Incidence and Molecular Identification of Begomoviruses Infecting Tomato and Pepper in Myanmar

 , ,
, ,
Abstract
1. Introduction
2. Results
2.1. Detection of Begomoviruses in Tomato and Pepper Plants
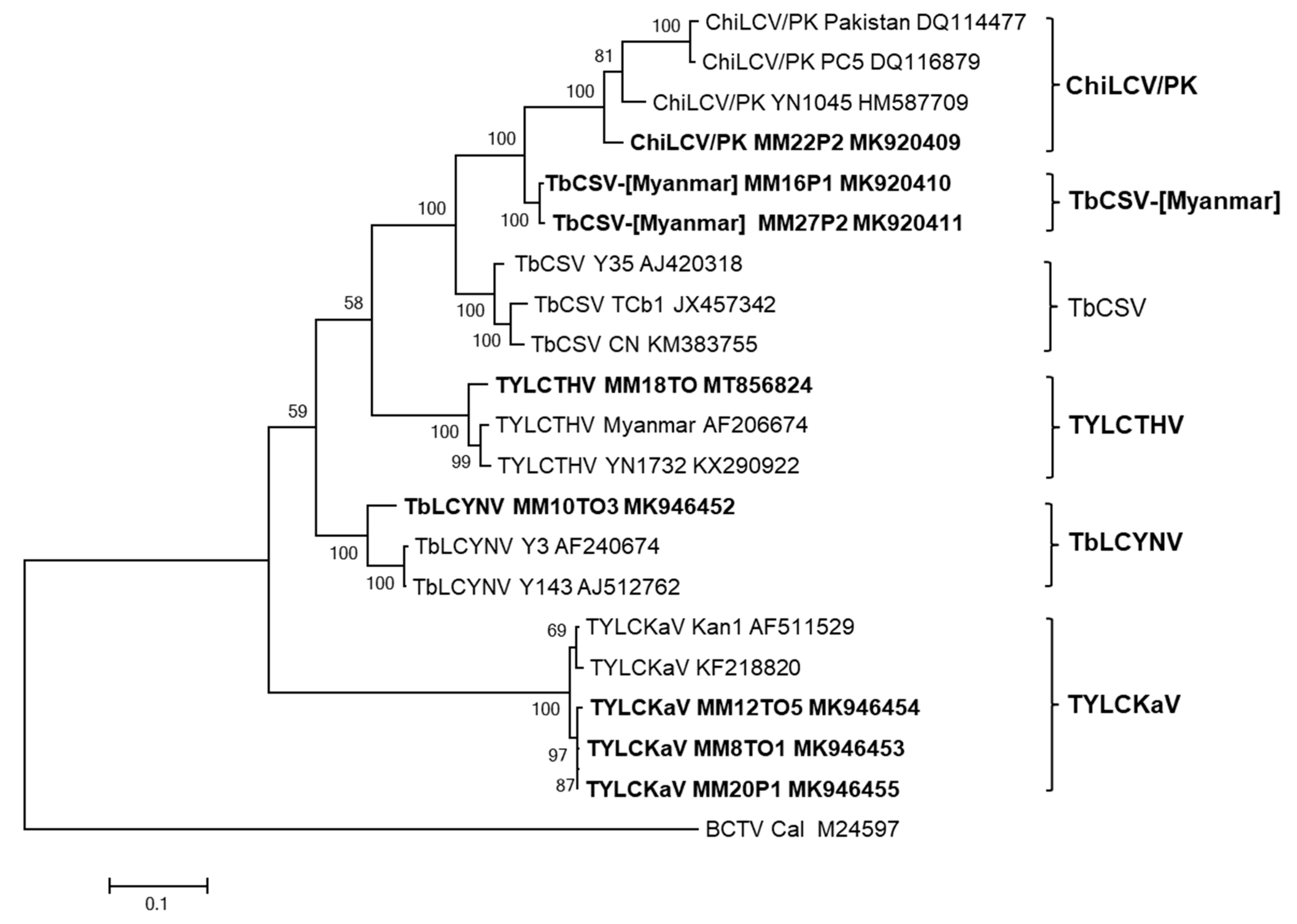
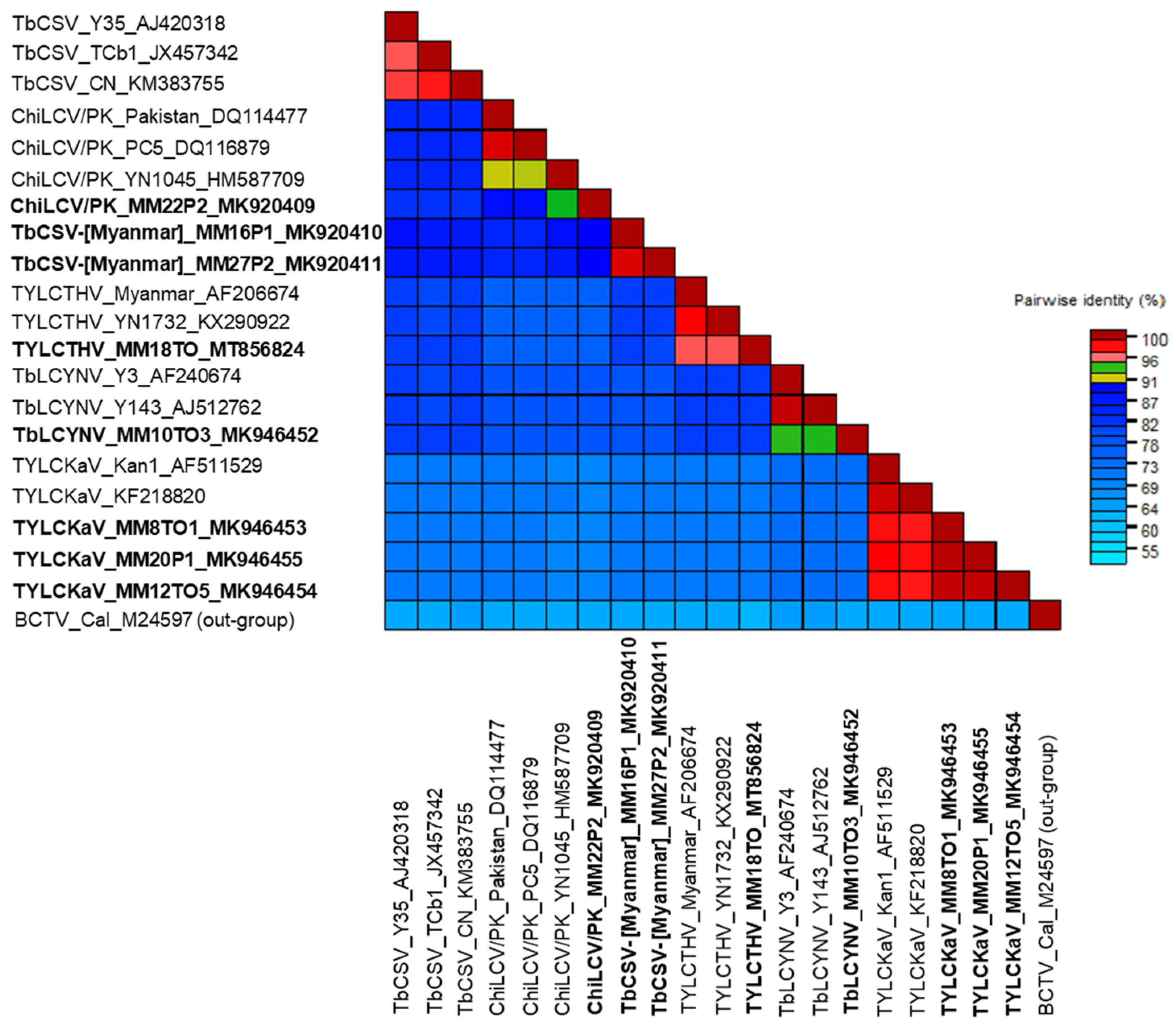
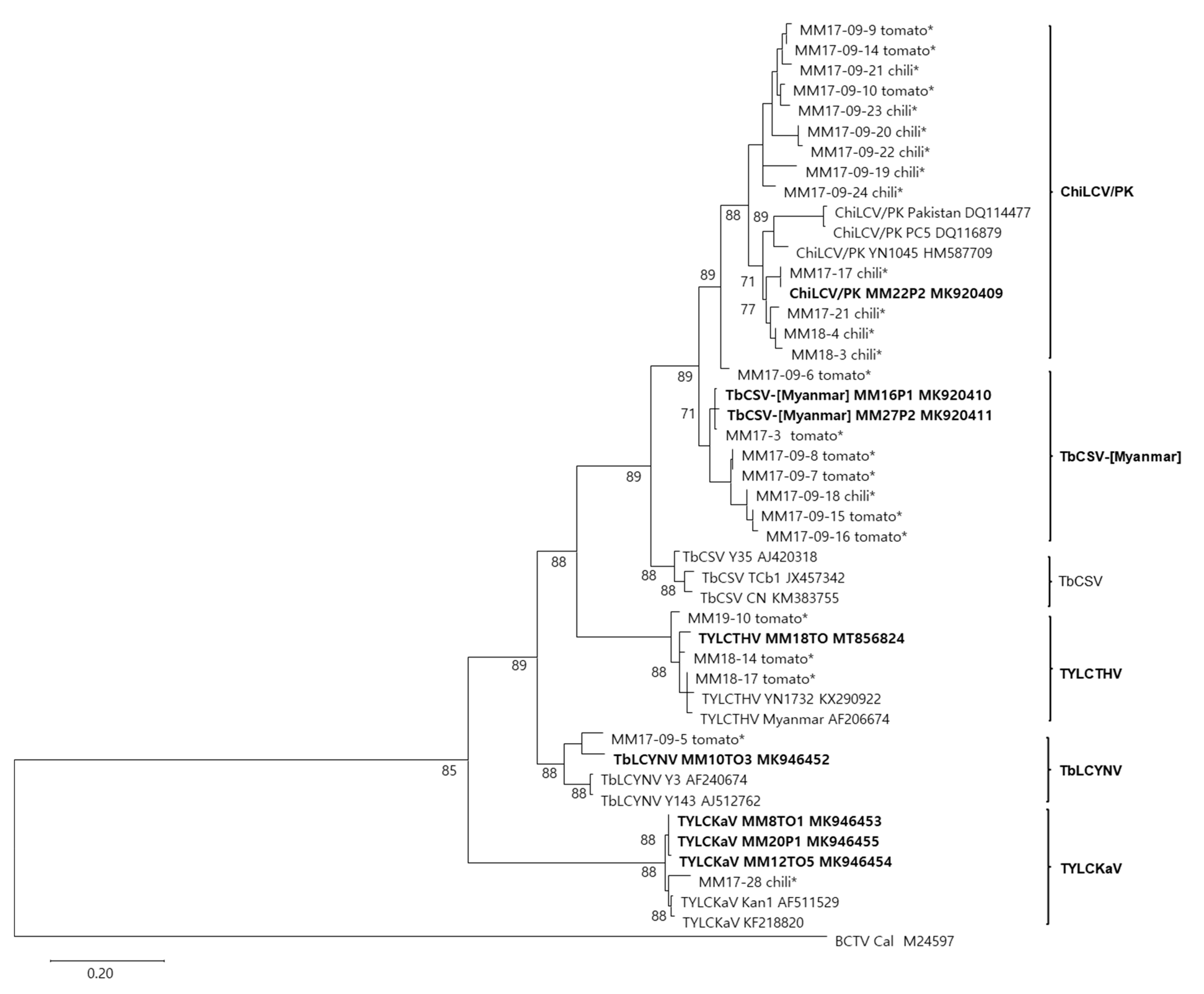
2.2. Pairwise Sequence Comparison and Phylogenetic Analysis of Begomovirus Species
2.3. Molecular Characterization of TbCSV-[Myanmar] Isolates
2.4. Diversity of Begomoviruses in Tomato and Pepper
3. Discussion
4. Materials and Methods
4.1. Sample Collection
4.2. Total Nucleic Acids Extraction
4.3. PCR, Cloning, and Sequencing
4.4. Sequence and Phylogenetic Analyses
4.5. Recombination Analysis of TbCSV-[Myanmar] Isolates
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Varma, A.; Malathi, V.G. Emerging geminivirus problems, a serious threat to crop production. Ann. Appl. Biol. 2003, 142, 145–164. [Google Scholar] [CrossRef]
- Kenyon, L.; Tsai, W.S.; Shih, S.L.; Lee, L.M. Emergence and diversity of begomoviruses infecting solanaceous crops in East and Southeast Asia. Virus Res. 2014, 186, 104–113. [Google Scholar] [CrossRef] [PubMed]
- Varma, A.; Mandal, B.; Singh, M.K. Emergence of begomoviruses, a major threat to vegetable production in Southeast Asia. In Proceedings of the SEAVEG 2012, Chiang Mai, Thailand, 24–26 January 2012. [Google Scholar]
- Navas-Castillo, J.; Fiallo-Olivé, E.; Sánchez-Campos, S. Emerging virus diseases transmitted by whiteflies. Annu. Rev. Phytopathol. 2011, 49, 219–248. [Google Scholar] [CrossRef] [PubMed]
- Gutierrez, C. Geminivirus DNA replication. Cell. Mol. Life Sci. CMLS 1999, 56, 313–329. [Google Scholar] [CrossRef] [PubMed]
- Hanley-Bowdoin, L.; Settlage, S.B.; Orozco, B.M.; Nagar, S.; Robertson, D. Geminiviruses, models for plant DNA replication, transcription, and cell cycle regulation. Crit. Rev. Plant Sci. 1999, 18, 71–106. [Google Scholar] [CrossRef]
- Stanley, J. Analysis of African cassava mosaic virus recombinants suggests strand nicking occurs within the conserved nonanucleotide motif during the initiation of rolling circle DNA replication. Virology 1995, 206, 707–712. [Google Scholar] [CrossRef]
- Stanley, J.; Bisaro, D.M.; Briddon, R.W.; Brown, J.K.; Fauquet, C.M.; Harrison, B.D.; Stenger, D.C. Family geminiviridae. In Virus Taxonomy, Eighth Report of the International Committee on Taxonomy of Viruses; Elsevier: Amsterdam, The Netherlands, 2005; pp. 301–326. [Google Scholar]
- Brown, J.K.; Zerbini, F.M.; Navas-Castillo, J.; Moriones, E.; Ramos-Sobrinho, R.; Silva, J.C. Revision of Begomovirus taxonomy based on pairwise sequence comparisons. Arch. Virol. 2015, 160, 1593–1619. [Google Scholar] [CrossRef]
- Annu, P.K.; Rani, R.; Rathi, A.S. Gemini Viruses-emerging threat to crops. J. Pharmacogn. Phytochem. 2019, 8, 2006–2012. [Google Scholar]
- Charoenvilaisiri, S.; Seepiban, C.; Phironrit, N.; Phuangrat, B.; Yoohat, K.; Deeto, R.; Gajanandana, O. Occurrence and distribution of begomoviruses infecting tomatoes, peppers and cucurbits in Thailand. Crop Protect. 2020, 127, 104948. [Google Scholar] [CrossRef]
- Kenyon, L.; Kumar, S.; Shih, S.L.; Lee, L.M.; Chan, Y.L. Diversity of begomoviruses causing disease in chilli peppers in Asia. In Proceedings of the International Congress of Plant Pathology (ICPP), Boston, MA, USA, 29 July–3 August 2018. [Google Scholar]
- Khatun, M.F.; Hwang, H.S.; Shim, J.K.; Kil, E.J.; Lee, S.; Lee, K.Y. Identification of begomoviruses from different cryptic species of Bemisia tabaci in Bangladesh. Microb. Pathog. 2020, 142, 104069. [Google Scholar] [CrossRef]
- Kumar, R.; Esakky, R.; Acharya, S. Molecular evidence of occurrence of Tomato Leaf Curl New Delhi Virus infecting cucurbits in several states in India. Arch. Phytopathol. Plant Prot. 2019, 52, 895–905. [Google Scholar] [CrossRef]
- Lukman, R.; Afifuddin, A.; Van Deynze, A.; Hill, T.; Jimenez, R. A survey of mixed Begomovirus infection in solanaceae and fabaceae at different altitudes in East Java, Indonesia. Arch. Phytopathol. Plant Prot. 2019, 52, 385–406. [Google Scholar] [CrossRef]
- Subiastuti, A.S.; Hartono, S.; Daryono, B.S. Detection and identification of Begomovirus infecting Cucurbitaceae and Solanaceae in Yogyakarta, Indonesia. Biodiversitas 2019, 20, 738–744. [Google Scholar] [CrossRef]
- Green, S.K.; Tsai, W.S.; Shih, S.L.; Black, L.L.; Rezaian, A.; Rashid, M.H.; Hong, L.T.A. Molecular characterization of begomoviruses associated with leafcurl diseases of tomato in Bangladesh, Laos, Malaysia, Myanmar, and Vietnam. Plant Dis. 2001, 85, 1286. [Google Scholar] [CrossRef] [PubMed]
- Brown, J.K.; Idris, A.M.; Torres-Jerez, I.; Banks, G.K.; Wyatt, S.D. The core region of the coat protein gene is highly useful for establishing the provisional identification and classification of begomoviruses. Arch. Virol. 2001, 146, 1581–1598. [Google Scholar] [CrossRef] [PubMed]
- Hernandez-Zepeda, C.; Idris, A.M.; Carnevali, G.; Brown, J.K.; Moreno-Valenzuela, O.A. Preliminary identification and coat protein gene phylogenetic relationships of begomoviruses associated with native flora and cultivated plants from the Yucatan Peninsula of Mexico. Virus Genes 2007, 25, 825–833. [Google Scholar] [CrossRef] [PubMed]
- Jan, F.J.; Green, S.K.; Shih, S.L.; Lee, L.M.; Ito, H.; Kimbara, J.; Tsai, W.S. First report of Tomato yellow leaf curl Thailand virus in Taiwan. Plant Dis. 2007, 91, 1363. [Google Scholar] [CrossRef] [PubMed]
- Green, S.K.; Tsai, W.S.; Shih, S.L.; Rezaian, M.A.; Duangsong, U. Molecular characterization of a new begomovirus associated with tomato yellow leaf curl and eggplant yellow mosaic diseases in Thailand. Plant Dis. 2003, 87, 446. [Google Scholar] [CrossRef]
- Bagewadi, B.; Naidu, R.A. First report of Tomato yellow leaf curl Kanchanaburi virus in eggplant and tomato in Cambodia. Plant Dis. 2016, 100, 233. [Google Scholar] [CrossRef]
- Ha, C.; Coombs, S.; Revill, P.; Harding, R.; Vu, M.; Dale, J. Molecular characterization of begomoviruses and DNA satellites from Vietnam, additional evidence that the New World geminiviruses were present in the Old World prior to continental separation. J. Gen. Virol. 2008, 89, 312–326. [Google Scholar] [CrossRef]
- Koeda, S.; Kesumawati, E.; Tanaka, Y.; Hosokawa, M.; Doi, M.; Kitajima, A. Mixed infection of begomoviruses on pepper plants at northern Sumatra, Indonesia. Trop. Agric. Dev. 2016, 60, 59–64. [Google Scholar]
- Tang, Y.F.; He, Z.F.; Du, Z.G.; Lu, L.H. First report of Tomato yellow leaf curl Kanchanaburi virus infecting eggplant in Laos. Plant Dis. 2014, 98, 428. [Google Scholar] [CrossRef] [PubMed]
- Zhou, X.P.; Xie, Y.; Zhang, Z.K. Molecular characterization of a distinct begomovirus infecting tobacco in Yunnan, China. Arch. Virol. 2001, 146, 1599–1606. [Google Scholar] [CrossRef] [PubMed]
- Thakur, H.; Jindal, S.K.; Sharma, A.; Dhaliwal, M.S. Chilli leaf curl virus disease, a serious threat for chilli cultivation. J. Plant Dis. Prot. 2018, 125, 239–249. [Google Scholar] [CrossRef]
- Khan, A.J.; Akhtar, S.; Al-Zaidi, A.M.; Singh, A.K.; Briddon, R.W. Genetic diversity and distribution of a distinct strain of Chili leaf curl virus and associated betasatellite infecting tomato and pepper in Oman. Virus Res. 2013, 177, 87–97. [Google Scholar] [CrossRef]
- Mubin, M.; Briddon, R.W.; Mansoor, S. Complete nucleotide sequence of chili leaf curl virus and its associated satellites naturally infecting potato in Pakistan. Arch. Virol. 2009, 154, 365–368. [Google Scholar] [CrossRef] [PubMed]
- Senanayake, D.M.J.B.; Mandal, B.; Lodha, S.; Varma, A. First report of Chilli leaf curl virus affecting chilli in India. Plant Pathol. 2007, 56, 343. [Google Scholar] [CrossRef]
- Shahid, M.S.; Al-Sadi, A.M.; Briddon, R.W. First report of Chilli leaf curl virus and tomato leaf curl betasatellite infecting watermelon (Citrullus lanatus) in Oman. Plant Dis. 2017, 101, 1063. [Google Scholar] [CrossRef]
- Shih, S.L.; Tsai, W.S.; Green, S.K.; Khalid, S.; Ahmad, I.; Rezaian, M.A.; Smith, J. Molecular characterization of tomato and chili leaf curl begomoviruses from Pakistan. Plant Dis. 2003, 87, 200. [Google Scholar] [CrossRef]
- Li, P.B.; Jing, C.C.; Wang, Z.Y.; Ma, M.G.; Li, M.J.; Wu, G.T.; Qing, L. First Report of Tobacco curly shoot virus Infecting Phaseolus vulgaris in China. Plant Dis. 2019, 103, 165. [Google Scholar] [CrossRef]
- Qing, L.; Xiong, Y.; Sun, X.C.; Yang, S.Y.; Zhou, C.Y. First report of Tobacco curly shoot virus infecting pepper in China. Plant Dis. 2010, 94, 637. [Google Scholar] [CrossRef] [PubMed]
- Xie, Y.; Zhou, X.P.; Zhang, Z.K.; Qi, Y.J. Tobacco curly shoot virus isolated in Yunnan is a distinct species of Begomovirus. Chin. Sci. Bull. 2002, 47, 197–200. [Google Scholar] [CrossRef]
- Zhao, L.L.; Ding, M.; Zhang, X.Y.; Yin, Y.Y.; Li, T.T.; Zhang, Z.K. First report of tobacco curly shoot virus (TBCSV) and its associated satellites from watermelon in China. J. Plant Pathol. 2017, 99, 761–764. [Google Scholar]
- Li, Z.; Xie, Y.; Zhou, X. Tobacco curly shoot virus DNAβ is not necessary for infection but intensifies symptoms in a host-dependent manner. Phytopathology 2005, 95, 902–908. [Google Scholar] [CrossRef] [PubMed]
- Kumar, R.; Palicherla, S.R.; Mandal, B.; Kadiri, S. PCR based detection of betasatellite associated with the begomoviruses using improved universal primers. Australas. Plant Pathol. 2018, 47, 115–118. [Google Scholar] [CrossRef]
- Hong, S.B.; Kwak, H.R.; Kim, M.K.; Seo, J.K.; Shin, J.S.; Han, J.H.; Choi, H.S. Simple and Rapid Detection for Rice stripe virus Using RT-PCR and Porous Ceramic Cubes. Res. Plant Dis. 2015, 21, 321–325. [Google Scholar] [CrossRef][Green Version]
- Lee, G.; Kim, S.; Jung, J.; Auh, C.K.; Choi, E.; Chang, M.; Lee, S. Agroinoculation of Nicotiana benthamiana with cloned honeysuckle yellow vein virus isolated from Lonicera japonica. Arch. Virol. 2011, 156, 785–791. [Google Scholar] [CrossRef]
- Thompson, J.D.; Higgins, D.G.; Gibson, T.J. CLUSTAL W: Improving the sensitivity of progressive multiple sequence alignment through sequence weighting, position-specific gap penalties and weight matrix choice. Nucleic Acids Res. 1994, 22, 4673–4680. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular evolutionary genetics analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Muhire, B.M.; Varsani, A.; Martin, D.P. SDT, a virus classification tool based on pairwise sequence alignment and identity calculation. PLoS ONE 2014, 9, e108277. [Google Scholar] [CrossRef]
- Martin, D.P.; Murrell, B.; Golden, M.; Khoosal, A.; Muhire, B. RDP4, Detection and analysis of recombination patterns in virus genomes. Virus Evol. 2015, 1, vev003. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Surveyed Vegetable Crops | Collected Year | Collected Area | No. Of Collected Samples | No. of Plant Positive for Begomovirus By PCR | No. of Begomovirus Species Identified Based on Blast Analysis of Amplified Product a | |||
|---|---|---|---|---|---|---|---|---|
| ChiLCV/PK | TYLCTHV | TYLCKaV | TbLCYnV | |||||
| Tomato | 2017 | Naypyitaw | 9 | 7 b | 1 (1) c | 1 | 2 | 1 |
| 2017 | Naypyitaw | 15 | 9 | 8 (5) c | 1 | |||
| 2018 | Naypyitaw | 3 | 1 | 1 | ||||
| 2018 | Tatkon | 4 | 1 | 1 | ||||
| 2018 | Mohnyin | 2 | 0 | |||||
| 2019 | Naypyitaw | 3 | 1 | 1 | ||||
| Subtotal | 36 | 19 (52.8%) | 9 (6) c | 4 | 2 | 2 | ||
| Pepper | 2017 | Naypyitaw | 5 | 5 | 4 (1) c | 1 | ||
| 2017 | Tatkon | 2 | 2 | 1 (1) c | 1 | |||
| 2017 | Naypyitaw | 9 | 7 | 7 (1) c | ||||
| 2018 | Naypyitaw | 3 | 0 | |||||
| 2018 | Tatkon | 4 | 2 | 2 | ||||
| Subtotal | 23 | 16 (69.6%) | 14 (3) c | 2 | ||||
| Total | 59 | 35 (59.3%) | 23 (9) c | 4 | 4 | 2 | ||
| Identities of the closely reported begomovirus in Genbank | 94–96%, HM587709 | 96–99%, KX290922 | 98%, AF511529 | 93%, AF240674 | ||||
| Virus a | Isolate b | Host Plant | Origin | Genome (nt) | NCBI Accession No. |
|---|---|---|---|---|---|
| ChiLCV/PK | MM22P2 | Pepper | Myanmar: Naypyitaw | 2759 | MK920409 |
| YN1045 | Pepper | China | 2752 | HM587709 | |
| Pakistan | Pepper | Pakistan | 2756 | DQ114477 | |
| PC5 | Pepper | Pakistan | 2756 | DQ116879 | |
| TYLCTHV | MM18TO | Tomato | Myanmar: Naypyitaw | 2747 | MT856824 |
| YN1732 | Tomato | China | 2744 | KX290922 | |
| D[MY:Yan:99] | Tomato | Myanmar | 2746 | AF206674 | |
| TYLCKaV | MM8TO1 | Tomato | Myanmar: Naypyitaw | 2753 | MK946453 |
| MM12TO5 | Tomato | Myanmar: Naypyitaw | 2753 | MK946454 | |
| MM20P1 | Pepper | Myanmar: Naypyitaw | 2753 | MK946455 | |
| TH:Kan1 | Tomato | Thailand | 2752 | AF511529 | |
| Laos | Eggplant | Laos | 2752 | KF218820 | |
| TbLCYnV | MM10TO3 | Tomato | Myanmar: Naypyitaw | 2753 | MK946452 |
| Y3 | Tobacco | China | 2744 | AF240674 | |
| Y143 | Tobacco | China | 2750 | AJ512762 | |
| TbCSV | Y35 | Tobacco | China | 2746 | AJ420318 |
| CN[BD:Raj:02:25:Tom:10] | Tomato | Bangladesh | 2746 | KM383755 | |
| TCb1 | Tomato | India | 2758 | JX457342 | |
| TbCSV-[Myanmar] | MM16P1 | Pepper | Myanmar: Naypyitaw | 2762 | MK920410 |
| MM27P2 | Pepper | Myanmar: Tetkone | 2762 | MK920411 |
| Virus Isolate | NCBI Accession No. | Full-Genome Nucleotide Sequence Identities (%) | Amino Acid Sequence Identities (%) | |||||
|---|---|---|---|---|---|---|---|---|
| V1 (CP) | V2 (MP) | C1 (Rep) | C2 (TrAP) | C3 (Ren) | C4 | |||
| TbCSV-[Myanmar]-MM27P2 | MK920411 | 99 | 99 | 100 | 98 | 97 | 96 | 96 |
| TbCSV-Y35 | AJ420318 | 86 | 97 | 94 | 88 | 84 | 83 | 90 |
| TbCSV-TCb1 | JX457342 | 85 | 96 | 93 | 87 | 86 | 84 | 89 |
| TbCSV-CN[BD:Raj:02:25:Tom:10] | KM383755 | 85 | 96 | 92 | 87 | 86 | 83 | 90 |
| ChiLCV/PK-MM22P2 | MK920409 | 80 | 98 | 96 | 80 | 97 | 93 | 32 |
| ChiLCV/PK-YN1045 | HM587709 | 82 | 98 | 94 | 78 | 92 | 90 | 35 |
| ChiLCV/PK-Pakistan | DQ114477 | 79 | 97 | 93 | 77 | 91 | 91 | 32 |
| ChiLCV/PK-PC5 | DQ116879 | 80 | 98 | 94 | 77 | 90 | 91 | 33 |
| Recombination Breakpoints | Parental Isolates a | RDP4 (p-Value) b | |
|---|---|---|---|
| Major | Minor | ||
| 16-2106 | TbCSV-CN[BD:Raj:02:25:Tom:10] | ChiLCV/PK-YN1045 | R, G, B, M, C, S, 3S (4.895 × 10−39) |
| (KM383755) | (HM587709) | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kwak, H.-R.; Hong, S.-B.; Byun, H.-S.; Park, B.; Choi, H.-S.; Myint, S.S.; Kyaw, M.M. Incidence and Molecular Identification of Begomoviruses Infecting Tomato and Pepper in Myanmar. Plants 2022, 11, 1031. https://doi.org/10.3390/plants11081031
Kwak H-R, Hong S-B, Byun H-S, Park B, Choi H-S, Myint SS, Kyaw MM. Incidence and Molecular Identification of Begomoviruses Infecting Tomato and Pepper in Myanmar. Plants. 2022; 11(8):1031. https://doi.org/10.3390/plants11081031
Chicago/Turabian StyleKwak, Hae-Ryun, Su-Bin Hong, Hee-Seong Byun, Bueyong Park, Hong-Soo Choi, Si Si Myint, and Mu Mu Kyaw. 2022. "Incidence and Molecular Identification of Begomoviruses Infecting Tomato and Pepper in Myanmar" Plants 11, no. 8: 1031. https://doi.org/10.3390/plants11081031
APA StyleKwak, H.-R., Hong, S.-B., Byun, H.-S., Park, B., Choi, H.-S., Myint, S. S., & Kyaw, M. M. (2022). Incidence and Molecular Identification of Begomoviruses Infecting Tomato and Pepper in Myanmar. Plants, 11(8), 1031. https://doi.org/10.3390/plants11081031