
Characterization of the Spanish Pomegranate Germplasm Collection Maintained at the Agricultural Experiment Station of Elche to Identify Promising Breeding Materials


Abstract
:1. Introduction
2. Results
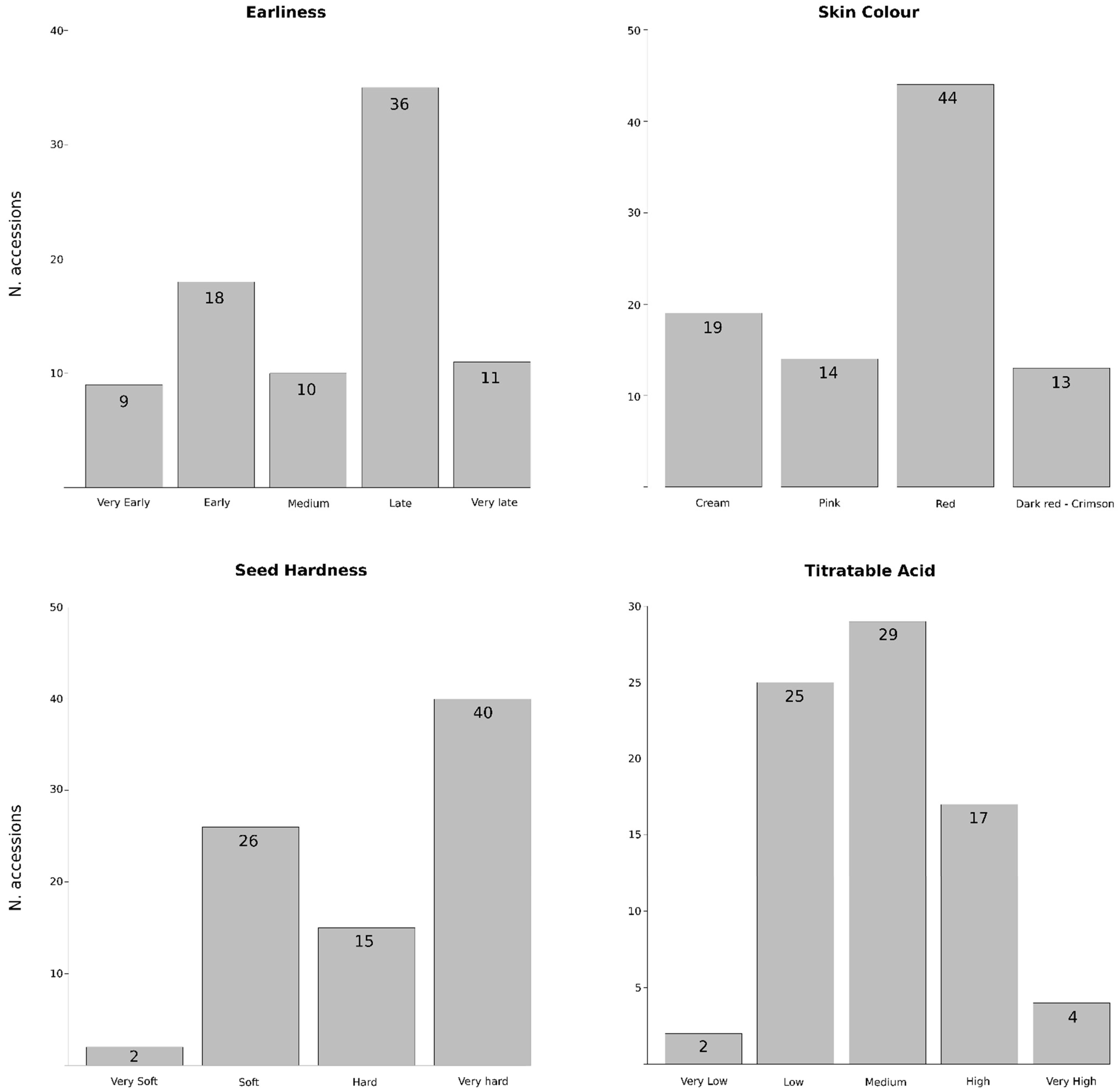
2.1. Germplasm Collection
2.2. Genetic Diversity, PIC and Cultivar Identification
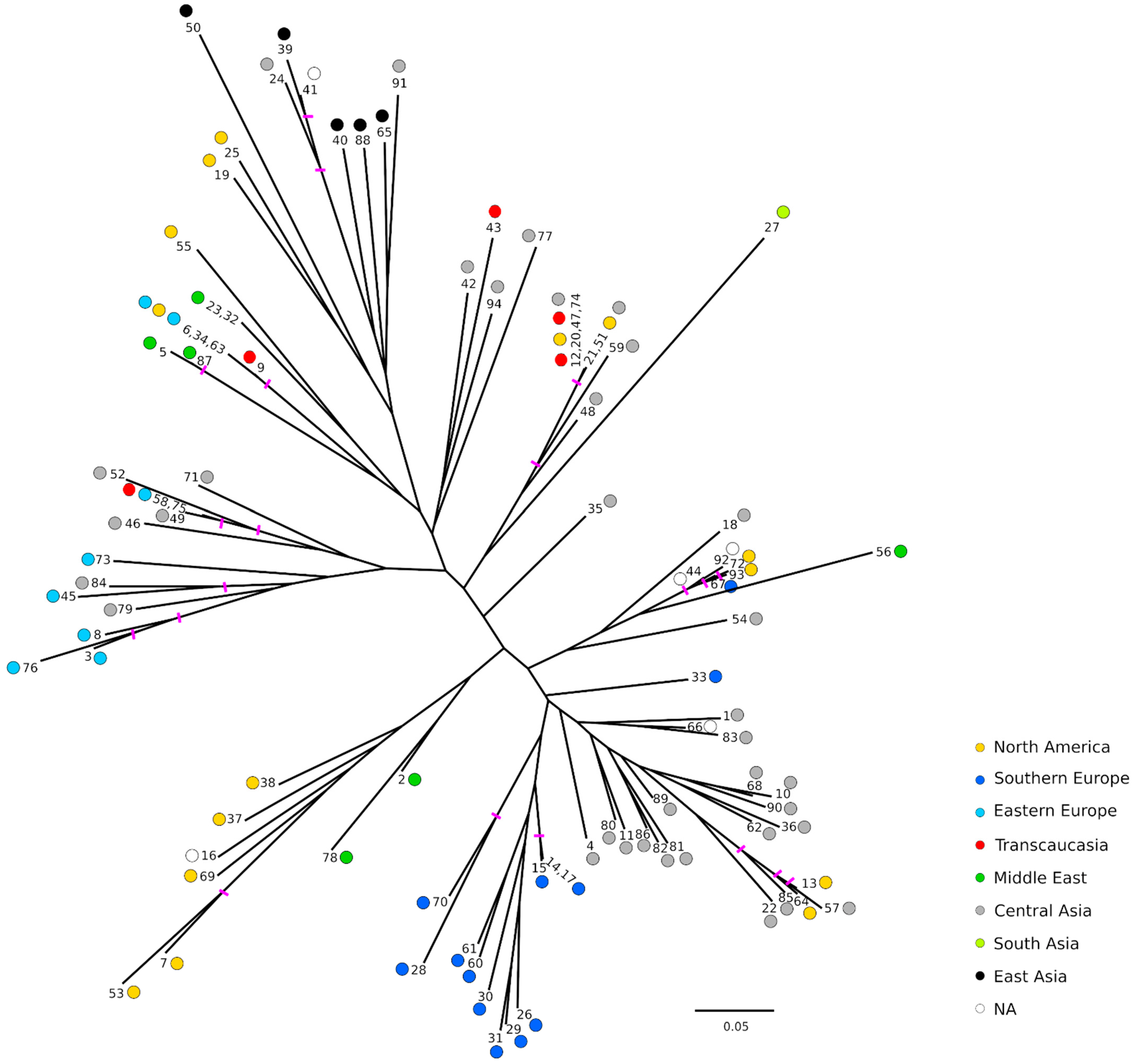
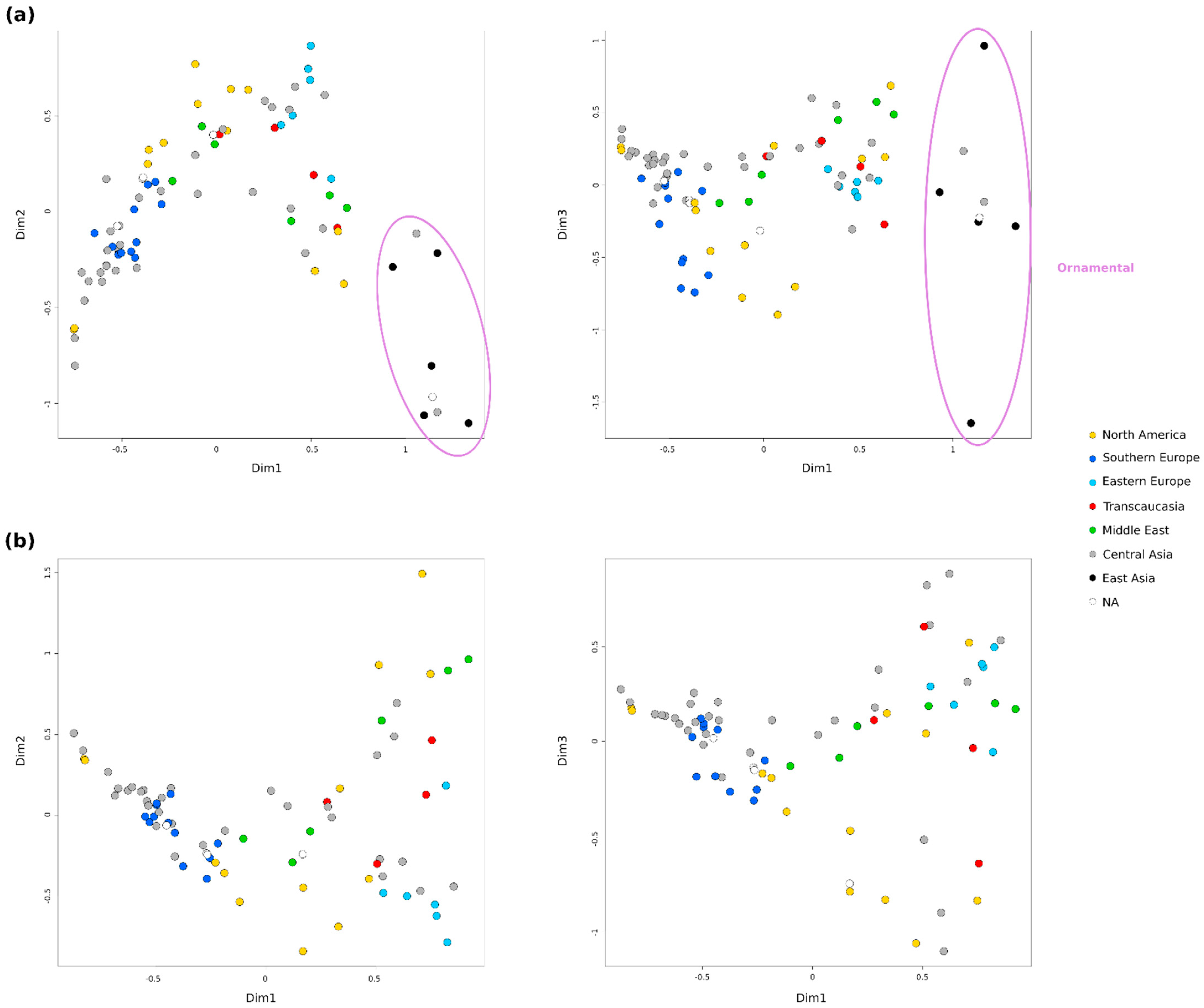
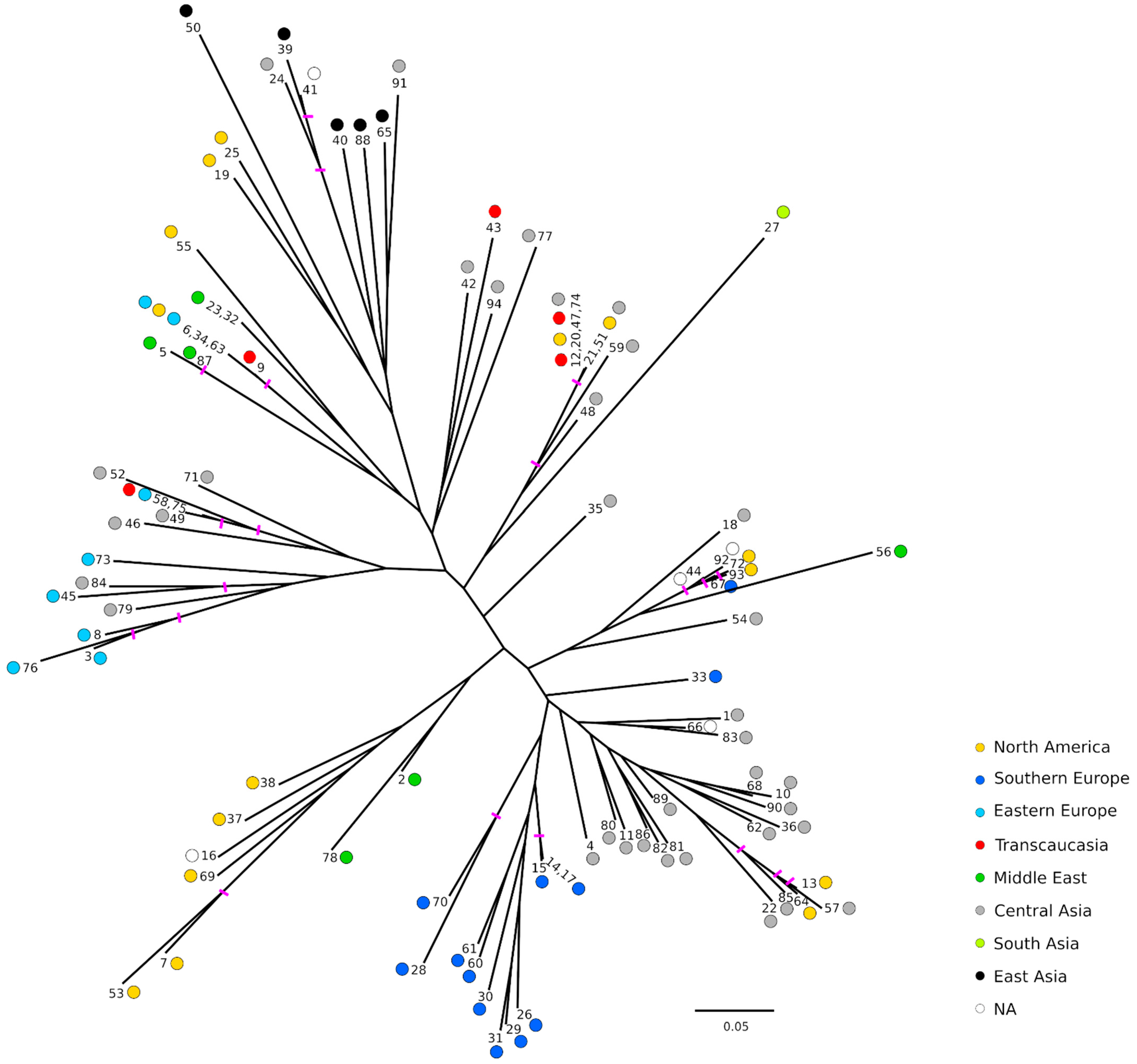
2.3. Population Structure and Genetic Relationships
2.4. Genetic Differentiation of Geographical Groups
3. Discussion
4. Materials and Methods
4.1. Plant Materials
4.2. Morphological and Pomological Characterization
4.3. DNA Isolation and Microsatellite Analysis
4.4. Data Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Rana, T.S.; Narzary, D.; Ranade, S.A. Systematics and taxonomic disposition of the genus Punica L. Fruit Veg. Cereal Sci. Biotechnol. 2010, 4, 51–55. [Google Scholar]
- Janick, J. The origins of fruits, fruit growing, and fruit breeding. Plant Breed. Rev. 2005, 25, 255–320. [Google Scholar] [CrossRef]
- Holland, D.; Hatib, K.; Bar-Ya’akov, I. Pomegranate: Botany, horticulture, breeding. Hortic. Rev. 2009, 35, 127–191. [Google Scholar] [CrossRef]
- Holland, D.; Bar-Ya’akov, I. Pomegranate (Punica granatum L.) breeding. In Advances in Plant Breeding Strategies: Fruits; Al-Khayri, J.M., Jain, S.M., Johnson, D.V., Eds.; Springer: Berlin/Heidelberg, Germany, 2018; pp. 601–647. [Google Scholar] [CrossRef]
- IPGRI. Regional Report CWANA 1999–2000. International Plant; Genetic Resources Institute: Rome, Italy, 2001; ISBN 92-9043-494-5. [Google Scholar]
- Caliskan, O.; Bayazıt, S. Morpho-pomological and chemical diversity of pomegranate accessions grown in eastern mediterranean region of turkey. J. Agric. Sci. Technol. 2013, 15, 1449–1460. [Google Scholar]
- Durgaç, C.; Özgen, M.; Simsek, Ö.; Kaçar, Y.A.; Kıyga, Y.; Çelebi, S.; Gündüz, K.; Serçe, S. Molecular and pomological diversity among pomegranate (Punica granatum L.) cultivars in Eastern Mediterranean region of Turkey. Afr. J. Biotechnol. 2008, 7, 1294–1301. [Google Scholar]
- Ferrara, G.; Cavoski, I.; Pacifico, A.; Tedone, L.; Mondelli, D. Morpho-pomological and chemical characterization of pomegranate (Punica granatum L.) genotypes in Apulia region, Southeastern Italy. Sci. Hortic. 2011, 130, 599–606. [Google Scholar] [CrossRef]
- Martinez-Nicolas, J.J.; Melgarejo, P.; Legua, P.; Garcia-Sanchez, F.; Hernández, F. Genetic diversity of pomegranate germplasm collection from Spain determined by fruit, seed, leaf and flower characteristics. PeerJ 2016, 4, e2214. [Google Scholar] [CrossRef] [Green Version]
- Varasteh, F.; Arzani, K.; Zamani, Z.; Mohseni, A. Evaluation of the most important fruit characteristics of some commercial pomegranate (Punica granatum L.) cultivars grown in Iran. Acta Hortic. 2009, 818, 103–108. [Google Scholar] [CrossRef]
- Drogoudi, P.D.; Tsipouridis, C.; Michailidis, Z. Physical and Chemical Characteristics of Pomegranates. Hortic. Sci. 2005, 40, 1200–1203. [Google Scholar] [CrossRef] [Green Version]
- Hasnaoui, N.; Jbir, R.; Mars, M.; Trifi, M.; Kamal-Eldin, A.; Melgarejo, P.; Hernandez, F. Organic acids, sugars, and anthocyanins contents in juices of tunisian pomegranate fruits. Int. J. Food Prop. 2011, 14, 741–757. [Google Scholar] [CrossRef] [Green Version]
- Still, D.W. Pomegranates: A botanical perspective. In Pomegranates: Ancient Roots to Modern Medicine; Seeram, N.P., Schulman, R.N., Heber, D., Eds.; Taylor & Francis: Boca Raton, FL, USA, 2006; pp. 199–209. [Google Scholar]
- Bartual, J.; Fernandez-Zamudio, M.A.; De-Miguel, M.D. Situation of the production, research and economics of the pomegranate industry in Spain. Acta Hortic. 2015, 1089, 345–349. [Google Scholar] [CrossRef]
- Soriano, J.M.; Zuriaga, E.; Rubio, P.; Llácer, G.; Infante, R.; Badenes, M.L. Development and characterization of microsatellite markers in pomegranate (Punica granatum L.). Mol. Breed. 2011, 27, 119–128. [Google Scholar] [CrossRef]
- Shahsavari, S.; Noormohammadi, Z.; Sheidai, M.; Farahani, F.; Vazifeshenas, M.R. Genetic structure, clonality and diversity in commercial pomegranate (Punica granatum L.) cultivars. Genet. Resour. Crop Evol. 2021, 68, 2943–2957. [Google Scholar] [CrossRef]
- Patil, P.G.; Singh, N.V.; Parashuram, S.; Bohra, A.; Mundewadikar, D.M.; Sangnure, V.R.; Babu, K.D.; Sharma, J. Genome wide identification, characterization and validation of novel miRNA-based SSR markers in pomegranate (Punica granatum L.). Physiol. Mol. Biol. Plants 2020, 26, 683–696. [Google Scholar] [CrossRef]
- Aziz, S.; Firdous, S.; Rahman, H.; Awan, S.I.; Michael, V.; Meru, G. Genetic diversity among wild pomegranate (Punica granatum) in Azad Jammu and Kashmir region of Pakistan. Electron. J. Biotechnol. 2020, 46, 50–54. [Google Scholar] [CrossRef]
- Višnjevec, A.M.; Ota, A.; Skrt, M.; Butinar, B.; Možina, S.S.; Cimerman, N.G.; Nečemer, M.; Arbeiter, A.B.; Hladnik, M.; Krapac, M.; et al. Genetic, biochemical, nutritional and antimicrobial characteristics of pomegranate (Punica granatum L.) grown in Istria. Food Technol. Biotechnol. 2017, 55, 151–163. [Google Scholar] [CrossRef]
- Ophir, R.; Sherman, A.; Rubinstein, M.; Eshed, R.; Sharabi Schwager, M.; Harel-Beja, R.; Bar-Ya’akov, I.; Holland, D. Single-nucleotide polymorphism markers from de-novo assembly of the pomegranate transcriptome reveal germplasm genetic diversity. PLoS ONE 2014, 9, e88998. [Google Scholar] [CrossRef] [Green Version]
- Nei, M. Estimation of average heterozygosity and genetic distance from a small number of individuals. Genetics 1978, 89, 583–590. [Google Scholar] [CrossRef]
- Bruvo, R.; Michiels, N.K.; D’souza, T.G.; Schulenburg, H. A simple method for the calculation of microsatellite genotype distances irrespective of ploidy level. Mol. Ecol. 2004, 13, 2101–2106. [Google Scholar] [CrossRef]
- Pritchard, J.K.; Stephens, M.; Donnelly, P. Inference of population structure using multilocus genotype data. Genetics 2000, 155, 945–959. [Google Scholar] [CrossRef]
- Evanno, G.; Regnaut, S.; Goudet, J. Detecting the number of clusters of individuals using the software STRUCTURE: A simulation study. Mol. Ecol. 2005, 14, 2611–2620. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Melgarejo, P.; Martínez-Valero, R. El Granado; Ediciones Mundi-Prensa: Madrid, Spain, 1992; p. 163. [Google Scholar]
- Giménez-Bastida, J.A.; Ávila-Gálvez, M.A.; Espín, J.C.; González-Sarrías, A. Evidence for health properties of pomegranate juices and extracts beyond nutrition: A critical systematic review of human studies. Trends Food Sci. Technol. 2021, 114, 410–423. [Google Scholar] [CrossRef]
- Stover, E.; Mercure, E.W. The Pomegranate: A New Look at the Fruit of Paradise. HortScience 2007, 42, 1088–1092. [Google Scholar] [CrossRef] [Green Version]
- Dandachi, F.; Hamadeh, B.; Youssef, H.; Chahine, H.; Chalak, L. Diversity assessment of the Lebanese germplasm of pomegranate (Punica granatum L.) by morphological and chemical traits. Ann. Agric. Sci. 2017, 62, 89–98. [Google Scholar] [CrossRef]
- Khadivi, A.; Arab, M. Identification of the superior genotypes of pomegranate (Punica granatum L.) using morphological and fruit characters. Food Sci. Nutr. 2021, 9, 4578–4588. [Google Scholar] [CrossRef]
- Vicent, A.; Mira, J.L.; Bartual, J.; Beltrán, V.; Taberner, V.; Palou, L. First Report of Black Heart of Pomegranate Caused by Alternaria alternata in Spain. Plant Dis. 2016, 100, 1952. [Google Scholar] [CrossRef]
- Doyle, J.J.; Doyle, J.L. A Rapid DNA Isolation Procedure for Small Quantities of Fresh Leaf Tissue. Phytochem. Bull. 1987, 19, 11–15. [Google Scholar]
- Schuelke, M. An economic method for the fluorescent labeling of PCR fragments. Nat. Biotechnol. 2000, 18, 233–234. [Google Scholar] [CrossRef]
- Belkhir, K.; Borsa, P.; Chikhi, L.; Raufaste, N.; Bonhomme, F. 1996–2004. GENETIX 4.05, Logiciel Sous Windows TM Pour la Génétique des Populations. Laboratoire Génome, Populations, Interactions, CNRS UMR 5171, Université de Montpellier II, Montpellier (France). Available online: https://kimura.univ-montp2.fr/genetix/ (accessed on 20 January 2021).
- Kamvar, Z.N.; Tabima, J.F.; Grünwald, N.J. Poppr: An R package for genetic analysis of populations with clonal, partially clonal, and/or sexual reproduction. PeerJ 2014, 2, e281. [Google Scholar] [CrossRef] [Green Version]
- Bingham, J.; Sudarsanam, S. Visualizing large hierarchical clusters in hyperbolic space. Bioinformatics 2000, 16, 660–661. [Google Scholar] [CrossRef] [Green Version]
- Earl, D.A.; vonHoldt, B.M. STRUCTURE HARVESTER: A website and program for visualizing STRUCTURE output and implementing the Evanno method. Conserv. Genet. Resour. 2012, 4, 359–361. [Google Scholar] [CrossRef]
- Jakobsson, M.; Rosenberg, N.A. CLUMPP: A cluster matching and permutation program for dealing with label switching and multimodality in analysis of population structure. Bioinformatics 2007, 23, 1801–1806. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ramasamy, R.K.; Ramasamy, S.; Bindroo, B.B.; Naik, V.G. STRUCTURE PLOT: A program for drawing elegant STRUCTURE bar plots in user friendly interface. SpringerPlus 2014, 3, 431. [Google Scholar] [CrossRef] [PubMed] [Green Version]


{kind=link}
{kind=link}
{kind=link}
| Acc | Name | Region | Country | Original Germplasm Bank |
|---|---|---|---|---|
| 1 | 15/4 Pamyati Rozanova | Central Asia | Turkmenistan | Davis, CA, USA |
| 2 | Acco | Middle East | Israel | Newe Ya’ar Research Center, Israel |
| 3 | Afganski | Eastern Europe | former Soviet Union | Davis, CA, USA |
| 4 | Agat | Central Asia | Turkmenistan | Davis, CA, USA |
| 5 | Alk Pust Ghermez Saveh | Middle East | Iran | Davis, CA, USA |
| 6 | Al-sirin-nar | Eastern Europe | former Soviet Union | Davis, CA, USA |
| 7 | Ambrosia | North America | United States | Davis, CA, USA |
| 8 | Apseronski | Eastern Europe | former Soviet Union | Davis, CA, USA |
| 9 | Apseronski krasnyj | Transcaucasia | former Soviet Union | Davis, CA, USA |
| 10 | Ariana | Central Asia | Turkmenistan | Davis, CA, USA |
| 11 | Azadi | Central Asia | Turkmenistan | Davis, CA, USA |
| 12 | Bala Miursal | Transcaucasia | former Soviet Union | Davis, CA, USA |
| 13 | Balegal | North America | USA | Davis, CA, USA |
| 14 | Borde-1 | Southern Europe | Spain | EEA d’Elx, Spain |
| 15 | Borde-2 | Southern Europe | Spain | EEA d’Elx, Spain |
| 16 | Cana | Unknown | Unknown | Davis, CA, USA |
| 17 | Casta del Reino | Southern Europe | Spain | EEA d’Elx, Spain |
| 18 | Chandyr | Central Asia | Turkmenistan | Davis, CA, USA |
| 19 | Chico | North America | United States | Davis, CA, USA |
| 20 | Crab | North America | United States | Davis, CA, USA |
| 21 | Cranberry | North America | United States | Davis, CA, USA |
| 22 | Desertnyi | Central Asia | Turkmenistan | Davis, CA, USA |
| 23 | Dorosht 5 hahanshahi Khoramabad | Middle East | Iran | Davis, CA, USA |
| 24 | Dotch Legrelley | Central Asia | Turkmenistan | Davis, CA, USA |
| 25 | Elf | North America | United States | Davis, CA, USA |
| 26 | Elx-11 | Southern Europe | Spain | EEA d’Elx, Spain |
| 27 | Elx-13 | South Asia | India | EEA d’Elx, Spain |
| 28 | Elx-6 | Southern Europe | Spain | EEA d’Elx, Spain |
| 29 | Elx-7 | Southern Europe | Spain | EEA d’Elx, Spain |
| 30 | Elx-8 | Southern Europe | Spain | EEA d’Elx, Spain |
| 31 | Elx-9 | Southern Europe | Spain | EEA d’Elx, Spain |
| 32 | Entek habi saveh | Middle East | Iran | Davis, CA, USA |
| 33 | Ermioni | Southern Europe | Greece | EEA d’Elx, Spain |
| 34 | Eve | North America | United States | Davis, CA, USA |
| 35 | Girkanets | Central Asia | Turkmenistan | Davis, CA, USA |
| 36 | Gissarskii Rozovyi | Central Asia | Turkmenistan | Davis, CA, USA |
| 37 | Golden Globe | North America | United States | Davis, CA, USA |
| 38 | Green Globe | North America | United States | Davis, CA, USA |
| 39 | Haku Batan | East Asia | Japan | Davis, CA, USA |
| 40 | Haku Taka | East Asia | Japan | Davis, CA, USA |
| 41 | How Sweet It Is | Unknown | Unknown | Davis, CA, USA |
| 42 | Hyrdanar x Goulosha | Central Asia | Turkmenistan | Davis, CA, USA |
| 43 | Hyrdanar x Kirmizy- Akbuh | Transcaucasia | former Soviet Union | Davis, CA, USA |
| 44 | Ink | Unknown | Unknown | Davis, CA, USA |
| 45 | Kaim Anor | Eastern Europe | former Soviet Union | Davis, CA, USA |
| 46 | Kaj Acik Anor | Central Asia | former Soviet Union | Davis, CA, USA |
| 47 | Kara bala miursal | Transcaucasia | former Soviet Union | Davis, CA, USA |
| 48 | Kara-Kalinskii | Central Asia | Turkmenistan | Davis, CA, USA |
| 49 | Kazake | Central Asia | former Soviet Union | Davis, CA, USA |
| 50 | Ki-Zakuro | East Asia | Japan | Davis, CA, USA |
| 51 | Koinekasyrskii Kislosladkii Krasnyi | Central Asia | Turkmenistan | Davis, CA, USA |
| 52 | Kunduzski | Central & South Asia | Afghanistan | Davis, CA, USA |
| 53 | Loffani | North America | United States | Davis, CA, USA |
| 54 | Machtumkuli | Central Asia | Turkmenistan | Davis, CA, USA |
| 55 | Mae | North America | United States | Davis, CA, USA |
| 56 | Mahali Dezful | Middle East | Iran | Davis, CA, USA |
| 57 | Medovyi Vahsha | Central Asia | Turkmenistan | Davis, CA, USA |
| 58 | Mejhos 6269 | Transcaucasia | former Soviet Union | Davis, CA, USA |
| 59 | Molla Nepes | Central Asia | Turkmenistan | Davis, CA, USA |
| 60 | Mollar-6 | Southern Europe | Spain | EEA d’Elx, Spain |
| 61 | Mollar-7 | Southern Europe | Spain | EEA d’Elx, Spain |
| 62 | Myatadzhy | Central Asia | Turkmenistan | Davis, CA, USA |
| 63 | Nikitski ranni | Eastern Europe | former Soviet Union | Davis, CA, USA |
| 64 | No name | North America | United States | Davis, CA, USA |
| 65 | Nochi shibori | East Asia | Japan | Davis, CA, USA |
| 66 | Orange | Unknown | Unknown | Davis, CA, USA |
| 67 | Palermo | Southern Europe | Italy | Davis, CA, USA |
| 68 | Parfyanka | Central Asia | Turkmenistan | Davis, CA, USA |
| 69 | Phoenicia | North America | United States | Davis, CA, USA |
| 70 | Piñon tierno Blanca | Southern Europe | Spain | EEA d’Elx, Spain |
| 71 | Podarok | Central Asia | Turkmenistan | Davis, CA, USA |
| 72 | Purple Heart | North America | United States | Davis, CA, USA |
| 73 | Saartuzski | Eastern Europe | former Soviet Union | Davis, CA, USA |
| 74 | Sakerdze | Central Asia | former Soviet Union | Davis, CA, USA |
| 75 | Salavatski | Eastern Europe | former Soviet Union | Davis, CA, USA |
| 76 | Sejanec 2–5/8 | Eastern Europe | former Soviet Union | Davis, CA, USA |
| 77 | Shainakskii | Central Asia | Turkmenistan | Davis, CA, USA |
| 78 | Shirin Pust Ghermez Saveh | Middle East | Iran | Davis, CA, USA |
| 79 | Shirin Zigar | Central Asia | Turkmenistan | Davis, CA, USA |
| 80 | Sirenevyi | Central Asia | Turkmenistan | Davis, CA, USA |
| 81 | Sogdiana | Central Asia | Turkmenistan | Davis, CA, USA |
| 82 | Sumbar | Central Asia | Turkmenistan | Davis, CA, USA |
| 83 | Sumbarskii | Central Asia | Turkmenistan | Davis, CA, USA |
| 84 | Surh-anor | Central Asia | former Soviet Union | Davis, CA, USA |
| 85 | Sverkhranniy | Central Asia | Turkmenistan | Davis, CA, USA |
| 86 | Syunt | Central Asia | Turkmenistan | Davis, CA, USA |
| 87 | Tabestani Males Biranden saveh | Middle East | Iran | Davis, CA, USA |
| 88 | Toryu-shibori | East Asia | Japan | Davis, CA, USA |
| 89 | Vishnevyi | Central Asia | Turkmenistan | Davis, CA, USA |
| 90 | Vkusnyi | Central Asia | Turkmenistan | Davis, CA, USA |
| 91 | White Flower | Central Asia | Turkmenistan | Davis, CA, USA |
| 92 | Wonderful-1 | Unknown | Unknown | Davis, CA, USA |
| 93 | Wonderful-2 | North America | United States | Davis, CA, USA |
| 94 | Zubejda | Central Asia | former Soviet Union | Davis, CA, USA |
| Locus | Melting Temp. (°C) | Fragment Length Range (bp) | No. Alleles | PIC | Ho | He |
|---|---|---|---|---|---|---|
| PGCT015 | 60 | 164–228 | 7 | 0.627 | 0.511 | 0.630 |
| PGCT016 | 57 | 196–204 | 3 | 0.607 | 0.538 | 0.611 |
| PGCT022 | 55 | 238–243 | 4 | 0.178 | 0.172 | 0.179 |
| PGCT028 | 57 | 228–240 | 4 | 0.407 | 0.352 | 0.409 |
| PGCT032 | 59 | 123–129 | 3 | 0.434 | 0.430 | 0.437 |
| PGCT038 | 57 | 265–284 | 5 | 0.460 | 0.380 | 0.460 |
| PGCT057 | 56 | 178–182 | 3 | 0.516 | 0.473 | 0.519 |
| PGCT066 | 59 | 124–131 | 3 | 0.481 | 0.355 | 0.483 |
| PGCT083 | 59 | 131–143 | 5 | 0.494 | 0.484 | 0.497 |
| PGCT087 | 58 | 253–263 | 5 | 0.671 | 0.457 | 0.674 |
| PGCT088A | 59 | 148–169 | 5 | 0.604 | 0.451 | 0.607 |
| PGCT088B | 59 | 246–269 | 5 | 0.596 | 0.348 | 0.600 |
| PGCT089 | 59 | 134–154 | 7 | 0.603 | 0.511 | 0.606 |
| PGCT091 | 56 | 203–223 | 9 | 0.638 | 0.462 | 0.642 |
| PGCT093A | 58 | 231–254 | 7 | 0.658 | 0.554 | 0.662 |
| PGCT093B | 60 | 182–208 | 7 | 0.599 | 0.511 | 0.602 |
| PGCT098 | 60 | 114–146 | 4 | 0.365 | 0.319 | 0.367 |
| PGCT110 | 52 | 104–188 | 13 | 0.618 | 0.500 | 0.622 |
| PGCT111 | 58 | 206–248 | 6 | 0.653 | 0.495 | 0.656 |
| Group | Accession Number (Region) |
|---|---|
| G1 | 6 (EE), 34 (NA), 63 (EE) |
| G2 | 12 (TR), 20 (NA), 47 (TR), 74 (CA) |
| G3 | 21 (NA), 51 (CA) |
| G4 | 23 (ME), 32 (ME) |
| G5 | 58 (TR), 75 (EE) |
| G6 | 14 (SE), 17(SE) |
| Group | No. Accs | MLG | eMLG | A | Exclusive Alleles | Hobs | Hexp | Corrected Simpson’s Index |
|---|---|---|---|---|---|---|---|---|
| North America | 15 | 15 | 10.00 | 3.74 | 5 | 0.370 (0.110) | 0.548 | 1.000 |
| Southern Europe | 13 | 12 | 9.42 | 2.47 | 2 | 0.531 (0.258) | 0.440 | 0.987 |
| Eastern Europe | 8 | 7 | 7.00 | 2.63 | 0 | 0.428 (0.284) | 0.410 | 0.964 |
| Transcaucasia | 5 | 4 | 4.00 | 2.63 | 0 | 0.558 (0.356) | 0.483 | 0.900 |
| Middle East | 7 | 6 | 6.00 | 2.42 | 0 | 0.471 (0.245) | 0.479 | 0.952 |
| Central Asia | 35 | 35 | 10.00 | 4.32 | 11 | 0.414 (0.116) | 0.486 | 1.000 |
| South Asia | 1 | 1 | 1.00 | 1.32 | 5 | 0.316 (0.478) | 0.316 | NA |
| East Asia | 5 | 5 | 5.00 | 2.84 | 6 | 0.463 (0.291) | 0.488 | 1.000 |
| Unknown | 5 | 5 | 5.00 | 2.84 | 0 | 0.421 (0.266) | 0.473 | 1.000 |
| Total | 94 | 85 | 9.87 | 5.47 | 29 | 0.540 | 0.997 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zuriaga, E.; Pintová, J.; Bartual, J.; Badenes, M.L. Characterization of the Spanish Pomegranate Germplasm Collection Maintained at the Agricultural Experiment Station of Elche to Identify Promising Breeding Materials. Plants 2022, 11, 1257. https://doi.org/10.3390/plants11091257
Zuriaga E, Pintová J, Bartual J, Badenes ML. Characterization of the Spanish Pomegranate Germplasm Collection Maintained at the Agricultural Experiment Station of Elche to Identify Promising Breeding Materials. Plants. 2022; 11(9):1257. https://doi.org/10.3390/plants11091257
Chicago/Turabian StyleZuriaga, Elena, Jitka Pintová, Julián Bartual, and María Luisa Badenes. 2022. "Characterization of the Spanish Pomegranate Germplasm Collection Maintained at the Agricultural Experiment Station of Elche to Identify Promising Breeding Materials" Plants 11, no. 9: 1257. https://doi.org/10.3390/plants11091257
APA StyleZuriaga, E., Pintová, J., Bartual, J., & Badenes, M. L. (2022). Characterization of the Spanish Pomegranate Germplasm Collection Maintained at the Agricultural Experiment Station of Elche to Identify Promising Breeding Materials. Plants, 11(9), 1257. https://doi.org/10.3390/plants11091257