
The First Genetic Linkage Map of Winged Bean [Psophocarpus tetragonolobus (L.) DC.] and QTL Mapping for Flower-, Pod-, and Seed-Related Traits

 , , , ,
, , , ,
Abstract
:1. Introduction
2. Results
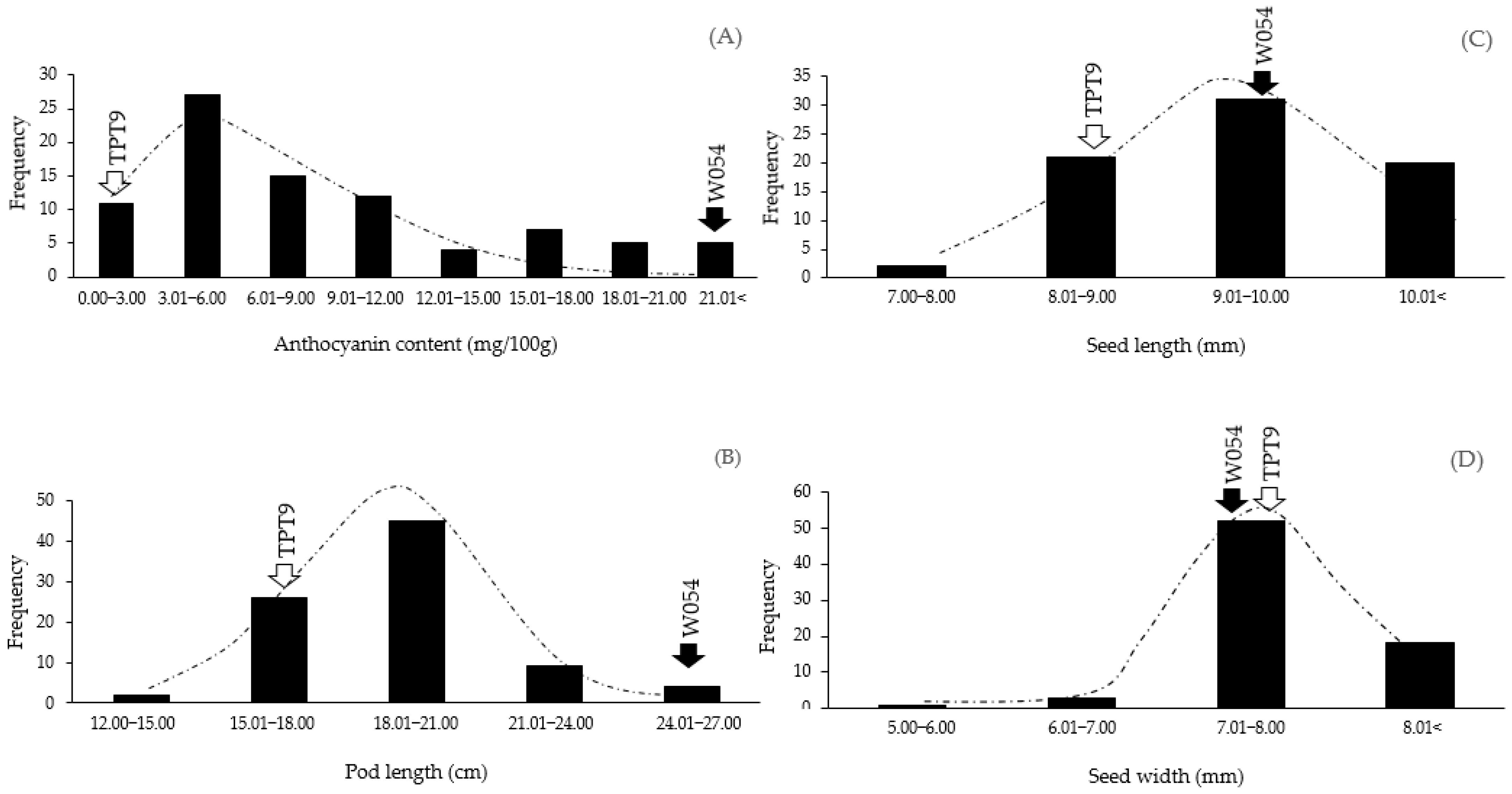
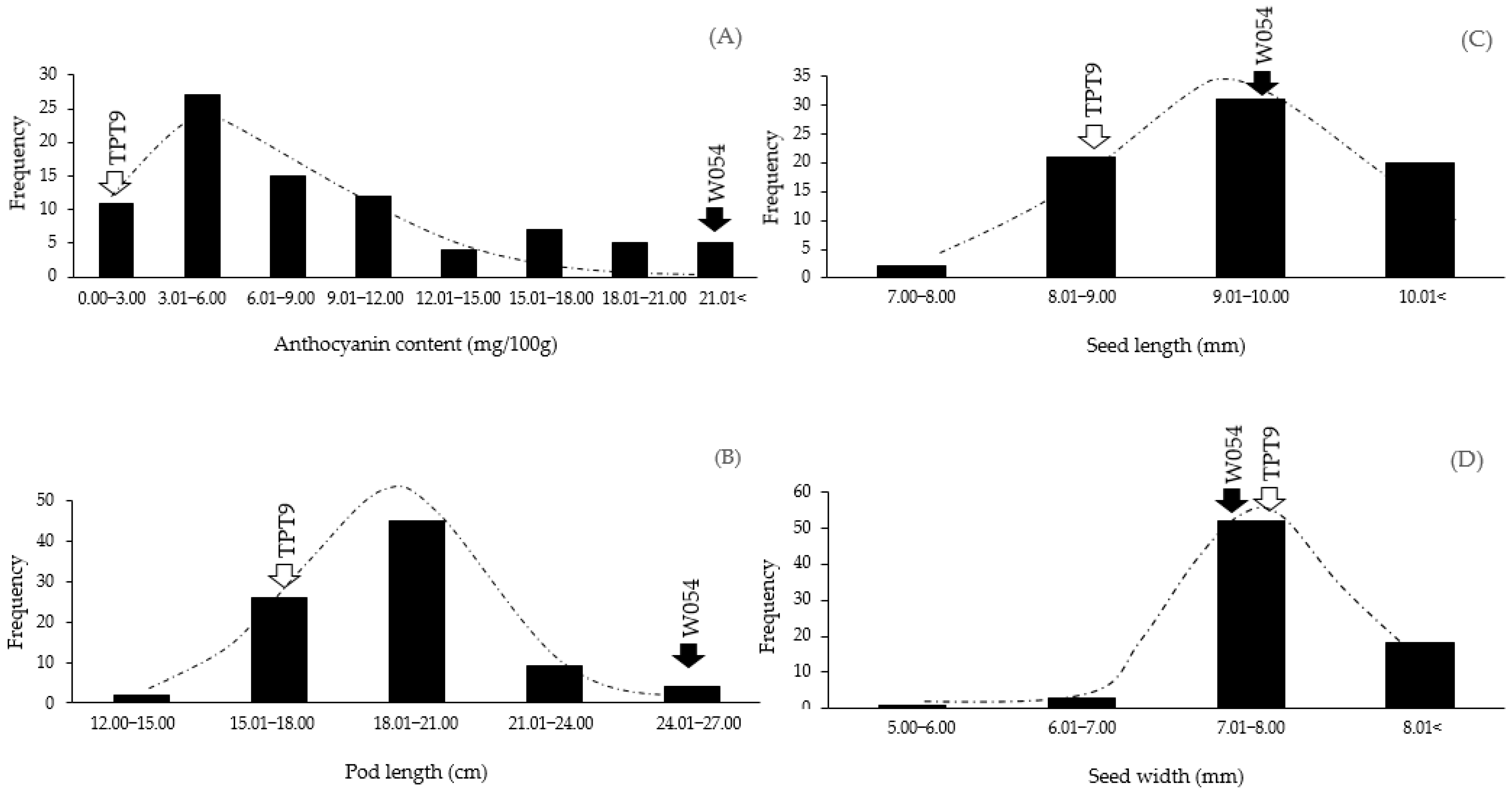
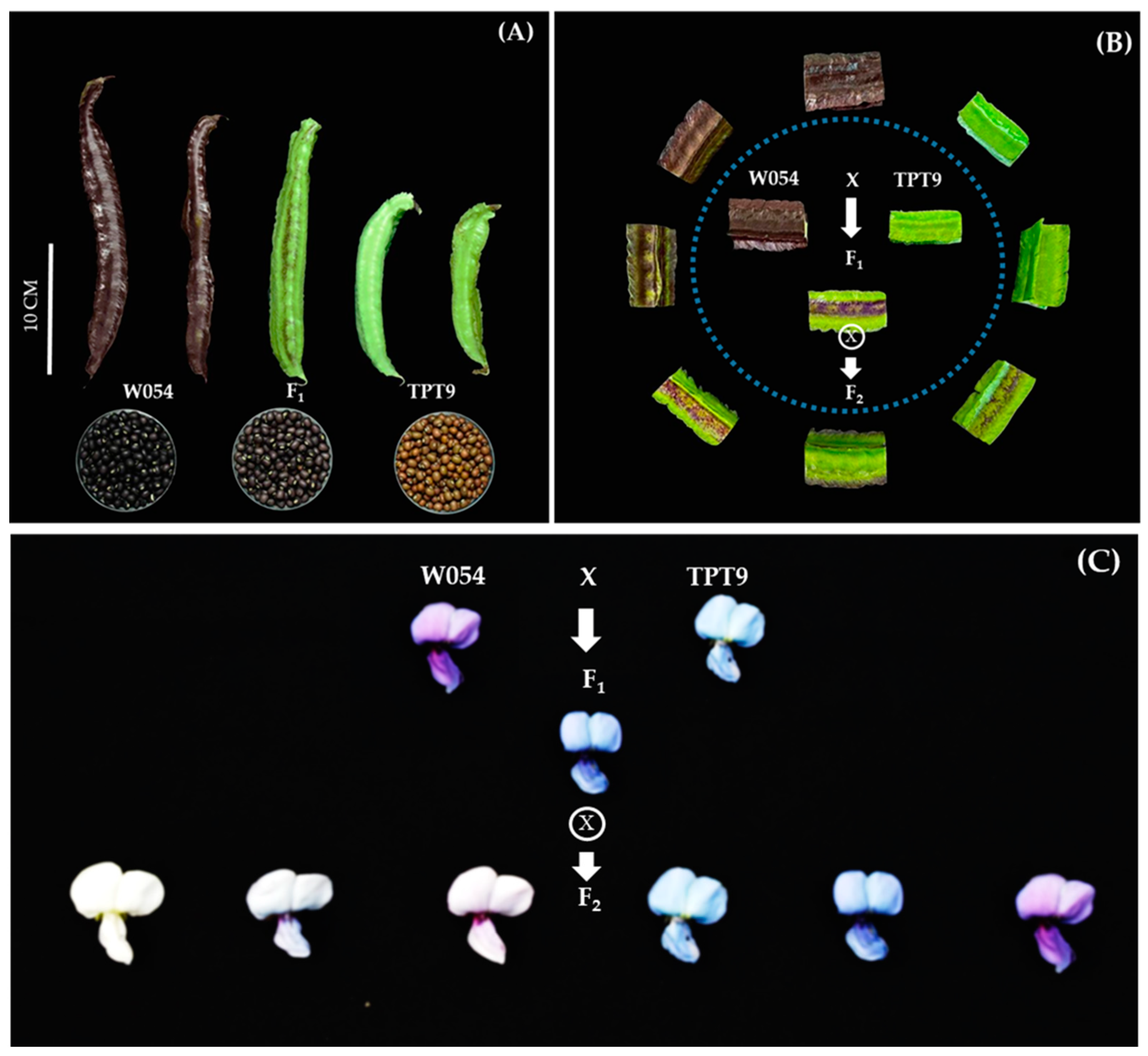
2.1. Variation in Pod Anthocyanin Content, Pod Length, Seed Length, and Seed Width
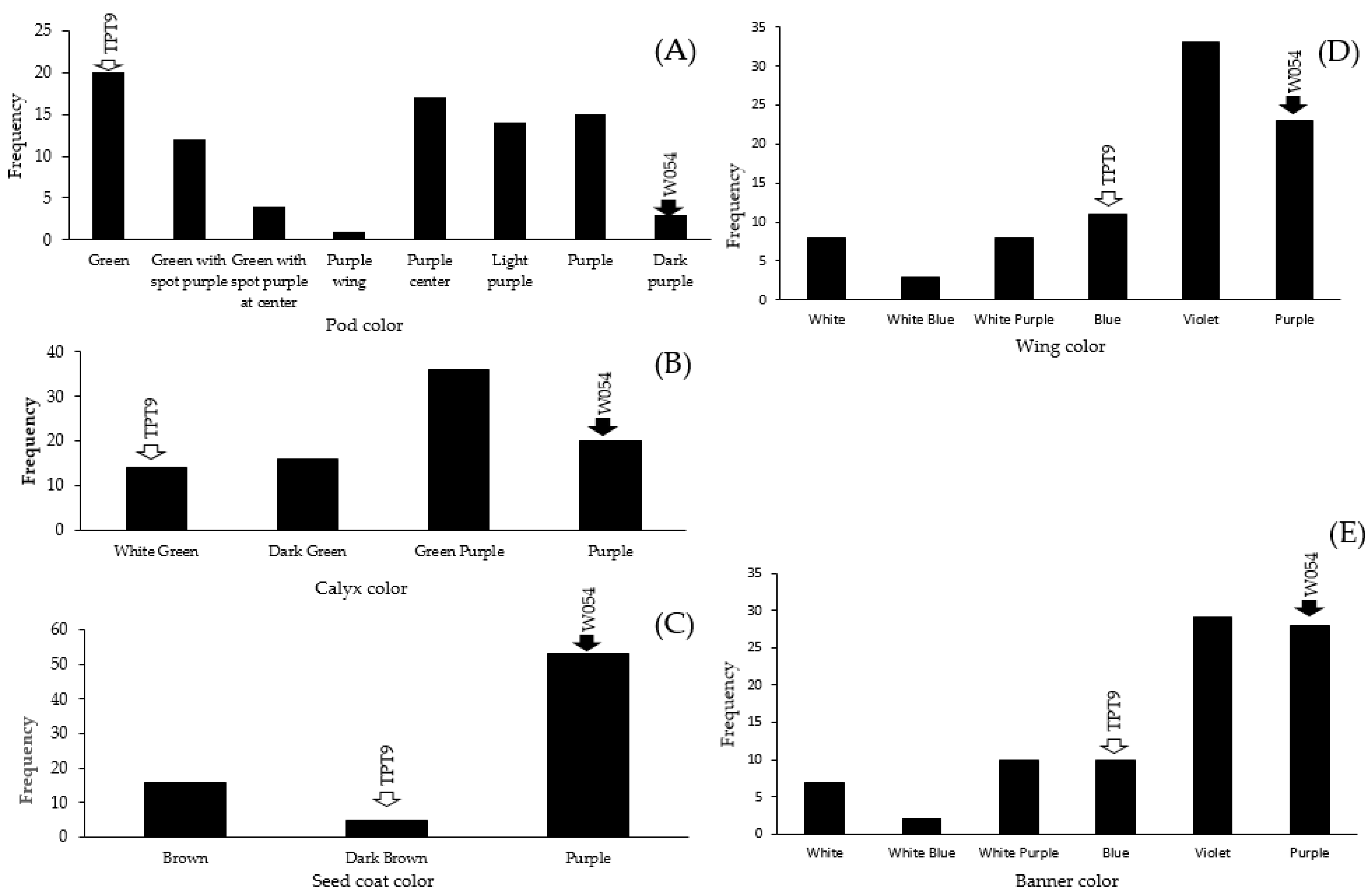
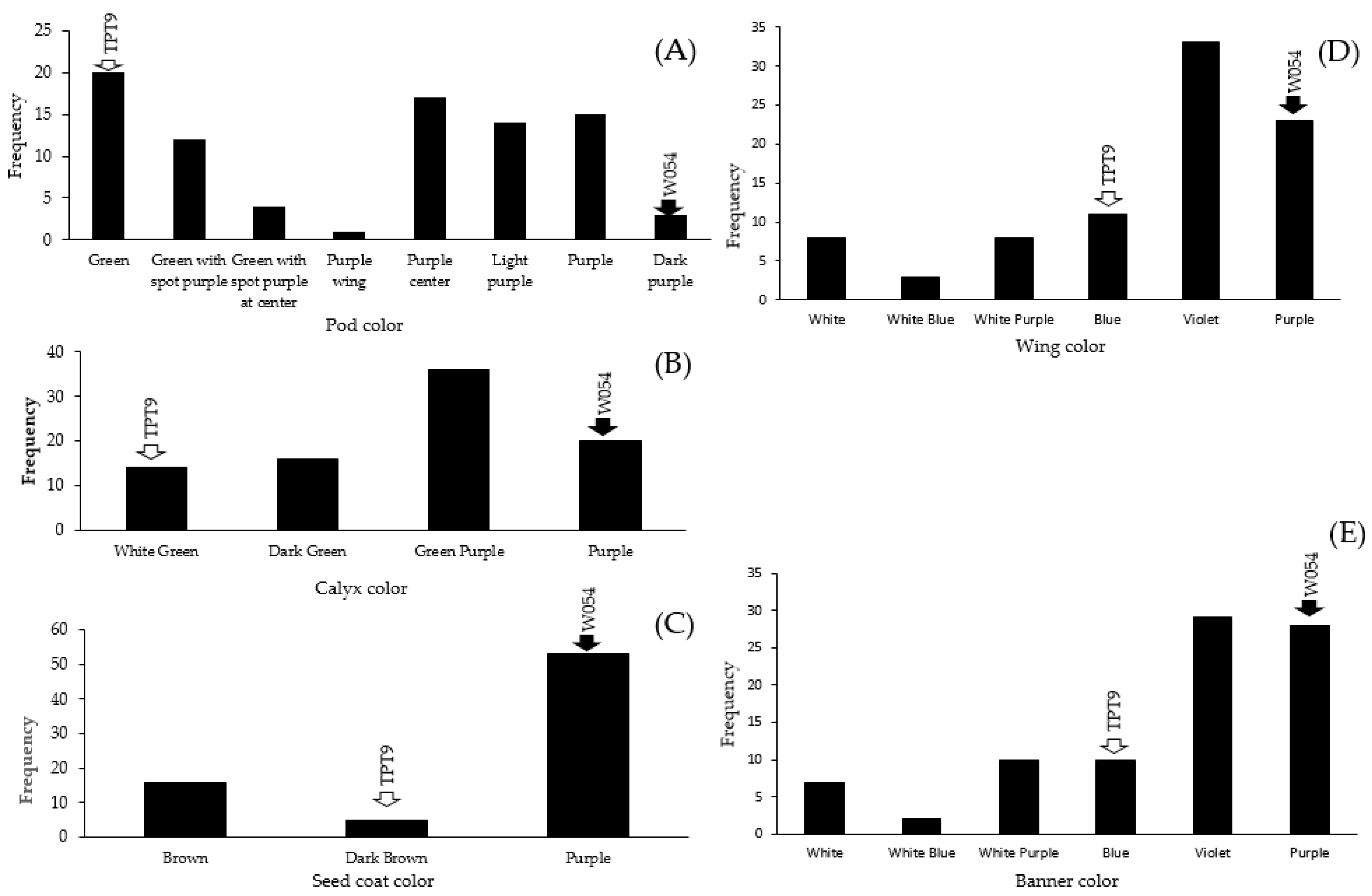
2.2. Variation in Organ Coloration
2.3. Construction of the Winged Bean Genetic Linkage Map
2.3.1. RAD-Seq Library Construction, Sequencing, and SNP Calling
2.3.2. Construction of the Winged Bean Genetic Linkage Map
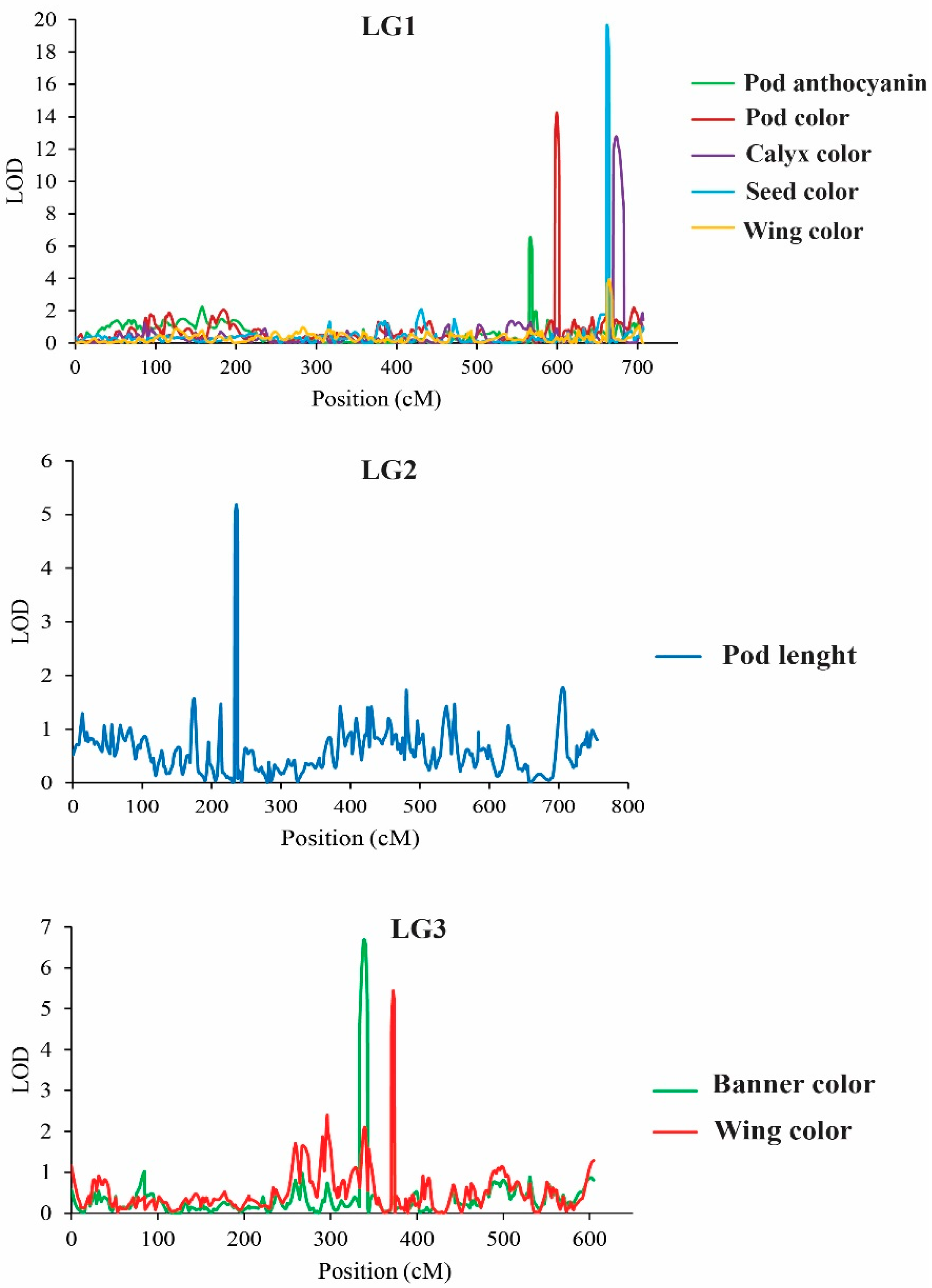
2.4. QTL Analysis for Flower-, Pod-, and Seed-Related Traits
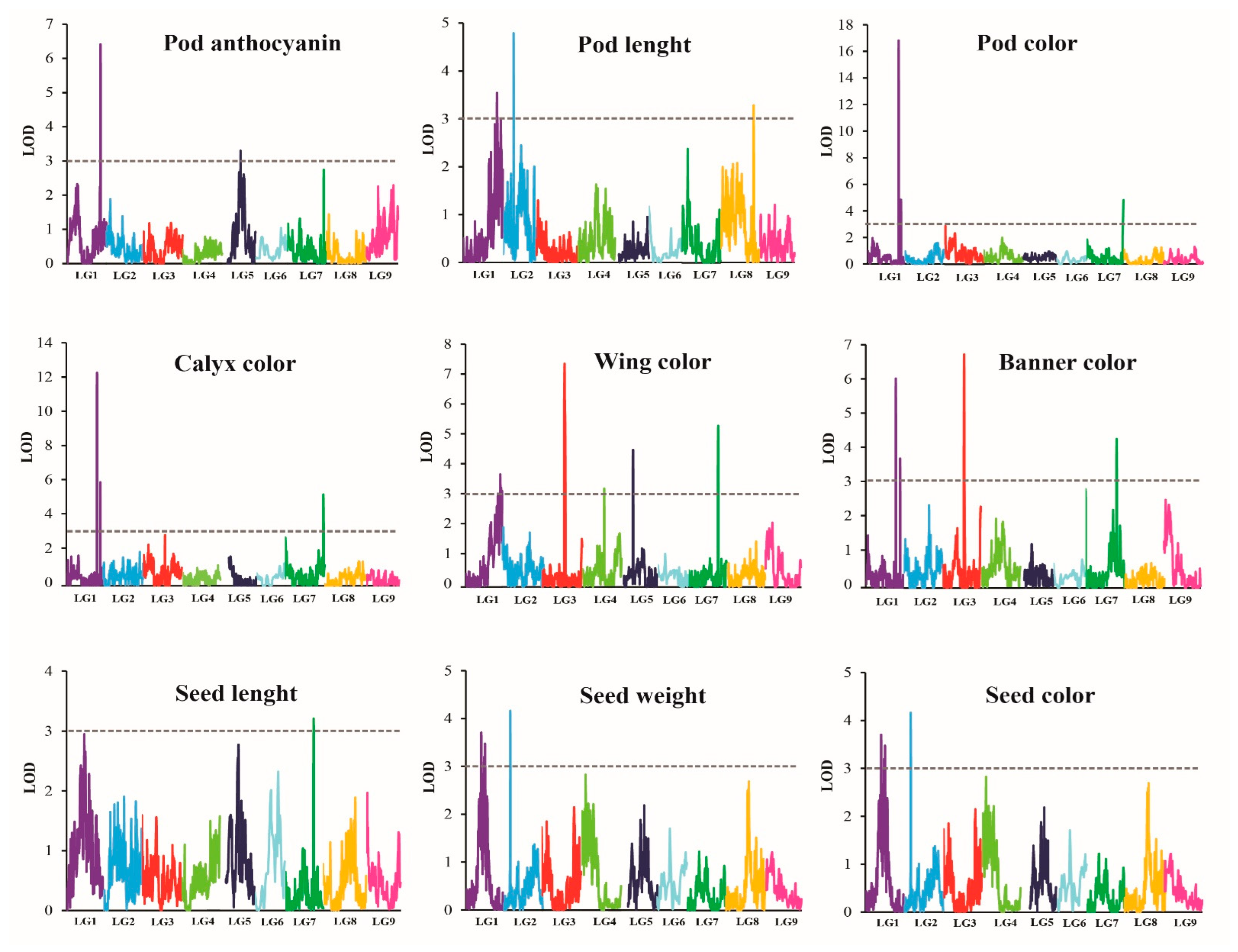
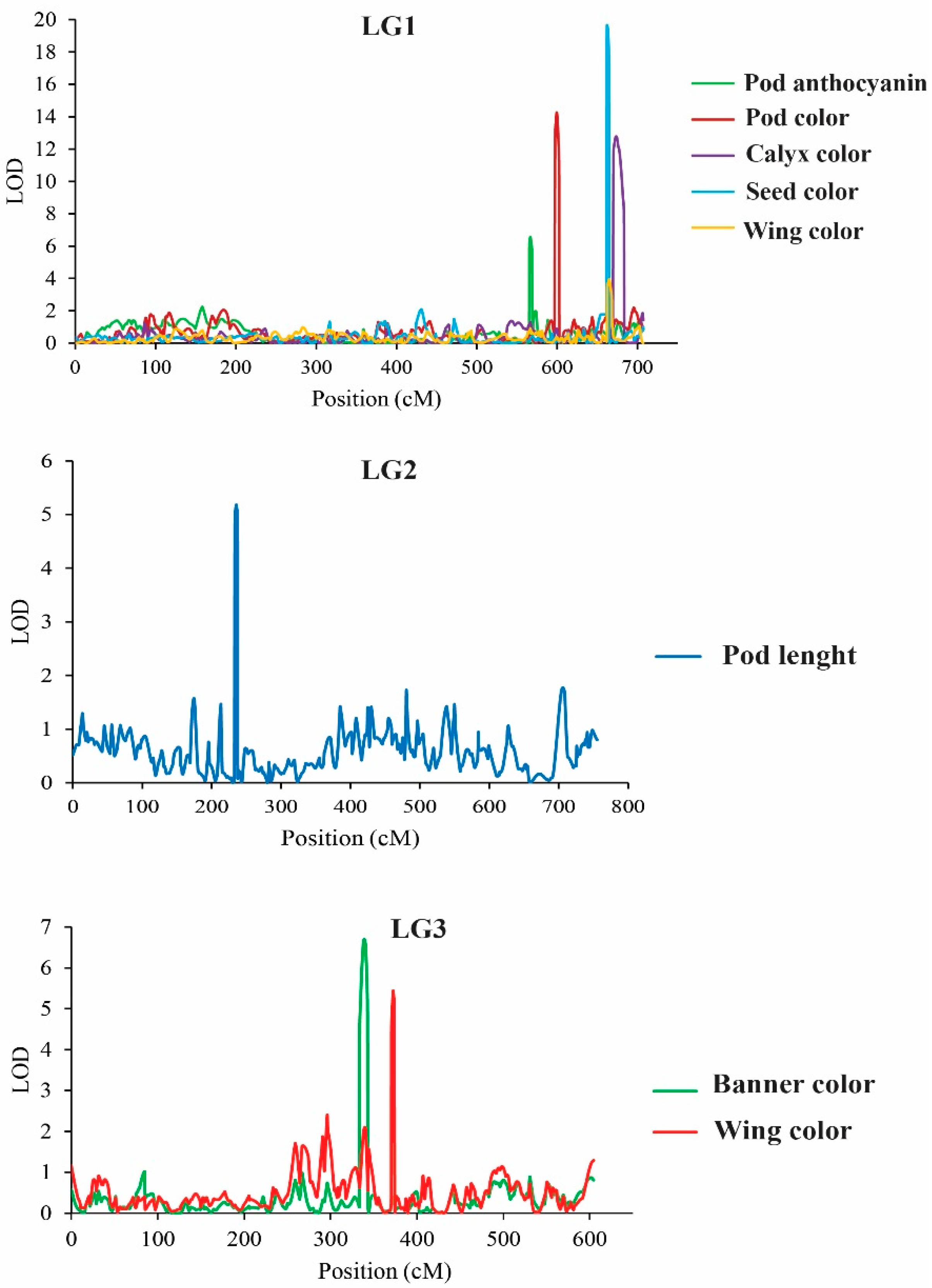
2.4.1. QTLs Controlling Organ Coloration
2.4.2. QTLs Controlling Organ Size
3. Discussion
3.1. Genetics Controlling Traits in Winged Bean
3.2. The Winged Bean Genetic Linkage Map
3.3. QTL Analysis for Flower-, Pod-, and Seed-Related Traits of the Winged Bean
4. Materials and Methods
4.1. Plant Materials
4.2. Field Evaluation
- TAcy = total anthocyanin (mg/100 g)
- OD = Absorbance 530 nm
- DV = Volume of diluted solution (mL)
- SV = Volume of extracted solution for diluted (mL)
- TEV = Total extracted volume (mL)
- SW = Sample weight (g)
- Ecf = Extinction coefficient (Ecf of cyaniding-3-glucoside is 449.2)
4.3. RAD-Seq
4.4. Linkage Map Construction
4.5. QTL Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Harder, D.K. Chromosome Counts in Psophocarpus. Kew Bull. 1992, 47, 529–534. [Google Scholar] [CrossRef]
- Peyachoknagul, S.; Matsui, T.; Shibata, H.; Hara, S.; Ikenaka, T.; Okada, Y.; Ohno, T. Sequence and expression of the mRNA encoding the chymotrypsin inhibitor in winged bean (Psophocarpus tetragonolobus (L.) DC.). Plant Mol. Biol. 1989, 12, 51–58. [Google Scholar] [CrossRef] [PubMed]
- Wilson, E. The Diversity of Life; The Belknap Press of Harvard University Press: Cambridge, UK, 1992. [Google Scholar]
- Khan, T.N. Papua New Guinea: A centre of genetic diversity in winged bean (Psophocarpus tetragonologus (L.) Dc.). Euphytica 1976, 25, 693–705. [Google Scholar] [CrossRef]
- Mohanty, C.S.; Verma, S.; Singh, V.; Khan, S.; Gaur, P.; Gupta, P.; Nizar, M.A.; Dikshit, N.; Pattanayak, R.; Shukla, A.; et al. Characterization of winged bean (Psophocarpus tetragonolobus (L.) DC.) based on molecular, chemical and physiological parameters. Am. J. Mol. Biol. 2013, 3, 187–197. [Google Scholar] [CrossRef] [Green Version]
- Adegboyega, T.T.; Abberton, M.T.; Abdelgadir, A.A.H.; Dianda, M.; Maziya-Dixon, B.; Oyatomi, O.A.; Ofodile, S.; Babalola, O.O. Nutrient and Antinutrient Composition of Winged Bean (Psophocarpus tetragonolobus (L.) DC.) Seeds and Tubers. J. Food Qual. 2019, 2019, 3075208. [Google Scholar] [CrossRef] [Green Version]
- NAS. The Winged Bean: High-Protein Crop for the Humid Tropics, 2nd ed.; National Academy Press: Washington, DC, USA, 1981; p. 41. [Google Scholar]
- Chen, D.; Yi, X.; Yang, H.; Zhou, H.; Yu, Y.; Tian, Y.; Lu, X. Genetic diversity evaluation of winged bean (Psophocarpus tetragonolobus (L.) DC.) using inter-simple sequence repeat (ISSR). Genet. Resour. Crop Evol. 2015, 62, 823–828. [Google Scholar] [CrossRef]
- Yang, S.; Grall, A.; Chapman, M.A. Origin and diversification of winged bean (Psophocarpus tetragonolobus (L.) DC.), a multipurpose underutilized legume. Am. J. Bot. 2018, 105, 888–897. [Google Scholar] [CrossRef]
- Laosatit, K.; Amkul, K.; Chankaew, S.; Somta, P. Molecular Genetic Diversity of Winged Bean Gene Pool in Thailand Assessed by SSR Markers. Hortic. Plant J. 2021, 8, 81–88. [Google Scholar] [CrossRef]
- Vatanparast, M.; Shetty, P.; Chopra, R.; Doyle, J.J.; Sathyanarayana, N.; Egan, A.N. Transcriptome sequencing and marker development in winged bean (Psophocarpus tetragonolobus; Leguminosae). Sci. Rep. 2016, 6, 29070. [Google Scholar] [CrossRef] [Green Version]
- Wong, Q.N.; Tanzi, A.S.; Ho, W.K.; Malla, S.; Blythe, M.; Karunaratne, A.; Massawe, F.; Mayes, S. Development of Gene-Based SSR Markers in Winged Bean (Psophocarpus tetragonolobus (L.) DC.) for Diversity Assessment. Genes 2017, 8, 100. [Google Scholar] [CrossRef] [Green Version]
- Sriwichai, S.; Monkham, T.; Sanitchon, J.; Jogloy, S.; Chankaew, S. Dual-Purpose of the Winged Bean (Psophocarpus tetragonolobus (L.) DC.), the Neglected Tropical Legume, Based on Pod and Tuber Yields. Plants 2021, 10, 1746. [Google Scholar] [CrossRef] [PubMed]
- Eagleton, G. Review: Winged bean (Psophocarpus tetragonolobus) cropping systems. Biodiversitas J. Biol. Divers. 2020, 21, 5927–5946. [Google Scholar] [CrossRef]
- Mahto, C.S.; Dua, R.P. Genetic Divergence for Yield Contributing Traits in Winged Bean. Indian J. Plant Genet. Resour. 2009, 22, 239–242. [Google Scholar]
- Peterson, B.K.; Weber, J.N.; Kay, E.H.; Fisher, H.S.; Hoekstra, H.E. Double Digest RADseq: An Inexpensive Method for De Novo SNP Discovery and Genotyping in Model and Non-Model Species. PLoS ONE 2012, 7, e37135. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kang, Y.J.; Kim, S.K.; Kim, M.Y.; Lestari, P.; Kim, K.H.; Ha, B.-K.; Jun, T.H.; Hwang, W.J.; Lee, T.; Lee, J.; et al. Genome sequence of mungbean and insights into evolution within Vigna species. Nat. Commun. 2014, 5, 5443. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lonardi, S.; Muñoz-Amatriaín, M.; Liang, Q.; Shu, S.; Wanamaker, S.I.; Lo, S.; Tanskanen, J.; Schulman, A.H.; Zhu, T.; Luo, M.-C.; et al. The genome of cowpea (Vigna unguiculata [L.] Walp.). Plant J. 2019, 98, 767–782. [Google Scholar] [CrossRef] [Green Version]
- Singh, A.K.; Velmurugan, A.; Gupta, D.S.; Kumar, J.; Kesari, R.; Konda, A.; Singh, N.P.; Roy, S.D.; Biswas, U.; Kumar, R.R.; et al. Draft genome sequence of a less-known wild Vigna: Beach pea (V. marina cv. ANBp-14-03). Crop J. 2019, 7, 660–666. [Google Scholar] [CrossRef]
- Sakai, H.; Naito, K.; Takahashi, Y.; Sato, T.; Yamamoto, T.; Muto, I.; Itoh, T.; Tomooka, N. The Vigna Genome Server, ‘VigGS’: A genomic knowledge base of the genus Vigna based on high-quality, annotated genome sequence of the azuki bean, Vigna angularis (Willd) Ohwi and Ohashi. Plant Cell Physiol. 2016, 57, e2. [Google Scholar] [CrossRef] [Green Version]
- Kreplak, J.; Madoui, M.-A.; Cápal, P.; Novák, P.; Labadie, K.; Aubert, G.; Bayer, P.E.; Gali, K.K.; Syme, R.A.; Main, D.; et al. A reference genome for pea provides insight into legume genome evolution. Nat. Genet. 2019, 51, 1411–1422. [Google Scholar] [CrossRef]
- Varshney, R.K.; Chen, W.; Li, Y.; Bharti, A.K.; Saxena, R.K.; Schlueter, J.A.; Donoghue, M.T.; Azam, S.; Fan, G.; Whaley, A.M.; et al. Draft genome sequence of pigeonpea (Cajanus cajan), an orphan legume crop of resource-poor farmers. Nat. Biotechnol. 2012, 30, 83–89. [Google Scholar] [CrossRef] [Green Version]
- Varshney, R.K.; Song, C.; Saxena, R.K.; Azam, S.; Yu, S.; Sharpe, A.G.; Cannon, S.; Baek, J.; Rosen, B.D.; Tar’An, B.; et al. Draft genome sequence of chickpea (Cicer arietinum) provides a resource for trait improvement. Nat. Biotechnol. 2013, 31, 240–246. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, Q.; Li, H.; Hong, Y.; Zhang, G.; Wen, S.; Li, X.; Zhou, G.; Li, S.; Liu, H.; Liu, H.; et al. Genome Sequencing and Analysis of the Peanut B-Genome Progenitor (Arachis ipaensis). Front. Plant Sci. 2018, 9, 604. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kajiya-Kanegae, H.; Nagasaki, H.; Kaga, A.; Hirano, K.; Ogiso-Tanaka, E.; Matsuoka, M.; Ishimori, M.; Ishimoto, M.; Hashiguchi, M.; Tanaka, H.; et al. Whole-genome sequence diversity and association analysis of 198 soybean accessions in mini-core collections. DNA Res. 2021, 28, dsaa032. [Google Scholar] [CrossRef] [PubMed]
- Hyten, D.L.; Cannon, S.B.; Song, Q.; Weeks, N.; Fickus, E.W.; Shoemaker, R.C.; Specht, J.E.; Farmer, A.D.; May, G.D.; Cregan, P.B. High-throughput SNP discovery through deep resequencing of a reduced representation library to anchor and orient scaffolds in the soybean whole genome sequence. BMC Genom. 2010, 11, 38. [Google Scholar] [CrossRef]
- Parchman, T.L.; Gompert, Z.; Mudge, J.; Schilkey, F.D.; Benkman, C.W.; Buerkle, C.A. Genome-wide association genetics of an adaptive trait in lodgepole pine. Mol. Ecol. 2012, 21, 2991–3005. [Google Scholar] [CrossRef]
- Baird, N.A.; Etter, P.D.; Atwood, T.S.; Currey, M.C.; Shiver, A.L.; Lewis, Z.A.; Selker, E.U.; Cresko, W.A.; Johnson, E.A. Rapid SNP Discovery and Genetic Mapping Using Sequenced RAD Markers. PLoS ONE 2008, 3, e3376. [Google Scholar] [CrossRef]
- Erskine, W.; Khan, T.N. Inheritance of pigmentation and pod shape in winged bean. Euphytica 1977, 26, 829–831. [Google Scholar] [CrossRef]
- Freyre, R.; Uzdevenes, C.; Gu, L.; Quesenberry, K.H. Genetics and Anthocyanin Analysis of Flower Color in Mexican Petunia. J. Am. Soc. Hortic. Sci. 2015, 140, 45–49. [Google Scholar] [CrossRef]
- Dekkers, J.C.M.; Hospital, F. The use of molecular genetics in the improvement of agricultural populations. Nat. Rev. Genet. 2002, 3, 22–32. [Google Scholar] [CrossRef]
- Miller, M.R.; Dunham, J.P.; Amores, A.; Cresko, W.A.; Johnson, E.A. Rapid and cost-effective polymorphism identification and genotyping using restriction site associated DNA (RAD) markers. Genome Res. 2006, 17, 240–248. [Google Scholar] [CrossRef] [Green Version]
- Hegarty, M.; Yadav, R.; Lee, M.; Armstead, I.; Sanderson, R.; Scollan, N.; Powell, W.; Skøt, L. Genotyping by RAD sequencing enables mapping of fatty acid composition traits in perennial ryegrass (Lolium perenne (L.)). Plant Biotechnol. J. 2013, 11, 572–581. [Google Scholar] [CrossRef] [PubMed]
- Pootakham, W.; Ruang-Areerate, P.; Jomchai, N.; Sonthirod, C.; Sangsrakru, D.; Yoocha, T.; Theerawattanasuk, K.; Nirapathpongporn, K.; Romruensukharom, P.; Tragoonrung, S.; et al. Construction of a high-density integrated genetic linkage map of rubber tree (Hevea brasiliensis) using genotyping-by-sequencing (GBS). Front. Plant Sci. 2015, 6, 367. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, Y.; Zhao, Y.; Guo, Y.; Su, K.; Shi, X.; Liu, D.; Zhang, J. High-density genetic linkage-map construction of hawthorn and QTL mapping for important fruit traits. PLoS ONE 2020, 15, e0229020. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khoo, H.E.; Azlan, A.; Tang, S.T.; Lim, S.M. Anthocyanidins and anthocyanins: Colored pigments as food, pharmaceutical ingredients, and the potential health benefits. Food Nutr. Res. 2017, 61, 1361779. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wallace, T.C.; Giusti, M.M. Anthocyanins—Nature’s Bold, Beautiful, and Health-Promoting Colors. Foods 2019, 8, 550. [Google Scholar] [CrossRef] [Green Version]
- Tanaka, Y.; Tsuda, S.; Kusumi, T. Metabolic Engineering to Modify Flower Color. Plant Cell Physiol. 1998, 39, 1119–1126. [Google Scholar] [CrossRef]
- Katsumoto, Y.; Fukuchi-Mizutani, M.; Fukui, Y.; Brugliera, F.; Holton, T.; Karan, M.; Nakamura, N.; Yonekura-Sakakibara, K.; Togami, J.; Pigeaire, A.; et al. Engineering of the Rose Flavonoid Biosynthetic Pathway Successfully Generated Blue-Hued Flowers Accumulating Delphinidin. Plant Cell Physiol. 2007, 48, 1589–1600. [Google Scholar] [CrossRef]
- Crow, J.F.; Crow, J.F. Dominance and Overdominance. In The Genetics and Exploitation of Heterosis in Crops; Coors, J.G., Pandey, S., Eds.; The American Society of Agronomy, Inc.: Madison, WI, USA; Crop Science Society of America, Inc.: Madison, WI, USA, 1999; pp. 49–58. [Google Scholar] [CrossRef]
- IBPGR. International Board for Plant Genetic Resources. Revised Winged Bean Descriptors; IBPGR: Rome, Italy, 1982; 24p. [Google Scholar]
- Ryu, S.N.; Park, S.Z.; Kang, S.S.; Han, S.J. Determination of C3G content in black purple rice using HPLC and UV-Vis spec-trophotometer. Korean J. Crop. Sci. 2003, 48, 369–371. [Google Scholar]
- Guisti, M.M.; Wrolstad, R.E. Characterization and Measurement of Anthocyanins by UV-Visible Spectroscopy. Current Protocols in Food Analytical Chemistry; John Wiley and Sons: Hoboken, NJ, USA, 2001; pp. F1.2.1–F1.2.13. [Google Scholar]
- Lodhi, M.A.; Ye, G.-N.; Weeden, N.F.; Reisch, B.I. A simple and efficient method for DNA extraction from grapevine cultivars andVitis species. Plant Mol. Biol. Rep. 1994, 12, 6–13. [Google Scholar] [CrossRef]
- Catchen, J.M.; Amores, A.; Hohenlohe, P.; Cresko, W.; Postlethwait, J.H. Stacks: Building and Genotyping Loci De Novo From Short-Read Sequences. G3 Genes Genomes Genet. 2011, 1, 171–182. [Google Scholar] [CrossRef] [Green Version]
- Meng, L.; Li, H.; Zhang, L.; Wang, J. QTL IciMapping: Integrated software for genetic linkage map construction and quantitative trait locus mapping in biparental populations. Crop. J. 2015, 3, 269–283. [Google Scholar] [CrossRef] [Green Version]
- Van Os, H.; Stam, P.; Visser, R.G.F.; Van Eck, H.J. RECORD: A novel method for ordering loci on a genetic linkage map. Theor. Appl. Genet. 2005, 112, 30–40. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Li, H.; Meng, L.; Wang, J. Ordering of high-density markers by the k-Optimal algorithm for the traveling-salesman problem. Crop. J. 2020, 8, 701–712. [Google Scholar] [CrossRef]
- Kosambi, D.D. The Estimation of Map Distances From Recombination Values. Ann. Eugen. 1943, 12, 172–175. [Google Scholar] [CrossRef]
- Li, H.; Ribaut, J.-M.; Li, Z.; Wang, J. Inclusive composite interval mapping (ICIM) for digenic epistasis of quantitative traits in biparental populations. Theor. Appl. Genet. 2007, 116, 243–260. [Google Scholar] [CrossRef] [PubMed]
- Van Ooijen, J. MAPQTL5.0, Software for the Mapping of Quantitative Trait Loci in Experimental Population; Kyazma, B.V., Ed.; Plant Research International: Wageningen, The Netherlands, 2004. [Google Scholar]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Traits | AnthoCy | PDL | PDC | CalyxC | WingC | BannerC | SeedL | SeedW |
|---|---|---|---|---|---|---|---|---|
| PDL | 0.248 * | |||||||
| PDC | 0.748 ** | 0.329 ** | ||||||
| CalyxC | 0.706 ** | 0.262 * | 0.812 ** | |||||
| WingC | 0.431 ** | 0.091 | 0.421 ** | 0.543 ** | ||||
| BannerC | 0.419 ** | 0.081 | 0.405 ** | 0.523 ** | 0.978 ** | |||
| SeedL | 0.158 | 0.151 | 0.110 | 0.095 | 0.082 | 0.114 | ||
| SeedW | −0.034 | −0.008 | 0.010 | −0.027 | −0.110 | −0.096 | 0.500 ** | |
| SeedC | 0.381 ** | 0.239 * | 0.547 ** | 0.595 ** | 0.400 ** | 0.378 ** | −0.018 | −0.197 |
| Linkage Group | Number of SNP Markers | Length (cM) | Average Marker Interval (cM) | Maximum Interval (cM) |
|---|---|---|---|---|
| 1 | 208 | 707.12 | 3.40 | 11.09 |
| 2 | 218 | 755.74 | 3.47 | 11.98 |
| 3 | 186 | 604.61 | 3.25 | 13.37 |
| 4 | 170 | 507.73 | 2.98 | 11.53 |
| 5 | 96 | 293.99 | 3.06 | 9.55 |
| 6 | 36 | 164.96 | 4.58 | 16.44 |
| 7 | 176 | 560.28 | 3.18 | 10.67 |
| 8 | 180 | 587.78 | 3.27 | 16.90 |
| 9 | 114 | 370.59 | 3.25 | 15.28 |
| Average | 153.78 | 505.87 | 3.38 | - |
| Total | 1384 | 4552.8 | - | - |
| Traits | QTLs | LGs | Positions | Marker Interval | LOD Score | PVE (%) | Add | Dom |
|---|---|---|---|---|---|---|---|---|
| Anthocyanin content | qAntho1.1 | 1 | 599.2 | 90362_173–86339_105 | 6.41 | 28.92 | 4.41 | 1.47 |
| qAntho5.1 | 5 | 137.4 | 129210_17–74111_70 | 3.29 | 11.61 | −2.16 | −2.81 | |
| Pod length | qPpl1.1 | 1 | 601.3 | 86339_105–88581_215 | 3.54 | 11.08 | 0.86 | 1.18 |
| qPdl1.1 | 1 | 609.8 | 88581_7–113679_228 | 3.03 | 9.49 | 0.88 | 1.04 | |
| qPdl2.1 | 2 | 236.6 | 65543_37–65543_66 | 4.79 | 16.17 | 0.44 | 1.79 | |
| qPdl8.1 | 8 | 503.0 | 758788_184–45825_50 | 3.39 | 11.22 | 1.39 | 0.01 | |
| Pod color | qPdc1.1 | 1 | 599.4 | 86339_105–88581_215 | 16.82 | 49.15 | 2.22 | 0.15 |
| qPdc1.2 | 1 | 643.9 | 50448_148–73516_244 | 4.84 | 10.35 | 0.06 | 1.56 | |
| qPdc7.1 | 7 | 560.2 | 6763_192–141967_88 | 4.81 | 9.69 | 1.02 | −0.47 | |
| Calyx color | qClxc1.1 | 1 | 599.4 | 86339_105–88581_215 | 12.26 | 38.28 | 0.76 | 0.05 |
| qClxc1.2 | 1 | 664.9 | 5563_75–37621_162 | 5.84 | 15.01 | 0.08 | 0.74 | |
| qClxc7.1 | 7 | 543.1 | 1977_174–6763_192 | 5.12 | 12.67 | 0.44 | −0.23 | |
| Wing color | qWingc1.1 | 1 | 569.0 | 49233_195–144410_63 | 6.01 | 16.33 | 0.76 | 0.03 |
| qWingc1.2 | 1 | 649.8 | 73516_104–64897_31 | 3.67 | 9.72 | −0.05 | 0.85 | |
| qWingc3.1 | 3 | 340.7 | 59095_37–59095_183 | 6.72 | 18.61 | −0.93 | 0.05 | |
| qWingc7.1 | 7 | 0.0 | 92184_83–54946_220 | 3.17 | 7.77 | −0.48 | 0.29 | |
| qWingc7.1 | 7 | 435.3 | 79824_126–57556_229 | 4.85 | 12.77 | 0.79 | −0.02 | |
| Banner color | qBannerc1.1 | 1 | 670.1 | 37621_240–101902_163 | 3.65 | 6.45 | 0.32 | 0.66 |
| qBannerc1.2 | 1 | 706.5 | 71361_136–71361_18 | 3.09 | 5.54 | 0.48 | 0.52 | |
| qBannerc3.1 | 3 | 340.1 | 120628_171–59095_37 | 7.34 | 17.75 | −1.02 | 0.10 | |
| qBannerc3.2 | 3 | 345.1 | 59095_229–542971_120 | 6.12 | 14.84 | 0.02 | 1.21 | |
| qBannerc4.1 | 4 | 283.3 | 29428_113–114955_133 | 3.17 | 6.66 | −0.18 | 0.79 | |
| qBannerc5.1 | 5 | 81.4 | 42289_151–31093_166 | 4.46 | 9.57 | −0.24 | 0.99 | |
| qBannerc7.1 | 7 | 435.3 | 79824_126–57556_229 | 5.27 | 11.51 | 0.85 | 0.32 | |
| Seed length | qSdl7.1 | 7 | 414.7 | 143811_217–2570_116 | 3.21 | 18.83 | 0.48 | 0.12 |
| Seed width | qSdw1.1 | 1 | 294.8 | 79225_198–130456_96 | 3.70 | 14.43 | 0.27 | 0.29 |
| qSdw1.2 | 1 | 347.9 | 88187_3–100448_14 | 3.19 | 11.47 | 0.23 | 0.29 | |
| qSdw1.3 | 1 | 367.2 | 46319_242–77017_78 | 3.47 | 12.41 | 0.20 | 0.30 | |
| qSdw2.1 | 2 | 134.6 | 10683_187–3208_238 | 4.16 | 18.52 | −0.09 | 0.51 | |
| Seed color | qSdc1.1 | 1 | 660.9 | 64897_73–73327_179 | 22.74 | 73.33 | 0.78 | 0.89 |
| qSdc3.1 | 3 | 277.3 | 74742_203–41532_216 | 3.27 | 5.45 | −0.30 | 0.01 |
| Traits | QTLs | LGs | Positions | Marker Interval | LOD Score | PVE (%) | Add | Dom |
|---|---|---|---|---|---|---|---|---|
| Anthocyanin | qAntho1.1 | 1 | 566.5 | 59413_160–49233_195 | 6.57 | 29.60 | 4.57 | 2.15 |
| Pod length | qPpl2.1 | 2 | 235.5 | 69533_212–65543_37 | 5.18 | 24.20 | 0.51 | 2.03 |
| Pod color | qPdc1.1 | 1 | 599.4 | 86339_105–88581_215 | 14.26 | 53.40 | 2.15 | 1.39 |
| Calyx color | qClxc1.1 | 1 | 673.03 | 37621_240–101902_163 | 12.78 | 49.60 | 0.82 | 0.83 |
| Wing color | qWingc1.1 | 1 | 664.83 | 73327_66–5563_75 | 3.94 | 19.00 | 0.72 | 0.68 |
| qWingc3.1 | 3 | 372.46 | 52207_209–15865_163 | 5.44 | 25.30 | −0.76 | 0.96 | |
| Banner color | qBannerc3.1 | 3 | 338.69 | 120628_171–59095_37 | 6.69 | 30.10 | −0.94 | 1.14 |
| Seed length | No QTL detected | |||||||
| Seed width | No QTL detected | |||||||
| Seed color | qSdc1.1 | 1 | 662.34 | 73327_179–73327_66 | 19.65 | 70.60 | 0.78 | 0.87 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chankaew, S.; Sriwichai, S.; Rakvong, T.; Monkham, T.; Sanitchon, J.; Tangphatsornruang, S.; Kongkachana, W.; Sonthirod, C.; Pootakham, W.; Amkul, K.; et al. The First Genetic Linkage Map of Winged Bean [Psophocarpus tetragonolobus (L.) DC.] and QTL Mapping for Flower-, Pod-, and Seed-Related Traits. Plants 2022, 11, 500. https://doi.org/10.3390/plants11040500
Chankaew S, Sriwichai S, Rakvong T, Monkham T, Sanitchon J, Tangphatsornruang S, Kongkachana W, Sonthirod C, Pootakham W, Amkul K, et al. The First Genetic Linkage Map of Winged Bean [Psophocarpus tetragonolobus (L.) DC.] and QTL Mapping for Flower-, Pod-, and Seed-Related Traits. Plants. 2022; 11(4):500. https://doi.org/10.3390/plants11040500
Chicago/Turabian StyleChankaew, Sompong, Sasiprapa Sriwichai, Teppratan Rakvong, Tidarat Monkham, Jirawat Sanitchon, Sithichoke Tangphatsornruang, Wasitthee Kongkachana, Chutima Sonthirod, Wirulda Pootakham, Kitiya Amkul, and et al. 2022. "The First Genetic Linkage Map of Winged Bean [Psophocarpus tetragonolobus (L.) DC.] and QTL Mapping for Flower-, Pod-, and Seed-Related Traits" Plants 11, no. 4: 500. https://doi.org/10.3390/plants11040500
APA StyleChankaew, S., Sriwichai, S., Rakvong, T., Monkham, T., Sanitchon, J., Tangphatsornruang, S., Kongkachana, W., Sonthirod, C., Pootakham, W., Amkul, K., Kaewwongwal, A., Laosatit, K., & Somta, P. (2022). The First Genetic Linkage Map of Winged Bean [Psophocarpus tetragonolobus (L.) DC.] and QTL Mapping for Flower-, Pod-, and Seed-Related Traits. Plants, 11(4), 500. https://doi.org/10.3390/plants11040500