
A Review of the Developmental Processes and Selective Pressures Shaping Aperture Pattern in Angiosperms


 , , and
, , and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Aperture Formation
2.1. Cellular Components Correlated with Aperture Location
2.2. Determination of Aperture Localization
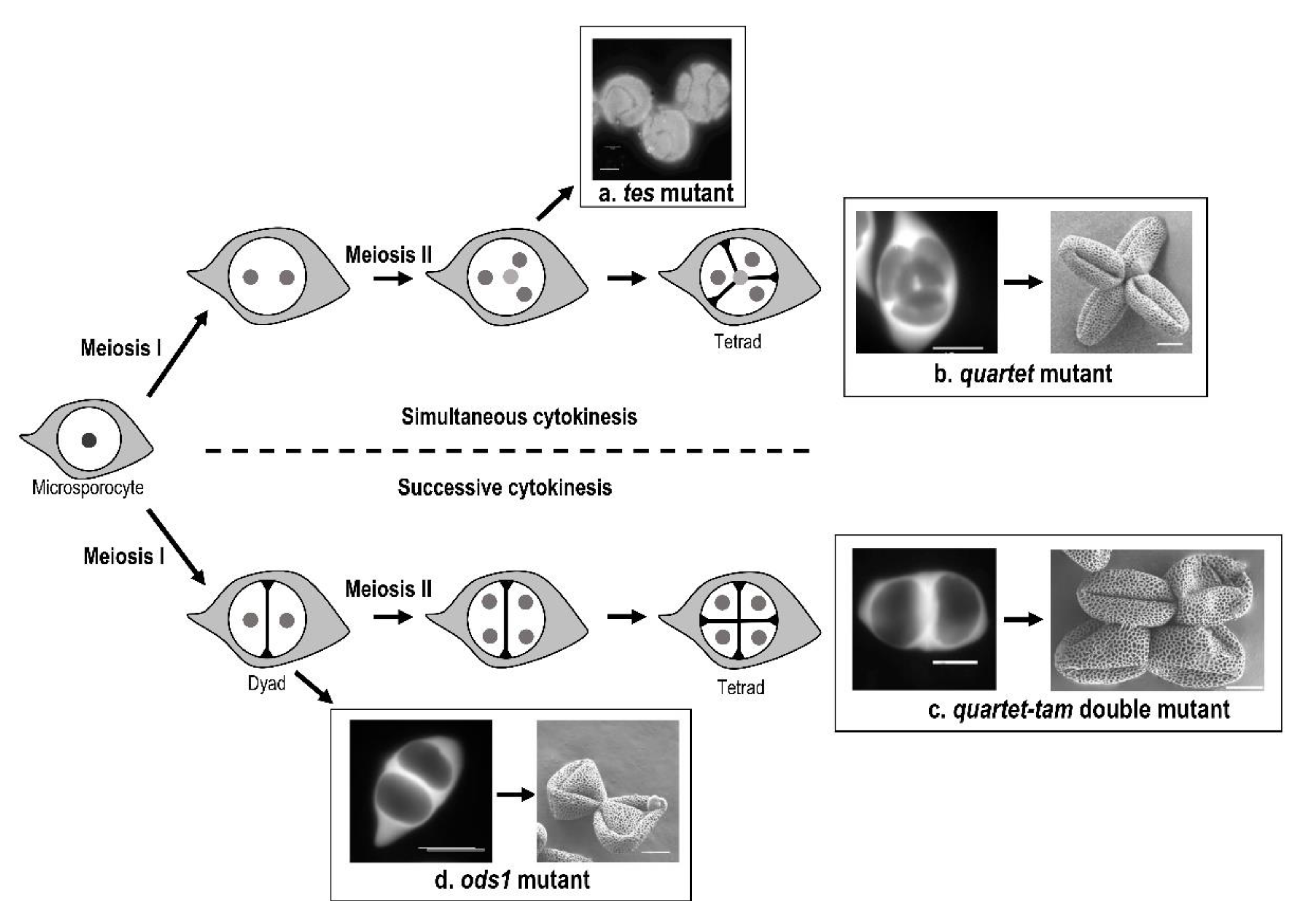
2.2.1. Role of Cytokinesis in Aperture Pattern Determination
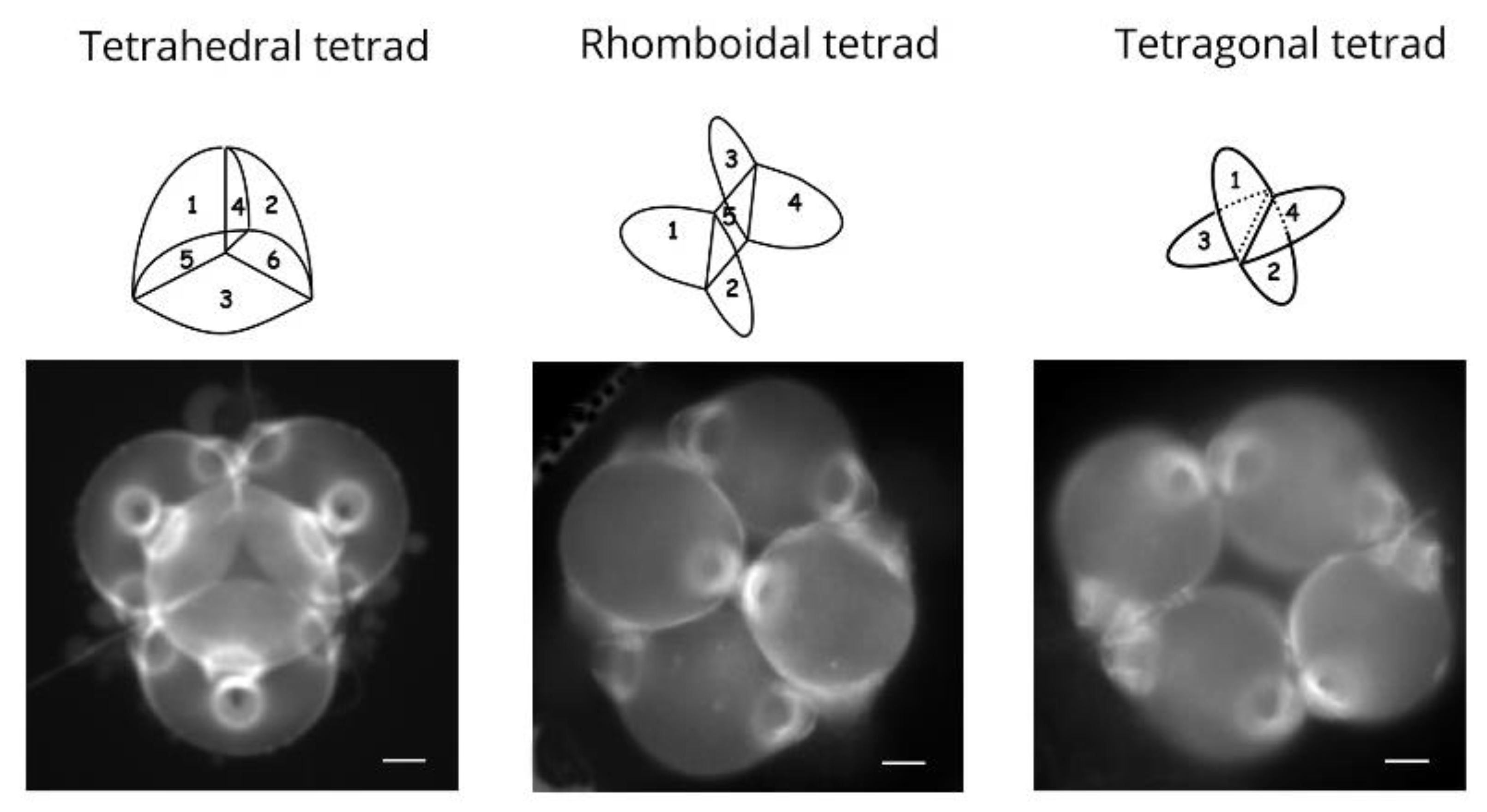
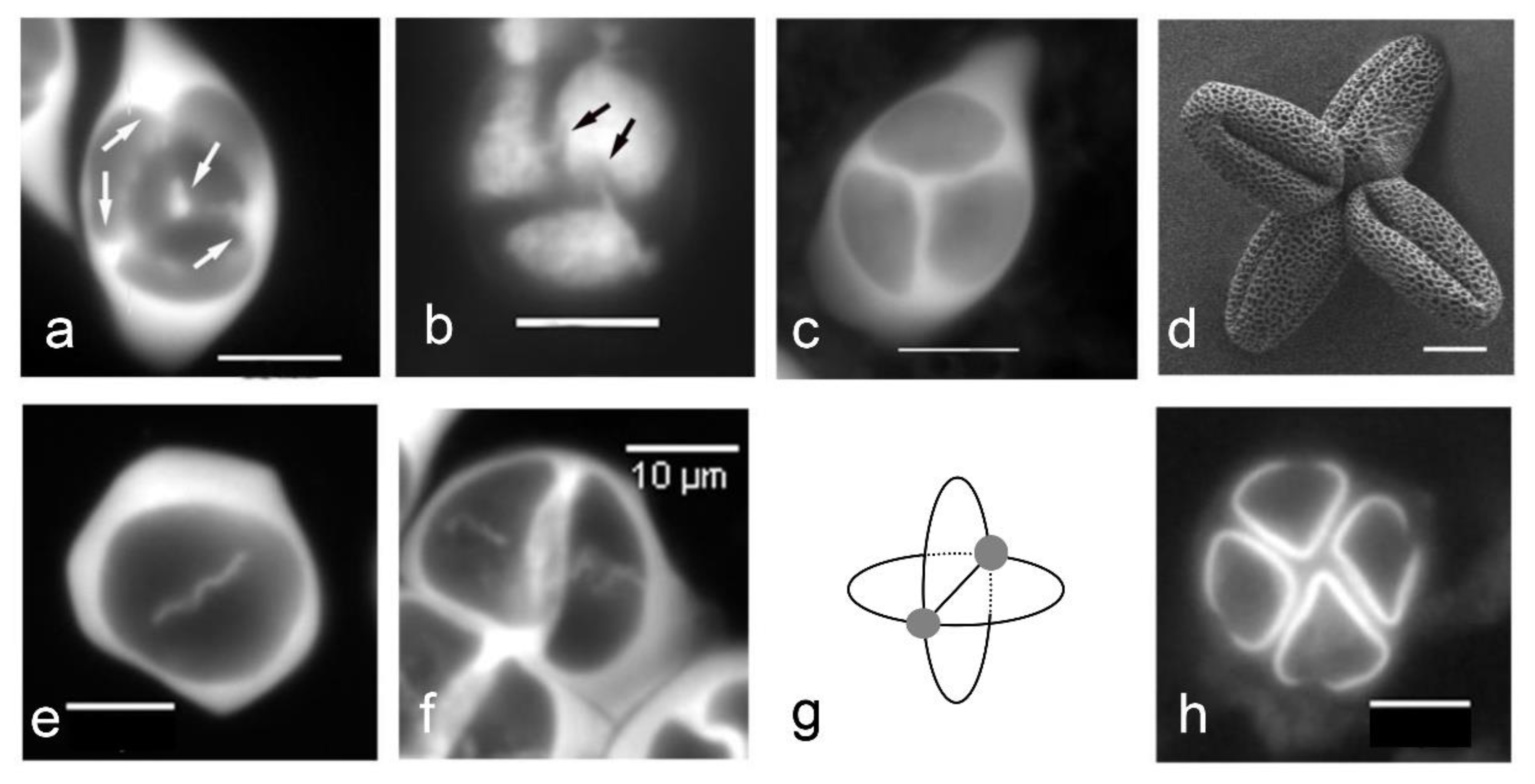
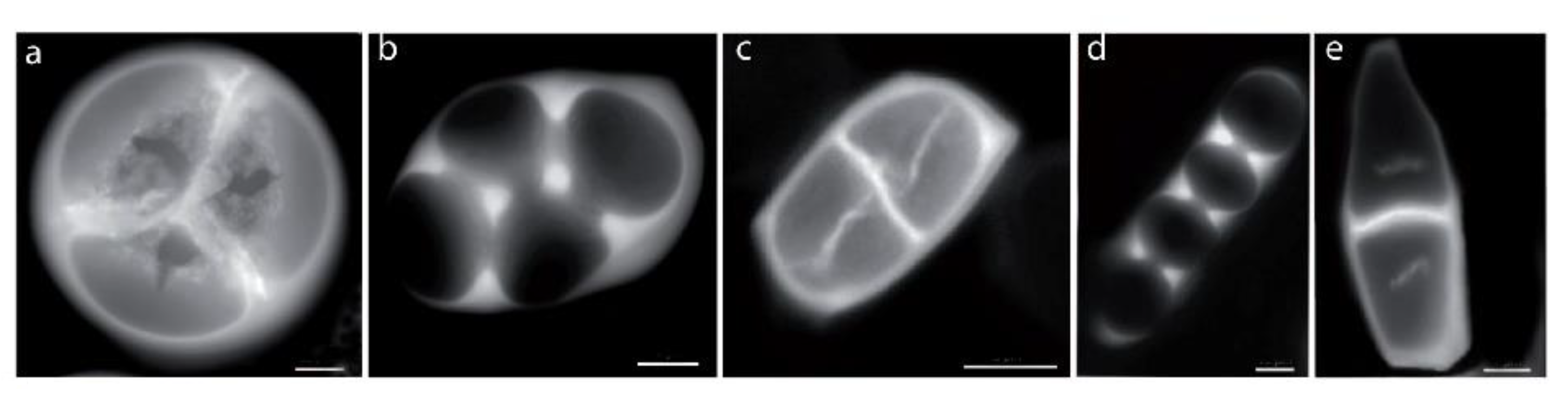
2.2.2. Role of Tetrad Form in Aperture Pattern Determination
2.2.3. The Role of Polyploidy or Hybridization in Aperture Patterns
2.2.4. Role of Callose Cleavage Wall Formation in Aperture Pattern Determination
3. Selection and Developmental Constraints Involved in the Evolution of Pollen Aperture Pattern
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Heslop-Harrison, J. The Pollen Wall: Structure and Development; Butterworth: London, UK, 1971. [Google Scholar]
- Edlund, A.F.; Swanson, R.; Preuss, D. Pollen and stigma structure and function: The role of diversity in pollination. Plant Cell 2004, 16, S84–S97. [Google Scholar] [CrossRef] [PubMed]
- Erdtman, G. Pollen Morphology and Plant Taxonomy: Angiosperms (An Introduction to Palynology); Almqvist & Wiksell: Stockholm, Sweden, 1952. [Google Scholar]
- PalDat. 2021. Available online: http://www.paldat.org/ (accessed on 22 December 2021).
- Mignot, A.; Hoss, C.; Dajoz, I.; Leuret, C.; Henry, J.; Dreuillaux, J.M.; Heberle-Bors, E.; Till-Bottraud, I. Pollen aperture polymorphism in the angiosperms: Importance, possible causes and consequences. Acta Bot. Gall. 1994, 141, 109–122. [Google Scholar] [CrossRef]
- Walker, J.; Doyle, J. The bases of angiosperm phylogeny, palynology. Ann. Mo. Bot. Gard. 1975, 62, 664–723. [Google Scholar] [CrossRef]
- Godwin, H.; Echlin, P.; Chapman, B. The development of the pollen grain wall in Ipomoea purpurea (L.) Roth. Rev. Palaeobot. Palyno 1967, 3, 181–195. [Google Scholar] [CrossRef]
- Vijayaraghavan, M.R.; Shukla, A.K. Absence of callose around the microspore tetrad and poorly developed exine in Pergularia daemia. Ann. Bot. 1977, 41, 923–926. [Google Scholar] [CrossRef]
- PaxsonSowders, D.M.; Owen, H.A.; Makaroff, C.A. A comparative ultrastructural analysis of exine pattern development in wild-type Arabidopsis and a mutant defective in pattern formation. Protoplasma 1997, 198, 53–65. [Google Scholar] [CrossRef]
- Doores, A.S.; Osborn, J.M.; El-ghazaly, G. Pollen ontogeny in Ephedra americana (Gnetales). Int. J. Plant Sci. 2007, 168, 985–997. [Google Scholar] [CrossRef][Green Version]
- Dong, X.; Hong, Z.; Sivaramakrishnan, M.; Mahfouz, M.; Verma, D.P.S. Callose synthase (CalS5) is required for exine formation during microgametogenesis and for pollen viability in Arabidopsis. Plant J. 2005, 42, 315–328. [Google Scholar] [CrossRef]
- Enns, L.C.; Kanaoka, M.M.; Torii, K.U.; Comai, L.; Okada, K.; Cleland, R.E. Two callose synthases, GSL1 and GSL5, play an essential and redundant role in plant and pollen development and in fertility. Plant Mol. Biol. 2005, 58, 333–349. [Google Scholar] [CrossRef]
- Nishikawa, S.-I.; Zinkl, G.; Swanson, R.; Maruyama, D.; Preuss, D. Callose (b-1,3 glucan) is essential for Arabidopsis pollen wall patterning, but not tube growth. BMC Plant Biol. 2005, 5, 22. [Google Scholar] [CrossRef]
- Suzuki, T.; Masaoka, K.; Nishi, M.; Nakamura, K.; Ishiguro, S. Identification of kaonashi mutants showing abnormal pollen exine structure in Arabidopsis thaliana. Plant Cell Physiol. 2008, 49, 1465–1477. [Google Scholar] [CrossRef] [PubMed]
- Zavada, M.S.; Anderson, G.J. The wall and aperture development of pollen from dioecious Solanum appendiculatum: What is inaperturate pollen? Grana 1997, 36, 129–134. [Google Scholar] [CrossRef]
- Pozhidaev, A.E. Hypothetical way of pollen aperture patterning. A family-based study of Krameriaceae. Rev. Palaeobot. Palynol. 2003, 127, 1–23. [Google Scholar] [CrossRef]
- Ariizumi, T.; Toriyama, K. Genetic Regulation of Sporopollenin Synthesis and Pollen Exine Development. Ann. Rev. Plant Biol. 2011, 62, 437–460. [Google Scholar] [CrossRef] [PubMed]
- Heslop-Harrison, J. An ultrastructural study of pollen wall ontogeny in Silene pendula. Grana Palynol. 1963, 4, 7–24. [Google Scholar] [CrossRef][Green Version]
- Skvarla, J.; Larson, D. Fine structural studies of Zea mays pollen I: Cell membranes and exine ontogeny. Am. J. Bot. 1966, 53, 1112–1125. [Google Scholar] [CrossRef]
- Christesen, J.E.; Horner, H.T. Pollen pore development and its spatial orientation during microsporogenesis in the grass Sorghum bicolor. Am. J. Bot. 1974, 61, 604–623. [Google Scholar] [CrossRef]
- Dickinson, H. Ultrastructural aspects of primexine formation in the microspore tetrad of Lilium longiflorum. Cytobiologie 1970, 1, 437–449. [Google Scholar]
- Blackmore, S.; Barnes, S. Pollen wall morphogenesis in Tragopogon porrifolius L. (Compositae: Lactuceae) and its taxonomic significance. Rev. Palaeobot. Palynol. 1987, 52, 233–246. [Google Scholar] [CrossRef]
- Echlin, P.; Godwin, H. The ultrastructure and ontogeny of pollen in Helleborus foetidus L. J. Cell Sci. 1968, 3, 175–186. [Google Scholar] [CrossRef]
- Horner, H.T.; Pearson, C.B. Pollen wall and aperture development in Helianthus annuus (compositae: Heliantheae). Am. J. Bot. 1978, 65, 293–309. [Google Scholar] [CrossRef]
- Waterkeyn, L.; Bienfait, A. On a possible function of the callosic special wall in Ipomea purpurea (L.) Roth. Grana 1970, 10, 13–20. [Google Scholar] [CrossRef]
- Albert, B.; Nadot, S. Aperture ontogeny in the Proteaceae Grevillea rosmarinifolia. In Pollen: Structure, Types and Effects; Kaiser, B.J., Ed.; Nova Science Publishers: Hauppauge, NY, USA, 2010. [Google Scholar]
- Albert, B.; Matamoro-Vidal, A.; Raquin, C.; Nadot, S. Formation and function of a new pollen aperture pattern in angiosperms: The proximal sulcus of Tillandsia leiboldiana (Bromeliaceae). Am. J. Bot. 2010, 97, 365–368. [Google Scholar] [CrossRef] [PubMed]
- Ressayre, A.; Triky-Teurtroy, S.; Forchioni, A.; Dreyer, L.; Nadot, S. Post-meiotic cytokinesis and pollen aperture pattern ontogeny: Comparison of development in four species differing in aperture pattern. Am. J. Bot. 2005, 92, 576–583. [Google Scholar] [CrossRef]
- Ressayre, A. Equatorial aperture pattern in monocots: Same definition rules as in eudicots? The example of two species of Pontederiaceae. Int. J. Plant Sci. 2001, 162, 1219–1224. [Google Scholar] [CrossRef]
- Albert, B.; Ressayre, A.; Nadot, S. Correlation between pollen aperture pattern and callose deposition in late tetrad stage in three species producing atypical pollen grains. Am. J. Bot. 2011, 98, 189–196. [Google Scholar] [CrossRef]
- Toghranegar, Z.; Nadot, S.; Albert, B. Variation of microsporogenesis in monocots producing monosulcate pollen grains. Ann. Bot. 2013, 112, 135–139. [Google Scholar] [CrossRef]
- Rowley, J.R. Germinal apertural formation in pollen. Taxon 1975, 24, 12–25. [Google Scholar] [CrossRef]
- Larson, D.; Lewis, C. Pollen wall development in Parkinsonia aculeata. Grana 1962, 3, 21–31. [Google Scholar]
- Guzzo, F.; Baldan, B.; Bracco, F.; Mariani, P. Pollen development in Liriodendron tulipifera: Some unusual features. Can. J. Bot. 1994, 72, 352–358. [Google Scholar] [CrossRef]
- Flynn, J.; Rowley, J.R. The primexine of Nelumbo nucifera. Experientia 1971, 27, 227–228. [Google Scholar] [CrossRef]
- Kreunen, S.S.; Osborn, J.M. Pollen and anther development in Nelumbo (Nelumbonaceae). Am. J. Bot. 1999, 86, 1662–1676. [Google Scholar] [CrossRef] [PubMed]
- Dobritsa, A.; Coerper, D. The novel plant protein inaperturate pollen1 marks distinct cellular domains and controls formation of apertures in the Arabidopsis pollen exine. Plant Cell 2012, 24, 4452–4464. [Google Scholar] [CrossRef] [PubMed]
- Li, P.; Ben-Menni Schuler, S.; Reeder, S.H.; Wang, R.; Suárez Santiago, V.N.; Dobritsa, A.A. INP1 involvement in pollen aperture formation is evolutionarily conserved and may require species-specific partners. J. Exp. Bot. 2018, 69, 983–996. [Google Scholar] [CrossRef]
- Zhang, X.; Zhao, G.; Tan, Q.; Yuan, H.; Betts, N.; Zhu, L.; Zhang, D.; Liang, W. Rice pollen aperture formation is regulated by the interplay between OsINP1 and OsDAF1. Nat. Plant 2020, 6, 394–403. [Google Scholar] [CrossRef]
- Mazuecos-Aguilera, I.; Romero-Garcia, A.; Klodova, B.; Honys, D.; Fernandez-fernandez, M.; Ben-Menni Schuler, S.; Dobritsa, A.A.; Suárez-Santiago, V.N. The role of inaperturate pollen1 as a pollen aperture factor is conserved in the basal eudicot Eschscholzia californica (Papaveraceae). Front. Plant Sci. 2021, 12, 1–14. [Google Scholar] [CrossRef]
- Lee, B.H.; Wang, R.; Moberg, I.; Reeder, S.; Amom, P.; Tan, M.H.; Amstutz, K.; Chandna, P.; Helton, A.; Andrianova, E.P.; et al. A species-specific functional module controls formation of pollen apertures. Nat. Plant 2021, 7, 966–978. [Google Scholar] [CrossRef]
- Lee, B.H.; Weber, Z.T.; Zourelidou, M.; Hofmeister, B.T.; Schmitz, R.J.; Schwechheimer, C.; Dobritsa, A.A. Arabidopsis Protein Kinase D6PKL3 Is Involved in the Formation of Distinct Plasma Membrane Aperture Domains on the Pollen Surface. Plant Cell 2018, 30, 2038–2056. [Google Scholar] [CrossRef]
- Lin, S.; Dong, H.; Zhang, F.; Qiu, L.; Wang, F.; Cao, J.; Huang, L. BcMF8, a putative arabinogalactan protein-encoding gene, contributes to pollen wall development, aperture formation and pollen tube growth in Brassica campestris. Ann. Bot. 2014, 113, 777–788. [Google Scholar] [CrossRef]
- Wodehouse, R. Pollen Grains: Their Structure, Identification and Significance; Medicine, S.a., Ed.; Hafner Publishing Co.: New York, NY, USA, 1935. [Google Scholar]
- Blackmore, S.; Crane, P. The systematic implications of pollen and spore ontogeny. In Ontogeny and Systematics; Humpries, C.J., Ed.; Columbia University Press: New York, NY, USA, 1988; pp. 83–115. [Google Scholar]
- Ressayre, A.; Mignot, A.; Siljak-Yakovlev, S.; Raquin, C. Post-meiotic cytokinesis and pollen aperture number determination in eudicots: Effect of the cleavage wall number. Protoplasma 2003, 221, 257–268. [Google Scholar] [CrossRef] [PubMed]
- Ressayre, A.; Godelle, B.; Raquin, C.; Gouyon, P.H. Aperture pattern ontogeny in angiosperms. J. Exp. Biol. (Mol. Dev. Evol.) 2002, 294, 122–135. [Google Scholar] [CrossRef] [PubMed]
- Ressayre, A.; Raquin, C.; Mignot, A.; Godelle, B.; Gouyon, P.H. Correlated variation in microtubule distribution, callose deposition during male post-meiotic cytokinesis, and pollen aperture number across Nicotiana species (Solanaceae). Am. J. Bot. 2002, 89, 393–400. [Google Scholar] [CrossRef] [PubMed]
- Hulskamp, M.; Parekh, N.S.; Grini, P.; Schneitz, K.; Zimmermann, I.; Lolle, S.J.; Pruitt, R.E. The STUD gene is required for male-specific cytokinesis after telophase II of meiosis in Arabidopsis thaliana. Dev. Biol. 1997, 187, 114–124. [Google Scholar] [CrossRef] [PubMed]
- Spielman, M.; Preuss, D.; Li, F.L.; Browne, W.E.; Scott, R.J.; Dickinson, H.G. TETRASPORE is required for male meiotic cytokinesis in Arabidopsis thaliana. Development 1997, 124, 2645–2657. [Google Scholar] [CrossRef]
- Yang, C.Y.; Spielman, M.; Coles, J.P.; Li, Y.; Ghelani, S.; Ghelani, S.; Bourdon, V.; Brown, R.C.; Lemmon, B.E.; Scott, R.J.; et al. TETRASPORE encodes a kinesin required for male meiotic cytokinesis in Arabidopsis. Plant J. 2003, 34, 229–240. [Google Scholar] [CrossRef]
- Albert, B.; Raquin, C.; Prigent, M.; Nadot, S.; Brisset, F.; Yang, M.; Ressayre, A. Successive microsporogenesis affects pollen aperture pattern in the tam mutant of Arabidopsis thaliana. Ann. Bot. 2011, 107, 1421–1426. [Google Scholar] [CrossRef]
- Magnard, J.-L.; Yang, M.; Chen, Y.-C.S.; Leary, M.; McCormick, S. The Arabidopsis gene tardy asynchronous meiosis is required for the normal pace and synchrony of cell division during male meiosis. Plant Physiol. 2001, 127, 1157–1166. [Google Scholar] [CrossRef]
- D’Erfurth, I.; Jolivet, S.; Froger, N.; Catrice, O.; Novatchkova, M.; Mercier, R. Turning meiosis into mitosis. PLoS Biol. 2009, 7, 1–10. [Google Scholar] [CrossRef]
- Prieu, C.; Matamoro-Vidal, A.; Raquin, C.; Dobritsa, A.; Mercier, R.; Gouyon, P.H.; Albert, B. Aperture number influences pollen survival in Arabidopsis mutants. Am. J. Bot. 2016, 103, 452–459. [Google Scholar] [CrossRef]
- Reeder, S.; Lee, B.; Fox, R.; Dobritsa, A. A ploidy-sensitive mechanism regulates aperture formation on the Arabidopsis pollen surface and guides localization of the aperture factor INP1. PLoS Genet. 2016, 12, e1006060. [Google Scholar] [CrossRef]
- Dover, G.A. The organization and polarity of pollen mother cells of Triticum aestivum. J. Cell Sci. 1972, II, 699–711. [Google Scholar] [CrossRef]
- Sheldon, J.; Dickinson, H. Determination of patterning in the pollen wall of Lilium henryi. J. Cell Sci. 1983, 63, 191–208. [Google Scholar] [CrossRef] [PubMed]
- Sheldon, J.; Dickinson, H. Pollen wall formation in Lilium: The effect of chaotropic agents, and the organization of the microtubular cytoskeleton during pattern development. Planta 1986, 168, 11–23. [Google Scholar] [CrossRef] [PubMed]
- Albert, B.; Nadot, S.; Dreyer, L.; Ressayre, A. The influence of tetrad shape and intersporal callose wall formation on pollen aperture pattern ontogeny in two eudicot species. Ann. Bot. 2010, 106, 557–564. [Google Scholar] [CrossRef]
- Blaise, S. Etude palynologique de deux espèces voisines de Plantains. Bull. Soc. Bot. Fr. 1963, 110, 91–107. [Google Scholar] [CrossRef]
- Erdtman, G. Handbook of Palynology; Munksgaard: Copenhagen, Denmark, 1969; pp. 78–81. [Google Scholar]
- Kessler, L.G.; Larson, D.A. Effects of polyploidy on pollen grains diameter and other exomorphic exine features in Tridax coronifolia. Pollen Spores 1969, 11, 203–211. [Google Scholar]
- Cartier, D. Etude biosystématique de quelques espèces du genre Plantago (Tourn) L. I. Historique, races chromosomiques du Plantago alpina L. et du Plantago serpentina All. Rev. Gén. Bot. 1971, 78, 493–556. [Google Scholar]
- Mignot, A. Contraintes et Sélection dans l'Évolution: Le Cas du Pollen; Université Paris-Saclay: Gif-sur-Yvette, France, 1995. [Google Scholar]
- Matamoro-Vidal, A.; Prieu, C.; Furness, C.; Albert, B.; Gouyon, P.H. Evolutionary stasis in pollen morphogenesis due to natural selection. New Phytol. 2016, 209, 376–394. [Google Scholar] [CrossRef]
- Williams, J.; Mazer, S. Pollen-Tiny and ephemeral but not forgotten: New ideas on their ecology and evolution. Am. J. Bot. 2016, 103, 365–374. [Google Scholar] [CrossRef]
- Van Campo, M. Patterns of pollen morphological variation within taxa. In The Evolutionary Significance of the Exine; Ferguson, I., Muller, M., Eds.; Academic Press: London, UK, 1976. [Google Scholar]
- Doyle, J.; Hotton, C. Diversification of early angiosperm pollen in a cladistic context. In Pollen and Spores: Patterns of Diversification; Clarendon Press: Oxford, UK, 1991; pp. 169–195. [Google Scholar]
- Mulcahy, G.B.; Mulcahy, D.L. A comparison of pollen tube growth in bi- and tri-nucleate pollen. In Pollen: Biology and Implications for Plant Breeding; Mulcahy, D.L., Ottaviano, E., Eds.; Elsevier Science: New York, NY, USA, 1983; pp. 35–41. [Google Scholar]
- Ottaviano, E.; Sari-gorla, M.; Frova, C.; Pe, E. Male gametophytic selection in higher plants. In Sexual Reproduction in Higher Plants; Cresti, M., Gori, P., Pacini, E., Eds.; Springer: Berlin/Heidelberg, Germany, 1988; pp. 35–42. [Google Scholar]
- Hoekstra, F.; Bruinsma, J. Respiration and vitality of binucleate and trinucleate pollen. Physiol. Plant. 1975, 34, 221–225. [Google Scholar] [CrossRef]
- Nadot, S.; Furness, C.A.; Sannier, J.; Penet, L.; Triki-Teurtroy, S.; Albert, B.; Ressayre, A. Phylogenetic comparative analysis of microsporogenesis in angiosperms with a focus on monocots. Am. J. Bot. 2008, 95, 1426–1436. [Google Scholar] [CrossRef] [PubMed]
- Penet, L.; Nadot, S.; Ressayre, A.; Forchioni, A.; Dreyer, L.; Gouyon, P.H. Multiple developmental pathways leading to a single morph: Monosulcate pollen (examples from the Asparagales). Ann. Bot. 2005, 95, 331–343. [Google Scholar] [CrossRef][Green Version]
- Sannier, J.; Nadot, S.; Forchioni, A.; Harley, M.M.; Albert, B. Variation in the microsporogenesis of monosulcate palm pollen. Bot. J. Linn. Soc. 2006, 151, 93–102. [Google Scholar] [CrossRef][Green Version]
- Toghranegar, Z. Diversité et Évolution de la Microsporogenèse chez les Angiospermes Basales et les Monocotylédones; Université Paris-Saclay: Gif-sur-Yvette, France, 2013. [Google Scholar]
- Albert, B.; Gouyon, P.-H.; Ressayre, A. Microsporogenesis variation in Codiaeum producing inaperturate pollen grain. CR Acad. Sci. Biol. 2009, 332, 507–516. [Google Scholar] [CrossRef] [PubMed]
- Matamoro-Vidal, A.; Furness, C.; Gouyon, P.; Wurdack, K.; Albert, B. Evolutionary stasis in Euphorbiaceae pollen: Selection and constraints. J. Evol. Biol. 2012, 25, 1077–1096. [Google Scholar] [CrossRef]
- Dajoz, I.; Till-Bottraud, I.; Gouyon, P.H. Evolution of pollen morphology. Science 1991, 253, 66–68. [Google Scholar] [CrossRef]
- Dajoz, I.; Till-Bottraud, I.; Gouyon, P.H. Pollen aperture polymorphism and gametophyte performance in Viola diversifolia. Evolution 1993, 47, 1080–1093. [Google Scholar] [CrossRef]
- Till-Bottraud, I.; Vincent, M.; Dajoz, I.; Mignot, A. Pollen aperture heteromorphism. Variation in pollen-type proportions along altitudinal transects in Viola calcarata. CR Acad. Sci. Biol. 1999, 322, 579–589. [Google Scholar] [CrossRef]
- Till-Bottraud, I.; Mignot, A.; Paepe, R.D.; Dajoz, I. Pollen Heteromorphism in Nicotiana tabacum (Solanaceae). Am. J. Bot. 1995, 82, 1040–1048. [Google Scholar] [CrossRef]
- Katifori, E.; Alben, S.; Cerda, E.; Nelson, D.; Dumais, J. Foldable structures and the natural design of pollen grains. Proc. Natl. Acad. Sci. USA 2010, 107, 7635–7639. [Google Scholar] [CrossRef]
- Matamoro-Vidal, A.; Raquin, C.; Brisset, F.; Colas, H.; Izac, B.; Albert, B.; Gouyon, P.H. Links between morphology and function of the pollen wall: An experimental approach. Bot. J. Linn. Soc. 2016, 180, 478–490. [Google Scholar] [CrossRef]
- Albert, B.; Ressayre, A.; Dillmann, C.; Carlson, A.L.; Swanson, R.J.; Gouyon, P.-H.; Dobritsa, A.A. Effect of aperture number on pollen germination, survival and reproductive success in Arabidopsis thaliana. Ann. Bot. 2018, 121, 733–740. [Google Scholar] [CrossRef]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Albert, B.; Matamoro-Vidal, A.; Prieu, C.; Nadot, S.; Till-Bottraud, I.; Ressayre, A.; Gouyon, P.-H. A Review of the Developmental Processes and Selective Pressures Shaping Aperture Pattern in Angiosperms. Plants 2022, 11, 357. https://doi.org/10.3390/plants11030357
Albert B, Matamoro-Vidal A, Prieu C, Nadot S, Till-Bottraud I, Ressayre A, Gouyon P-H. A Review of the Developmental Processes and Selective Pressures Shaping Aperture Pattern in Angiosperms. Plants. 2022; 11(3):357. https://doi.org/10.3390/plants11030357
Chicago/Turabian StyleAlbert, Beatrice, Alexis Matamoro-Vidal, Charlotte Prieu, Sophie Nadot, Irène Till-Bottraud, Adrienne Ressayre, and Pierre-Henri Gouyon. 2022. "A Review of the Developmental Processes and Selective Pressures Shaping Aperture Pattern in Angiosperms" Plants 11, no. 3: 357. https://doi.org/10.3390/plants11030357
APA StyleAlbert, B., Matamoro-Vidal, A., Prieu, C., Nadot, S., Till-Bottraud, I., Ressayre, A., & Gouyon, P.-H. (2022). A Review of the Developmental Processes and Selective Pressures Shaping Aperture Pattern in Angiosperms. Plants, 11(3), 357. https://doi.org/10.3390/plants11030357