Li, H.; Cheng, L.; Wang, P.; Bao, Z.; Liu, Z.; Feng, S.; Zhu, X.; et al. Genome evolution and diversity of wild and cultivated potatoes. Nature 2022, 606, 535–541. [Google Scholar] [CrossRef] [PubMed]
- Hosaka, K. Who Is the Mother of the Potato?—Restriction Endonuclease Analysis of Chloroplast DNA of Cultivated Potatoes. Theor. Appl. Genet. 1986, 72, 606–618. [Google Scholar] [CrossRef] [PubMed]
- Hosaka, K.; Hanneman, R.E. The origin of the cultivated tetraploid potato based on chloroplast DNA. Theor. Appl. Genet. 1988, 76, 172–176. [Google Scholar] [CrossRef] [PubMed]
- Sukhotu, T.; Kamijima, O.; Hosaka, K. Nuclear and chloroplast DNA differentiation in Andean potatoes. Genome 2004, 47, 46–56. [Google Scholar] [CrossRef] [PubMed]
- Lössl, A.; Adler, N.; Horn, R.; Frei, U.; Wenzel, G. Chondriome-type characterization of potato: Mt α, β, γδε and novel plastid-mitochondrial configurations in somatic hybrids. Theor. Appl. Genet. 1999, 99, 1–10. [Google Scholar] [CrossRef]
- Hosaka, K.; Sanetomo, R. Development of a rapid identification method for potato cytoplasm and its use for evaluating Japanese collections. Theor. Appl. Genet. 2012, 125, 1237–1251. [Google Scholar] [CrossRef]
- Lössl, A.; Götz, M.; Braun, A.; Wenzel, G. Molecular markers for cytoplasm in potato: Male sterility and contribution of different plastid-mitochondrial configurations to starch production. Euphytica 2000, 116, 221–230. [Google Scholar] [CrossRef]
- Kawagoe, Y.; Kikuta, Y. Chloroplast DNA evolution in potato (Solanum tuberosum L.). Theor. Appl. Genet. 1991, 81, 13–20. [Google Scholar] [CrossRef]
- Hosaka, K. T-type Chloroplast DNA in Solanum tuberosum L. ssp. tuberosum was conferred from some populations of S. tarijense Hawkes. Am. J. Potato Res. 2003, 80, 21–32. [Google Scholar] [CrossRef]
- Hosaka, K. Evolutionary Pathway of T–type Chloroplast DNA in Potato. Am. J. Potato Res. 2004, 81, 153–158. [Google Scholar] [CrossRef]
- Hosaka, K.; Sanetomo, R. Comparative differentiation in mitochondrial and chloroplast DNA among cultivated potatoes and closely related wild species. Genes Genet. Syst. 2009, 84, 371–378. [Google Scholar] [CrossRef] [PubMed]
- Ghislain, M.; Núñez, J.; Herrera, M.D.R.; Spooner, D.M. The single Andigenum origin of Neo-Tuberosum potato materials is not supported by microsatellite and plastid marker analyses. Theor. Appl. Genet. 2009, 118, 963–969. [Google Scholar] [CrossRef] [PubMed]
- Gavrilenko, T.; Antonova, O.; Shuvalova, A.; Krylova, E.; Alpatyeva, N.; Spooner, D.; Novikova, L. Genetic diversity and origin of cultivated potatoes based on plastid microsatellite polymorphism. Genet. Resour. Crop Evol. 2013, 60, 1997–2015. [Google Scholar] [CrossRef]
- Hosaka, K. Distribution of the 241 bp deletion of chloroplast DNA in wild potato species. Am. J. Potato Res. 2002, 79, 119–123. [Google Scholar] [CrossRef]
- Hardigan, M.A.; Laimbeer, F.P.E.; Newton, L.; Crisovan, E.; Hamilton, J.P.; Vaillancourt, B.; Wiegert-Rininger, K.; Wood, J.C.; Douches, D.S.; Farré, E.M.; et al. Genome diversity of tuber-bearing Solanum uncovers complex evolutionary history and targets of domestication in the cultivated potato. Proc. Natl. Acad. Sci. USA 2017, 114, E9999–E10008. [Google Scholar] [CrossRef]
- Rodríguez, F.; Ghislain, M.; Clausen, A.M.; Jansky, S.H.; Spooner, D.M. Hybrid origins of cultivated potatoes. Theor. Appl. Genet. 2010, 121, 1187–1198. [Google Scholar] [CrossRef] [PubMed]
- Spooner, D.M.; Jansky, S.; Clausen, A.; Del Rosario Herrera, M.; Ghislain, M. The Enigma of Solanum maglia in the Origin of the Chilean Cultivated Potato, Solanum tuberosum Chilotanum Group. Econ. Bot. 2012, 66, 12–21. [Google Scholar] [CrossRef]
- Cox, A.E.; Large, E.C. Plant pathology Laboratory, Ministry of Agriculture, Fisheries and Food. England. In Potato Blight Epidemics throughout the World; Agriculture Handbook; United State Department of Agriculture: Washington, DC, USA, 1960; Volume 174, pp. 160–165. [Google Scholar]
- Arentsen, S. History and development of phytopathology in Chile. In VII Congreso Latinoamericano de Fitopatología; Universidad Catolica de Chile: Santiago, Chile, 1994; pp. 126–128. [Google Scholar]
- Niederhauser, J.S. Phytophthora infestans: The Mexican connection. In Phytophthora; Lucas, J.A., Shattock, R.C., Shaw, D.S., Cooke, L.R., Eds.; Cambridge University Press: Cambridge, UK, 1991; pp. 25–45. [Google Scholar]
- Dodds, K.S. Classification of cultivated potatoes. In The Potato and Its Wild Relatives; Correll, D.S., Ed.; Texas Research Foundation: Renner, TX, USA, 1962; Volume 4, pp. 517–539. [Google Scholar]
- Zykin, A.G. Chilian cultivated potatoes and its wild relatives. Bull. Appl Bot Genet. Plant Breed 1973, 50, 216–243. [Google Scholar]
- Ochoa, C.M. Potato collecting expeditions in Chile, Bolivia and Peru, and the genetic erosion of indigenous cultivars. In Crop Genetic Resources for Today and Tomorrow; International Biological Programme 2; Frankel, O.H., Hawkes, J.G., Eds.; Cambridge University Press: Cambridge, UK, 1975; pp. 167–173. [Google Scholar]
- Spooner, D.M.; Contreras, M.A.; Bamberg, J.B. Potato germplasm collecting expedition to Chile, 1989, and utility of the Chilean species. Am. Potato J. 1991, 68, 681–690. [Google Scholar] [CrossRef]
- United States Department of Agriculture. Seeds and plants imported during the period from April 1 to June 30, 1908. Nos. 22511 to 23322. Bur. Plant Ind.—Bull. 1909, 15, 58–65. Available online: https://archive.org/details/plantinventoryno142unit/mode/2up (accessed on 11 November 2022).
- United States Department of Agriculture. Plant Material Introduced by the Division of Foreign Plant Introduction, Bureau of Plant Industry, April 1 to June 30, 1932 (nos. 98257–100467); United States Department of Agriculture: Washington DC, NY, USA, 1934; Volume 111, pp. 1–93. Available online: https://www.ars-grin.gov/npgs/pi_books/scans/pi111.pdf (accessed on 11 November 2022).
- Núñez, P.B.G. Determinación de Tipos Citoplasmáticos y Diversidad Genética a Nivel Nuclear de la Papa Nativa Chilena y Cultivares Europeos (Solanum tuberosum ssp. tuberosum). Ph.D. Thesis, Universidad Austral de Chile, Valdivia, Chile, 2007. [Google Scholar]
- Gutaker, R.M.; Weiß, C.L.; Ellis, D.; Anglin, N.L.; Knapp, S.; Luis Fernández-Alonso, J.; Prat, S.; Burbano, H.A. The origins and adaptation of European potatoes reconstructed from historical genomes. Nat. Ecol. Evol. 2019, 3, 1093–1101. [Google Scholar] [CrossRef] [PubMed]
- López, M.; Riegel, R.; Lizana, C.; Behn, A. Identification of virus and nematode resistance genes in the Chilota Potato Genebank of the Universidad Austral de Chile. Chil. J. Agric. Res. 2015, 75, 320–327. [Google Scholar] [CrossRef]
- Esnault, F.; Solano, J.; Perretant, M.-R.; Hervé, M.; Label, A.; Pellé, R.; Dantec, J.-P.; Boutet, G.; Brabant, P.; Chauvin, J.-E. Genetic diversity analysis of a potato (Solanum tuberosum L.) collection including Chiloé Island landraces and a large panel of worldwide cultivars. Plant Genet. Resour. 2014, 12, 74–82. [Google Scholar] [CrossRef]
- Glendinning, D.R. Chilean potatoes; an appraisal. Potato Res. 1975, 18, 306–307. [Google Scholar] [CrossRef]
- Contreras, A.; Castro, I. Catálogo de Variedades de Papas Nativas de Chile; Universidad Austral de Chile: Valdivia, Chile, 2008; 234p. [Google Scholar]
- Nagel, M.; Dulloo, M.; Bissessur, P.; Gavrilenko, T.; Bamberg, J.; Ellis, D.; Giovannini, P. Global Strategy for the Conservation of Potato; Global Crop Diversity Trust: Bonn, Germany, 2022; 160p. [Google Scholar] [CrossRef]
- Juzepczuk, S.V. About the South American Expedition of the All–Union Institute of Applied Botany and New Crops. News State Inst. Exp. Agron. 1928, 6, 11–12. [Google Scholar]
- Lechnovich, V.S. New wild potato species from the Pacific Coastal Islands of South America. Proc. Appl. Bot. Genet. Breed. 1978, 62, 36–54. [Google Scholar]
- Ovchinnikova, A.B.; Chukhina, I.G.; Gavrilenko, T.A. Type specimens of cultivated species of Potato section Petota Dumort. genus Solanum L., kept in the herbaria state scientific centre N.I.Vavilov All-Russian Research Institute of Plant Industry (WIR). Proc. Appl. Bot. Genet. Breed 2012, 169, 300–304. [Google Scholar]
- Chukhina, I.; Dorofeyev, V.; Gavrilenko, T.; Krylova, E.; Ovchinnikova, A. Type specimens of Solanum section Petota Dumort. (Solanaceae) taxa in the N. I. Vavilov Institute of Plant Genetic Resources (WIR) and the V. L. Komarov Botanical Institute (LE): New findings and updates. Turczaninowia 2017, 20, 97–105. [Google Scholar] [CrossRef][Green Version]
- Zykin, A.G. About the wild relatives of the Chilean potato. Plant Resour. 1971, 7, 242–252. [Google Scholar]
- Sanetomo, R.; Hosaka, K. A maternally inherited DNA marker, descended from Solanum demissum (2n=6x=72) to S. tuberosum (2n=4x=48). Breed. Sci. 2011, 61, 426–434. [Google Scholar] [CrossRef]
- Hosaka, K.; Sanetomo, R. Application of a PCR-Based Cytoplasm Genotyping Method for Phylogenetic Analysis in Potato. Am. J. Potato Res. 2014, 91, 246–253. [Google Scholar] [CrossRef]
- Heberling, J.M.; Prather, L.A.; Tonsor, S.J. The Changing Uses of Herbarium Data in an Era of Global Change: An Overview Using Automated Content Analysis. BioScience 2019, 69, 812–822. [Google Scholar] [CrossRef]
- Ames, M.; Spooner, D.M. DNA from herbarium specimens settles a controversy about origins of the European potato. Am. J. Bot. 2008, 95, 252–257. [Google Scholar] [CrossRef] [PubMed]
- Sanetomo, R.; Hosaka, K. Pollen transcriptome analysis of Solanum tuberosum (2n = 4x = 48), S. demissum (2n = 6x = 72), and their reciprocal F1 hybrids. Plant Cell Rep. 2013, 32, 623–636. [Google Scholar] [CrossRef] [PubMed]
- Ross, H. Potato Breeding—Problems and Perspectives; Verlag Paul Parey: Berlin, German, 1986; p. 132. [Google Scholar]
- Turner, R.S. After the famine: Plant pathology, Phytophthora infestans, and the late blight of potatoes, 1845–1960. Hist. Stud. Phys. Biol. Sci. 2005, 35, 341–370. [Google Scholar] [CrossRef]
- Bradshaw, J.E.; Bryan, G.J.; Ramsay, G. Genetic Resources (Including Wild and Cultivated Solanum Species) and Progress in their Utilisation in Potato Breeding. Potato Res. 2006, 49, 49–65. [Google Scholar] [CrossRef]
- Bradshaw, J.E. Potato Breeding: Theory and Practice; Springer: Cham, Switzerland, 2021; 553p. [Google Scholar]
- Bethke, P.C.; Halterman, D.A.; Jansky, S.H. Potato Germplasm Enhancement Enters the Genomics Era. Agronomy 2019, 9, 575. [Google Scholar] [CrossRef]
- Gavrilenko, T.A.; Klimenko, N.S.; Alpatieva, N.V.; Kostina, L.I.; Lebedeva, V.A.; Evdokimova, Z.Z.; Apalikova, O.V.; Novikova, L.Y.; Antonova, O. Cytoplasmic genetic diversity of potato varieties bred in Russia and FSU countries. Vavilov J. Genet. Breed. 2019, 23, 753–764. [Google Scholar] [CrossRef]
- Sanetomo, R.; Gebhardt, C. Cytoplasmic genome types of European potatoes and their effects on complex agronomic traits. BMC Plant Biol. 2015, 15, 162. [Google Scholar] [CrossRef]
- Mihovilovich, E.; Sanetomo, R.; Hosaka, K.; Ordoñez, B.; Aponte Villadoma, M.; Bonierbale, M. Cytoplasmic Diversity in Potato Breeding: Case Study from the International Potato Center. Mol. Breed. 2015, 35, 137. [Google Scholar] [CrossRef]
- Dionne, L.A. Cytoplasmic sterility in derivatives of Solanum demissum. Am. Potato J. 1961, 38, 117–120. [Google Scholar] [CrossRef]
- Irikura, Y. Studies on interspecific crosses of tuber-bearing Solanums. I. Overcoming and cross-incompatibility between Solanum tuberosum and other Solanum species by means of induced polyploids and haploids. Res. Bull. Hokkaido. Nat. Agr. Exp. Stat. 1968, 92, 23–27. [Google Scholar]
- Santayana, M.; Aponte, M.; Kante, M.; Eyzaguirre, R.; Gastelo, M.; Lindqvist-Kreuze, H. Cytoplasmic Male Sterility Incidence in Potato Breeding Populations with Late Blight Resistance and Identification of Breeding Lines with a Potential Fertility Restorer Mechanism. Plants 2022, 11, 3093. [Google Scholar] [CrossRef]
- Anisimova, I.; Gavrilenko, T. Cytoplasmic male sterility and prospects for its utilization in breeding, genetic studies and seed production of potato. Vavilovskii Zhurnal Genetiki i Selektsii 2017, 21, 83–95. [Google Scholar] [CrossRef]
- Anisimova, I.N.; Alpatieva, N.V.; Karabitsina, Y.I.; Gavrilenko, T.A. Nucleotide sequence polymorphism in the RFL-PPR genes of potato. J. Genet. 2019, 98, 87. [Google Scholar] [CrossRef] [PubMed]
- Kuang, H.; Wei, F.; Marano, M.R.; Wirtz, U.; Wang, X.; Liu, J.; Shum, W.P.; Zaborsky, J.; Tallon, L.J.; Rensink, W.; et al. The R1 resistance gene cluster contains three groups of independently evolving, type I R1 homologues and shows substantial structural variation among haplotypes of Solanum demissum. Plant J. 2005, 44, 37–51. [Google Scholar] [CrossRef] [PubMed]
- Sokolova, E.; Pankin, A.; Beketova, M.; Kuznetsova, M.; Spiglazova, S.; Rogozina, E.; Yashina, I.; Khavkin, E. SCAR markers of the R-genes and germplasm of wild Solanum species for breeding late blightresistant potato cultivars. Plant Genet. Res. 2011, 9, 309–312. [Google Scholar] [CrossRef]
- Aguilera-Galvez, C.; Champouret, N.; Rietman, H.; Lin, X.; Wouters, D.; Chu, Z.; Jones, J.D.G.; Vossen, J.H.; Visser, R.G.F.; Wolters, P.J.; et al. Two different R gene loci co-evolved with Avr2 of Phytophthora infestans and confer distinct resistance specificities in potato. Stud. Mycol. 2018, 89, 105–115. [Google Scholar] [CrossRef] [PubMed]
- The Potato Genome Sequencing Consortium. Genome sequence and analysis of the tuber crop potato. Nature 2011, 475, 189–195. [Google Scholar] [CrossRef]
- Song, Y.-S.; Schwarzfischer, A. Development of STS markers for selection of extreme resistance (Rysto) to PVY and maternal pedigree analysis of extremely resistant cultivars. Am. J. Potato Res. 2008, 85, 159–170. [Google Scholar] [CrossRef]
- Kostina, L. Aboriginal varieties of Solanum chilotanum Hawk. Bull. Appl. Bot. Genet. Breed. 1978, 62, 55–82. [Google Scholar]
- Chimote, V.P.; Chakrabarti, S.K.; Pattanayak, D.; Pandey, S.K.; Naik, P.S. Molecular analysis of cytoplasm type in Indian potato varieties. Euphytica 2008, 162, 69–80. [Google Scholar] [CrossRef]
- Iwanaga, M.; Ortiz, R.; Cipar, M.S.; Peloquin, S.J. A Restorer gene for genetic-cytoplasmic male sterility in cultivated potatoes. Am. Potato J. 1991, 68, 19–28. [Google Scholar] [CrossRef]
- Ortiz, R.; Iwanaga, M.; Peloquin, S.J. Male sterility and 2n pollen in 4x progenies derived from 4x × 2x and 4x × 4x crosses in potatoes. Potato Res. 1993, 36, 227–236. [Google Scholar] [CrossRef]
- Grun, P. Changes of cytoplasmic factors during the evolution of the cultivated potato. Evolution 1970, 24, 188–198. [Google Scholar] [CrossRef] [PubMed]
- Grun, P. Cytoplasmic sterilities that separate the cultivated potato from its putative diploid ancestors. Evolution 1970, 24, 750–758. [Google Scholar] [CrossRef] [PubMed]
- Hoopes, R.W.; Plaisted, R.L.; Cubillos, A.G. Yield and fertility of reciprocal-cross Tuberosum-Andigena hybrids. Am. Potato J. 1980, 57, 275–284. [Google Scholar] [CrossRef]
- Hermundstad, S.A.; Peloquin, S.J. Male fertility and 2n pollen production in haploid-wild species hybrids. Am. Potato J. 1985, 62, 479–487. [Google Scholar] [CrossRef]
- Tucci, M.; Carputo, D.; Bile, G.; Frusciante, L. Male fertility and freezing folerance of hybrids involving Solanum tuberosum haploids and diploid Solanum species. Potato Res. 1996, 39, 345–353. [Google Scholar] [CrossRef]
- Santini, M.; Camadro, E.L.; Marcellán, O.N.; Erazzú, L.E. Agronomic characterization of diploid hybrid families derived from crosses between haploids of the common potato and three wild Argentinian tuber-bearing species. Am. J. Pot. Res. 2000, 77, 211–218. [Google Scholar] [CrossRef]
- Hilali, A.; Lauer, F.I.; Veilleux, R.E. Reciprocal differences between hybrids of Solanum tuberosum Groups Tuberosum (haploid) and Phureja. Euphytica 1987, 36, 631–639. [Google Scholar] [CrossRef]
- Contreras, A.; Ciampi, L.; Padulosi, S.; Spooner, D.M. Potato germplasm collecting expedition to the Guaitecas and Chonos Archipelagos, Chile, 1990. Potato Res. 1993, 36, 309–316. [Google Scholar] [CrossRef]
- Ristaino, J.B.; Pfister, D.H. “What a Painfully Interesting Subject”: Charles Darwin’s Studies of Potato Late Blight. BioScience 2016, 66, 1035–1045. [Google Scholar] [CrossRef]
- United States Department of Agriculture. Bureau of plant industry. Inventory of seeds and plants imported by the office of foreign seed and plant introduction during the period from July 1 to September 30,1913 (no. 36; nos, 35667 to 36258). In Bureau of Plant Industry—Bulletin; United States Department of Agriculture: Washington DC, NY, USA, 1915; p. 53. Available online: https://www.ars-grin.gov/npgs/pi_books/scans/pi036.pdf (accessed on 20 November 2022).
- Rabinowitz, D.; Linder, C.; Ortega, R.; Begazo, D.; Murguia, H.; Douches, D.; Quiros, C. High levels of interspecific hybridization between Solanum Sparsipilum and S. Stenotomum in experimental plots in the Andes. Am. J. Potato Res. 1990, 67, 73–81. [Google Scholar] [CrossRef]
- Turland, N.J.; Wiersema, J.H.; Barrie, F.R.; Greuter, W.; Hawksworth, D.L.; Herendeen, P.S.; Knapp, S.; Kusber, W.-H.; Li, D.-Z.; Marhold, K.; et al. International Code of Nomenclature for Algae, Fungi, and Plants (Shenzhen Code) Adopted by the Nineteenth International Botanical Congress Shenzhen, China, July 2017; Koeltz Botanical Books: Shenzhen, China, 2018. [Google Scholar]
- Bryan, G.J.; McNicoll, J.; Ramsay, G.; Meyer, R.C.; De Jong, W.S. Polymorphic simple sequence repeat markers in chloroplast genomes of Solanaceous plants. Theor. Appl. Genet. 1999, 99, 859–867. [Google Scholar] [CrossRef]
- Ballvora, A.; Ercolano, M.R.; Weiss, J.; Meksem, K.; Bormann, C.A.; Oberhagemann, P.; Salamini, F.; Gebhardt, C. The R1 gene for potato resistance to late blight (Phytophthora Infestans) belongs to the leucine zipper/NBS/LRR class of plant resistance genes. Plant J. 2002, 30, 361–371. [Google Scholar] [CrossRef]
- Mori, K.; Sakamoto, Y.; Mukojima, N.; Tamiya, S.; Nakao, T.; Ishii, T.; Hosaka, K. Development of a multiplex PCR method for simultaneous detection of diagnostic DNA markers of five disease and pest resistance genes in potato. Euphytica 2011, 180, 347–355. [Google Scholar] [CrossRef]
- Huang, S.; van der Vossen, E.A.G.; Kuang, H.; Vleeshouwers, V.G.A.A.; Zhang, N.; Borm, T.J.A.; van Eck, H.J.; Baker, B.; Jacobsen, E.; Visser, R.G.F. comparative genomics enabled the isolation of the R3a late blight resistance gene in potato. Plant J. 2005, 42, 251–261. [Google Scholar] [CrossRef]
- Song, Y.-S.; Hepting, L.; Schweizer, G.; Hartl, L.; Wenzel, G.; Schwarzfischer, A. Mapping of extreme resistance to PVY (Rysto) on chromosome XII using anther-culture-derived primary dihaploid potato lines. Theor. Appl. Genet. 2005, 111, 879–887. [Google Scholar] [CrossRef]
- Hawkes, J.G.; Hjerting, J.P. The Potatoes of Bolivia: Their Breeding Value and Evolutionary Relationships, 1st ed; Oxford University Press: Oxford, UK, 1989. [Google Scholar]
- Ochoa, C.M. Las Papas de Sudamérica: Peru; Centro Internacional de la Papa (CIP): Peru, Lima, 1999. [Google Scholar]
- Khoury, C.K.; Brush, S.; Costich, D.E.; Curry, H.A.; de Haan, S.; Engels JM, M.; Guarino, L.; Hoban, S.; Mercer, K.L.; Miller, A.J.; et al. Crop Genetic Erosion: Understanding and Responding to Loss of Crop Diversity. New Phytol. 2022, 233, 84–118. [Google Scholar] [CrossRef]
- van de Wouw, M.; Kik, C.; van Hintum, T.; van Treuren, R.; Visser, B. Genetic Erosion in Crops: Concept, Research Results and Challenges. Plant Genet. Resour. 2010, 8, 1–15. [Google Scholar] [CrossRef]


{kind=link}
| Name of Species | Number of Analyzed Herbarium Samples | Number (Frequency) of Specimens with Different | Name of Species According to | Number of Analyzed Herbarium Samples | Number (Frequency) of Herbarium Specimens with Different cpDNA Types and cpSSR Haplotypes | |||
|---|---|---|---|---|---|---|---|---|
| cpDNA Type | Plastid SSR Haplotypes | Hawkes, 1990 [3] | Spooner et al., 2007; Ovchinnikova et al., 2011 [4,5] | |||||
| Chilean cultivated potato | Chilean cultivated potato | |||||||
| S. tuberosum L. (S. tuberosum s.str.) | 49 1 | T—47 (0.96) A—2 (0.04) | #III—47 (0.96) #II—2 (0.04) | S. tuberosum subsp. tuberosum | S. tuberosum Chilotanum Group | 59 | T (#III)—57 (0.97) A (#II)—2 (0.03) | |
| Wild Chilean species according to Russian taxonomists: | ||||||||
| S. molinae Juz. | 4 1 | T—4 | #III—4 | |||||
| S. leptostigma Juz. | 2 1 | T—2 | #III—2 | |||||
| Total number of samples collected in Chile in 1928 | 55 1 | T—53 (0.96) A—2 (0.04) | #III—53 (0.96) #II—2 (0.04) | |||||
| S. ochoanum Lechn. | 2 2 | T—2 | #III—2 | |||||
| S. zykinii Lechn. | 2 2 | T—2 | #III—2 | |||||
| Wild species: S. maglia Schltdl. | 2 | A—2 | #II—2 | Wild species: S. maglia Schltdl. | 2 | A (#II)—1 3 | ||
| Total number of herbarium specimens | 61 | 61 | ||||||
| Gene Bank Collection—Source of Analyzed Accessions (Reference) | No. of Analyzed Accessions | Number (Frequency, %) of Accessions with Specific Organelle DNA Haplotypes | Nomenclature of Cytoplasm Types of Lössl et al., 2000 1 [30] | |||
|---|---|---|---|---|---|---|
| cpDNA with the 241 bp Deletion/without This Deletion | cpDNA-Type | mtDNA-Type | Modern Nomenclature of Potato Cytoplasm Types of Hosaka and Sanetomo, 2012 [29] | |||
| (a) Present Study | ||||||
| VIR, Russia VIR field gene bank Potato collection (Present study) | 46 | with deletion—83.0 | T-type—84.8 | β-type—89.1 | T (T/β)—84.8 | T β—84.8 |
| without deletion—17.0 | A-type—4.3 | A (A/β)—4.3 | without deletion/β—4.3 | |||
| W-type—10.9 | α-type—10.9 | D (W/α)—10.9 | without deletion/α—10.9 | |||
| other types of cpDNA—0 | other types of mtDNA—0 | other cytoplasm types—0 | ||||
| (b) Data from the Literature | ||||||
| UACH, Chile [50] | 114 2 | with deletion—80.7 | T-type –80.7 | β-type—82.5 | n.d. | T/β—78.1 |
| α-type—17.5 | T/α—2.6 | |||||
| without deletion—19.3 | other types of cpDNA—n.d. | without deletion/β—4.4 | ||||
| other types of mtDNA—0 | without deletion/α—14.9 | |||||
| NRSP-6, USA [26] | 24 | with deletion—87.5 | T-type–87.5 | n.d. | n.d. | n.d. |
| without deletion—12.5 | A-type—8.3 | |||||
| W-type—4.2 | ||||||
| other types of cpDNA—0 | ||||||
| NRSP-6, USA [29,34] | 8 | with deletion—87.5 | T-type—87.5 | β-type—100 | T (T/β)—87.5 | T/β—87.5 |
| without deletion—12.5 | A-type—12.5 | A (A/β)—12.5 | without deletion/β—12.5 | |||
| other types of cpDNA—0 | α-type—0 | 0 | ||||
| CIP, Peru [4] | 27 | with deletion—81.5 | T-type—81.5 | n.d. | n.d. | n.d. |
| without deletion—18.5 | other types of cpDNA—n.d. | |||||
| CIP, Peru [35] Supplementary Table | 134 3 | with deletion—82.1 | T-type—82.1 | n.d. | n.d. | n.d. |
| without deletion—17.9 | other types of cpDNA—n.d. | |||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gavrilenko, T.; Chukhina, I.; Antonova, O.; Krylova, E.; Shipilina, L.; Oskina, N.; Kostina, L. Comparative Analysis of the Genetic Diversity of Chilean Cultivated Potato Based on a Molecular Study of Authentic Herbarium Specimens and Present-Day Gene Bank Accessions. Plants 2023, 12, 174. https://doi.org/10.3390/plants12010174
Gavrilenko T, Chukhina I, Antonova O, Krylova E, Shipilina L, Oskina N, Kostina L. Comparative Analysis of the Genetic Diversity of Chilean Cultivated Potato Based on a Molecular Study of Authentic Herbarium Specimens and Present-Day Gene Bank Accessions. Plants. 2023; 12(1):174. https://doi.org/10.3390/plants12010174
Chicago/Turabian StyleGavrilenko, Tatjana, Irena Chukhina, Olga Antonova, Ekaterina Krylova, Liliya Shipilina, Natalia Oskina, and Ludmila Kostina. 2023. "Comparative Analysis of the Genetic Diversity of Chilean Cultivated Potato Based on a Molecular Study of Authentic Herbarium Specimens and Present-Day Gene Bank Accessions" Plants 12, no. 1: 174. https://doi.org/10.3390/plants12010174
APA StyleGavrilenko, T., Chukhina, I., Antonova, O., Krylova, E., Shipilina, L., Oskina, N., & Kostina, L. (2023). Comparative Analysis of the Genetic Diversity of Chilean Cultivated Potato Based on a Molecular Study of Authentic Herbarium Specimens and Present-Day Gene Bank Accessions. Plants, 12(1), 174. https://doi.org/10.3390/plants12010174