
Novel QTL for Low Seed Cadmium Accumulation in Soybean

 and
and 
Abstract
:1. Introduction
2. Results
2.1. Population Phenotyping
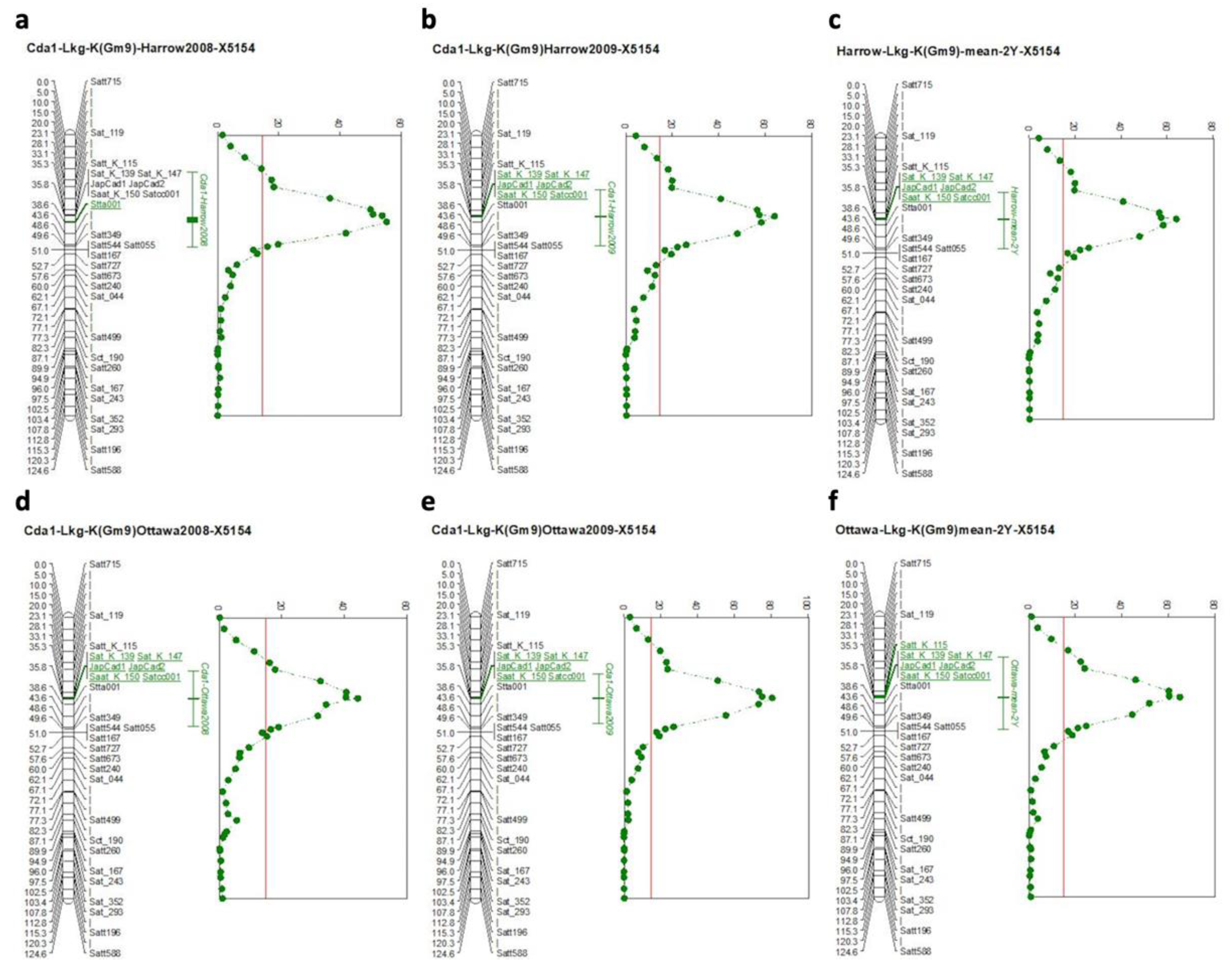
2.2. Genetic Map and QTL Results
2.3. Candidate Gene Search
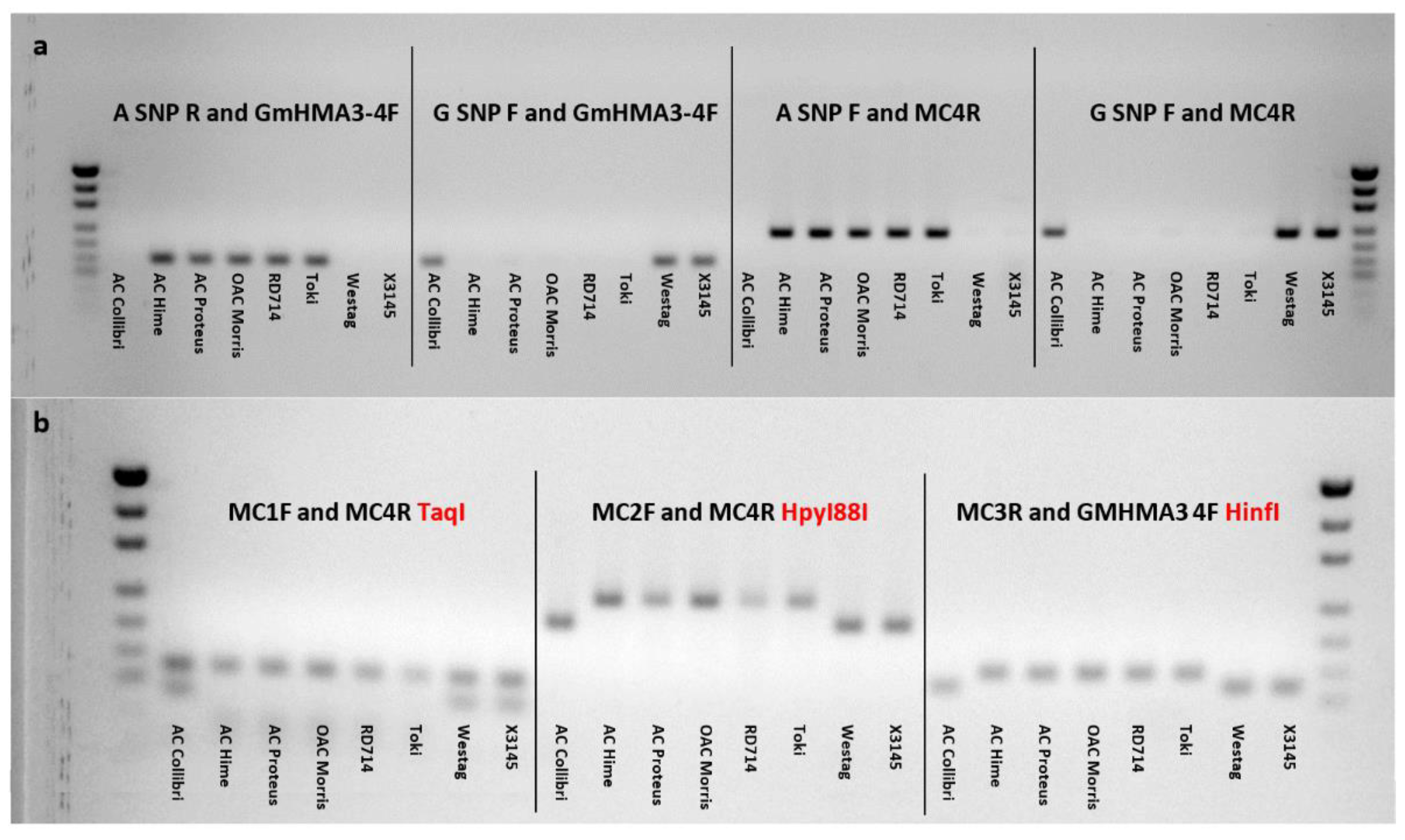
2.4. Allele-Specific Marker Development for Cda1
3. Discussion
4. Materials and Methods
4.1. Population Development and Field Evaluation
4.2. Sample Preparation and Cd Analysis
4.3. DNA Extraction, SSR Markers and Primer/SSR and PCR Conditions
4.4. Genetic Map Construction and QTL Analysis
4.5. Candidate Gene Search
4.6. Marker Development for Cda1
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Dhaliwal, S.S.; Singh, J.; Taneja, P.K.; Mandal, A. Remediation Techniques for Removal of Heavy Metals from the Soil Contaminated through Different Sources: A Review. Environ. Sci. Pollut. Res. 2020, 27, 1319–1333. [Google Scholar] [CrossRef] [PubMed]
- Pan, L.; Ma, J.; Wang, X.; Hou, H. Chemosphere Heavy Metals in Soils from a Typical County in Shanxi Province, China: Levels, Sources and Spatial Distribution. Chemosphere 2016, 148, 248–254. [Google Scholar] [CrossRef] [PubMed]
- Baliardini, C.; Meyer, C.L.; Salis, P.; Saumitou-Laprade, P.; Verbruggen, N. CATION EXCHANGER1 Cosegregates with Cadmium Tolerance in the Metal Hyperaccumulator Arabidopsis Halleri and Plays a Role in Limiting Oxidative Stress in Arabidopsis Spp. Plant Physiol. 2015, 169, 549–559. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Borges, K.L.R.; Salvato, F.; Alcântara, B.K.; Nalin, R.S.; Piotto, F.Â.; Azevedo, R.A. Temporal Dynamic Responses of Roots in Contrasting Tomato Genotypes to Cadmium Tolerance. Ecotoxicology 2018, 27, 245–258. [Google Scholar] [CrossRef]
- Fattahi, B.; Arzani, K.; Souri, M.K.; Barzegar, M. Effects of Cadmium and Lead on Seed Germination, Morphological Traits, and Essential Oil Composition of Sweet Basil (Ocimum Basilicum L.). Ind. Crops Prod. 2019, 138, 111584. [Google Scholar] [CrossRef]
- Shah, V.; Daverey, A. Phytoremediation: A Multidisciplinary Approach to Clean up Heavy Metal Contaminated Soil. Environ. Technol. Innov. 2020, 18, 100774. [Google Scholar] [CrossRef]
- Kumar, S.; Sharma, A. Cadmium Toxicity: Effects on Human Reproduction and Fertility. Rev. Environ. Health 2019, 34, 327–338. [Google Scholar] [CrossRef]
- Huff, J.; Lunn, R.M.; Waalkes, M.P.; Tomatis, L.; Infante, P.F. Cadmium-Induced Cancers in Animals and in Humans. Int. J. Occup. Environ. Health 2007, 13, 202–212. [Google Scholar] [CrossRef] [Green Version]
- Djordjevic, V.R.; Wallace, D.R.; Schweitzer, A.; Boricic, N.; Knezevic, D.; Matic, S.; Grubor, N.; Kerkez, M.; Radenkovic, D.; Bulat, Z.; et al. Environmental Cadmium Exposure and Pancreatic Cancer: Evidence from Case Control, Animal and in vitro Studies. Environ. Int. 2019, 128, 353–361. [Google Scholar] [CrossRef]
- Baba, H.; Tsuneyama, K.; Yazaki, M.; Nagata, K.; Minamisaka, T.; Tsuda, T.; Nomoto, K.; Hayashi, S.; Miwa, S.; Nakajima, T.; et al. The Liver in Itai-Itai Disease (Chronic Cadmium Poisoning): Pathological Features and Metallothionein Expression. Mod. Pathol. 2013, 26, 1228–1234. [Google Scholar] [CrossRef]
- Grant, C.A.; Clarke, J.M.; Duguid, S.; Chaney, R.L. Selection and Breeding of Plant Cultivars to Minimize Cadmium Accumulation. Sci. Total Environ. 2008, 390, 301–310. [Google Scholar] [CrossRef] [PubMed]
- Zook, E.G.; Greene, F.E.; Morris, E.R. Nutrient Composition of Selected Wheats and Wheat Products. VI. Distribution of Manganese, Copper, Nickel, Zinc, Magnesium, Lead, Tin, Cadmium, Chromium, and Selenium as Determined by Atomic Absorption Spectroscopy and Colorimetry. Cereal Chem. 1970, 47, 720–731. [Google Scholar]
- Petersen, A. Expert Advice on Appropriate Criteria and Limits for Contaminants in Ready to Use Therapeutic Foods (RUTF). 2018. Available online: https://www.fao.org/fao-who-codexalimentarius/sh-proxy/en/?lnk=1&url=https%253A%252F%252Fworkspace.fao.org%252Fsites%252Fcodex%252FMeetings%252FCX-720-40%252FMisc%252FFinal%2BReport%2Bcontaminants%2Bin%2BReady%2Bto%2BUse%2BTherapeutic%2BFoods%2B%2528RUFT%2529%2BUNICEF%2B-WFP.pdf (accessed on 8 April 2022).
- WHO. Report of the 33rd Session of the Codex Committee on Food Additives and Contaminants, FAO, The Hague, The Netherlands, 12–16 March 2001. 2001. Available online: https://agris.fao.org/agris-search/search.do?recordID=XF2016007533 (accessed on 8 April 2022).
- Smolders, E. Revisiting and Updating the Effect of Phosphorus Fertilisers on Cadmium Accumulation in European Agricultural Soils. Proc. Int. Fertil. Society. Int. Fertil. Soc. 2013, 724, 1–33. [Google Scholar]
- Yang, Z. Cadmium and Lead Low Accumulation among Chinese Soybean Cultivars and the Evaluation of Quality Difference and Correlation. Ph.D. Thesis, Northeastern University, Shenyang, China, 2015. Available online: https://global.cnki.net/kcms/detail/detail.aspx?dbcode=CDFD&dbname=CDFD2017&filename=1016011464.nh (accessed on 8 April 2022).
- Zhang, S.; Song, J.; Wu, L.; Chen, Z. Worldwide Cadmium Accumulation in Soybean Grains and Feasibility of Food Prouction on Contaminated Calcareous Soils. Environ. Pollut. 2021, 269, 116153. [Google Scholar] [CrossRef]
- Sun, X.; Lan, J.; Zhang, R. Study on Residues of Heavy Metals in Heilongjiang Soybean and Its Dietary Risk. J. Agric. Prod. Qual. Saf. 2019, 5, 49–57. [Google Scholar]
- Zhi, Y.; Sun, T.; Zhou, Q.; Leng, X. Screening of Safe Soybean Cultivars for Cadmium Contaminated Fields. Sci. Rep. 2020, 10, 12965. [Google Scholar] [CrossRef]
- Dong, W.Q.Y.; Cui, Y.; Liu, X. Instances of Soil and Crop Heavy Metal Contamination in China. Soil Sediment Contam. An Int. J. 2001, 10, 497–510. [Google Scholar] [CrossRef]
- Liu, J.; Qian, M.; Cai, G.; Yang, J.; Zhu, Q. Uptake and Translocation of Cd in Different Rice Cultivars and the Relation with Cd Accumulation in Rice Grain. J. Hazard. Mater. 2007, 143, 443–447. [Google Scholar] [CrossRef]
- Morrison, M.J.; Cober, E.; Poysa, V.; Gawalko, E. Soil Cadmium Concentration Influences Soybean Seed Concentration. In Proceedings of the Joint Meeting of the CSA and CSSA, Joint Annual Meeting of the Canadian Society of Agronomy and the Canadian Society of Soil Science, Guelph, ON, Canada, 5–7 August 2009; University of Guelph: Guelph, ON, Canada, 2009. Available online: https://agronomycanada.com/joint-annual-meeting-2009/ (accessed on 8 April 2022).
- Morrison, M.J.; Mongeon, V.; Cober, E.; Poysa, V.; Gawalko, E. A Method for Selecting Low Cadmium (Cd) Accumulating Soybean Varieties. In Proceedings of the Joint Meeting of the CSA and CSSA, Joint Annual Meeting of the Canadian Society of Agronomy and the Canadian Society of Soil Science, Saskatoon, SK, Canada, 20–24 June 2010; University of Saskatchewan: Saskatoon, SK, Canada, 2010. Available online: https://agronomycanada.com/2010-joint-annual-meeting/ (accessed on 8 April 2022).
- Jegadeesan, S.; Yu, K.; Poysa, V.; Gawalko, E.; Morrison, M.J.; Shi, C.; Cober, E. Mapping and Validation of Simple Sequence Repeat Markers Linked to a Major Gene Controlling Seed Cadmium Accumulation in Soybean [Glycine Max (L.) Merr]. Theor. Appl. Genet. 2010, 121, 283–294. [Google Scholar] [CrossRef]
- Benitez, E.R.; Hajika, M.; Yamada, T.; Takahashi, K.; Oki, N.; Yamada, N.; Nakamura, T.; Kanamaru, K. A Major QTL Controlling Seed Cadmium Accumulation in Soybean. Crop Sci. 2010, 50, 1728–1734. [Google Scholar] [CrossRef]
- Bradley, D.; Carpenter, R.; Copsey, L.; Vincent, C.; Rothstein, S.; Coen, E. Control of inflorescence architecture in Antirrhinum. Nature 1996, 379, 791–797. [Google Scholar] [CrossRef] [PubMed]
- Bi, X.; van Esse, W.; van Esse, W.; Mulki, M.A.; Kirschner, G.; Zhong, J.; Simon, R.; von Korff, M. Centroradialis Interacts with Flowering Locus T-like Genes to Control Floret Development and Grain Number. Plant Physiol. 2019, 180, 1013–1030. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Danilevskaya, O.N.; Meng, X.; Hou, Z.; Ananiev, E.V.; Simmons, C.R. A Genomic and Expression Compendium of the Expanded PEBP Gene Family from Maize. Plant Physiol. 2008, 146, 250–264. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miyata, S.I.; Urao, T.; Yamaguchi-Shinozaki, K.; Shinozaki, K. Characterization of Genes for Two-Component Phosphorelay Mediators with a Single HPt Domain in Arabidopsis Thaliana. FEBS Lett. 1998, 437, 11–14. [Google Scholar] [CrossRef] [Green Version]
- Urao, T.; Yakubov, B.; Yamaguchi-Shinozaki, K.; Shinozaki, K. Stress-Responsive Expression of Genes for Two-Component Response Regulator-like Proteins in Arabidopsis Thaliana. FEBS Lett. 1998, 427, 175–178. [Google Scholar] [CrossRef] [Green Version]
- Hwang, I.; Chen, H.-C.; Sheen, J. Two-Component Signal Transduction Pathways in Arabidopsis. Plant Physiol. 2002, 129, 500–515. [Google Scholar] [CrossRef] [Green Version]
- Atencio, L.; Salazar, J.; Moran Lauter, A.N.; Gonzales, M.D.; O’Rourke, J.A.; Graham, M.A. Characterizing Short and Long Term Iron Stress Responses in Iron Deficiency Tolerant and Susceptible Soybean (Glycine Max L. Merr.). Plant Stress 2021, 2, 100012. [Google Scholar] [CrossRef]
- Colangelo, E.P.; Guerinot, M. Lou. The Essential Basic Helix-Loop-Helix Protein FIT1 Is Required for the Iron Deficiency Response. Plant Cell 2004, 16, 3400–3412. [Google Scholar] [CrossRef] [Green Version]
- Chen, J.; Li, L.G.; Liu, Z.H.; Yuan, Y.J.; Guo, L.L.; Mao, D.D.; Tian, L.F.; Chen, L.B.; Luan, S.; Li, D.P. Magnesium Transporter AtMGT9 Is Essential for Pollen Development in Arabidopsis. Cell Res. 2009, 19, 887–898. [Google Scholar] [CrossRef]
- Lusk, J.E.; Kennedy, E.P. Magneisum Transport in Escherichia Coli. J. Biol. Chem. 1969, 244, 1653–1655. [Google Scholar] [CrossRef]
- Snavely, M.D.; Florer, J.B.; Miller, C.G.; Maguire, M.E. Magnesium Transport in Salmonella Typhimurium: Expression of Cloned Genes for Three Distinct Mg2+ Transport Systems. J. Bacteriol. 1989, 171, 4752–4760. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, L.; Tutone, A.F.; Drummond, R.S.M.; Gardner, R.C.; Luan, S. A Novel Family of Magnesium Transport Genes in Arabidopsis. Plant Cell 2001, 13, 2761–2775. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schock, I.; Gregan, J.; Steinhauser, S.; Schweyen, R.; Brennicke, A.; Knoop, V. A Member of a Novel Arabidopsis Thaliana Gene Family of Candidate Mg2+ Ion Transporters Complements a Yeast Mitochondrial Group II Intron-Splicing Mutant. Plant J. 2000, 24, 489–501. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.Z.; Chen, Y.; Sun, Z.H.; Zhou, Q.; Sui, S.F. Escherichia Coli CorA Periplasmic Domain Functions as a Homotetramer to Bind Substrate. J. Biol. Chem. 2006, 281, 26813–26820. [Google Scholar] [CrossRef] [Green Version]
- Smith, R.L.; Maguire, M.E. Microbial Magnesium Transport: Unusual Transporters Searching for Identity. Mol. Microbiol. 1998, 28, 217–226. [Google Scholar] [CrossRef]
- Moncrief, M.B.; Maguire, M.E. Magnesium Transport in Prokaryotes. JBIC J. Biol. Inorg. Chem. 1999, 4, 523–527. [Google Scholar] [CrossRef]
- Coleman, Z.; Boelter, J.; Espinosa, K.; Goggi, A.S.; Palmer, R.G.; Sandhu, D. Isolation and Characterization of the Aconitate Hydratase 4 (Aco4) Gene from Soybean. Can. J. Plant Sci. 2017, 97, 684–691. [Google Scholar] [CrossRef]
- Matsushima, A.; Sato, Y.; Otsuka, M.; Watanabe, T.; Yamamoto, H.; Hirata, T. An Enone Reductase from Nicotiana Tabacum: CDNA Cloning, Expression in Escherichia Coli, and Reduction of Enones with the Recombinant Proteins. Bioorg. Chem. 2008, 36, 23–28. [Google Scholar] [CrossRef]
- Youn, B.; Kim, S.J.; Moinuddin, S.G.A.; Lee, C.; Bedgar, D.L.; Harper, A.R.; Davin, L.B.; Lewis, N.G.; Kang, C.H. Mechanistic and Structural Studies of Apoform, Binary, and Ternary Complexes of the Arabidopsis Alkenal Double Bond Reductase At5g16970. J. Biol. Chem. 2006, 281, 40076–40088. [Google Scholar] [CrossRef] [Green Version]
- Kasahara, H.; Jiao, Y.; Bedgar, D.L.; Kim, S.-J.; Patten, A.M.; Xia, Z.-Q.; Davin, L.B.; Lewis, N.G. Pinus Taeda Phenylpropenal Double-Bond Reductase: Purification, CDNA Cloning, Heterologous Expression in Escherichia Coli, and Subcellular Localization in P. Taeda. Phytochemistry 2006, 67, 1765–1780. [Google Scholar] [CrossRef]
- Lewis, N.G.; Davin, L.B. 1.25—Lignans: Biosynthesis and Function; Barton, S.D., Nakanishi, K., Meth-Cohn, O., Eds.; Pergamon: Oxford, UK, 1999; pp. 639–712. [Google Scholar] [CrossRef]
- Davin, L.B.; Lewis, N.G. Dirigent Phenoxy Radical Coupling: Advances and Challenges. Curr. Opin. Biotechnol. 2005, 16, 398–406. [Google Scholar] [CrossRef] [PubMed]
- Voldeng, H.D.; Guillemette, R.J.D.; Leonard, D.A.; Cober, E.R. AC Brant Soybean. Can. J. Plant Sci. 1996, 76, 149–150. [Google Scholar] [CrossRef]
- Samanfar, B.; Cober, E.R.; Charette, M.; Tan, L.H.; Bekele, W.A.; Morrison, M.J.; Kilian, A.; Belzile, F.; Molnar, S.J. Genetic Analysis of High Protein Content in ‘AC Proteus’ Related Soybean Populations Using SSR, SNP, DArT and DArTseq Markers. Sci. Rep. 2019, 9, 19657. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Voldeng, H.D.; Fregeau-Reid, J.A.; Guillemette, R.J.D.; Leonard, D.A.; Cober, E.R. AC Colibri Soybean. Can. J. Plant Sci. 1997, 77, 113–114. [Google Scholar] [CrossRef] [Green Version]
- Gawalko, E.; Garrett, R.G.; Warkentin, T.; Wang, N.; Richter, A. Trace Elements in Canadian Field Peas: A Grain Safety Assurance Perspective. Food Addit. Contam. Part A 2009, 26, 1002–1012. [Google Scholar] [CrossRef]
- Molnar, S.J.; Rai, S.; Charette, M.; Cober, E.R. Simple Sequence Repeat (SSR) Markers Linked to E1, E3, E4, and E7 Maturity Genes in Soybean. Genome 2003, 46, 1024–1036. [Google Scholar] [CrossRef]
- Cregan, P.; Jarvik, T.; Bush, A.; Shoemaker, R.; Lark, K.; Kahler, A.; Kaya, N.; Van Toai, T.; Lohnes, D.; Chung, J.; et al. An Integrated Genetic Linkage Map of the Soybean Genome. Crop Sci. 1999, 39, 1464–1490. [Google Scholar] [CrossRef] [Green Version]
- Lander, E.S.; Green, P.; Abrahamson, J.; Barlow, A.; Daly, M.J.; Lincoln, S.E.; Newberg, L.A. MAPMAKER: An Interactive Computer Package for Constructing Primary Genetic Linkage Maps of Experimental and Natural Populations. Genomics 1987, 1, 174–181. [Google Scholar] [CrossRef]
- Morales, A.M.A.P.; O’Rourke, J.A.; van de Mortel, M.; Scheider, K.T.; Bancroft, T.J.; Borém, A.; Nelson, R.T.; Nettleton, D.; Baum, T.J.; Shoemaker, R.C.; et al. Transcriptome Analyses and Virus Induced Gene Silencing Identify Genes in the Rpp4-Mediated Asian Soybean Rust Resistance Pathway. Funct. Plant Biol. 2013, 40, 1029–1047. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| GO_Id | GO_Description | Gene_Id (Wm82.a2.v1) |
|---|---|---|
| GO:0000041 | Transition metal ion transport | Glyma.13G308000 |
| Glyma.13G317100 | ||
| Glyma.13G317900 | ||
| Glyma.13G322100 | ||
| GO:0046686 | Response to cadmium ion | Glyma.13G308700 |
| Glyma.13G309100 | ||
| GO:0030001 | Metal ion transport | Glyma.13G320800 |
| GO:0070838 | Divalent metal ion transport | Glyma.13G320800 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nissan, N.; Hooker, J.; Pattang, A.; Charette, M.; Morrison, M.; Yu, K.; Hou, A.; Golshani, A.; Molnar, S.J.; Cober, E.R.; et al. Novel QTL for Low Seed Cadmium Accumulation in Soybean. Plants 2022, 11, 1146. https://doi.org/10.3390/plants11091146
Nissan N, Hooker J, Pattang A, Charette M, Morrison M, Yu K, Hou A, Golshani A, Molnar SJ, Cober ER, et al. Novel QTL for Low Seed Cadmium Accumulation in Soybean. Plants. 2022; 11(9):1146. https://doi.org/10.3390/plants11091146
Chicago/Turabian StyleNissan, Nour, Julia Hooker, Arezo Pattang, Martin Charette, Malcolm Morrison, Kangfu Yu, Anfu Hou, Ashkan Golshani, Stephen J. Molnar, Elroy R. Cober, and et al. 2022. "Novel QTL for Low Seed Cadmium Accumulation in Soybean" Plants 11, no. 9: 1146. https://doi.org/10.3390/plants11091146
APA StyleNissan, N., Hooker, J., Pattang, A., Charette, M., Morrison, M., Yu, K., Hou, A., Golshani, A., Molnar, S. J., Cober, E. R., & Samanfar, B. (2022). Novel QTL for Low Seed Cadmium Accumulation in Soybean. Plants, 11(9), 1146. https://doi.org/10.3390/plants11091146