
Fruit Anatomy of the Canarieae (Burseraceae)

Abstract
:1. Introduction
2. Results
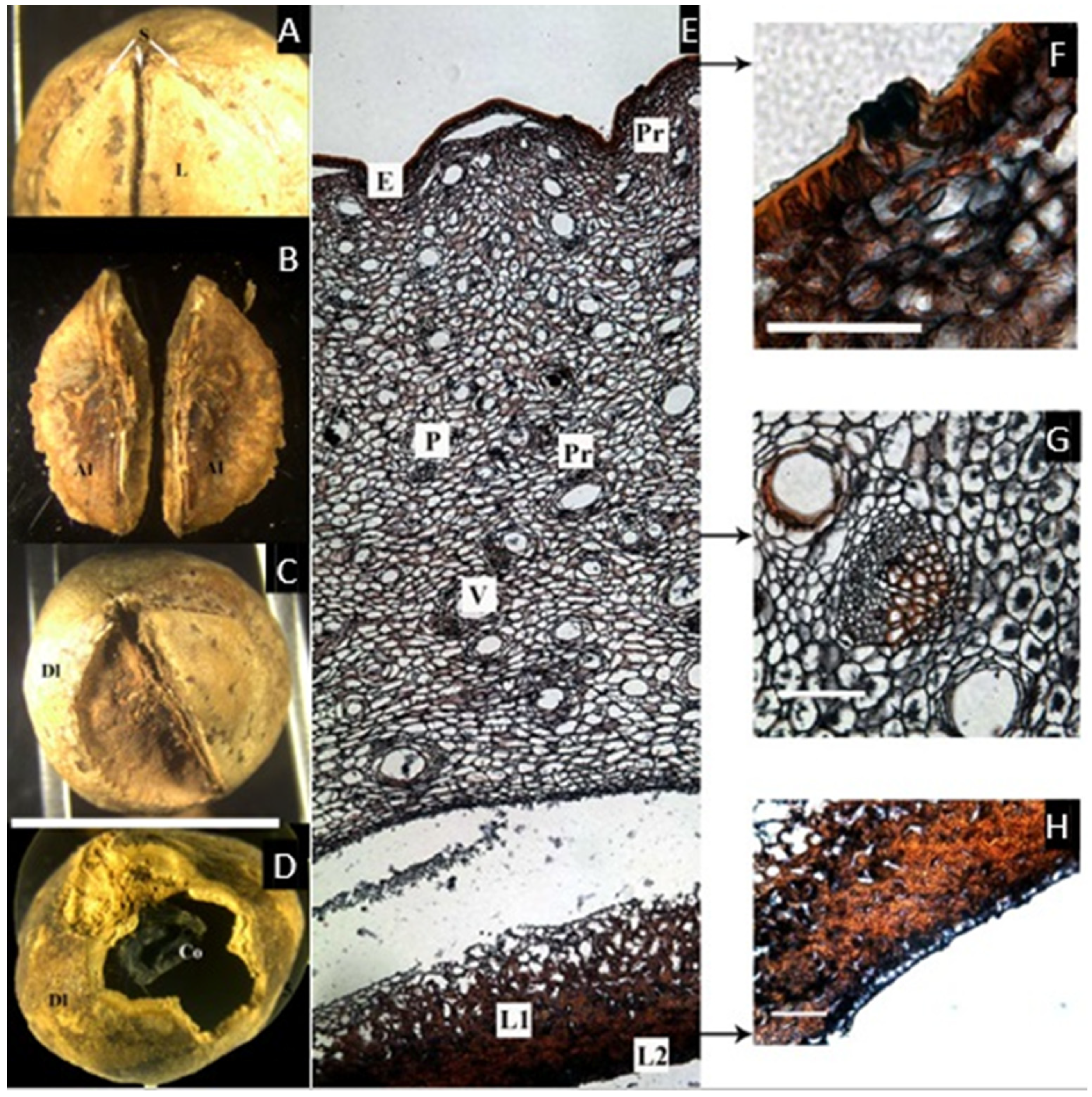
2.1. Canarium L.
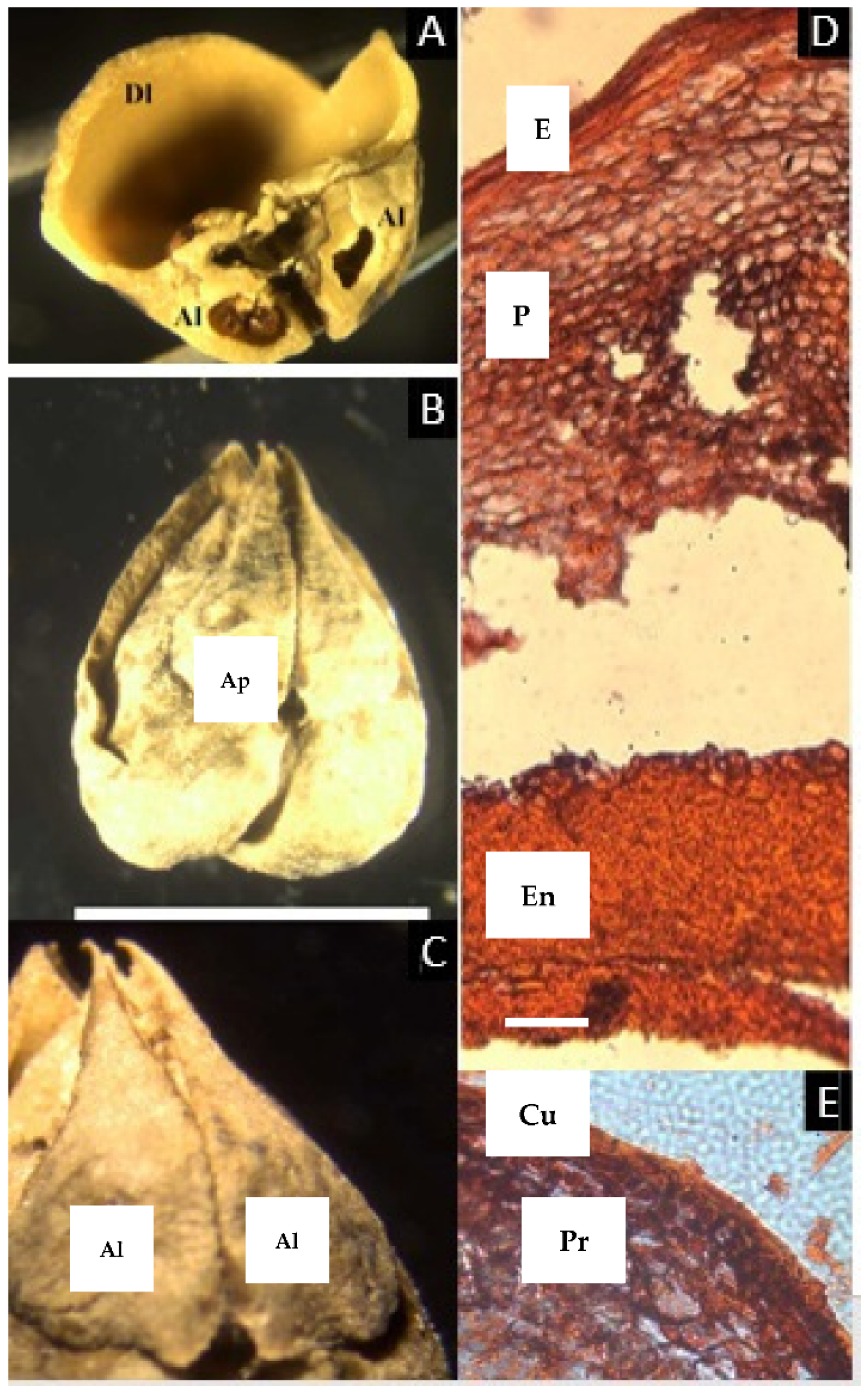
2.2. Trattinnickia Willd.
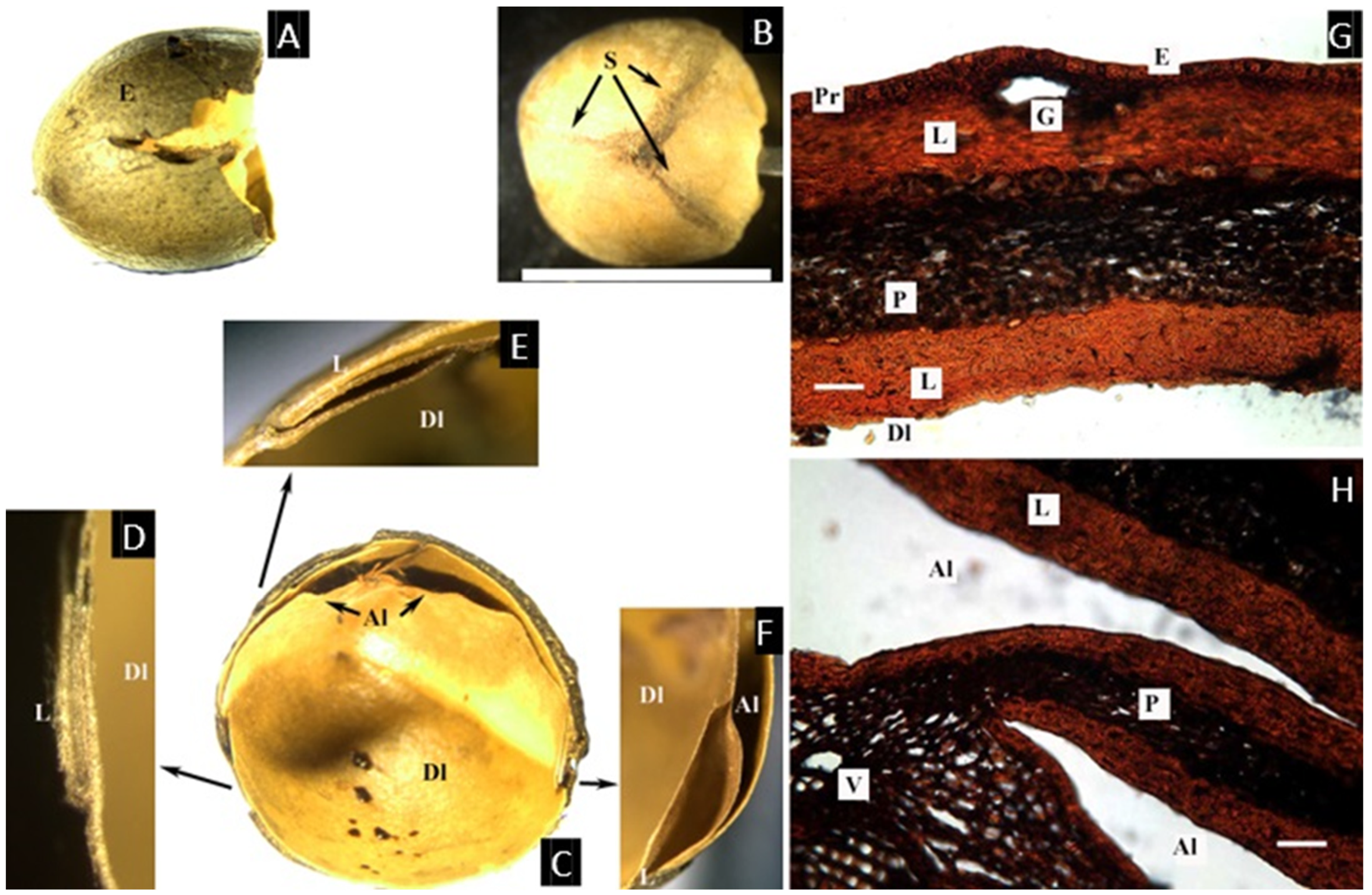
2.3. Dacryodes Vahl
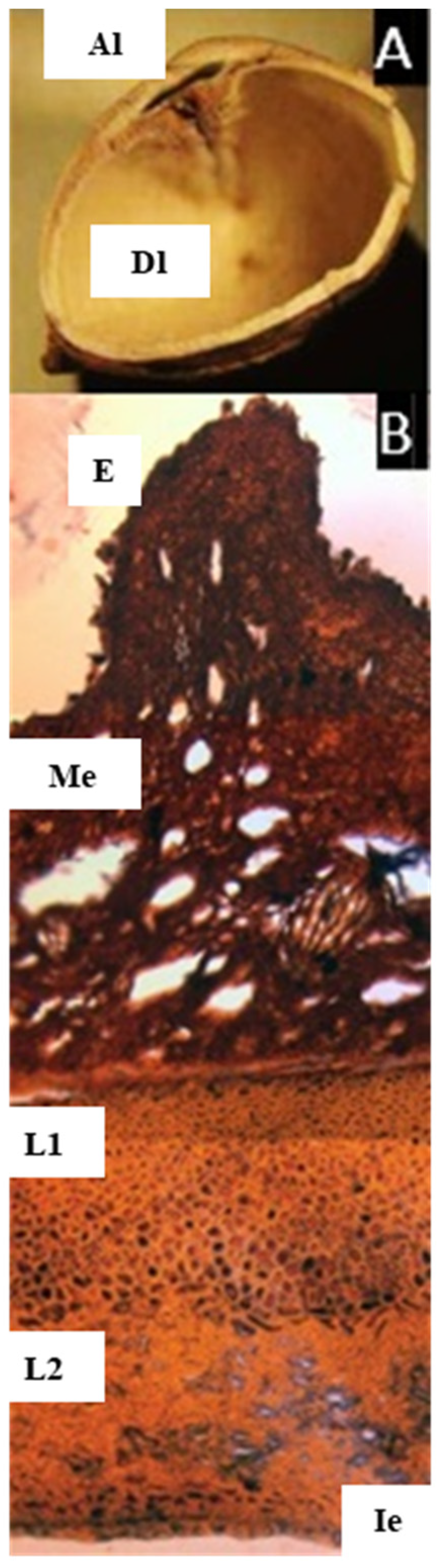
2.4. Pseudodacryodes Pierlot
2.5. Santiria Blume
2.6. Rosselia Forman
2.7. Haplolobus H. J. Lam
2.8. Scutinanthe Thwaites
2.9. Garuga Roxb.
3. Discussion
4. Materials and Methods
5. Conclusions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Engler, A. Burseraceae. D.C. Mon. Phan. 1883, 4, 1–169. [Google Scholar]
- Engler, A. Naturlichen Pflanzenfamilien; W. Engelmann: Leipzig, Germany, 1931; pp. 405–456. [Google Scholar]
- Daly, D.C.; Harley, M.M.; Martínez-Habibe, M.C.; Weeks, A. Burseraceae. In The Families and Genera of Vascular Plants. Flowering Plants. Eudicots: Sapindales, Cucurbitales, Myrtaceae, Vol. X; Kubitzki, K., Ed.; Springer: Berlin/Heidelberg, Germany, 2011; pp. 76–104. [Google Scholar] [CrossRef]
- Thulin, M.; Beier, B.-A.; Razafimandimbison, S.G.; Banks, H.I. Ambilobea, a New Genus from Madagascar, the Position of Aucoumea, and Comments on the Tribal Classification of the Frankincense and Myrrh Family (Burseraceae). Nord. J. Bot. 2008, 26, 218–229. [Google Scholar] [CrossRef] [Green Version]
- Weeks, A.; Zapata, F.; Pell, S.K.; Daly, D.C.; Mitchell, J.D.; Fine, P.V.A. To move or to evolve: Contrasting patterns of intercontinental connectivity and climatic niche evolution in “Terebinthaceae” (Anacardiaceae and Burseraceae). Front. Genet. 2014, 5, 409. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lam, H.J. The Burseraceae of the Malay archipelago and peninsula. Bull. Jard. Bot. Buitenzorg III 1932, 12, 281–561. [Google Scholar]
- Cuatrecasas, J. Notas a La Flora de Colombia. Rev. Acad. Colomb. Cienc. Exactas Físicas Nat. 1952, 8, 473–475. [Google Scholar]
- Cuatrecasas, J. The American Species Of Dacryodes. Trop. Woods 1957, 106, 46–65. [Google Scholar]
- Daly, D.C.; Martínez-Habibe, M.C. Notes on Dacryodes Vahl, including a new species from the Rio negro basin in Amazonia. studies in neotropical Burseraceae. XI. Brittonia 2002, 54, 266–274. [Google Scholar] [CrossRef]
- Leenhouts, P.W. Burseraceae. Flora Males. Ser. I 1956, 5, 209–296. [Google Scholar]
- Porter, D.M.; Pool, A. Burseraceae. In Flora de Nicaragua; Monographs in systematic botany from the Missouri Botanical Garden; Stevens, W.D., Ulloa, C., Pool, A., Montiel, O., Eds.; Missouri Botanical Garden Press: St. Louis, MO, USA, 2001; Volume 85. [Google Scholar]
- Spjut, R.W. A systematic treatment of fruit types. Mem. N. Y. Bot. Gard. 1994, 70, 1–182. [Google Scholar]
- Clarkson, J.J.; Chase, M.W.; Harley, M.M. Phylogenetic relationships in Burseraceae based on plastid rps16 intron sequences. Kew Bull. 2002, 57, 183–193. [Google Scholar] [CrossRef]
- Roth, I. Fruits of angiosperms. Handb. der Pflanz. 1977, 10, 1–675. [Google Scholar]
- Troupin, G. Burseraceae. In Flore du Congo Belge et du Ruanda-Urundi; Publications de l’Institut National pour l’Étude Agronomique du Congo Belge (I.N.É.A.C.): Bruxelles, Belgium, 1958; pp. 132–133. [Google Scholar]
- Whitmore, T. Tree flora of Malaya: A manual for foresters. Malay. Fo 1973, 2, 10–18. [Google Scholar]
- Ng, F.S.P. Manual of Forest Fruits, Seeds and Seedlings. Malayan Forest Record No 34; Center for International Forestry Research (CIFOR): Bogor, Indonesia, 1996. [Google Scholar]
- Hill, A.W. The method of germination of seeds enclosed in a stony endocarp. II. Ann. Bot. 1937, 1, 239–256. [Google Scholar] [CrossRef]
- Wannan, B.S.; Quinn, C.J. Pericarp structure and generic affinities in the Anacardiaceae. Bot. J. Linn. Soc. 1990, 102, 225–252. [Google Scholar] [CrossRef]
- Daly, D.C. Studies in neotropical Burseraceae. ii. generic limits in new world Protieae and Canarieae. Brittonia 1989, 41, 17–27. [Google Scholar] [CrossRef]
- Federman, S.; Dornburg, A.; Downie, A.; Richard, A.F.; Daly, D.C.; Donoghue, M.J. The biogeographic origin of a radiation of trees in Madagascar: Implications for the assembly of a tropical forest biome. BMC Evol. Biol. 2015, 15, 216. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pierlot, R. Pseudodacryodes Pierlot genre nouveau de Burseraceae de l’est de la rep. dem. du Congo. Bull. Jard. Bot. Nat. Belg. 1997, 66, 175–186. [Google Scholar] [CrossRef]
- Kochummen, K.M. Burseraceae. In Tree Flora of Sabah and Sarawak; Longman, Ed.; Forest Research Institute of Malaysia: Kuala Lumpur, Malaysia, 1972; Volume 1, pp. 46–47. [Google Scholar]
- Weeks, A.; Daly, D.C.; Simpson, B.B. The phylogenetic history and biogeography of the frankincense and myrrh family (Burseraceae) based on nuclear and chloroplast sequence data. Mol. Phylogenet. Evol. 2005, 35, 85–101. [Google Scholar] [CrossRef] [PubMed]
- Carlquist, S. The Use of Ethylenediamine in Softening Hard Plant Structures for Paraffin Sectioning. Stain Technol. 1982, 57, 311–317. [Google Scholar] [CrossRef] [PubMed]
- Sharman, B. Tannic Acid and Iron Alum with Safranin and Orange G in Studies of the Shoot Apex. Stain Technol. 1943, 18, 105–111. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Taxa | Axial Intrusion | Lignified Hypodermis | Articulated Plate | Pyrene Surface | Outermost Layer in Endocarp | Innermost Layer in Endocarp | Inner Epidermis in Developed Locule |
|---|---|---|---|---|---|---|---|
| Canarium asperum | + | - | - | smooth/ rugose | lignified | lignified | not observed |
| Dacryodes edilsonii | - | + | + | smooth | parenchymatic | lignified | not observed |
| Garuga floribunda | - | - | - | rugose | lignified | lignified | not observed |
| Haplolobus floribundus | - | - | - | smooth | parenchymatic | lignified | not observed |
| Pseudodacryodes leonardiana | - | - | - | smooth | parenchymatic | parenchymatic | observed |
| Rosselia bracteata | - | - | + | smooth | lignified | lignified | not observed |
| Santiria laevigata | - | - | + | smooth | - | lignified | observed |
| Scutinanthe brunnea | - | - | - | smooth | lignified | lignified | observed |
| Trattinnickia rhoifolia | - | - | - | rugose | parenchymatic | lignified | not observed |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Martínez-Habibe, M.C. Fruit Anatomy of the Canarieae (Burseraceae). Plants 2022, 11, 253. https://doi.org/10.3390/plants11030253
Martínez-Habibe MC. Fruit Anatomy of the Canarieae (Burseraceae). Plants. 2022; 11(3):253. https://doi.org/10.3390/plants11030253
Chicago/Turabian StyleMartínez-Habibe, María C. 2022. "Fruit Anatomy of the Canarieae (Burseraceae)" Plants 11, no. 3: 253. https://doi.org/10.3390/plants11030253
APA StyleMartínez-Habibe, M. C. (2022). Fruit Anatomy of the Canarieae (Burseraceae). Plants, 11(3), 253. https://doi.org/10.3390/plants11030253