


Soil and Foliar Applications of Wood Distillate Differently Affect Soil Properties and Field Bean Traits in Preliminary Field Tests



Abstract
1. Introduction
2. Results
2.1. Effect on Soil Quality and Nutrient Content
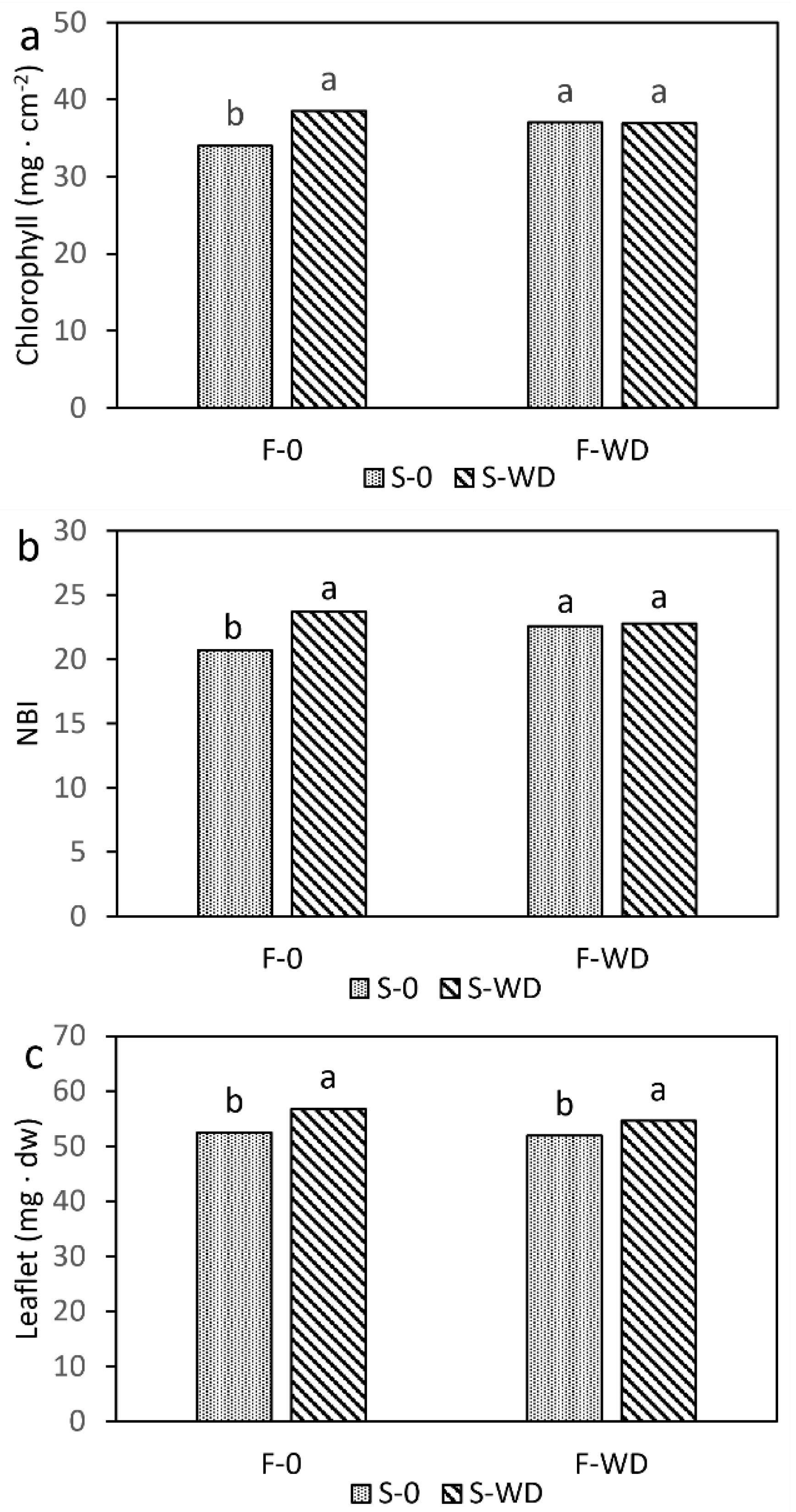
2.2. Effects on Field Bean Plants
2.3. Effects on Roots
3. Discussion
4. Materials and Methods
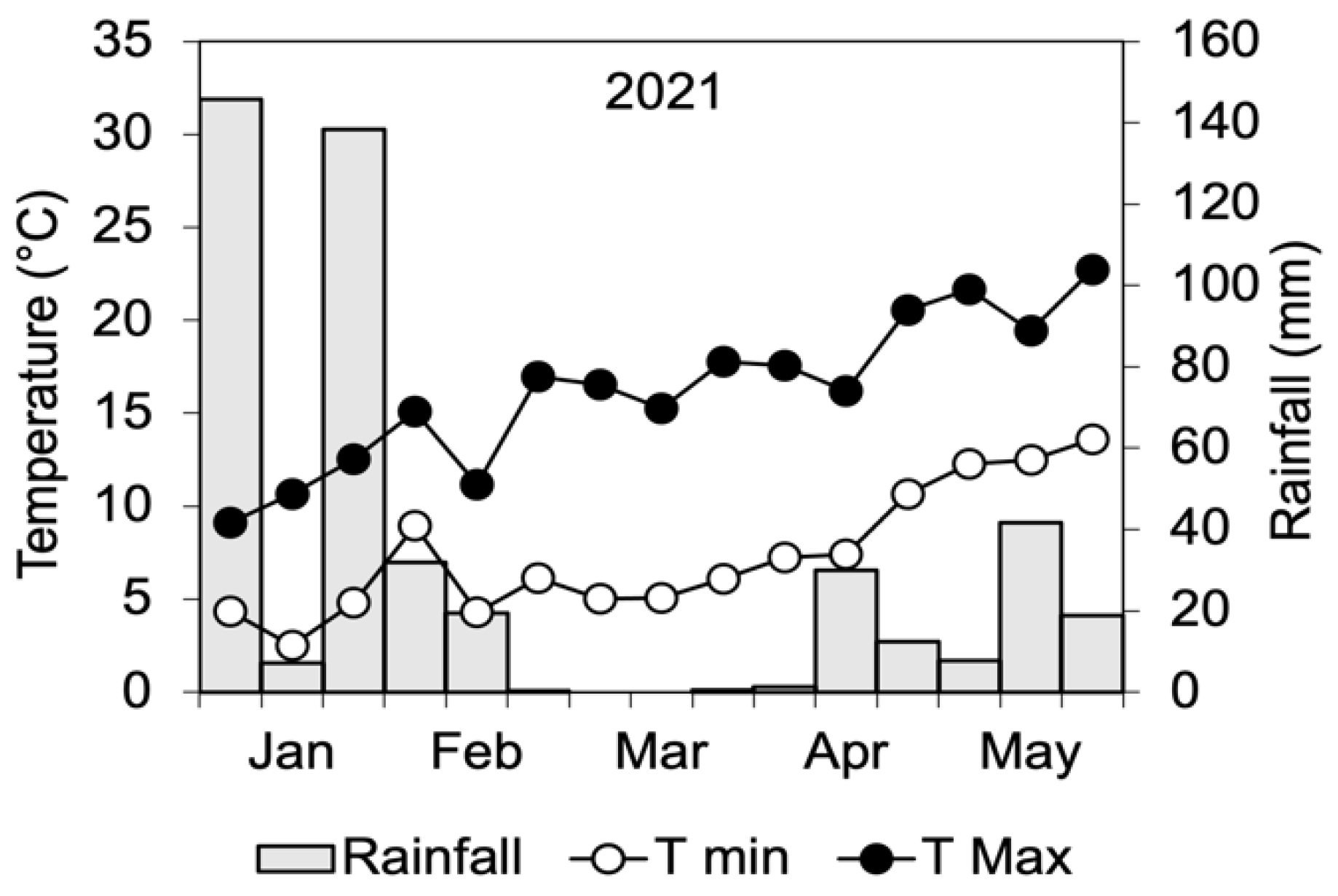
4.1. Field Experiment
4.1.1. Experimental Setup
4.1.2. Plant Harvest and Measurements
4.1.3. Soil Sampling and Analysis
4.2. Pot Experiment
4.2.1. Experimental Setup
4.2.2. Treatments
4.2.3. Harvest and Measurements
4.3. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Grewal, A.; Abbey Gunupuru, L.R. Production, prospects and potential application of pyroligneous acid in agriculture. J. Anal. Appl. Pyrolysis 2018, 135, 152–159. [Google Scholar] [CrossRef]
- Ma, X.; Wei, Q.; Zhang, S.; Shi, L.; Zhao, Z. Isolation and bioactivities of organic acids and phenols from walnut shell pyroligneous acid. J. Anal. Appl. Pyrolysis 2011, 91, 338–343. [Google Scholar] [CrossRef]
- Nelson, D.C.; Flematti, G.R.; Ghisalberti, E.L.; Dixon, K.W.; Smith, S.M. Regulation of seed germination and seedling growth by chemical signals from burning vegetation. Annu. Rev. Plant Biol. 2012, 63, 107–130. [Google Scholar] [CrossRef]
- Simma, B.; Polthanee, A.; Goggi, A.S. Wood vinegar seed priming improves yield and suppresses weeds in dryland direct-seeding rice under rainfed production. Agron. Sustain. Dev. 2017, 37, 56. [Google Scholar] [CrossRef]
- Wang, Y.; Qiu, L.; Song, Q.; Wang, S.; Wang, Y.; Ge, Y. Root Proteomics Reveals the Effects of Wood Vinegar on Wheat Growth and Subsequent Tolerance to Drought Stress. Int. J. Mol. Sci. 2019, 20, 943. [Google Scholar] [CrossRef]
- Mao, K.; Li, S.; Li, B.; Wu, W.; Wei, C.; Yuan, S.; Niu, Y.; Du, H.; Zhang, L. Effect of wood vinegar on growth, yield and quality of upper leaves of flue-cured tobacco of Nanzheng. SW China J. Agric. Sci. 2019, 32, 645–652. [Google Scholar]
- Zhu, K.; Gu, S.; Liu, J.; Luo, T.; Khan, Z.; Zhang, K.; Hu, L. Wood vinegar as a complex growth regulator promotes the growth, yield, and quality of rapeseed. Agronomy 2021, 11, 510. [Google Scholar] [CrossRef]
- Vannini, A.; Fedeli, R.; Guarnieri, M.; Loppi, S. Damage in Lettuce (Lactuca sativa L.). Toxics 2022, 10, 178. [Google Scholar] [CrossRef]
- Sun, H.; Feng, Y.; Xue, L.; Mandal, S.; Wang, H.; Shi, W.; Yang, L. Responses of ammonia volatilization from rice paddy soil to application of wood vinegar alone or combined with biochar. Chemosphere 2020, 242, 125247. [Google Scholar] [CrossRef]
- Duc, G. Faba bean (Vicia faba L.). F. Crop. Res. 1997, 53, 99–109. [Google Scholar] [CrossRef]
- Mariotti, M.; Andreuccetti, V.; Arduini, I.; Minieri, S.; Pampana, S. Field Bean for Forage and Grain in Short-Season Rainfed Mediterranean Conditions. Ital. J. Agron. 2018, 13, 208–215. [Google Scholar] [CrossRef]
- Köpke, U.; Nemecek, T. Ecological Services of Faba Bean. F. Crop. Res. 2010, 115, 217–233. [Google Scholar] [CrossRef]
- Egli, D.B. Flowering, Pod Set and Reproductive Success in Soya Bean. J. Agron. Crop. Sci. 2005, 191, 283–291. [Google Scholar] [CrossRef]
- De Costa, W.A.J.M.; Dennett, M.D.; Ratnaweera, U.; Nyalemegbe, K. Effects of Different Water Regimes on Field-Grown Determinate and Indeterminate Faba Bean (Vicia faba L.). I. Canopy Growth and Biomass Production. F. Crop. Res. 1997, 49, 83–93. [Google Scholar] [CrossRef]
- Salon, C.; Munier-Jolain, N.G.; Duc, G.; Voisin, A.S.; Grandgirard, D.; Larmure, A.; Emery, R.J.N.; Ney, B. Grain Legume Seed Filling in Relation to Nitrogen Acquisition: A Review and Prospects with Particular Reference to Pea. Agronomie 2001, 21, 539–552. [Google Scholar] [CrossRef]
- Pampana, S.; Masoni, A.; Arduini, I. Grain Legumes Differ in Nitrogen Accumulation and Remobilisation during Seed Filling. Acta Agric. Scand. Sect. B Soil Plant Sci. 2016, 66, 127–132. [Google Scholar] [CrossRef]
- Cheema, Z.A.; Ahmad, A. Effects of Urea on the Nitrogen Fixing Capacity and Growth of Grain Legumes. Int. J. Agric. Biol. 2000, 2, 388–394. [Google Scholar]
- Caliskan, S.; Ozkaya, I.; Caliskan, M.E.; Arslan, M. The Effects of Nitrogen and Iron Fertilization on Growth, Yield and Fertilizer Use Efficiency of Soybean in a Mediterranean-Type Soil. F. Crop. Res. 2008, 108, 126–132. [Google Scholar] [CrossRef]
- Kocon, A. The effect of foliar or soil top-dressing of urea on some physiological processes and seed yield of faba bean. Pol. J. Agron. 2010, 3, 15–19. [Google Scholar]
- Namvar, A.; Sharifi, R.S.; Sedghi, M.; Zakaria, R.A.; Khandan, T.; Eskandarpour, B. Study on the effects of organic and inorganic nitrogen fertilizer on yield, yield components and nodulation state of chickpea (Cicer arietinum L.). Commun. Soil Sci. Plant Anal. 2011, 42, 1097–1109. [Google Scholar] [CrossRef]
- Pampana, S.; Masoni, A.; Mariotti, M.; Ercoli, L.; Arduini, I. Nitrogen Fixation of Grain Legumes Differs in Response to Nitrogen Fertilisation. Exp. Agric. 2018, 54, 66–82. [Google Scholar] [CrossRef]
- Salvagiotti, F.; Cassman, K.G.; Specht, J.E.; Walters, D.T.; Weiss, A.; Dobermann, A. Nitrogen uptake, fixation and response to fertilizer N in soybeans: A review. Field Crops Res. 2008, 108, 1–13. [Google Scholar] [CrossRef]
- Pampana, S.; Scartazza, A.; Cardelli, R.; Saviozzi, A.; Guglielminetti, L.; Vannacci, G.; Mariotti, M.; Masoni, A.; Arduini, I. Biosolids Differently Affect Seed Yield, Nodule Growth, Nodule-Specific Activity, and Symbiotic Nitrogen Fixation of Field Bean. Crop. Pasture Sci. 2017, 68, 735–745. [Google Scholar] [CrossRef]
- Stoddard, F.L.; Nicholas, A.H.; Rubiales, D.; Thomas, J.; Villegas-Fernández, A.M. Integrated Pest Management in Faba Bean. F. Crop. Res. 2010, 115, 308–318. [Google Scholar] [CrossRef]
- De Vita, P.; Colecchia, S.A.; Pecorella, I.; Saia, S. Reduced inter-row distance improves yield and competition against weeds in a semi-dwarf durum wheat variety. Eur. J. Agron. 2017, 85, 69–77. [Google Scholar] [CrossRef]
- Tiilikkala, K.; Fagernäs, L.; Tiilikkala, J. History and Use of Wood Pyrolysis Liquids as Biocide and Plant Protection Product. Open Agric. J. 2014, 4, 111–118. [Google Scholar] [CrossRef]
- Mourant, D.; Yang, D.Q.; Lu, X.; Roy, C. Anti-Fungal Properties of the Pyroligneous Liquors from the Pyrolysis of Softwood Bark. Wood Fiber Sci. 2005, 37, 542–548. [Google Scholar]
- Andersson, G.K.S.; Ekroos, J.; Stjernman, M.; Rundlöf, M.; Smith, H.G. Effects of Farming Intensity, Crop Rotation and Landscape Heterogeneity on Field Bean Pollination. Agric. Ecosyst. Environ. 2014, 184, 145–148. [Google Scholar] [CrossRef]
- Bailes, E.J.; Pattrick, J.G.; Glover, B.J. An Analysis of the Energetic Reward Offered by Field Bean (Vicia faba) Flowers: Nectar, Pollen, and Operative Force. Ecol. Evol. 2018, 8, 3161–3171. [Google Scholar] [CrossRef]
- Cook, B.D.; Allan, D.L. Dissolved organic carbon in old field soils: Compositional changes during the biodegradation of soil organic matter. Soil Biol. Biochem. 1992, 24, 595–600. [Google Scholar] [CrossRef]
- Ma, C.; Li, W.; Zu, Y.; Yang, L.; Li, J. Antioxidant properties of pyroligneous acid obtained by thermochemical conversion of schisandra chinensis baill. Molecules 2014, 19, 20821–20838. [Google Scholar] [CrossRef] [PubMed]
- Wei, D.; Houtman, C.J.; Kapich, A.N.; Hunt, C.G.; Cullen, D.; Hammel, K.E. Laccase and its role in production of extracellular reactive oxygen species during wood decay by the brown rot basidiomycetes Postia placenta. Appl. Environ. Microbiol. 2010, 76, 2091–2097. [Google Scholar] [CrossRef] [PubMed]
- Juric, S.; Juric, M.; Režek-Jambrak, A.; Vincekovi’c, M. Tailoring alginate/chitosan microparticles loaded with chemical and biological agents for agricultural application and production of value-added foods. Appl. Sci. 2021, 11, 4061. [Google Scholar] [CrossRef]
- Rauha, J.P.; Remes, S.; Heinonen, M.; Hopia, A.; Kähkönen, M.; Kujala, T.; Pihlaja, K.; Vuorela, H.; Vuorela, P. Antimicrobial effects of Finnish plant extracts containing flavonoids and other phenolic com- pounds. Int. J. Food Microbiol. 2000, 56, 3–12. [Google Scholar] [CrossRef]
- Silva, T.A.; Castro, J.S.; Ribeiro, V.J.; Ribeiro, J.I., Jr.; Tavares, G.P.; Calijuri, M.L. Microalgae biomass as a renewable biostimulant: Meat processing industry effluent treatment, soil health improvement, and plant growth. Environ. Technol. 2021, 23, 1–17. [Google Scholar] [CrossRef]
- Zhang, R.; Wei, D.; Yao, Z.; Chao, Z.; Xiaojuan, A. Effects of wood vinegar on the soil microbial characteristics. J. Chem. Pharm. Res. 2014, 6, 1254–1260. [Google Scholar]
- Koç, I.; Öğün, E.; Namli, A.; Mendeş, M.; Kutlu, E.; Yardim, E.N. The effects of wood vinegar on some soil microorganisms. Appl. Ecol. Environ. Res. 2019, 17, 2437–2447. [Google Scholar] [CrossRef]
- Cardelli, R.; Becagli, M.; Marchini, F.; Saviozzi, A. Soil biochemical activities after the application of pyroligneous acid to soil. Soil Res. 2020, 58, 461–467. [Google Scholar] [CrossRef]
- Liang, B.; Lehmann, J.; Sohi, S.; Thies, J.; O’Neill, B.; Trujillo, L.; Gaunt, J.; Solomon, D.; Grossman, J.; Neves, E.; et al. Black carbon affects the cycling of non-black carbon in soil. Org. Geochem. 2010, 41, 206–213. [Google Scholar] [CrossRef]
- Meyer, A.H.; Wooldridge, J.; Dames, J.F. Relationship between soil alteration index three (AI3), soil organic matter and tree performance in a Cripps Pink/M7 apple orchard. S. Afr. J. Plant Soil 2014, 31, 173–175. [Google Scholar] [CrossRef]
- Puglisi, E.; Del Re, A.A.M.; Rao, M.A.; Gianfreda, L. Development and validation of numerical indexes integrating enzyme activities of soils. Soil Biol. Biochem. 2006, 38, 1673–1681. [Google Scholar] [CrossRef]
- Paz-Ferreiro, J.; Trasar-Cepeda, C.; Leirós, M.C.; Seoane, S.; Gil-Sotres, F. Biochemical properties in managed grassland soils in a temperate humid zone: Modifications of soil quality as a consequence of intensive grassland use. Biol. Fert. Soils 2009, 45, 711–722. [Google Scholar] [CrossRef]
- Ghosh, A.; Singh, A.B.; Kumar, R.V.; Manna, M.C.; Bhattacharyya, R.; Rahman, M.M.; Sharma, P.; Rajput, P.S.; Misra, S. Soil enzymes and microbial elemental stoichiometry as bio-indicators of soil quality in diverse cropping systems and nutrient management practices of Indian Vertisols. Appl. Soil Ecol. 2020, 145, 103304. [Google Scholar] [CrossRef]
- Van Huyssteen, I.; Mulidzi, A.R.; Meyer, A.H.; Wooldridge, J. Alteration Index Three Facilitates Interpretation of β-Glucosidase, Acid-phosphatase and Urease Activities in Soils Irrigated with Diluted Winery Wastewater. S. Afr. J. Enol. Vitic. 2020, 14, 238–244. [Google Scholar] [CrossRef]
- Togoro, A.H.; Silva, J.A.S.; Cazetta, J.O. Chemical changes in an oxisol treated with pyroligneous acid. Ciênc. Agrotec. 2014, 38, 113–121. [Google Scholar] [CrossRef]
- Becagli, M.; Santin, M.; Cardelli, R. Co-application of wood distillate and biochar improves soil quality and plant growth in basil (Ocimum basilicum). J. Plant Nutr. Soil Sci. 2022, 185, 120–131. [Google Scholar] [CrossRef]
- Zhang, F.; Shao, J.; Yang, H.; Guo, D.; Chen, Z.; Zhang, S.; Chen, H. Effects of biomass pyrolysis derived wood vinegar on microbial activity and communities of activated sludge. Bioresour. Technol. 2019, 279, 252–261. [Google Scholar] [CrossRef]
- Lashari, M.S.; Ye, Y.; Ji, H.; Li, L.; Kibue, G.W.; Lu, H.; Pan, G. Biochar-manure compost in conjunction with pyroligneous solution alleviated salt stress and improved leaf bioactivity of maize in a saline soil from central china: A 2-year field experiment. J. Sci. Food Agr. 2015, 95, 1321–1327. [Google Scholar] [CrossRef]
- Utsumi, Y.; Utsumi, C.; Tanaka, M.; Van Ha, C.; Takahashi, S.; Matsui, A.; Matsunaga, T.M.; Matsunaga, S.; Kanno, Y.; Seo, M.; et al. Acetic acid treatment enhances drought avoidance in cassava (Manihot esculenta Crantz). Front. Plant Sci. 2019, 10, 1–12. [Google Scholar] [CrossRef]
- Fedeli, R.; Vannini, A.; Guarnieri, M.; Monaci, F.; Loppi, S. Bio-Based Solutions for Agriculture: Foliar Application of Wood Distillate Alone and in Combination with Other Plant-Derived Corroborants Results in Different Effects on Lettuce (Lactuca sativa L.). Biology 2022, 11, 404. [Google Scholar] [CrossRef]
- Stiller, A.; Garrison, K.; Gurdyumov, K.; Kenner, J.; Yasmin, F.; Yates, P.; Song, B.H. From Fighting Critters to Saving Lives: Polyphenols in Plant Defense and Human Health. Int. J. Mol. Sci. 2021, 22, 8995. [Google Scholar] [CrossRef] [PubMed]
- Boeckx, T.; Winters, A.L.; Webb, K.J.; Kingston-Smith, A.H. Polyphenol Oxidase in Leaves: Is There Any Significance to the Chloroplastic Localization? J. Exp. Bot. 2015, 66, 3571–3579. [Google Scholar] [CrossRef] [PubMed]
- Bapatla, R.B.; Saini, D.; Aswani, V.; Rajsheel, P.; Sunil, B.; Timm, S.; Raghavendra, A.S. Modulation of Photorespiratory Enzymes by Oxidative and Photo-Oxidative Stress Induced by Menadione in Leaves of Pea (Pisum sativum). Plants 2021, 10, 987. [Google Scholar] [CrossRef] [PubMed]
- Fernández, V.; Gil-Pelegrín, E.; Eichert, T. Foliar Water and Solute Absorption: An Update. Plant J. 2021, 105, 870–883. [Google Scholar] [CrossRef]
- Ferrarin, M.; Becagli, M.; Guglielmineeti, L.; Cardelli, R. Wood distillate, a review over past application and future perspective on soil and plant research. Agrochimica 2021, 65, 17–24. [Google Scholar] [CrossRef]
- Fedeli, R.; Vannini, A.; Celletti, S.; Maresca, V.; Munzi, S.; Cruz, C.; Alexandrov, D.; Guarnieri, M.; Loppi, S. Foliar Application of Wood Distillate Boosts Plant Yield and Nutritional Parameters of Chickpea. Ann. Appl. Biol. 2022, 1–8. [Google Scholar] [CrossRef]
- Meier, U. Growth Stages of Mono and Dicotyledonous Plants; BBCH Monograph; Julius Kuhn-Institut (JKI): Quedlinburg, Germany, 2018. [Google Scholar] [CrossRef]
- Colombo, C.; Miano, T. Metodi di Analisi Chimica del Suolo; Società Italiana della Scienza del Suolo (SISS): Modugno, Italy, 2015; ISBN 978-88-940679-0-3. [Google Scholar]
- Vance, E.D.; Brookes, P.C.; Jenkinson, D.S. An extraction method for measuring soil microbial biomass C. Soil Biol. Biochem. 1987, 19, 703–707. [Google Scholar] [CrossRef]
- Tabatabai, M. Soil enzymes. Methods Soil Anal. Microbiol. Biochem. Prop. 1994, 5 Pt 2, 775–833. [Google Scholar]
- Eivazi, F.; Tabatabai, M.A. Glucosidases and galactosidases in soils. Soil Biol. Biochem. 1988, 20, 601–606. [Google Scholar] [CrossRef]
- Eivazi, F.; Tabatabai, M.A. Phosphatases in soil. Soil Biol. Biochem. 1977, 9, 167–172. [Google Scholar] [CrossRef]
- Tabatabai, M.A.; Bremner, J.M. Arylsulfatase activity of soils. Soil Sci. Soc. Am. Pro. 1970, 34, 225–229. [Google Scholar] [CrossRef]
- Kandeler, E.; Gerber, H. Short-term assay of soil urease activity using colorimetric determination of ammonium. Biol. Fert. Soils 1988, 6, 68–72. [Google Scholar] [CrossRef]
- Yakhin, O.I.; Lubyanov, A.A.; Yakhin, I.A.; Brown, P.H. Biostimulants in plant science: A global perspective. Front. Plant Sci. 2017, 7, 2049. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Treatments | DOC (mg kg−1) | DOC (% TOC) |
|---|---|---|
| Foliar spray | ||
| F-0 | 192.5 a | 1.19 a |
| F-WD | 123.2 b | 1.13 b |
| Soil irrigation | ||
| S-0 | 122.3 b | 1.12 b |
| S-WD | 130.4 a | 1.20 a |
| Treatments | MB-C (mg kg−1) | MB-C/TOC (%) | ||
|---|---|---|---|---|
| S-0 | S-WD | S-0 | S-WD | |
| F-0 | 144 b | 158 a | 1.34 b | 1.45 a |
| F-WD | 143 b | 139 b | 1.32 b | 1.33 b |
| Treatments | S-0 | S-WD | S-0 | S-WD |
|---|---|---|---|---|
| APase (µg p-nitrophenol g−1 h−1) | β—glu (µg p-nitrophenol g−1 h−1) | |||
| F-0 | 321 b | 376 a | 51.0 b | 69.7 a |
| F-WD | 335 ab | 338 ab | 64.3 ab | 63.0 ab |
| Deh (µg TTF g−1 h−1) | Ure (µg NH4+-N g−1 2 h−1) | |||
| F-0 | 3.87 b | 5.61 a | 16 b | 24 a |
| F-WD | 4.82 ab | 5.26 ab | 20 ab | 21 ab |
| Treatments | SAI3 | |
|---|---|---|
| S-0 | S-WD | |
| F-0 | −16.4 b | −18.7 a |
| F-WD | −16.5 ab | −16.8 b |
| Treatment | Soil | Leaves | Pods | |||
|---|---|---|---|---|---|---|
| S-0 | S-WD | S-0 | S-WD | S-0 | S-WD | |
| NO3−-N (mg kg−1) | N concentration (%) | |||||
| F-0 | 4.3 b | 7.3 a | 4.1 b | 4.4 ab | 3.7 b | 4.1 a |
| F-WD | 4.9 b | 7.4 a | 4.6 a | 4.6 a | 3.8 b | 4.0 a |
| Available P (mg kg−1) | P concentration (mg g−1) | |||||
| F-0 | 7.3 b | 11.0 a | 3.3 b | 3.8 a | 3.2 a | 2.7 b |
| F-WD | 7.9 b | 8.6 ab | 3.3 b | 3.5 ab | 2.7 b | 2.7 b |
| Treatments | Stem Height | Fertile Nodes | 1st Fertile Node | Pods | ||
|---|---|---|---|---|---|---|
| (cm) | (n stem−1) | (Node Ranking) | (n stem−1) | (mg pod−1) | (g m−2) | |
| F-0 | 105.3 a | 7.3 b | 5.3 a | 9.0 b | 283.7 a | 299.1 a |
| F-WD | 95.8 b | 9.0 a | 4.6 a | 12.1 a | 189.9 b | 237.6 b |
| Treatments | Shoot (g plant−1) | Roots (g plant−1) | Root/Shoot Ratio | Nodule Number (n plant−1) | Nodule Mass (mg plant−1) | Nod. Density (n g root−1) | Nod./Root (mg g−1) |
|---|---|---|---|---|---|---|---|
| S-0 | 3.9 ± 0.27 | 2.1 ± 0.31 | 0.54 ± 0.07 | 170 ± 15.4 | 103 ± 19.4 | 81.2 ± 4.9 | 49.4 ± 3.8 |
| S-WD | 4.0 ± 0.49 | 1.9 ± 0.20 | 0.47 ± 0.07 | 168 ± 22.9 | 96.3 ± 13.4 | 89.8 ± 9.6 | 51.4 ± 5.1 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Becagli, M.; Arduini, I.; Cantini, V.; Cardelli, R. Soil and Foliar Applications of Wood Distillate Differently Affect Soil Properties and Field Bean Traits in Preliminary Field Tests. Plants 2023, 12, 121. https://doi.org/10.3390/plants12010121
Becagli M, Arduini I, Cantini V, Cardelli R. Soil and Foliar Applications of Wood Distillate Differently Affect Soil Properties and Field Bean Traits in Preliminary Field Tests. Plants. 2023; 12(1):121. https://doi.org/10.3390/plants12010121
Chicago/Turabian StyleBecagli, Michelangelo, Iduna Arduini, Valentina Cantini, and Roberto Cardelli. 2023. "Soil and Foliar Applications of Wood Distillate Differently Affect Soil Properties and Field Bean Traits in Preliminary Field Tests" Plants 12, no. 1: 121. https://doi.org/10.3390/plants12010121
APA StyleBecagli, M., Arduini, I., Cantini, V., & Cardelli, R. (2023). Soil and Foliar Applications of Wood Distillate Differently Affect Soil Properties and Field Bean Traits in Preliminary Field Tests. Plants, 12(1), 121. https://doi.org/10.3390/plants12010121