
DEFHAZ: A Mechanistic Weather-Driven Predictive Model for Diaporthe eres Infection and Defective Hazelnut Outbreaks

 ,
, 
Abstract
1. Introduction
2. Results
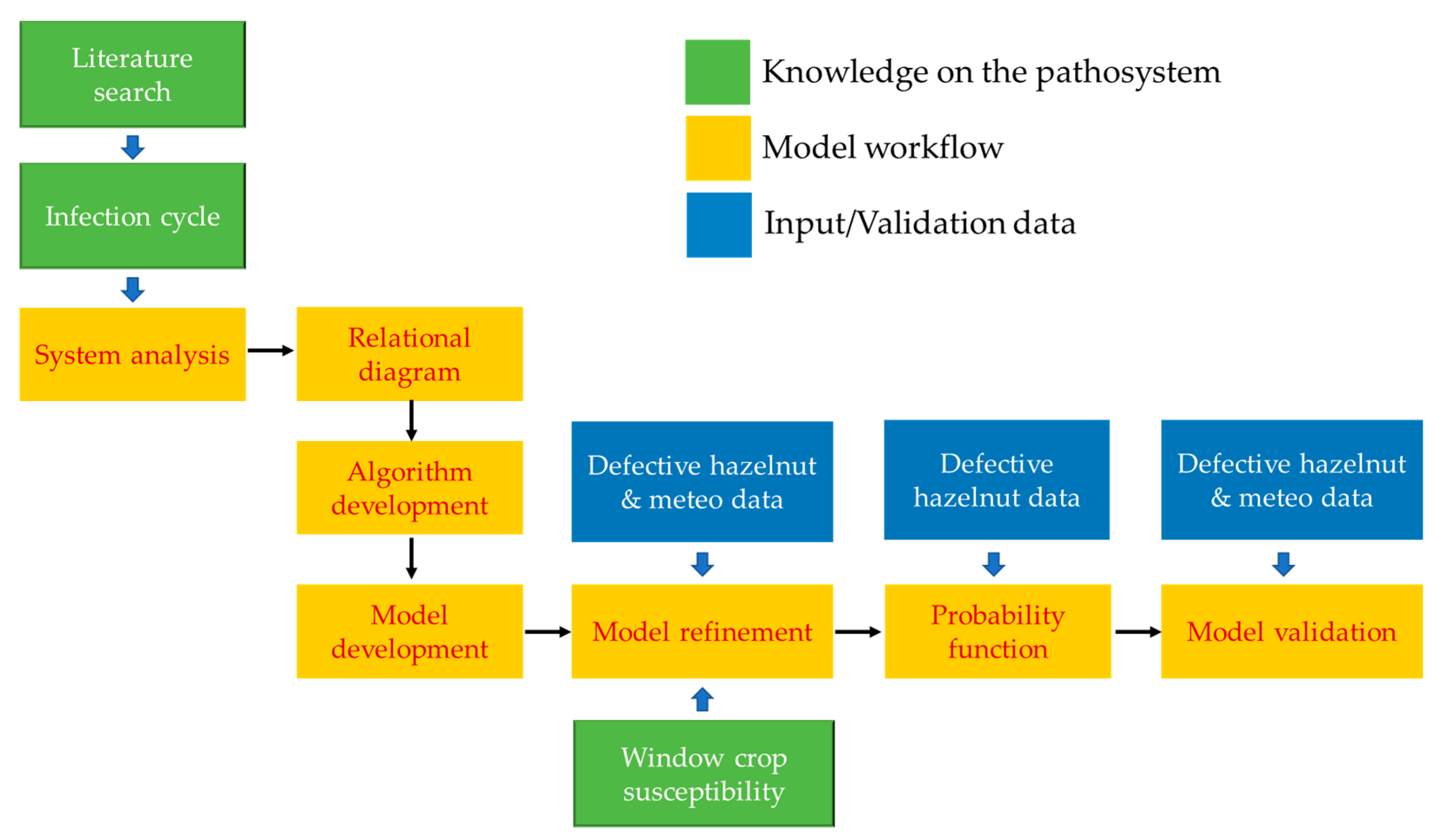
2.1. System Analysis of D. eres Lifecycle
2.2. Algorithm and Model Development
2.2.1. Overwintering Inoculum (Oi), Pycnidia Production Rate (PydR), and Conidiation Rate (CoR)
2.2.2. Dispersal Rate (DisR)
2.2.3. Germination Rate (GeR)
2.2.4. Infection Rate (IR)
2.2.5. Growth Rate (GR)
2.2.6. Defective Hazelnut Rate (DhR)
2.3. Model Refinement
Crop Susceptibility to Infection
2.4. Probability Function
Binary Logistic Regressions for HD and TD Estimations in Hazelnuts: Internal Validation
2.5. Model Internal Validation
3. Discussion
4. Materials and Methods
4.1. System Analysis and Model Development
4.1.1. Infection Cycle
4.1.2. Relation Diagram
4.2. Algorithm and Model Development
Functions and Algorithm
4.3. Model Input
4.4. Model Output
4.5. Field Data Collection
4.6. Model Refinement
4.6.1. Window for Crop Susceptibility to Infection
4.6.2. Probability Function
4.6.3. Model Internal Validation
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- FAOSTAT. Faostat Database Collections. Food and Agriculture Organization of the United Nations. Available online: http://www.fao.org/faostat/en/#data/QC (accessed on 4 February 2022).
- Teviotdale, B.L.; Michailides, T.J.; Pscheidt, J.W. Compendium of Nut Crop Diseases in Temperate Zones; APS Press, The American Phytopathological Society: St. Paul, MN, USA, 2002. [Google Scholar]
- ECE/TRADE/C/WP.7/2009/1. Specialized Section on Standardization of Dry and Dried Produce. Available online: https://unece.org/fileadmin/DAM/trade/agr/meetings/ge.02/2010/WP7_2009_19_Rev_HazelnutKernels.doc (accessed on 7 November 2022).
- Arciuolo, R.; Santos, C.; Soares, C.; Castello, G.; Spigolon, N.; Chiusa, G.; Lima, N.; Battilani, P. Molecular characterization of Diaporthe species associated with hazelnut defects. Front. Plant Sci. 2020, 11, 611655. [Google Scholar] [CrossRef]
- Battilani, P.; Chiusa, G.; Arciuolo, R.; Somenzi, M.; Fontana, M.; Castello, G.; Spigolon, N. Diaporthe as the main cause of hazelnut defects in the caucasus region. Phytopathol. Medit. 2018, 57, 320–333. [Google Scholar]
- Arciuolo, R.; Chiusa, G.; Castello, G.; Camardo Leggieri, M.; Spigolon, N.; Battilani, P. Diaporthe spp. Is confirmed as the main fungus associated with defective turkish hazelnuts. Plant Health Prog. 2022. [Google Scholar] [CrossRef]
- Rossman, A.Y.; Adams, G.C.; Cannon, P.F.; Castlebury, L.A.; Crous, P.W.; Gryzenhout, M.; Jaklitsch, W.M.; Mejia, L.C.; Stoykov, D.; Udayanga, D.; et al. Recommendations of generic names in Diaporthales competing for protection or use. IMA Fungus 2015, 6, 145–154. [Google Scholar] [CrossRef]
- Anco, D.J.; Madden, L.V.; Ellis, M.A. Temporal patterns of sporulation potential of Phomopsis viticola on infected grape shoots, canes, and rachises. Plant Dis. 2012, 96, 1297–1302. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Pscheidt, J.W.; Pearson, R.C. Phomopsis cane and leaf spot of grape. Dis. Identif. 1991, 6, 1–2. [Google Scholar]
- Nakatami, F.; Fujita, K. Diaporthe canker. In Compendium of Apple and Pear Diseases; Jones, A.L., Aldwinkle, H.S., Eds.; APS Press, The American Phytopathological Society: St. Paul, MN, USA, 1990; p. 38. [Google Scholar]
- Gonzalez-Dominguez, E.; Caffi, T.; Paolini, A.; Mugnai, L.; Latinović, N.; Latinović, J.; Languasco, L.; Rossi, V. Development and validation of a mechanistic model that predicts infection by Diaporthe ampelina, the causal agent of phomopsis cane and leaf spot of grapevines. Front. Plant Sci. 2022, 13, 872333. [Google Scholar] [CrossRef]
- Rosenberger, D.A. Phomopsis canker, rough bark, and fruit decay. In Compendium of Apple and Pear Diseases; Jones, A.L., Aldwinkle, H.S., Eds.; APS Press: St. Paul, MN, USA, 1990; pp. 38–39. [Google Scholar]
- Hewitt, W.B.; Pearson, R.C. Phomopsis cane and leaf spot. In Compendium of Grape Diseases; Pearson, R.C., Goheen, A.C., Eds.; APS Press: St. Paul, MN, USA, 1994; pp. 17–18. [Google Scholar]
- Erincik, O.; Madden, L.V.; Ferree, D.C.; Ellis, M.A. Temperature and wetness-duration requirements for grape leaf and cane infection by Phomopsis viticola. Plant Dis. 2003, 87, 832–840. [Google Scholar] [CrossRef]
- Wiman, N.G.; Webber, J.B., III; Wiseman, M.; Merlet, L. Identity and pathogenicity of some fungi associated with hazelnut (Corylus avellana L.) trunk cankers in oregon. PLoS ONE 2019, 14, e0223500. [Google Scholar] [CrossRef]
- Guerrero-Contreras, J.; Galdames-Gutierrez, R.; Ogass-Contreras, K.; Pérez-Fuentealba, S. First report of Diaporthe foeniculina causing black tip and necrotic spot on hazelnut kernel in chile. Plant Dis. 2020, 104, 975. [Google Scholar] [CrossRef]
- Pscheidt, J.W.; Heckert, S.; Wiseman, M.; Jones, L. Fungi associated with and influence of moisture on development of kernel mold of hazelnut. Plant Dis. 2019, 103, 922–928. [Google Scholar] [CrossRef] [PubMed]
- Arciuolo, R.; Leggieri, M.C.; Chiusa, G.; Castello, G.; Genova, G.; Spigolon, N.; Battilani, P. Ecology of Diaporthe eres, the causal agent of hazelnut defects. PLoS ONE 2021, 16, e0247563. [Google Scholar] [CrossRef] [PubMed]
- Rossi, V.; Sperandio, G.; Caffi, T.; Simonetto, A.; Gilioli, G. Critical success factors for the adoption of decision tools in IPM. Agronomy 2019, 9, 710. [Google Scholar] [CrossRef]
- Commission, E. Directive 2009/128/ec of the european parliament and of the council of 21 october 2009 establishing a framework for community action to achieve the sustainable use of pesticides. Off. J. Eur. Union 2009, 2009, 16. [Google Scholar]
- Pscheidt, J.W.; Heckert, S. Progression of kernel mold on hazelnut. Plant Disease 2021, 105, 1320–1327. [Google Scholar] [CrossRef] [PubMed]
- Valeriano, T.; Fischer, K.; Ginaldi, F.; Giustarini, L.; Castello, G.; Bregaglio, S. Rotten hazelnuts prediction via simulation modeling—A case study on the turkish hazelnut sector. Front. Plant Sci. 2022, 13, 766493. [Google Scholar] [CrossRef]
- Caffi, T.; Rossi, V.; Cossu, A.; Fronteddu, F. Empirical vs mechanistic model for primary infections of Plasmopora viticola. EPPO Bull. 2007, 37, 261–271. [Google Scholar] [CrossRef]
- Camardo Leggieri, M.; Fels-Klerx, H.J.v.d.; Battilani, P. Cross-validation of predictive models for deoxynivalenol in wheat at harvest. World Mycotox. J. 2013, 6, 389–397. [Google Scholar] [CrossRef]
- Lefferlaar, P.A. Basic elements of dynamic simulation. In On system Analysis and Simulation of Ecological Processes; Leffelaar, P.A., Ed.; Kluwer Academic Publishers: Dordrecht, The Netherlands, 1993; pp. 11–28. [Google Scholar]
- Rossi, V.; Onesti, G.; Legler, S.; Caffi, T. Use os system analysis to develop plant sidease models based on literature data: Grape black-rot as a case study. Europ. J. Plant Path. 2015, 141, 427–444. [Google Scholar] [CrossRef]
- Camardo Leggieri, M.; Mazzoni, M.; Battilani, P. Machine learning for predicting mycotoxin occurrence in maize. Front. Microbiol. 2021, 12, 661132. [Google Scholar] [CrossRef]
- Liakos, K.G.; Busato, P.; Moshou, D.; Pearson, S.; Bochtis, D. Machine learning in agriculture: A review. Sensors 2018, 18, 2674. [Google Scholar] [CrossRef] [PubMed]
- Battilani, P.; Arciuolo, R.; Chiusa, G.; (Università Cattolica del Sacro Cuore, Piacenza, 29121, PC, Italy). Diaporthe eres pycnidia development and conidia release in different temperature regimes. Personal communication, 2022. [Google Scholar]
- Analytis, S. Über die relation zwischen biologischer entwicklung und temperatur bei phytopathogenen pilzen. Phytopathol. Z. 1977, 90, 64–76. [Google Scholar] [CrossRef]
- Paradinas, A.; Ramade, L.; Mulot-Greffeuille, C.; Hamidi, R.; Thomas, M.; Toillon, J. Phenological growth stages of ‘barcelona’ hazelnut (Corylus avellana L.) described using an extended bbch scale. Sci. Hortic. 2022, 296, 110902. [Google Scholar] [CrossRef]
- Cristofori, V.; Ferramondo, S.; Bertazza, G.; Bignami, C. Nut and kernel traits and chemical composition of hazelnut (Corylus avellana L.) cultivars. J. Sci. Food Agric. 2008, 88, 1091–1098. [Google Scholar] [CrossRef]
- Karaogladinis, G.S.; Bardas, G. First report of phomopsis fruit decay on apple caused by Phomopsis mali in Greece. Plant Dis. 2006, 90, 375. [Google Scholar] [CrossRef]
- Sezer, A.; Dolar, F.S. Prevalence of Botrytis cinerea pers. Ex fr. Causing disease on fruit clusters in hazelnut growing areas of Ordu, Giresun and Trabzon provinces and determination of the reactions of some cultivars against pathogen. Bitki Koruma Bülteni 2012, 52, 93–110. [Google Scholar]
- Simşek, O.; Arici, M.; Demir, C. Mycoflora of hazelnut (Corylus avellana L.) and aflatoxin content in hazelnut kernels artificially infected with Aspergillus parasiticus. Nahrung 2002, 46, 194–196. [Google Scholar] [CrossRef]
- Aloi, F.; Zamora-Ballesteros, C.; Martín-García, J.; Diez, J.J.; Cacciola, S.O. Co-infections by Fusarium circinatum and Phytophthora spp. on Pinus radiata: Complex phenotypic and molecular interactions. Plants 2021, 10, 1976. [Google Scholar] [CrossRef]
- Weiland, J.E.; Benedict, C.; Zasada, I.A.; Scagel, C.R.; Beck, B.R.; Davis, A.; Graham, K.; Peetz, A.; Martin, R.R.; Dung, J.K.S.; et al. Late-summer disease symptoms in western washington red raspberry fields associated with co-occurrence of Phytophthora rubi, Verticillium dahliae, and Pratylenchus penetrans, but not raspberry bushy dwarf virus. Plant Dis. 2018, 102, 938–947. [Google Scholar] [CrossRef]
- Giorni, P.; Bertuzzi, T.; Battilani, P. Impact of fungi co-occurrence on mycotoxin contamination in maize during the growing season. Front. Microbiol. 2019, 10, 1265. [Google Scholar] [CrossRef]
- Camardo Leggieri, M.; Lanubile, A.; Dall’Asta, C.; Pietri, A.; Battilani, P. The impact of seasonal weather variation on mycotoxins: Maize crop in 2014 in northern italy as a case study. World Mycotox. J. 2020, 13, 25–36. [Google Scholar] [CrossRef]
- Lanubile, A.; Giorni, P.; Bertuzzi, T.; Marocco, A.; Battilani, P. Fusarium verticillioides and Aspergillus flavus co-occurrence influences plant and fungal transcriptional profiles in maize kernels and in vitro. Toxins 2021, 13, 680. [Google Scholar] [CrossRef] [PubMed]
- Houshyarfard, M.; Davood, J. The Aspergillus flavus susceptibility of hazelnut varieties (Corylus avellana L.) in laboratory conditions. J. Nuts 2018, 9, 181–188. [Google Scholar]
- Kabak, B. Aflatoxins in hazelnuts and dried figs: Occurrence and exposure assessment. Food Chem. 2016, 211, 8–16. [Google Scholar] [CrossRef] [PubMed]
- Ozay, G.; Seyhan, F.; Pembeci, C.; Saklar, S.; Yilmaz, A. Factors influencing fungal and aflatoxin levels in turkish hazelnuts (Corylus avellana L.) during growth, harvest, drying and storage: A 3-year study. Food Addit. Contam. 2008, 25, 209–218. [Google Scholar] [CrossRef]
- Agrios, G.N. Plant Pathology, 5th ed.; Elsevier Academic Press: Burlington, MA, USA, 2005. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
| Rate | Variable | Function | Reference in Text | Parameter | |||
|---|---|---|---|---|---|---|---|
| a | b | c | R2 | ||||
| PydR | GDD_T | Logistic | 1 | 6.27 | −0.01 | 0.96 | 0.950 |
| PydR | GDD_aw | Logistic | 1 | 9.64 | −0.02 | 0.98 | 0.971 |
| GerT | T | Bete | 2–3 | 13.49 | 1.10 | 3.69 | 0.793 |
| GerRH | RH | Polynomial | 4 | 11.85 | 1110.80 | * n.a. | 0.975 |
| GRT | T | Bete | 2–3 | 5.06 | 1.39 | 3.46 | 0.964 |
| GRaw | aw | Logistic | 1 | 73.59 | −79.05 | 0.97 | 0.987 |
| b | S.E. | Wald a | df | Prob. b | Exp (b) c | |
|---|---|---|---|---|---|---|
| Hidden defects | ||||||
| BBCH81 vs. BBCH81 | ||||||
| Dh-I 30-5 | 0.000 d | 0.000 | 0.810 | 1 | 0.368 | 1.000 |
| Constant | −2.449 | 0.621 | 15.533 | 1 | 0.000 | 0.086 |
| BBCH89 vs. BBCH89 | ||||||
| Dh-I 30-5 | 0.000 | 0.000 | 1.340 | 1 | 0.247 | 1.000 |
| Constant | −1.890 | 0.374 | 25.475 | 1 | 0.000 | 0.151 |
| BBCH81 vs. BBCH89 | ||||||
| Dh-I 30-5 | 0.000 | 0.000 | 1.860 | 1 | 0.173 | 1.000 |
| Constant | −1.908 | 0.366 | 27.244 | 1 | 0.000 | 0.148 |
| Total defects | ||||||
| BBCH81 vs. BBCH81 | ||||||
| Dh-I 30-5 | 0.000 | 0.000 | 3.914 | 1 | 0.048 | 1.000 |
| Constant | −2.546 | 0.476 | 28.647 | 1 | 0.000 | 0.078 |
| BBCH89 vs. BBCH89 | ||||||
| Dh-I 30-5 | 0.000 | 0.000 | 8.708 | 1 | 0.003 | 1.000 |
| Constant | −1.033 | 0.297 | 12.127 | 1 | 0.000 | 0.356 |
| BBCH81 vs. BBCH89 | ||||||
| Dh-I 30-5 | 0.000 | 0.000 | 5.861 | 1 | 0.015 | 1.000 |
| Constant | −0.904 | 0.280 | 10.424 | 1 | 0.001 | 0.405 |
| Hidden Defective Hazelnuts | Observed | Predicted | |
|---|---|---|---|
| Negative | Positive | ||
| BBCH81 vs. BBCH81 | Negative | 95 | 0 |
| Positive | 5 | 0 | |
| BBCH89 vs. BBCH89 | Negative | 84 | 0 |
| Positive | 16 | 0 | |
| BBCH81 vs. BBCH89 | Negative | 84 | 0 |
| Positive | 16 | 0 | |
| Total Defective Hazelnuts | |||
| BBCH81 vs. BBCH81 | Negative | 89 | 0 |
| Positive | 11 | 0 | |
| BBCH89 vs. BBCH89 | Negative | 56 | 5 |
| Positive | 23 | 16 | |
| BBCH81 vs. BBCH89 | Negative | 59 | 3 |
| Positive | 26 | 12 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Camardo Leggieri, M.; Arciuolo, R.; Chiusa, G.; Castello, G.; Spigolon, N.; Battilani, P. DEFHAZ: A Mechanistic Weather-Driven Predictive Model for Diaporthe eres Infection and Defective Hazelnut Outbreaks. Plants 2022, 11, 3553. https://doi.org/10.3390/plants11243553
Camardo Leggieri M, Arciuolo R, Chiusa G, Castello G, Spigolon N, Battilani P. DEFHAZ: A Mechanistic Weather-Driven Predictive Model for Diaporthe eres Infection and Defective Hazelnut Outbreaks. Plants. 2022; 11(24):3553. https://doi.org/10.3390/plants11243553
Chicago/Turabian StyleCamardo Leggieri, Marco, Roberta Arciuolo, Giorgio Chiusa, Giuseppe Castello, Nicola Spigolon, and Paola Battilani. 2022. "DEFHAZ: A Mechanistic Weather-Driven Predictive Model for Diaporthe eres Infection and Defective Hazelnut Outbreaks" Plants 11, no. 24: 3553. https://doi.org/10.3390/plants11243553
APA StyleCamardo Leggieri, M., Arciuolo, R., Chiusa, G., Castello, G., Spigolon, N., & Battilani, P. (2022). DEFHAZ: A Mechanistic Weather-Driven Predictive Model for Diaporthe eres Infection and Defective Hazelnut Outbreaks. Plants, 11(24), 3553. https://doi.org/10.3390/plants11243553