
The Assembly of Tropical Dry Forest Tree Communities in Anthropogenic Landscapes: The Role of Chemical Defenses

 ,
,  ,
,  ,
,  ,
,  ,
,  and
and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Site
2.2. Vegetation Sampling and Plant Functional Traits Measurements
Quantification of Total Phenols, Tannins, and Flavonoids
2.3. Environmental Characterization
2.3.1. Environmental and Biogeochemical Soil Factors Analyses
2.3.2. Environmental Data Collection
2.4. Data Analysis
2.4.1. Relationships between Leaf Defense Traits and Plant Functional Traits
2.4.2. Contrasting OGF and SEF Plant Communities in Terms of Their Functional Composition
2.4.3. Evaluation of Potential Assembly Rules Underlying Species Coexistence in OGF and SEF Sites
2.4.4. Identification of Potential External Environmental Filters Modulating Plant Community Assembly
3. Results
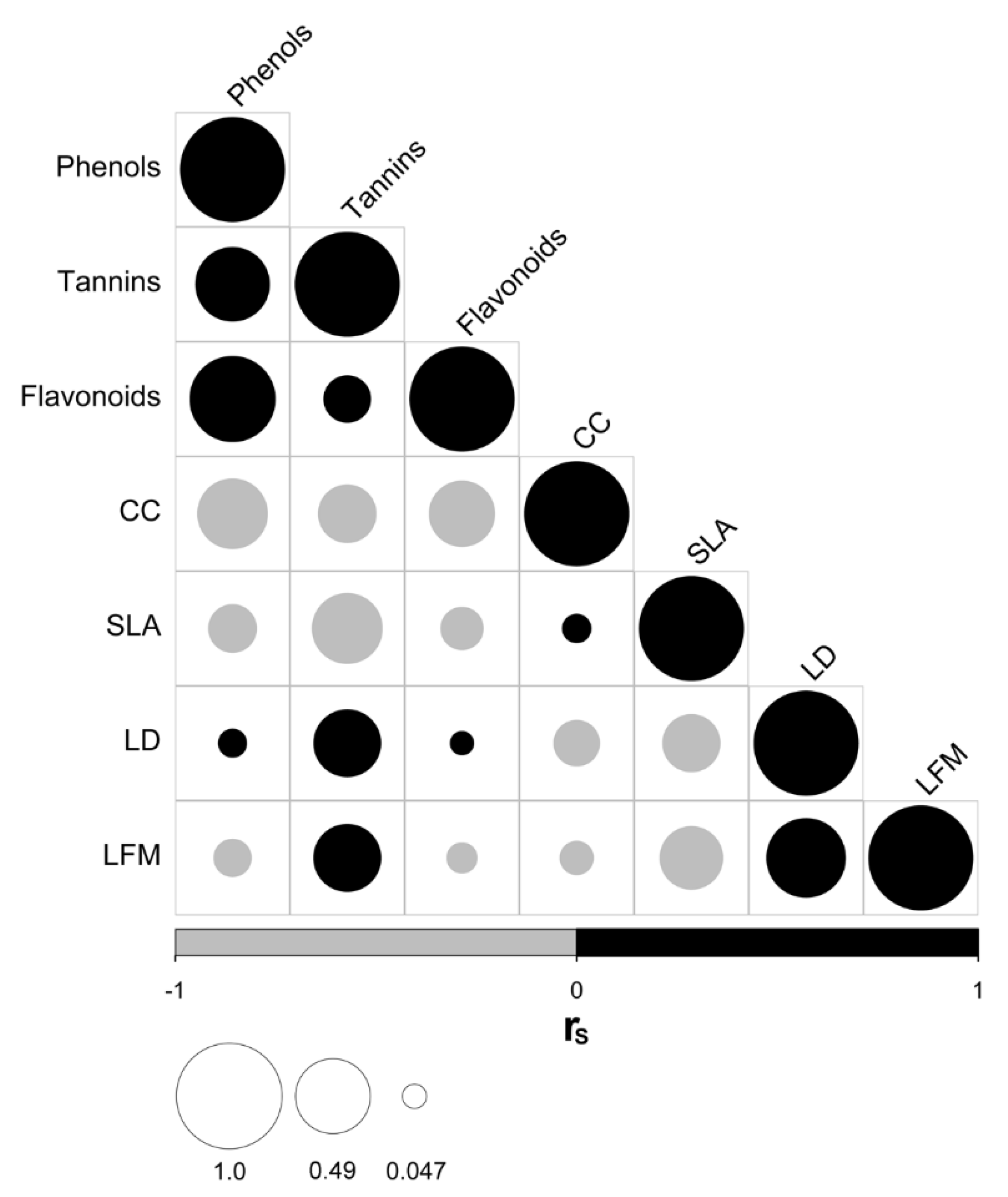
3.1. Relationships between Traits Related to Leaf Defense, Water Use, Light Acquisition, and Heat Load Regulation
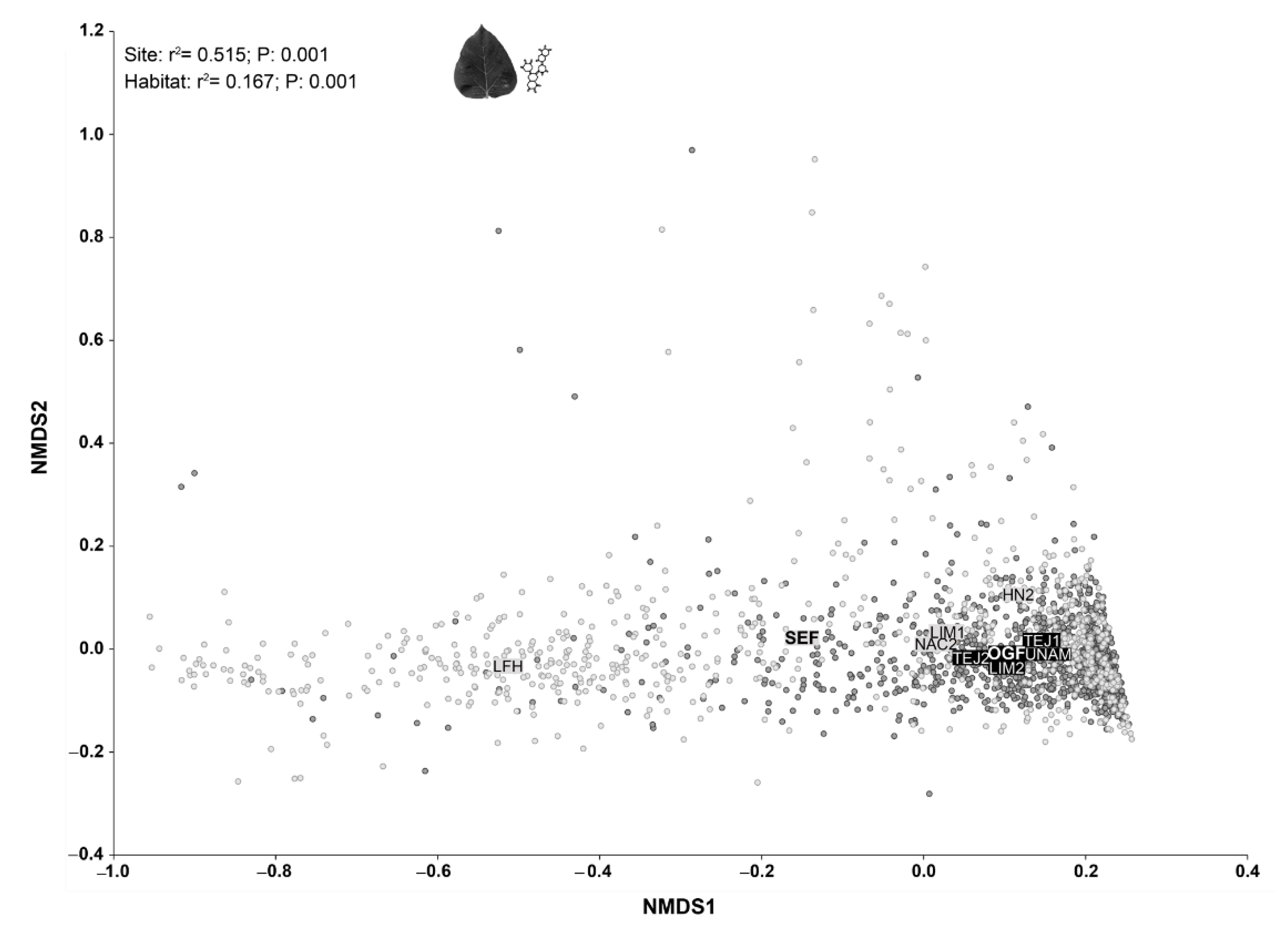
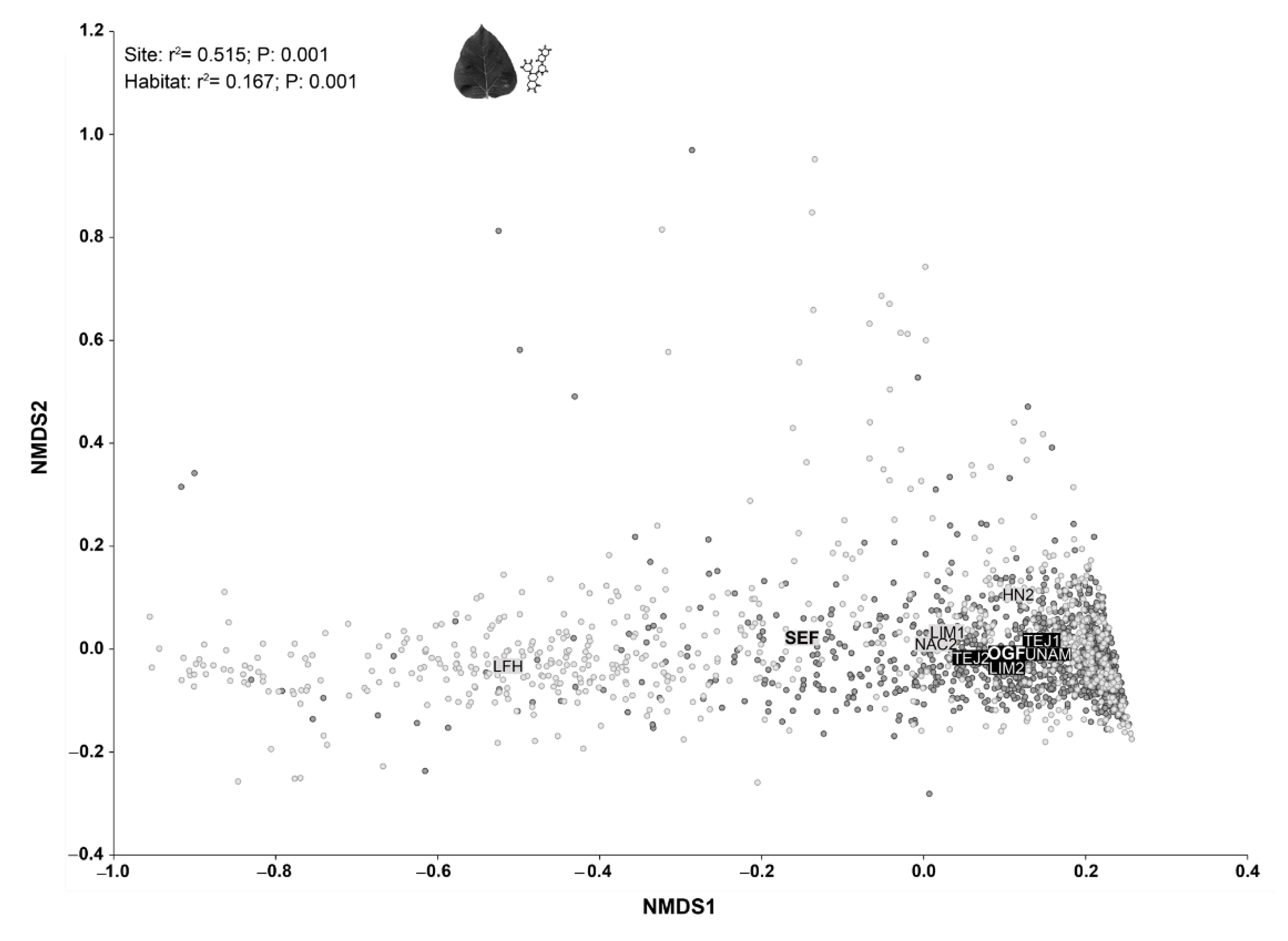
3.2. Contrasting OGF vs. SEF Plant Communities in Terms of Traits Composition
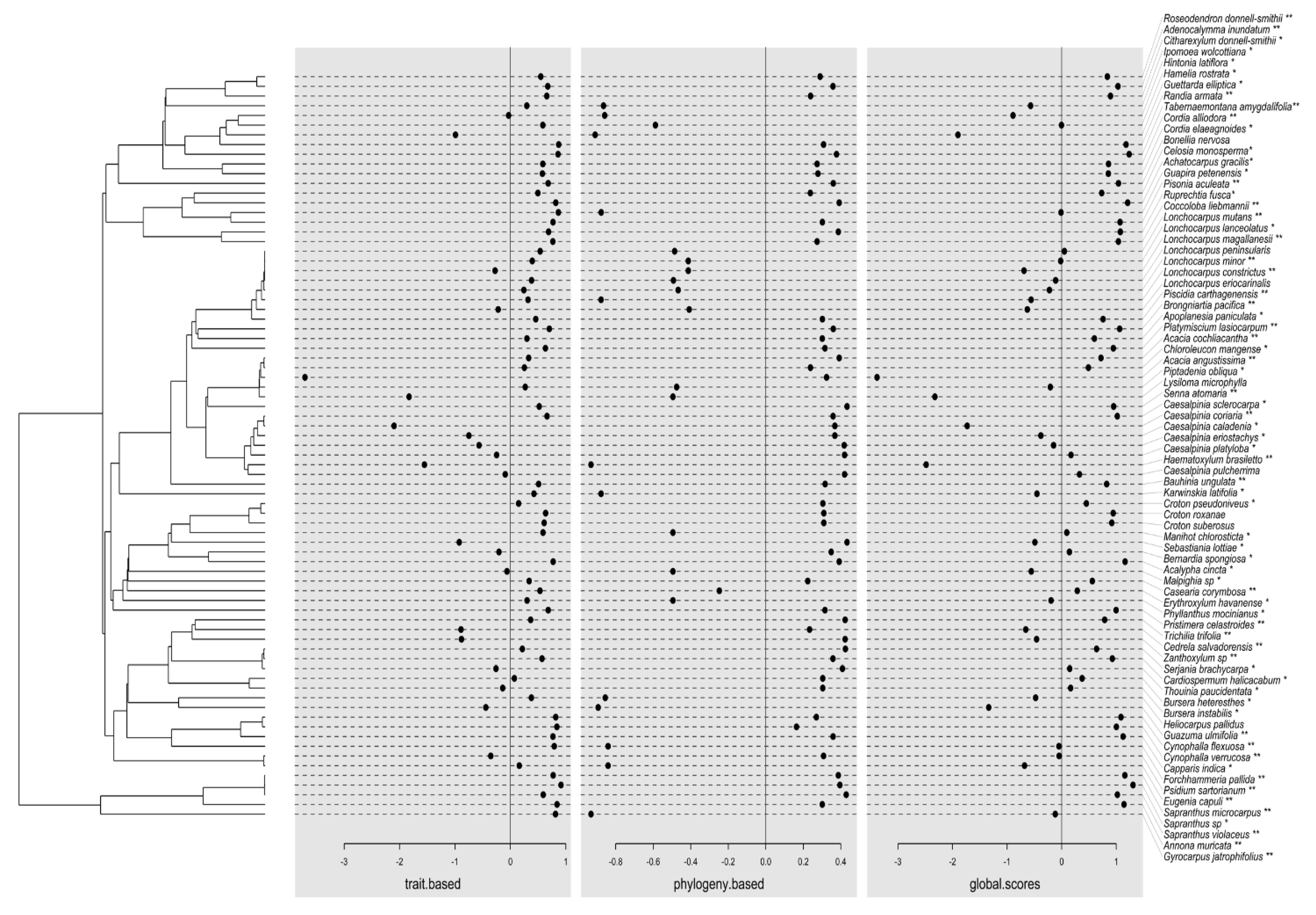
3.3. External and Internal Filters Underlying Species Coexistence in Old-Growth Forests and Secondary Forests
3.4. Potential Environmental Drivers of Defensive Traits Variation
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| Average of photosynthetically active radiatio | (PARavg) |
| Average soil temperatur | (STavg) |
| Chlorophyll conten | (CC) |
| Community-weighted mea | (CWM) |
| Diameter at breast heigh | (DBH) |
| Environmental/air average humidit | (EHavg) |
| Environmental/air average temperatur | (ETavg) |
| Growth-differentiation balanc | (GDB) hypothesis |
| Leaf densit | (LD) |
| Leaf fresh mas | (LFM) |
| Maximum sun radiatio | (SRmax) |
| Nitroge | (N) |
| Non-Metric Multidimensional Scalin | (NMDS) |
| Old-growth fores | (OGF) |
| Phosphomonoesteras | (PME) |
| Phosphoru | (Pho) |
| p-nitropheno | (pNP) |
| Secondary fores | (SEF) |
| Soil water conten | (SWC) |
| Specific leaf are | (SLA) |
| Standardized effect siz | (SES) |
| Total carbo | (TC) |
| Total flavonoid | (Flavonoids) |
| Total nitroge | (TN) |
| Total organic carbo | (TOC) |
| Total phenol | (Phenols) |
| Total phosphoru | (TPho) |
| Total tannin | (Tannis) |
| Tropical dry fores | (TDF) |
| T-statistic | (TIP/IC, TIC/IR, and TPC/PR) |
| Universal buffe | (MUB) |
| β-1, 4-glucosidas | (BG) |
| β-1, 4-N-acetylglucosaminidas | (NAG) |
Appendix A. Reagents Used for Secondary Metabolites Quantification
Appendix A.1. Reagents
- -
- Aluminum chloride (Sigma-Aldrich 206911)
- -
- (+)-Catechin hydrate ((Sigma-Aldrich C1251)
- -
- Folin and Ciocalteu’s phenol reagent (Sigma-Aldrich F9252)
- -
- Gallic acid monohydrate (Sigma-Aldrich 398225)
- -
- Methanol (Sigma-Aldrich 179337)
- -
- Polyvinylpyrrolidone (PVP) (Sigma-Aldrich PVP40)
- -
- Sodium carbonate (Sigma-Aldrich S7795)
- -
- Sodium nitrite (Sigma-Aldrich S2252)
- -
- Sodium hydroxide (Sigma-Aldrich S5881)
- -
- Water deionized and distilled (dd H2O)
Appendix A.2. Reagents Setup
- -
- Aluminum chloride: Prepare a solution of 10% in methanol.
- -
- Folin–Ciocalteu reagent: Prepare a solution of 10% (v/v) F&C in water.
- -
- Methanol: Prepare a solution of 95% (v/v) methanol in water.
- -
- Sodium carbonate: Prepare a solution of 700 mM Na2CO3 in water.
- -
- Sodium hydroxide: Prepare a solution of 1 M NaOH in water.
- -
- Sodium nitrite: Prepare a solution of 5% NaN02 in water.
References
- Westoby, M.; Wright, I.J. Land-plant ecology on the basis of functional traits. Trends Ecol. Evol. 2006, 21, 261–268. [Google Scholar] [CrossRef]
- Pérez-Harguindeguy, N.; Díaz, S.; Garnier, E.; Lavorel, S.; Poorter, H.; Jaureguiberry, P.; Bret-Harte, M.S.; Cornwell, W.K.; Craine, J.M.; Gurvich, D.E.; et al. New handbook for standardised measurement of plant functional traits worldwide. Aust. J. Bot. 2013, 61, 167–234. [Google Scholar] [CrossRef]
- Engelbrecht, B.M.J.; Comita, L.S.; Condit, R.; Kursar, T.A.; Tyree, M.T.; Turner, B.L.; Hubbell, S.P. Drought sensitivity shapes species distribution patterns in tropical forests. Nature 2007, 447, 80–82. [Google Scholar] [CrossRef] [PubMed]
- Kraft, N.J.B.; Adler, P.B.; Godoy, O.; James, E.C.; Fuller, S.; Levine, J.M. Community assembly, coexistence and the environmental filtering metaphor. Funct. Ecol. 2015, 29, 592–599. [Google Scholar] [CrossRef]
- Callaway, R.M.; Brooker, R.W.; Choler, P.; Kikvidze, Z.; Lortie, C.J.; Michalet, R.; Paolini, L.; Pugnaire, F.I.; Newingham, B.; Aschehoug, E.T.; et al. Positive interactions among alpine plants increase with stress. Nature 2002, 417, 844–848. [Google Scholar] [CrossRef]
- Benítez-Malvido, J. Fungal diseases in Neotropical forests disturbed by humans. In New Directions in Conservation Medicine: Applied Cases of Ecological Health; Aguirre, A., Daszak, P., Ostfeld, R.S., Eds.; Oxford University Press: New York, NY, USA, 2012; p. 302. [Google Scholar]
- Pagán, I.; González-Jara, P.; Moreno-Letelier, A.; Rodelo-Urrego, M.; Fraile, A.; Piñero, D.; García-Arenal, F. Effect of biodiversity changes in disease risk: Exploring disease emergence in a plant-virus system. PLoS Pathog. 2012, 8, e1002796. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ostfeld, R.S.; Keesing, F. Effects of Host Diversity on Infectious Disease. Annu. Rev. Ecol. Evol. Syst. 2012, 43, 157–182. [Google Scholar] [CrossRef]
- Richards, L.A.; Dyer, L.A.; Forister, M.L.; Smilanich, A.M.; Dodson, C.D.; Leonard, M.D.; Jeffrey, C.S. Phytochemical diversity drives plant-insect community diversity. Proc. Natl. Acad. Sci. USA 2015, 112, 10973–10978. [Google Scholar] [CrossRef] [Green Version]
- Kessler, A.; Kalske, A. Plant secondary metabolite diversity and species interactions. Annu. Rev. Ecol. Evol. Syst. 2018, 49, 115–138. [Google Scholar] [CrossRef]
- Lämke, J.S.; Unsicker, S.B. Phytochemical variation in treetops: Causes and consequences for tree-insect herbivore interactions. Oecologia 2018, 187, 377–388. [Google Scholar] [CrossRef]
- Herms, D.A.; Mattson, W.J. The Dilemma of Plants: To Grow or Defend. Q. Rev. Biol. 1992, 67, 283–335. [Google Scholar] [CrossRef] [Green Version]
- Loomis, W.E. Growth-differentiation balance vs. carbohydrate-nitrogen ratio. Proc. Am. Soc. Hortic. Sci. 1932, 29, 240–245. [Google Scholar]
- Salleo, S.; Nardini, A. Sclerophylly: Evolutionary advantage or mere epiphenomenon? Plant Biosyst.—Int. J. Deal. Asp. Plant Biol. 2000, 134, 247–259. [Google Scholar] [CrossRef]
- Fine, P.V.A.; Miller, Z.J.; Mesones, I.; Irazuzta, S.; Appel, H.M.; Stevens, M.H.H.; Sääksjärvi, I.; Schultz, J.C.; Coley, P.D. The growth-defense trade-off and habitat specialization by plants in Amazonian forests. Ecology 2006, 87, 150–162. [Google Scholar] [CrossRef] [Green Version]
- Wright, I.J.; Reich, P.B.; Westoby, M.; Ackerly, D.D.; Baruch, Z.; Bongers, F.; Cavender-Bares, J.; Chapin, T.; Cornelissen, J.H.C.; Diemer, M.; et al. The worldwide leaf economics spectrum. Nature 2004, 428, 821–827. [Google Scholar] [CrossRef]
- Stamp, N. Out of the quagmire of plant defense hypotheses. Q. Rev. Biol. 2003, 78, 23–55. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Massad, T.J.; Fincher, R.M.; Smilanich, A.M.; Dyer, L. A quantitative evaluation of major plant defense hypotheses, nature versus nurture, and chemistry versus ants. Arthropod. Plant Interact. 2011, 5, 125–139. [Google Scholar] [CrossRef]
- Massad, T.J.; Dyer, L.A.; Vega, G.C. Costs of Defense and a Test of the Carbon-Nutrient Balance and Growth-Differentiation Balance Hypotheses for Two Co-Occurring Classes of Plant Defense. PLoS ONE 2012, 7, e47554. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alvarez-Añorve, M.Y.; Quesada, M.; Arturo Sánchez-Azofeifa, G.; Avila-Cabadilla, L.D.; Gamon, J.A. Functional regeneration and spectral reflectance of trees during succession in a highly diverse tropical dry forest ecosystem. Am. J. Bot. 2012, 99, 816–826. [Google Scholar] [CrossRef] [PubMed]
- Lojka, B.; Krausová, J.; Kubík, Š.; Polesný, Z. Assessment of insect biological diversity in various land use systems in the Peruvian Amazon. In Amazon Basin: Plant Life, Wildlife and Environment; Nova Science Publishers: New York, NY, USA, 2010; pp. 103–121. [Google Scholar]
- Lebrija-Trejos, E.; Pérez-García, E.A.; Meave, J.A.; Poorter, L.; Bongers, F. Environmental changes during secondary succession in a tropical dry forest in Mexico. J. Trop. Ecol. 2011, 27, 477–489. [Google Scholar] [CrossRef] [Green Version]
- Albert, C.H.; Thuiller, W.; Yoccoz, N.G.; Douzet, R.; Aubert, S.; Lavorel, S. A multi-trait approach reveals the structure and the relative importance of intra- vs. interspecific variability in plant traits. Funct. Ecol. 2010, 24, 1192–1201. [Google Scholar] [CrossRef]
- Messier, J.; McGill, B.J.; Lechowicz, M.J. How do traits vary across ecological scales? A case for trait-based ecology. Ecol. Lett. 2010, 13, 838–848. [Google Scholar] [CrossRef]
- Violle, C.; Enquist, B.J.; McGill, B.J.; Jiang, L.; Albert, C.H.; Hulshof, C.; Jung, V.; Messier, J. The return of the variance: Intraspecific variability in community ecology. Trends Ecol. Evol. 2012, 27, 244–252. [Google Scholar] [CrossRef]
- Olson, D.M.; Dinerstein, E.; Wikramanayake, E.D.; Burgess, N.D.; Powell, G.V.N.; Underwood, E.C.; D’amico, J.A.; Itoua, I.; Strand, H.E.; Morrison, J.C. Terrestrial Ecoregions of the World: A New Map of Life on Earth. Bioscience 2001, 51, 933–938. [Google Scholar] [CrossRef]
- Lott, E.J.; Atkinson, T.H. Mexican and Central American Seasonally Dry Tropical Forests: Chamela-Cuixmala, Jalisco, as a Focal Point for Comparison. In Neotropical Savannas and Seasonally Dry Forests: Plant Diversity, Biogeography, and Conservation; CRC Press: Boca Raton, FL, USA, 2006; pp. 307–334. [Google Scholar]
- Flores-Casas, R.; Ortega-Huerta, M.A. Modelling land cover changes in the tropical dry forest surrounding the Chamela-Cuixmala biosphere reserve, Mexico. Int. J. Remote Sens. 2019, 40, 6948–6974. [Google Scholar] [CrossRef]
- Jimenez-Rodríguez, D.L.; Alvarez-Añorve, M.Y.; Flores-Puerto, J.I.; Oyama, K.; Avila-Cabadilla, L.D.; Pineda-Cortes, M.; Benítez-Malvido, J. Structural and functional traits predict short term response of tropical dry forests to a high intensity hurricane. For. Ecol. Manag. 2018, 426, 101–114. [Google Scholar] [CrossRef]
- Cornelissen, J.H.C.; Lavorel, S.; Garnier, E.; Díaz, S.; Buchmann, N.; Gurvich, D.E.; Reich, P.B.; Ter Steege, H.; Morgan, H.D.; Van Der Heijden, M.G.A.; et al. A handbook of protocols for standardised and easy measurement of plant functional traits worldwide. Aust. J. Bot. 2003, 51, 335–380. [Google Scholar] [CrossRef] [Green Version]
- Koricheva, J.; Larsson, S.; Haukioja, E.; Keinänen, M.; Keinanen, M. Regulation of Woody Plant Secondary Metabolism by Resource Availability: Hypothesis Testing by Means of Meta-Analysis. Oikos 1998, 83, 212. [Google Scholar] [CrossRef]
- Ainsworth, E.A.; Gillespie, K.M. Estimation of total phenolic content and other oxidation substrates in plant tissues using Folin-Ciocalteu reagent. Nat. Protoc. 2007, 2, 875–877. [Google Scholar] [CrossRef]
- Bravo-Monzón, A.E.; Montiel-González, C.; Avila-Cabadilla, L.D.; Alvarez-Añorve, M.Y. Small-scale determination of total phenols, tannins, and flavonoids from foliar tissue using colorimetric assays. Protoc. Exch. 2021. [Google Scholar] [CrossRef]
- Makkar, H.P.S.; Siddhuraju, P.; Becker, K. Tannins. In Plant Secondary Metabolites; Humana Press: Totowa, NJ, USA, 2007; pp. 67–81. ISBN 978-1-59745-425-4. [Google Scholar]
- Marinova, D.; Ribarova, F.; Atanassova, M. Total phenolic and total flavonoids in Bulgarian fruits and vegetables. J. Univ. Chem. Technol. Metall. 2005, 40, 255–260. [Google Scholar]
- Reynolds, S.G. The gravimetric method of soil moisture determination Part I A study of equipment, and methodological problems. J. Hydrol. 1970, 11, 258–273. [Google Scholar] [CrossRef]
- Jones, D.L.; Willett, V.B. Experimental evaluation of methods to quantify dissolved organic nitrogen (DON) and dissolved organic carbon (DOC) in soil. Soil Biol. Biochem. 2006, 38, 991–999. [Google Scholar] [CrossRef]
- Huffman, E.W.D. Performance of a new automatic carbon dioxide coulometer. Microchem. J. 1977, 22, 567–573. [Google Scholar] [CrossRef]
- Bremner, M. Chapter 37: Nitrogen-Total. In Methods of Soil Analysis: Part 3 Chemical Methods 5; Wiley Online Library: Hoboken, NJ, USA, 1996; pp. 1085–1121. [Google Scholar]
- Murphy, J.; Riley, J.P. A modified single solution method for the determination of phosphate in natural waters. Anal. Chem. Acta 1962, 27, 31–36. [Google Scholar] [CrossRef]
- Tabatabai, M.A.; Bremner, J.M. Use of p-nitrophenyl phosphate for assay of soil phosphatase activity. Soil Biol. Biochem. 1969, 1, 301–307. [Google Scholar] [CrossRef]
- Eivazi, F.; Tabatabai, M.A. Phosphatases in soils. Soil Biol. Biochem. 1977, 2, 167–172. [Google Scholar] [CrossRef]
- Eivazi, F.; Tabatabai, M.A. Glucosidases and galactosidases in soils. Soil Biol. Biochem. 1988, 20, 601–606. [Google Scholar] [CrossRef]
- Verchot, L.V.; Borelli, T. Application of para-nitrophenol (pNP) enzyme assays in degraded tropical soils. Soil Biol. Biochem. 2005, 37, 625–633. [Google Scholar] [CrossRef]
- Keck, F.; Rimet, F.; Bouchez, A.; Franc, A. phylosignal: An R package to measure, test, and explore the phylogenetic signal. Ecol. Evol. 2016, 6, 2774–2780. [Google Scholar] [CrossRef] [PubMed]
- Zanne, A.E.; Tank, D.C.; Cornwell, W.K.; Eastman, J.M.; Smith, S.A.; FitzJohn, R.G.; McGlinn, D.J.; O’Meara, B.C.; Moles, A.T.; Reich, P.B. Three keys to the radiation of angiosperms into freezing environments. Nature 2014, 506, 89–92. [Google Scholar] [CrossRef]
- Qian, H.; Jin, Y. An updated megaphylogeny of plants, a tool for generating plant phylogenies and an analysis of phylogenetic community structure. J. Plant Ecol. 2016, 9, 233–239. [Google Scholar] [CrossRef] [Green Version]
- Zheng, L.; Ives, A.R.; Garland, T.; Larget, B.R.; Yu, Y.; Cao, K. New multivariate tests for phylogenetic signal and trait correlations applied to ecophysiological phenotypes of nine Manglietia species. Funct. Ecol. 2009, 23, 1059–1069. [Google Scholar] [CrossRef]
- Veech, J.A. Significance testing in ecological null models. Theor. Ecol. 2012, 5, 611–616. [Google Scholar] [CrossRef]
- Gotelli, N.J.; Graves, G.R. Null Models in Ecology; Smithsonian Institution Press: Washington, DC, USA, 1996; ISBN 1560986573. [Google Scholar]
- Gurevitch, J.; Morrow, L.L.; Wallace, A.; Walsh, J.S. A Meta-Analysis of Competition in Field Experiments. Am. Nat. 1992, 140, 539–572. [Google Scholar] [CrossRef]
- Gotelli, N.J.; McCabe, D.J. Species co-occurrence: A meta-analysis of J. M. Diamond’s assembly rules model. Ecology 2002, 83, 2091–2096. [Google Scholar] [CrossRef]
- Pavoine, S.; Vela, E.; Gachet, S.; de Bélair, G.; Bonsall, M.B. Linking patterns in phylogeny, traits, abiotic variables and space: A novel approach to linking environmental filtering and plant community assembly. J. Ecol. 2011, 99, 165–175. [Google Scholar] [CrossRef]
- Dray, S.; Choler, P.; Dolédec, S.; Peres-Neto, P.R.; Thuiller, W.; Pavoine, S.; ter Braak, C.J.F. Combining the fourth-corner and the RLQ methods for assessing trait responses to environmental variation. Ecology 2014, 95, 14–21. [Google Scholar] [CrossRef] [Green Version]
- Dray, S.; Legendre, P. Testing the species traits-environment relationships: The fourth-corner problem revisited. Ecology 2008, 89, 3400–3412. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2019. [Google Scholar]
- Wei, T.; Simko, V. R Package “Corrplot”: Visualization of a Correlation Matrix; The Comprehensive R Archive Network, Vienna University of Economics and Business: Vienna, Austria, 2017. [Google Scholar]
- Harrell, F.E., Jr. Hmisc: Harrell Miscellaneous; The Comprehensive R Archive Network, Vienna University of Economics and Business: Vienna, Austria, 2021. [Google Scholar]
- Paradis, E.; Schliep, K. Ape 5.0: An environment for modern phylogenetics and evolutionary analyses in R. Bioinformatics 2019, 35, 526–528. [Google Scholar] [CrossRef]
- Oksanen, J.; Blanchet, F.G.; Friendly, M.; Kindt, R.; Legendre, P.; McGlinn, D.; Minchin, P.R.; O’Hara, R.B.; Simpson, G.L.; Solymos, P.; et al. Vegan: Community Ecology Package; The Comprehensive R Archive Network, Vienna University of Economics and Business: Vienna, Austria, 2019. [Google Scholar]
- Taudiere, A.; Violle, C. Cati: An R package using functional traits to detect and quantify multi-level community assembly processes. Ecography 2016, 39, 699–708. [Google Scholar] [CrossRef]
- Dray, S.; Dufour, A.-B. The {ade4} Package: Implementing the Duality Diagram for Ecologists. J. Stat. Softw. 2007, 22, 1–20. [Google Scholar] [CrossRef] [Green Version]
- Pavoine, S. Adiv: An R package to analyse biodiversity in ecology. Methods Ecol. Evol. 2020, 11, 1106–1112. [Google Scholar] [CrossRef]
- McCune, B.; Grace, J.B.; Urban, D.L. Analysis of Ecological Communities; MjM software design: Gleneden Beach, OR, USA, 2002. [Google Scholar]
- Quesada, M.; Álvarez-Añorve, M.; Ávila-Cabadilla, L.; Castillo, A.; Lopezaraiza-Mikel, M.; Martén-Rodríguez, S.; Rosas-Guerrero, V.; Sáyago, R.; Sánchez-Montoya, G.; Contreras-Sánchez, J.M.; et al. Tropical dry forest ecological succession in Mexico: Synthesis of a long-term study. In Tropical Dry Forests in the Americas; CRC Press: Boca Raton, FL, USA, 2013; pp. 17–34, ISBN 978-1-4665-1201-6; 978-1-4665-1200-9. [Google Scholar]
- Jones, C.G.; Hartley, S.E. A Protein Competition Model of Phenolic Allocation. Oikos 1999, 86, 27. [Google Scholar] [CrossRef]
- Meyer, S.; Cerovic, Z.G.; Goulas, Y.; Montpied, P.; Demotes-Mainard, S.; Bidel, L.P.R.; Moya, I.; Dreyer, E. Relationships between optically assessed polyphenols and chlorophyll contents, and leaf mass per area ratio in woody plants: A signature of the carbon-nitrogen balance within leaves? Plant Cell Environ. 2006, 29, 1338–1348. [Google Scholar] [CrossRef]
- Ibrahim, M.H.; Jaafar, H.Z.E.; Rahmat, A.; Rahman, Z.A. Effects of nitrogen fertilization on synthesis of primary and secondary metabolites in three varieties of kacip fatimah (Labisia pumila blume). Int. J. Mol. Sci. 2011, 12, 5238–5254. [Google Scholar] [CrossRef]
- Caretto, S.; Linsalata, V.; Colella, G.; Mita, G.; Lattanzio, V. Carbon fluxes between primary metabolism and phenolic pathway in plant tissues under stress. Int. J. Mol. Sci. 2015, 16, 26378–26394. [Google Scholar] [CrossRef] [Green Version]
- Kitajima, K.; Llorens, A.M.; Stefanescu, C.; Timchenko, M.V.; Lucas, P.W.; Wright, S.J. How cellulose-based leaf toughness and lamina density contribute to long leaf lifespans of shade-tolerant species. New Phytol. 2012, 195, 640–652. [Google Scholar] [CrossRef]
- Witkowski, E.T.F.; Lamont, B.B. Leaf specific mass confounds leaf density and thickness. Oecologia 1991, 88, 486–493. [Google Scholar] [CrossRef]
- Garnier, E.; Cordonnier, P.; Guillerm, J.-L.; Sonié, L. Specific leaf area and leaf nitrogen concentration in annual and perennial grass species growing in Mediterranean old-fields. Oecologia 1997, 111, 490–498. [Google Scholar] [CrossRef]
- Hanley, M.E.; Lamont, B.B. Relationships between physical and chemical attributes of congeneric seedlings: How important is seedling defence? Funct. Ecol. 2002, 16, 216–222. [Google Scholar] [CrossRef]
- Read, J.; Sanson, G.D.; Caldwell, E.; Clissold, F.J.; Chatain, A.; Peeters, P.; Lamont, B.B.; De Garine-Wichatitsky, M.; Jaffré, T.; Kerr, S. Correlations between leaf toughness and phenolics among species in contrasting environments of Australia and New Caledonia. Ann. Bot. 2009, 103, 757–767. [Google Scholar] [CrossRef]
- García-Guzmán, G.; Morales, E. Life-History Strategies of Plant Pathogens. Ecology 2007, 88, 589–596. [Google Scholar] [CrossRef] [PubMed]
- Gilbert, G.S.; Parker, I.M. The Evolutionary Ecology of Plant Disease: A Phylogenetic Perspective. Annu. Rev. Phytopathol. 2016, 54, 549–578. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Chen, L.; Liu, M.; García-Guzmán, G.; Gilbert, G.S.; Zhou, S. Dilution effect of plant diversity on infectious diseases: Latitudinal trend and biological context dependence. Oikos 2020, 129, 457–465. [Google Scholar] [CrossRef] [Green Version]
- Bazzaz, F.A. The physiological ecology of plant succession. Annu. Rev. Ecol. Syst. 1979, 10, 351–371. [Google Scholar] [CrossRef] [Green Version]
- Kalisz, S.; Oszmiański, J.; Wojdyło, A. Increased content of phenolic compounds in pear leaves after infection by the pear rust pathogen. Physiol. Mol. Plant Pathol. 2015, 91, 113–119. [Google Scholar] [CrossRef]
- Fonseca, M.B.; Silva, J.O.; Falcão, L.A.D.; Dupin, M.G.V.; Melo, G.A.; Espírito-Santo, M.M. Leaf damage and functional traits along a successional gradient in Brazilian tropical dry forests. Plant Ecol. 2018, 219, 403–415. [Google Scholar] [CrossRef]
- García-Guzmán, G.; Trejo, I.; Sánchez-Coronado, M.E. Foliar diseases in a seasonal tropical dry forest: Impacts of habitat fragmentation. For. Ecol. Manag. 2016, 369, 126–134. [Google Scholar] [CrossRef]
- Hubbell, S.P. Tree dispersion, abundance, and diversity in a tropical dry forest. Science 1979, 203, 1299–1309. [Google Scholar] [CrossRef] [Green Version]
- Clark, J.S. Individuals and the Variation Needed for High Species Diversity in Forest Trees. Science 2010, 327, 1129–1132. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clark, J.S.; Dietze, M.; Chakraborty, S.; Agarwal, P.K.; Ibanez, I.; LaDeau, S.; Wolosin, M. Resolving the biodiversity paradox. Ecol. Lett. 2007, 10, 647–659. [Google Scholar] [CrossRef] [PubMed]
- Neves, F.S.; Silva, J.O.; Espírito-Santo, M.M.; Fernandes, G.W. Insect Herbivores and Leaf Damage along Successional and Vertical Gradients in a Tropical Dry Forest. Biotropica 2014, 46, 14–24. [Google Scholar] [CrossRef]
- Keesing, F.; Belden, L.K.; Daszak, P.; Dobson, A.; Harvell, C.D.; Holt, R.D.; Hudson, P.; Jolles, A.; Jones, K.E.; Mitchell, C.E.; et al. Impacts of biodiversity on the emergence and transmission of infectious diseases. Nature 2010, 468, 647–652. [Google Scholar] [CrossRef] [PubMed]
- Keesing, F.; Holt, R.D.; Ostfeld, R.S. Effects of species diversity on disease risk. Ecol. Lett. 2006, 9, 485–498. [Google Scholar] [CrossRef]
- Lankau, R.A. Specialist and generalist herbivores exert opposing selection on a chemical defense. New Phytol. 2007, 175, 176–184. [Google Scholar] [CrossRef]
- Benton, J.J.J. Plant Nutrition and Soil Fertility Manual; CRC Press: Boca Raton, FL, USA, 2012; ISBN 9781439816103. [Google Scholar]
- Somma, F.; Hopmans, J.W.; Clausnitzer, V. Transient three-dimensional modeling of soil water and solute transport with simultaneous root growth, root water and nutrient uptake. Plant Soil 1998, 202, 281–293. [Google Scholar] [CrossRef]
- Bruijnzeel, L.A.; Waterloo, M.J.; Proctor, J.; Kuiters, A.T.; Kotterink, B. Hydrological observations in montane rain forests on Gunung Silam, Sabah, Malaysia, with special reference to the “Massenerhebung’’ effect. J. Ecol. 1993, 81, 145–167. [Google Scholar] [CrossRef]
- Mariano, N.A.; Martínez-Garza, C.; Alcalá, R.E. Differential herbivory and successional status in five tropical tree species. Rev. Mex. Biodivers. 2018, 89, 1107–1114. [Google Scholar] [CrossRef]
- Viani, R.A.G.; Rodrigues, R.R.; Dawson, T.E.; Lambers, H.; Oliveira, R.S. Soil pH accounts for differences in species distribution and leaf nutrient concentrations of Brazilian woodland savannah and seasonally dry forest species. Perspect. Plant Ecol. Evol. Syst. 2014, 16, 64–74. [Google Scholar] [CrossRef]
- Lott, E.J. Annotated checklist of the vascular flora of the Chamela Bay Region, Jalisco, Mexico. Occas. Pap. Calif. Acad. Sci. USA 1993, 148, 60. [Google Scholar]
- Silva, J.O.; Espírito-Santo, M.M.; Morais, H.C. Leaf traits and herbivory on deciduous and evergreen trees in a tropical dry forest. Basic Appl. Ecol. 2015, 16, 210–219. [Google Scholar] [CrossRef]
- Schimel, J.P.; Van Cleve, K.; Cates, R.G.; Clausen, T.P.; Reichardt, P.B. Effects of balsam poplar (Populus balsamifera) tannins and low molecular weight phenolics on microbial activity in taiga floodplain soil: Implications for changes in N cycling during succession. Can. J. Bot. 1996, 74, 84–90. [Google Scholar] [CrossRef]
- Fierer, N.; Schimel, J.P.; Cates, R.G.; Zou, J. Influence of balsam poplar tannin fractions on carbon and nitrogen dynamics in Alaskan taiga floodplain soils. Soil Biol. Biochem. 2001, 33, 1827–1839. [Google Scholar] [CrossRef]
- Kraus, T.E.C.; Zasoski, R.J.; Dahlgren, R.A.; Horwath, W.R.; Preston, C.M. Carbon and nitrogen dynamics in a forest soil amended with purified tannins from different plant species. Soil Biol. Biochem. 2004, 36, 309–321. [Google Scholar] [CrossRef]
- Portillo-Quintero, C.A.; Sánchez-Azofeifa, G.A. Extent and conservation of tropical dry forests in the Americas. Biol. Conserv. 2010, 143, 144–155. [Google Scholar] [CrossRef]
- Zhang, H.; Chen, Y.; Huang, L.; Ni, H. Predicting potential geographic distribution of Mikania micrantha planting based on ecological niche models in China. Nongye Gongcheng Xuebao/Trans. Chin. Soc. Agric. Eng. 2011, 27, 413–418. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
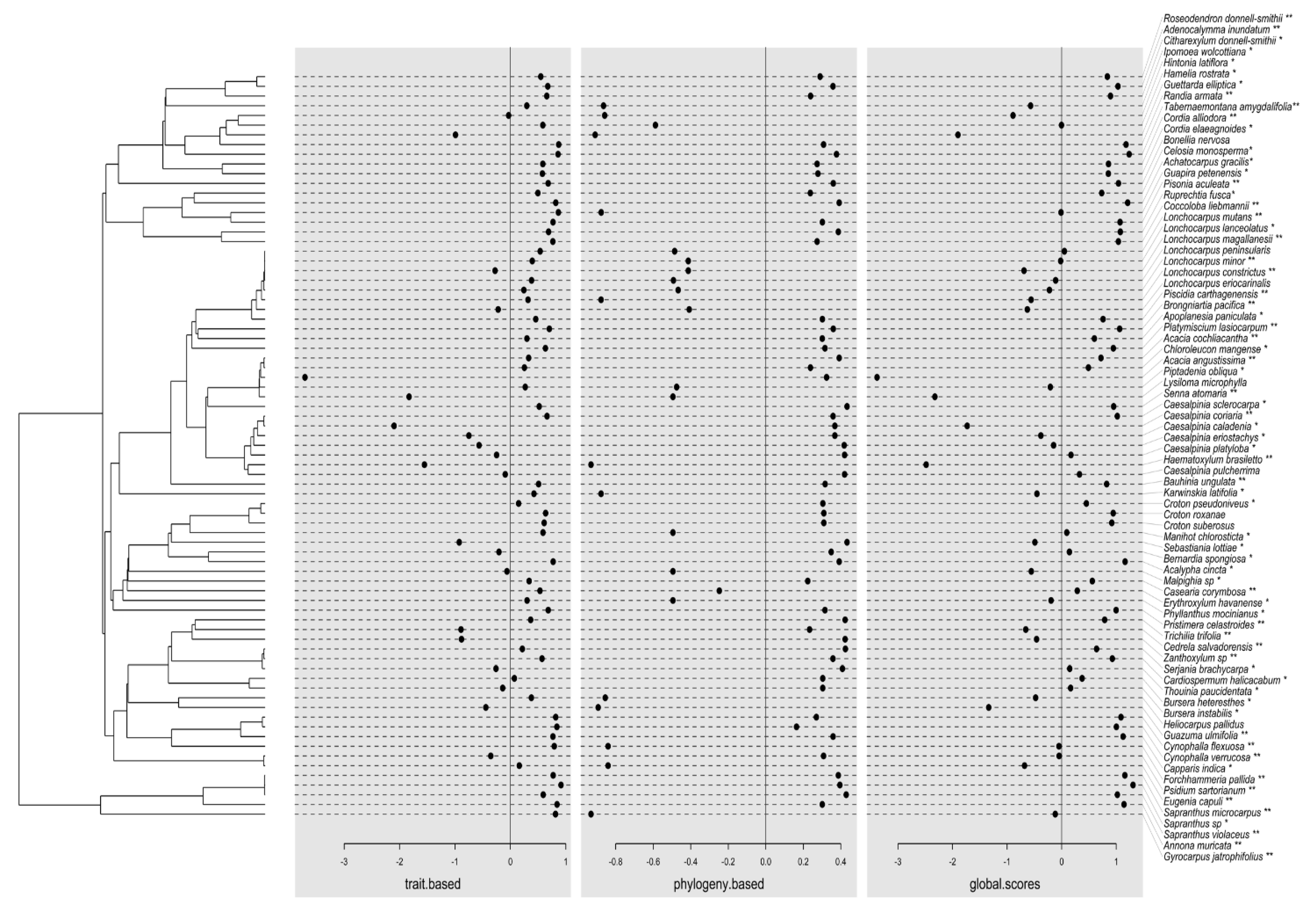{kind=link}
| Habitat Sites | NInd | S | Phenols | Tannins | Flavonoids | CC | SLA | LD | LFM |
|---|---|---|---|---|---|---|---|---|---|
| SF | |||||||||
| LFH | 256 | 2 | 140.47 | 36.52 | 26.14 | 32.30 | 125.47 | 0.13 | 0.02 |
| NAC2 | 81 | 8 | 35.46 | 9.58 | 20.41 | 37.39 | 137.85 | 0.22 | 0.02 |
| HN2 | 135 | 21 | 11.14 | 4.97 | 2.81 | 37.37 | 190.74 | 0.10 | 0.05 |
| LIM1 | 238 | 23 | 31.48 | 11.78 | 11.54 | 39.38 | 149.02 | 0.06 | 0.02 |
| Total | 710 | 43 | |||||||
| Mean/SD | 54.64/58.20 | 15.71/14.16 | 15.22/10.23 | 36.61/3.02 | 150.77/28.33 | 0.13/0.07 | 0.03/0.02 | ||
| OGF | |||||||||
| TEJ1 | 234 | 12 | 18.97 | 7.36 | 10.30 | 39.29 | 176.94 | 0.54 | 0.02 |
| TEJ2 | 244 | 11 | 31.99 | 9.92 | 13.83 | 36.76 | 174.01 | 0.10 | 0.03 |
| LIM2 | 206 | 13 | 25.45 | 8.71 | 7.84 | 34.46 | 119.00 | 0.11 | 0.02 |
| UNAM | 359 | 31 | 23.89 | 7.73 | 7.23 | 38.44 | 194.59 | 0.09 | 0.02 |
| Total | 1043 | 42 | – | – | – | ||||
| Grand total | 1753 | 77 | – | – | – | ||||
| Mean/SD | 25.08/5.37 | 8.43/1.15 | 9.8/3.00 | 37.24/2.13 | 166.14/32.71 | 0.21/0.22 | 0.02/0 | ||
| F/p-value | 1.02/0.35 | 1.05/0.0.35 | 1.04/0.35 | 0.12/0.75 | 0.50/0.50 | 0.51/0.50 | 0.4/0.55 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bravo-Monzón, Á.E.; Montiel-González, C.; Benítez-Malvido, J.; Arena-Ortíz, M.L.; Flores-Puerto, J.I.; Chiappa-Carrara, X.; Avila-Cabadilla, L.D.; Alvarez-Añorve, M.Y. The Assembly of Tropical Dry Forest Tree Communities in Anthropogenic Landscapes: The Role of Chemical Defenses. Plants 2022, 11, 516. https://doi.org/10.3390/plants11040516
Bravo-Monzón ÁE, Montiel-González C, Benítez-Malvido J, Arena-Ortíz ML, Flores-Puerto JI, Chiappa-Carrara X, Avila-Cabadilla LD, Alvarez-Añorve MY. The Assembly of Tropical Dry Forest Tree Communities in Anthropogenic Landscapes: The Role of Chemical Defenses. Plants. 2022; 11(4):516. https://doi.org/10.3390/plants11040516
Chicago/Turabian StyleBravo-Monzón, Ángel E., Cristina Montiel-González, Julieta Benítez-Malvido, María Leticia Arena-Ortíz, José Israel Flores-Puerto, Xavier Chiappa-Carrara, Luis Daniel Avila-Cabadilla, and Mariana Yolotl Alvarez-Añorve. 2022. "The Assembly of Tropical Dry Forest Tree Communities in Anthropogenic Landscapes: The Role of Chemical Defenses" Plants 11, no. 4: 516. https://doi.org/10.3390/plants11040516
APA StyleBravo-Monzón, Á. E., Montiel-González, C., Benítez-Malvido, J., Arena-Ortíz, M. L., Flores-Puerto, J. I., Chiappa-Carrara, X., Avila-Cabadilla, L. D., & Alvarez-Añorve, M. Y. (2022). The Assembly of Tropical Dry Forest Tree Communities in Anthropogenic Landscapes: The Role of Chemical Defenses. Plants, 11(4), 516. https://doi.org/10.3390/plants11040516