
Entanglement of Arabidopsis Seedlings to a Mesh Substrate under Microgravity Conditions in KIBO on the ISS


 ,
, 
Abstract
:1. Introduction
2. Results
3. Discussion
4. Materials and Methods
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kolesnikov, Y.S.; Kretynin, S.V.; Volotovsky, I.D.; Kordyum, E.L.; Ruelland, E.; Kravets, V.S. Erratum to: Molecular mechanisms of gravity perception and signal transduction in plants. Protoplasma 2016, 253, 1005. [Google Scholar] [CrossRef] [Green Version]
- Joo, J.H.; Bae, Y.S.; Lee, J.S. Role of auxin-induced reactive oxygen species in root gravitropism. Plant Physiol. 2001, 126, 1055–1060. [Google Scholar] [CrossRef] [Green Version]
- Fasano, J.M.; Swanson, S.J.; Blancaflor, E.B.; Dowd, P.E.; Kao, T.H.; Gilroy, S. Changes in root cap pH are required for the gravity response of the Arabidopsis root. Plant Cell 2001, 13, 907–921. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scott, A.C.; Allen, N.S. Changes in cytosolic pH within Arabidopsis root columella cells play a key role in the early signaling pathway for root gravitropism. Plant Physiol. 1999, 121, 1291–1298. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fasano, J.M.; Massa, G.D.; Gilroy, S. Ionic signaling in plant responses to gravity and touch. J. Plant Growth Regul. 2002, 21, 71–88. [Google Scholar] [CrossRef] [PubMed]
- Perera, I.Y.; Heilmann, I.; Chang, S.C.; Boss, W.F.; Kaufman, P.B. A role for inositol 1,4,5-trisphosphate in gravitropic signaling and the retention of cold-perceived gravistimulation of oat shoot pulvini. Plant Physiol. 2001, 125, 1499–1507. [Google Scholar] [CrossRef] [Green Version]
- Sinclair, W.; Trewavas, A.J. Calcium in gravitropism. A re-examination. Planta 1997, 203, S85–S90. [Google Scholar] [CrossRef] [PubMed]
- Toyota, M.; Gilroy, S. Gravitropism and mechanical signaling in plants. Am. J. Bot. 2013, 100, 111–125. [Google Scholar] [CrossRef] [PubMed]
- Chen, R.; Rosen, E.; Masson, P.H. Gravitropism in higher plants. Plant Physiol. 1999, 120, 343–350. [Google Scholar] [CrossRef] [Green Version]
- Morita, M.T.; Tasaka, M. Gravity sensing and signaling. Curr. Opin. Plant Biol. 2004, 7, 712–718. [Google Scholar] [CrossRef]
- Nakano, M.; Furuichi, T.; Sokabe, M.; Iida, H.; Tatsumi, H. The gravistimulation-induced very slow Ca2+ increase in Arabidopsis seedlings requires MCA1, a Ca2+-permeable mechanosensitive channel. Sci. Rep. 2021, 11, 227. [Google Scholar] [CrossRef] [PubMed]
- Toyota, M.; Furuichi, T.; Tatsumi, H.; Sokabe, M. Cytoplasmic calcium increases in response to changes in the gravity vector in hypocotyls and petioles of Arabidopsis seedlings. Plant Physiol. 2008, 146, 505–514. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Toyota, M.; Furuichi, T.; Sokabe, M.; Tatsumi, H. Analyses of a gravistimulation-specific Ca2+ signature in Arabidopsis using parabolic flights. Plant Physiol. 2013, 163, 543–554. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakagawa, Y.; Katagiri, T.; Shinozaki, K.; Qi, Z.; Tatsumi, H.; Furuichi, T.; Kishigami, A.; Sokabe, M.; Kojima, I.; Sato, S.; et al. Arabidopsis plasma membrane protein crucial for Ca2+ influx and touch sensing in roots. Proc. Natl. Acad. Sci. USA 2007, 104, 3639–3644. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamanaka, T.; Nakagawa, Y.; Mori, K.; Nakano, M.; Imamura, T.; Kataoka, H.; Terashima, A.; Iida, K.; Kojima, I.; Katagiri, T.; et al. MCA1 and MCA2 that mediate Ca2+ uptake have distinct and overlapping roles in Arabidopsis. Plant Physiol. 2010, 152, 1284–1296. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haswell, E.S.; Meyerowitz, E.M. MscS-like proteins control plastid size and shape in Arabidopsis thaliana. Curr. Biol. 2006, 16, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Haswell, E.S.; Peyronnet, R.; Barbier-Brygoo, H.; Meyerowitz, E.M.; Frachisse, J.M. Two MscS homologs provide mechanosensitive channel activities in the Arabidopsis root. Curr. Biol. 2008, 18, 730–734. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maathuis, F.J.M. Vacuolar two-pore K+ channels act as vacuolar osmosensors. New Phytol. 2011, 191, 84–91. [Google Scholar] [CrossRef] [PubMed]
- Coste, B.; Mathur, J.; Schmidt, M.; Earley, T.J.; Ranade, S.; Petrus, M.J.; Dubin, A.E.; Patapoutian, A. Piezo1 and Piezo2 are essential components of distinct mechanically activated cation channels. Science 2010, 330, 55–60. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yuan, F.; Yang, H.; Xue, Y.; Kong, D.; Ye, R.; Li, C.; Zhang, J.; Theprungsirikul, L.; Shrift, T.; Krichilsky, B.; et al. OSCA1 mediates osmotic-stress-evoked Ca2+ increases vital for osmosensing in Arabidopsis. Nature 2014, 514, 367–371. [Google Scholar] [CrossRef]
- Becker, D.; Geiger, D.; Dunkel, M.; Roller, A.; Bertl, A.; Latz, A.; Carpaneto, A.; Dietrich, P.; Roelfsema, M.R.; Voelker, C.; et al. AtTPK4, an Arabidopsis tandem-pore K+ channel, poised to control the pollen membrane voltage in a pH- and Ca2+-dependent manner. Proc. Natl. Acad. Sci. USA 2004, 101, 15621–15626. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Furuichi, T.; Iida, H.; Sokabe, M.; Tatsumi, H. Expression of Arabidopsis MCA1 enhanced mechanosensitive channel activity in the Xenopus laevis oocyte plasma membrane. Plant Signal. Behav. 2012, 7, 1022–1026. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shigematsu, H.; Iida, K.; Nakano, M.; Chaudhuri, P.; Iida, H.; Nagayama, K. Structural characterization of the mechanosensitive channel candidate MCA2 from Arabidopsis thaliana. PLoS ONE 2014, 9, e87724. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Veley, K.M.; Marshburn, S.; Clure, C.E.; Haswell, E.S. Mechanosensitive channels protect plastids from hypoosmotic stress during normal plant growth. Curr. Biol. 2012, 22, 408–413. [Google Scholar] [CrossRef] [Green Version]
- Yoshimura, K.; Iida, K.; Iida, H. MCAs in Arabidopsis are Ca2+-permeable mechanosensitive channels inherently sensitive to membrane tension. Nat. Commun. 2021, 12, 6074. [Google Scholar] [CrossRef]
- Hattori, T.; Otomi, Y.; Nakajima, Y.; Soga, K.; Wakabayashi, K.; Iida, H.; Hoson, T. MCA1 and MCA2 Are Involved in the Response to Hypergravity in Arabidopsis Hypocotyls. Plants 2020, 9, 590. [Google Scholar] [CrossRef]
- Iida, H.; Furuichi, T.; Nakano, M.; Toyota, M.; Sokabe, M.; Tatsumi, H. New candidates for mechano-sensitive channels potentially involved in gravity sensing in Arabidopsis thaliana. Plant Biol. 2014, 16 (Suppl. 1), 39–42. [Google Scholar] [CrossRef] [PubMed]
- Ruyters, G.; Braun, M. Plant biology in space: Recent accomplishments and recommendations for future research. Plant Biol. 2014, 16 (Suppl 1), 4–11. [Google Scholar] [CrossRef]
- Scherer, G.F.; Pietrzyk, P. Gravity-dependent differentiation and root coils in Arabidopsis thaliana wild type and phospholipase-A-I knockdown mutant grown on the International Space Station. Plant Biol. 2014, 16 (Suppl 1), 97–106. [Google Scholar] [CrossRef] [PubMed]
- Migliaccio, F.; Tassone, P.; Fortunati, A. Circumnutation as an autonomous root movement in plants. Am. J. Bot. 2013, 100, 4–13. [Google Scholar] [CrossRef] [Green Version]
- Millar, K.D.; Johnson, C.M.; Edelmann, R.E.; Kiss, J.Z. An endogenous growth pattern of roots is revealed in seedlings grown in microgravity. Astrobiology 2011, 11, 787–797. [Google Scholar] [CrossRef]
- Massa, G.D.; Gilroy, S. Touch modulates gravity sensing to regulate the growth of primary roots of Arabidopsis thaliana. Plant J. 2003, 33, 435–445. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Okada, K.; Shimura, Y. Reversible root tip rotation in Arabidopsis seedlings induced by obstacle-touching stimulus. Science 1990, 250, 274–276. [Google Scholar] [CrossRef] [PubMed]
- Yano, S.; Masuda, D.; Kasahara, H.; Omori, K.; Higashibata, A.; Asashima, M.; Ohnishi, T.; Yatagai, F.; Kamisaka, S.; Furusawa, T.; et al. Excellent Thermal Control Ability of Cell Biology Experiment Facility (CBEF) for Ground-Based Experiments and Experiments Onboard the Kibo Japanese Experiment Module of International Space Station. Biol. Sci. Space 2012, 26, 12–20. [Google Scholar] [CrossRef] [Green Version]
- Soga, K.; Yano, S.; Matsumoto, S.; Hoson, T. Hypergravity experiments to evaluate gravity resistance mechanisms in plants. Methods Mol. Biol. 2015, 1309, 307–319. [Google Scholar] [CrossRef]
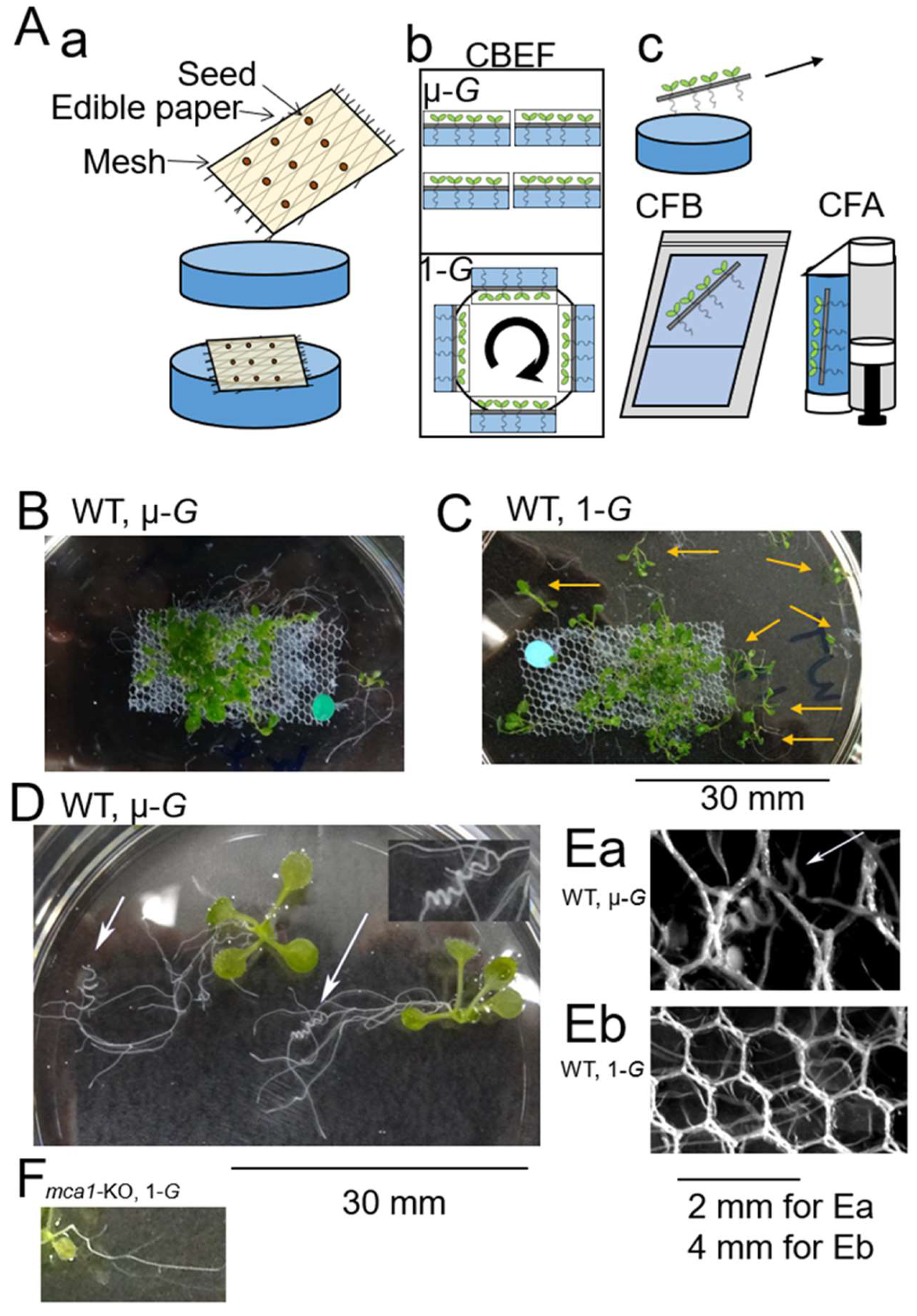

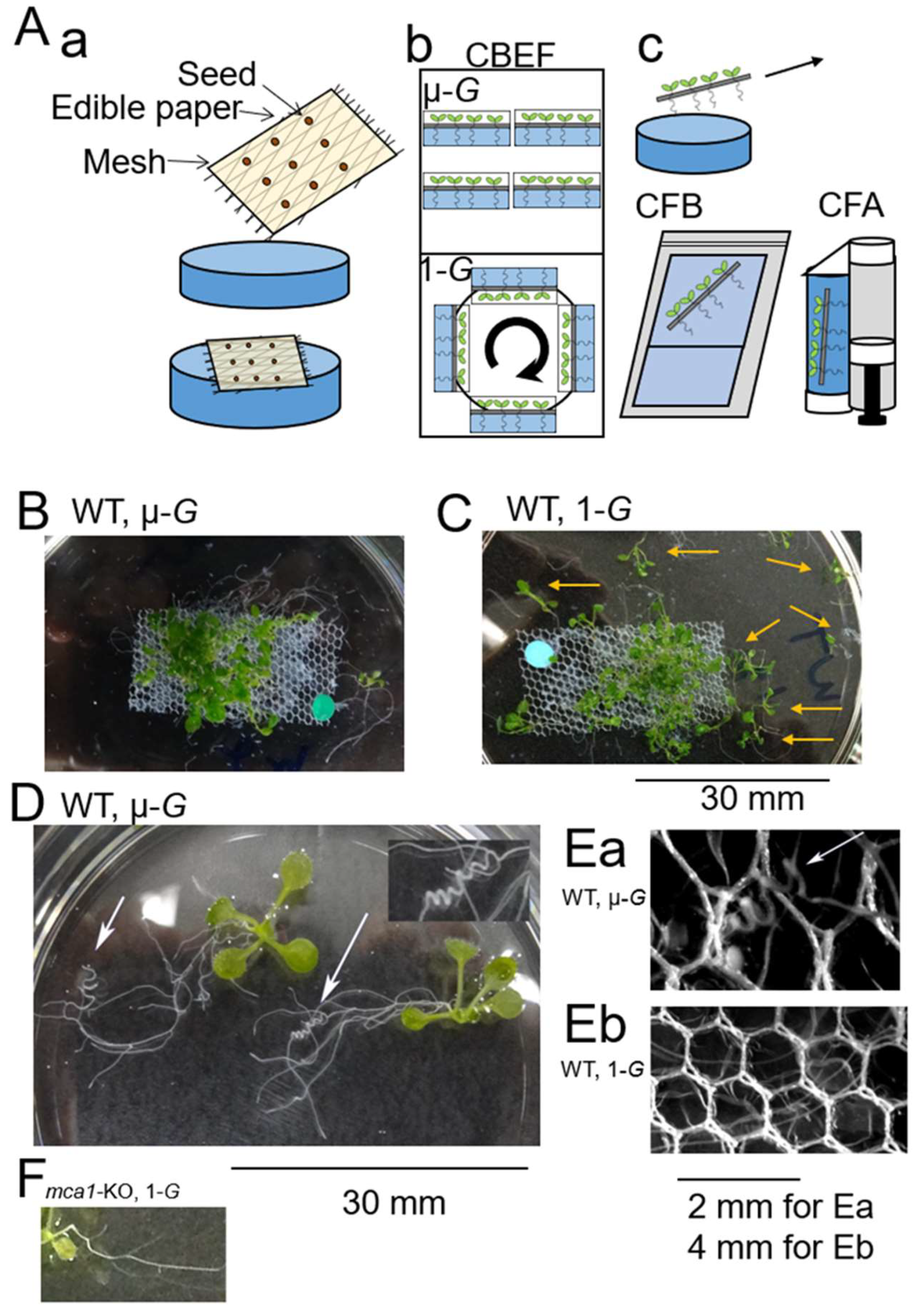{kind=link}
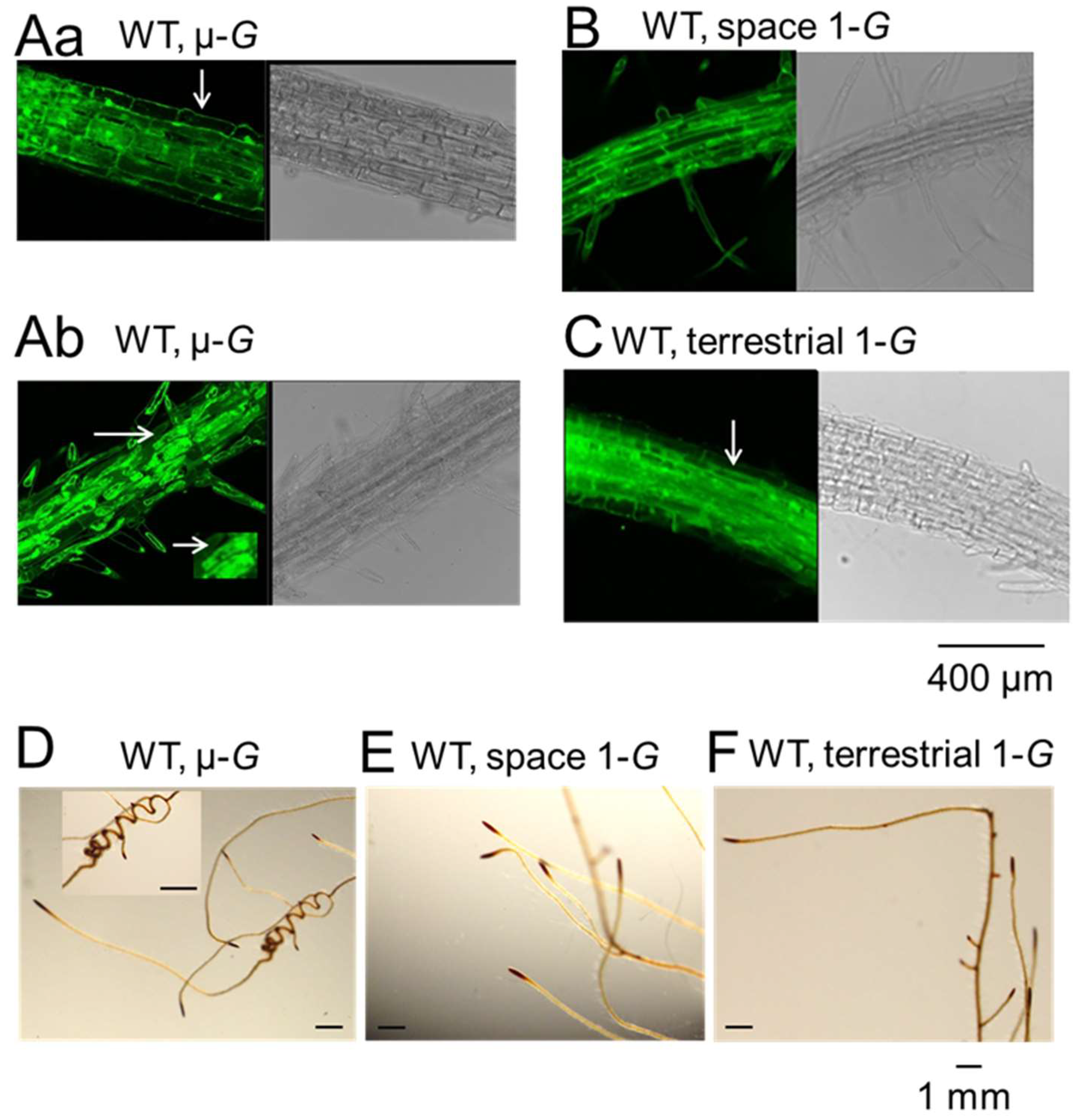
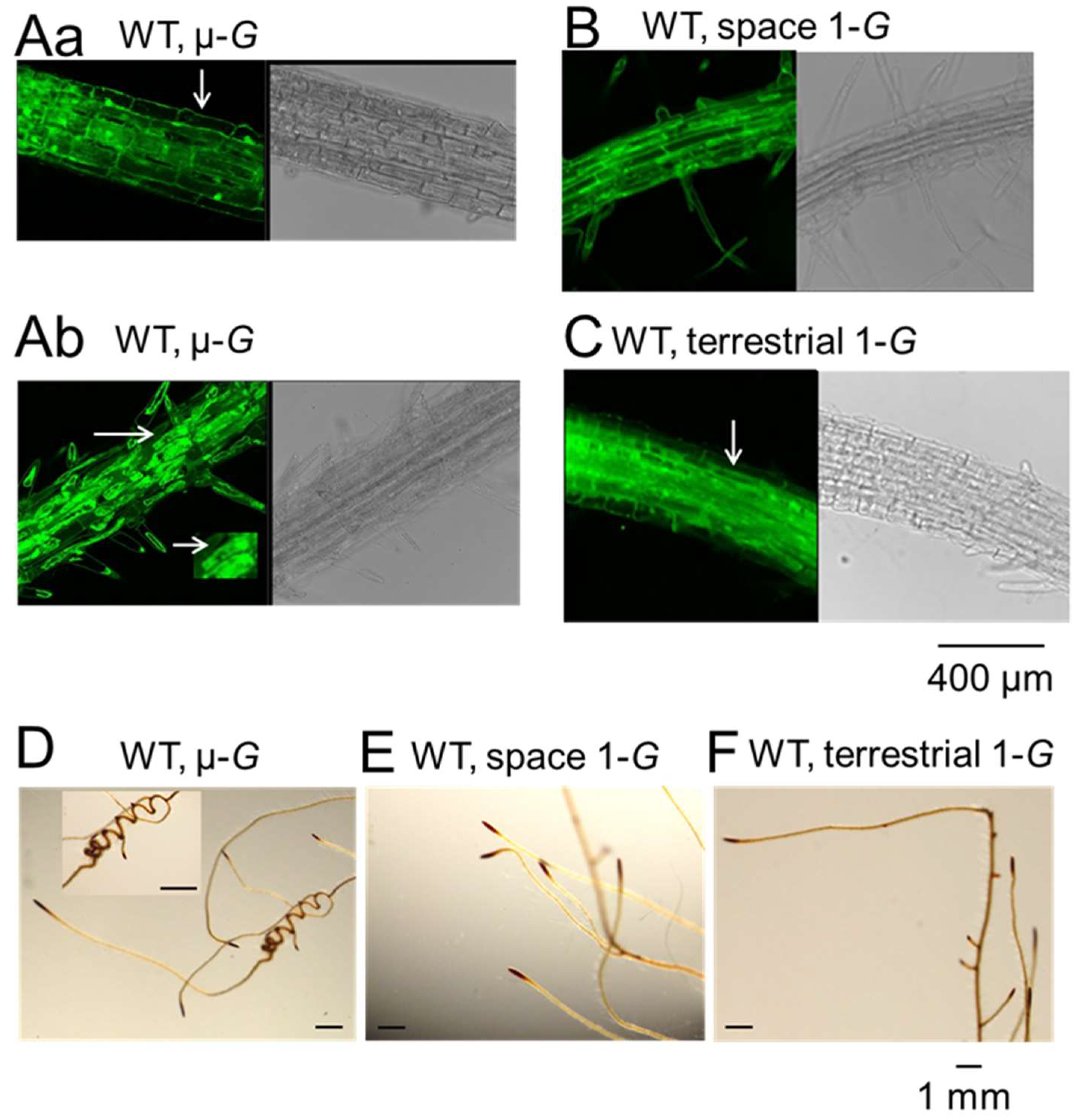{kind=link}
| Space Micro-G | Space Artificial 1-G | |||
|---|---|---|---|---|
| Seedlings on Mesh | Seedlings Dispersed | Seedlings on Mesh | Seedlings Dispersed | |
| WT | 20 | 3 (13%) | 20 | 11 (35%) “c” |
| mca1-KO | 15 | 2 (11.7%) “a” | 3 | 17 (85%) “b” |
| MCA1-OX | 24 | 1 (4%) | 19 | 6 (24%) “d” |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nakano, M.; Furuichi, T.; Sokabe, M.; Iida, H.; Yano, S.; Tatsumi, H. Entanglement of Arabidopsis Seedlings to a Mesh Substrate under Microgravity Conditions in KIBO on the ISS. Plants 2022, 11, 956. https://doi.org/10.3390/plants11070956
Nakano M, Furuichi T, Sokabe M, Iida H, Yano S, Tatsumi H. Entanglement of Arabidopsis Seedlings to a Mesh Substrate under Microgravity Conditions in KIBO on the ISS. Plants. 2022; 11(7):956. https://doi.org/10.3390/plants11070956
Chicago/Turabian StyleNakano, Masataka, Takuya Furuichi, Masahiro Sokabe, Hidetoshi Iida, Sachiko Yano, and Hitoshi Tatsumi. 2022. "Entanglement of Arabidopsis Seedlings to a Mesh Substrate under Microgravity Conditions in KIBO on the ISS" Plants 11, no. 7: 956. https://doi.org/10.3390/plants11070956
APA StyleNakano, M., Furuichi, T., Sokabe, M., Iida, H., Yano, S., & Tatsumi, H. (2022). Entanglement of Arabidopsis Seedlings to a Mesh Substrate under Microgravity Conditions in KIBO on the ISS. Plants, 11(7), 956. https://doi.org/10.3390/plants11070956