Abstract
While studying aromas produced by the edible flowers of Tulbaghia violacea, we noticed a different production of (Z)-3-Hexenyl acetate (a green-leaf volatile, GLV) by purple (var. ‘Violacea’) and white (var. ‘Alba’) flowers. The white Tulbaghia flowers constantly emits (Z)-3-Hexenyl acetate, which is instead produced in a lower amount by the purple-flowered variety. Thus, we moved to analyze the production of (Z)-3-Hexenyl acetate by whole plants of the two varieties by keeping them confined under a glass bell for 5 h together with a SPME (Solid Phase Micro Extraction) fiber. Results show that six main volatile compounds are emitted by T. violacea plants: (Z)-3-Hexenyl acetate, benzyl alcohol, nonanal, decanal, (Z)-3-Hexenyl-α-methylbutyrate, and one unknown compound. By cutting at half-height of the leaves, the (Z)-3-Hexenyl acetate is emitted in high quantities from both varieties, while the production of (Z)-3-Hexenyl-α-methylbutyrate increases. (Z)-3-Hexenyl acetate is a GLV capable of stimulating plant defenses, attracting herbivores and their natural enemies, and it is also involved in plant-to-plant communication and defense priming. Thus, T. violacea could represent a useful model for the study of GLVs production and a ‘signal’ plant capable of stimulating natural defenses in the neighboring plants.
1. Introduction
Tulbaghia violacea Harv. (1837) is a perennial-rhizomatous plant native to South Africa, now used as an ornamental plant and variously distributed throughout the world. Linnaeus coined the genus in honor of governor the Dutch Cape Colony, Ryjk Tulbagh (1699–1771), who sent him numerous specimens of the local flora [1]. The species name naturally refers to the purplish color of the flowers. Both the leaves and the flowers have a garlic-like flavor and are used for flavoring salads and other dishes; in fact, common T. violacea names recall garlic: society garlic, sweet garlic, and wild garlic. The popularity of the species is linked to the mistaken belief that its consumption does not leave an unpleasant breath as happens after garlic consumption [2]. A common characteristic between plants of the Tulbaghia and Allium genera is a ‘garlic’ smell that is produced when leaves or rhizomes are damaged; the precursors of the typical odor are cysteine-derived amino acids present in the cytoplasm, and S-(methylthiomethyl)-cysteine-4-oxide (marasmin) in the case of Tulbaghia and S-allyl-cysteine sulfoxide (alliin) in the case of Allium. When the tissues are damaged, a C-S lyase, present in the vacuoles of the cells, comes into contact with marasmin (or alliin), cleaving the compound into thiosulphinate marasmicin (or allicin) [3]. Marasmicin is not stable and decomposes giving origin of other volatile sulfur compounds, especially if Tulbalghia organs or tissues are subjected to heat treatment [4,5].
We became interested in T. violacea in the course of a study focusing on volatile compounds emitted by edible flowers and coincidentally noticed differences in the emission of volatile compounds between white and purple flowers of different varieties of T. violacea (var ‘Violacea’ and ‘Alba’); our attention was drawn to the emission of (Z)-3-Hexenyl acetate (Z3HAC), a compound part of the green leaf volatiles (GLVs), emitted by plants both constitutively and in response to wounding and attacks by herbivores [6], which are also involved in triggering plant-defense responses via jasmonic acid [7,8].
GLVs, with their typical smell of freshly cut grass, are produced by almost all plants and constitute an important group of volatile organic compounds emitted by plants. They are C6 aldehydes and acetylated C6 aldehydes deriving from the lipid oxidation of fatty acids in the oxylipin pathway (common to jasmonic acid) and are rapidly emitted as a result of tissue damage due to biotic or abiotic stress [6]. GLVs as Z-3-Hexenal, (E)-2-Hexenal, Z-3-Hexenol, and Z3HAC represent communication and defense signals that can activate defensive reactions in the same plant and neighboring plants against herbivores and pathogens, and repel or attract herbivores and their natural enemies, developing in addition some toxicity towards bacteria and fungi [6,9]. In addition, GLVs represent a pleasant powerful green-note/fruity-aroma component of apples and tropical fruits [10,11] so as to be efficiently employed e.g., in a recipe for the artificial imitation of mango flavor [12].
The aim of this study was to find differences of GLVs, if any, between white and purple flowers of T. violacea, both in normal conditions and in response to wounding.
2. Results
2.1. Main Volatile Compounds Emitted by Tulbaghia Flowers
Edible T. violacea flowers (Figure 1) show an emission of similar volatile compounds but in different ratio depending on the variety/flower color. In fact, in both flower types, we detected mainly the emission of 2,4,5,7-Tetrathiaoctane and Z3HAC in a ratio, calculated as a peak area, of approximately 4:1 for the purple flowers and of 2:1 for white flowers. Table 1 and Figure 2 summarize more clearly the identified volatile organic compounds (VOCs) emitted by flowers of the two varieties of Tulbaghia.

Figure 1.
Purple and white flowers of T. violacea from the two varieties. (A): var. ‘Violacea’: (B): var. ‘Alba’.
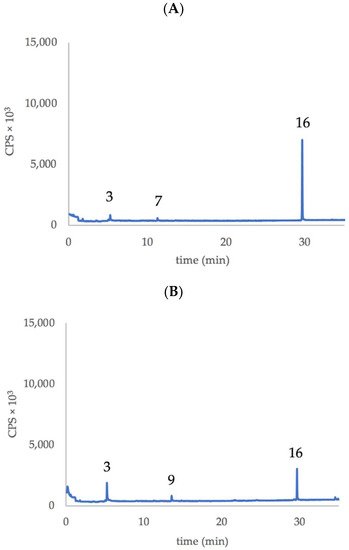
Figure 2.
Representative chromatograms showing the main VOCs emitted by the flowers T. violacea. (A) purple flowers of var. ‘Violacea’, (B) white flowers of var. ‘Alba’. Numbers indicate the compounds listed in Table 1.

Table 1.
Main VOCs emitted by T. violacea flowers var. ‘Violacea’ and ‘Alba’, purple and white flowers, respectively. Numbering of volatile compounds respects the order of elution considering the set of compounds identified after the analysis of flowers and of the whole plants (see Table 2) so as to keep the numbering univocal. The values given are the averages of three repetitions; standard-error values are not reported but were all values within 9%.
Table 2.
VOCs emitted by whole plants of T. violacea and captured by SPME fiber in 5 h wounding indicates that 50% of the leaves were cut at half of their height (+: presence; -: absence/trace amount). Numbering of volatile compounds respects the order of elution considering the set of compounds identified after the analysis of flowers and of the whole plants (see Table 1) so as to keep the numbering univocal.
The relevant finding is the diverse emission ratio between Z3HAC and 2,4,5,7-Tetrathiaoctane by the two different flowers (Figure 2), suggesting a different pattern between the two varieties of Tulbaghia regarding the emission of GLVs and the production of metabolites derived from marasmicin. In fact, 2,4,5,7-Tetrathiaoctane and 2,3,5-Trithiahexane are the result of flower-peduncle cutting (about 5–6 flowers make 1 g), manipulation and subsequent heating of the sample and fiber during the analytical procedure. The damaged (or homogenized) Tulbaghia tissues allow the contact between marasmin and a C-S lyase that cleaves the compound, yielding (methylthio)methane-sulfenic acid, a short-lived compound that condenses into marasmicin (2,4,5,7-tetrathiaoctane 4-oxide), which in turn is transformed to 2,4,5,7-Tetrathiaoctane and 2,3,5-Trithiahexane, particularly upon boiling of T. violacea rhizomes [4,5].
In addition to the previous results, a semiquantitative determination of the Z3HAC emitted by Tulbaghia flowers confirms that ‘Alba’ flowers emit approximately 2.5 times more than those of the var. ‘Violacea’ per g fresh weight (Table 3).
Table 3.
Semi-quantitative determination of (Z)-3-Hexenyl acetate in white and purple flowers of T. violacea (ng/g fresh weight).
2.2. Volatile Compounds Emitted by Whole Tulbaghia Plants
Thus, the next step was to check whether the emission of Z3HAC by whole Tulbaghia plants varied according to variety and/or wounding treatment; for this purpose, we proceeded to collect, via SPME fibers, the volatile compounds emitted by plants kept for several hours inside a glass bell (Figure 3).

Figure 3.
T. violacea plant in a pot (A) and under a glass bell in the presence of SPME fiber (B).
Table 2 presents the compounds released by whole plants of the two varieties of Tulbalghia before and after wounding (50% of the leaves cut at half of their height). None of the 16 compounds detected appeared to be variety-specific, but five of them appeared to be produced specifically as a result of wounding, although in a very low amount: (Z)-3-Hexen-1-ol,propanoate, (Z)-1,7-Octadiene-3,6-diol,2,6-dimethyl, (Z)-Butanoic acid,3-hexenyl ester, (Z)-1,6-Octadien-3-ol, 3,7-dimethyl-formate, and (Z)-Hex-3-enyl(E)-2-methylbut-2-enoate. In fact, of these, only (Z)-1,7-Octadiene-3,6-diol,2,6-dimethyl (compound no. 8) appeared evident in representative chromatograms of volatile compounds emitted by plants of the two varieties, both control plants and after wounding (Figure 4B,D).
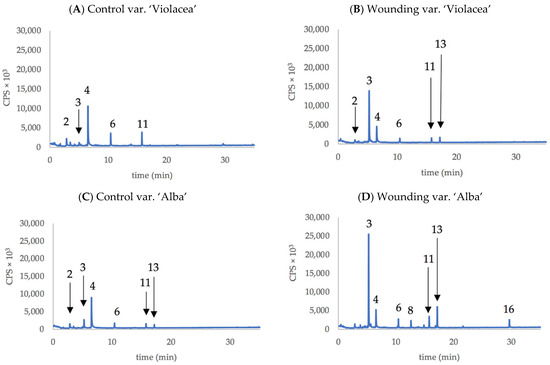
Figure 4.
Chromatograms of main VOCs produced by the whole plant of purple-flowered T. violacea ((A): Control; (B): after wounding) and white-flowered T. violacea ((C): Control; (D): after wounding). Numbers indicate the compounds listed in Table 2.
In both Tulbaghia varieties, there was an increase in the production of Z3HAC and of some related compounds, such as (Z)-3-Hexenyl-α-methylbutyrate (compound 13 in Figure 4).
The main volatile compounds, such as benzyl alcohol, nonanal, and decanal, also persisted, while most of the sulfur compounds deriving from marasmin, which are emitted by flowers, were not detected in this analysis, except in trace amounts. This is because, in this case, the ratio of damaged tissues/whole biomass was very small.
Regarding the production of Z3HAC, the wounding treatment resulted in a significant increase in emission in both Tulbaghia varieties, over 4000% for the ‘Violacea’ and about a quarter for the ‘Alba’, with the latter, however, emitting about twice as much following wounding treatment (Table 4).
Table 4.
Semi-quantitative determination of (Z)-3-Hexenyl acetate emitted after wounding by whole plants of white-flowered and purple flowered T. violacea (ng/plant in 5 h).
3. Discussion
The Z3HAC, which are derived from C6-aldehydes as (Z)-3-Hexenal and n-Hexenal, are metabolites produced from oxylipins (oxygenated compounds derived from fatty acids) along one of the two main oxylipin-pathway branches regulated by hydroperoxide lyase [13,14,15]. The other oxylipin-pathway branch, key-enzyme, allene-oxide-synthase, gives origin to jasmonates which are involved in a huge amount of defense responses [16,17,18].
We focused our work by finding that T. violacea plants emits significant amounts of Z3HAC; such compound was easily detected employing a solid-phase, micro-extraction methodology, both from the flowers and from the whole plant. The emission was lower in the purple flowers than in white flowers, whereas, by cutting at about half-height of the plant leaves, the response of the two Tulbaghia varieties was an emission of Z3HAC of approximately 10 to 50 times (up to 260 and 440 ng plant−1 h−1, respectively), compared with the non-wounding plants (Table 4). In addition, the production of the related compound cis-3-Hexenyl-α-methylbutyrate increased slightly (Figure 4).
More than fifteen years ago, Z3HAC and (Z)-3-Hexenol were identified as volatile compounds present in very low amounts (0.1–0.6%) in oil obtained from aerial parts of T. violacea by Pino et al. [19], but thereafter, such presence was not associated with GLVs. The only somehow related reference is the use of oil extract of T. violacea to strongly repel aphids [20].
An increase in the emission of Z3HAC as a result of wounding or mechanical damage was also detected in other plants. For instance, in corn plants, Z3HAC was released in greater quantities by plants damaged by insects (Elasmopalpus lignosellus larvae) than by plants not damaged or damaged manually (using a needle); notably, the volatile compound was released in large quantities after 24 and 96 h after treatment [21]. Instead, an increase of 7 or 18 times in the amount of Z3HAC emitted was observed using a SPME fiber for soybean leaves or pods mechanically damaged (with an entomological needle), in comparison to tissues damaged by insects (Rhyssomatus nigerrimus, Mexican weevil of soy) or healthy plants [22].
In addition to being involved in the internal communication of the plant and with neighboring plants as a trigger for immune-defense responses [6], Z3HAC appeared to be an attractive compound for both beneficial and harmful insects.
Concerning the attraction of beneficial insects, Arimura et al. [23] and Ozawa et al. [24] have observed that Lotus japonica plants infested by the spider mites Tetranychus urticae start to emit several volatiles, including a significant amount of Z3HAC and methyl salicylate, attracting Phytoseiuus persimilis (a predatory mites). Chehab et al. [13] indicated that Z3HAC is the predominant compound in the volatile blend emitted by Arabidopsis after aphid attack that mediates the attraction of aphid natural enemies (parasitoid wasp Aphidius colemani); Xavier et al. [21] revealed that the parasitoid, Trichogramma pretiosum, is attracted by VOCs released (mainly Z3HAC) by corn plants damaged by the larva of the insect, Elasmopalpus lignosellus, while Hegde et al. [25] reported that cotton plants, emitting a volatile blend including Z3HAC, repels the cotton aphid (Aphis gossypii). Also, it was described that Z3HAC is one of the major volatile components emitted by wheat seedlings, particularly when infested with aphids, and that spraying it on wheat field determines an attraction of ladybugs, reducing the abundance of aphids [26]. Interestingly, it is also demonstrated by electroantennograms and Y-tube olfactometer experiments that Z3HAC alone or cis-jasmone alone are attractive to Campoletis chlorideae, the parasitoid of the cotton bollworm, Helicoverpa armigera, but not the mixture of the two compounds [27].
Unfortunately, Z3HAC attracts harmful insects, such as the cutworm P. saucia [28] and the Holotrichia oblita beetle [29], but this phenomenon turns to be a vantage of pheromone traps. One case is the attraction of Plutella xylostella (cruciferous moth) which, in field experiments, were caught in greater numbers with synthetic-sex-pheromone traps containing Z3HAC alone or a mixture of Z3HAC, (Z)-3-Hexen-1-ol, and (E)-2-Hexenal, compared to sex pheromone alone [30]; other cases are the efficacy of sex-pheromone traps in a pear orchard where more males of Grapholita molesta (oriental fruit moth) are captured with traps containing pheromone plus Z3HAC then with the addition of 1-undecanol [31], and the increased catches of Agriotes brevis (click beetles, harmful to maize) when a mix of Z3HAC, methyl benzoate, (Z)-3-Hexenol, and methyl salicylate is added to the synthetic pheromone [32]. In field tests, Z3HAC is also a specific attractant of female Holotrichia parallela (dark black chafers, an important pest of several crops), enabling the capture of six times more insects than the pheromone-L-leucine-methyl ester; on the contrary, for the capture of H. parallela males, pheromone is found to be about three times more effective [33].
On the other hand, it has been observed that plants with higher emission levels of Z3HAC can attract female insects for oviposition: in Xin et al. [34], the insect, Empoasca vitis, chooses tea-plants varieties with higher Z3HAC emission levels; in another study [35], it is observed that Cnaphalocrocis medinalis has a negligible impact on corn whether there are rice plants in the vicinity, speculating that this could be due to different emission levels of VOCs, including Z3HAC. This feature could be exploited by the employment of ‘bait’ plants, removing them from the field after the oviposition stage, or using them for monitoring and detecting the abundance of phytophagous.
Finally, a study by Najdabbasi et al. [36] addresses the potential role of Z3HAC in decreasing the severity of late blight, the most important fungal disease of potato plants; Z3HAC provides a protection to pre-exposed plants against five genotypes of P. infestans, reducing late-blight severity by around 70% in a very susceptible potato cultivar. Similarly, Ameye et al. [37] demonstrated that exposure of Triticum aestivum plants to Z3HAC (priming) before artificial infection with Fusarium graminearum reduced the incidence of the relative pathology of Fusarium head blight. They observe a significant reduction of both fungal biomass and spikelets showing necrotic lesions in preexposed plants and a different expression of jasmonic acid- and salicylic acid-responsive genes. This suggests new opportunities for sustainable control of fungal diseases through activation of plant immunity.
There are slight increases of (Z)-3-Hexenyl-α-methylbutyrate after wounding (Figure 4); a similar increase for this Z3HAC derivate (and for the simpler butyrate form) was disclosed by McCall et al. [38] for heavily damaged cotton seedlings (by five Helicoverpa zea caterpillars/plant).
Thus, the two varieties of T. violacea, particularly the ‘Alba’ variety, represent a tool for studying the bio-synthetic pathway leading to Z3HAC production as well as ‘signal’ organisms capable of stimulating natural defenses in neighboring plants. Additionally, the two varieties of Tulbaghia could be employed for screening insects attracted to or repelled by Z3HAC with the aim of developing effective, low-cost, biological-control protocols.
4. Materials and Methods
4.1. Plant Culture Conditions
Plants of T. violacea var. ‘Violacea’ and var. ‘Alba’ were obtained from the Ortogourmet company (Laterza, Taranto, Italy) in pots of 18 cm in diameter and then transferred and grown in pots of 10 cm in diameter in an unheated greenhouse from October to April (and for the rest of the year in the open air) in the Botanical Garden of the University of Salento, Lecce, Italy.
4.2. Analysis of Volatile Organic Compounds
The analyses were carried out by solid-phase micro extraction (SPME) methodology, as described previously by Negro et al. [39] and Dimita et al. [40]. Tulbaghia flowers were collected at full bloom and 5–6 of them (approximately 1g FW) were immediately sealed into 20 mL SPME vials (Agilent Technologies, Palo Alto, CA, USA) by metal screw-caps with pre-notched, Teflon-silicone septa. The vials were then placed at 40 °C for 10 min in a thermostatically controlled bath to allow the evaporation of the compounds; hereafter, a SPME syringe was inserted, and the fiber (50/30 µm Divinylbenzene/Carboxen/Polydimethylsiloxane, Supelco/Merck KGaA, Darmstadt, Germany), which was previously conditioned for 5 min at 235 °C in the gas-chromatograph injector, was exposed for 10 min to absorb the volatile compounds.
In a wounding experiment, whole plants with approximately 18–22 leaves and no flowers (both in the case of control plant and after wounding with 50% of the leaves cut at half their height) were confined under a glass bell, resting on a wooden base with a hole in the middle, so that the epigeal part of the plant could be isolated from the pot (Figure 4). Additionally, in order to avoid contaminating the SPME fiber with soil emissions, an aluminum foil was placed between the wooden base and the soil, surrounding the root collar. The SPME syringe (previously conditioned for 5 min at 235 °C in the gas-chromatograph injector) was tied in the middle of the bell to a stick and exposed for 5 h at room temperature (Figure 3).
Subsequently, the fiber was inserted into the injector port of a gas chromatography with a mass-spectrometry detector (Agilent 7890B coupled with MS single quadrupole Agilent 5977A), and the desorption of the volatile compounds performed at 235 °C for 4 min. At this point, the chromatographic run was started with an Agilent HP-5 ms column (30 m × 0.25 mm, 0.25 µm) (where temperature was raised from 60 °C to 230 °C with a constant increase of 3 °C/minute), with a helium (purity >99.999%) constant flow of 1.0 mL/min. Compounds were identified by library search and analytical standard if available. The mass spectrum of an unknown compound was searched in a data-processing system [41]. Substances with a score above 800, both for identity and purity, were putatively identified after comparing the detected compound with the one in the NIST Computational Chemistry Comparison and Benchmark database [41]. Retention index (RI) was obtained, as reported by Zhao et al. [42], being employed as a reference of the retention times of a series of C8–C20 alkanes separated under the GC-MS conditions mentioned above; the following formula was applied:
where, ta is the retention time of the unknown peak a; tn the retention time of n-alkane Cn; and tn+1 the retention time of n-alkane Cn+1; n = carbon number of the alkane which elutes before the unknown peak a.
The semi-quantitative analysis of volatile compounds was carried out in the same experimental conditions, using known quantities of (Z)-3-Hexenyl acetate.
4.3. Statistics
All data were reported as the mean ± standard deviation (SD), with at least three replications for each sample. Statistical evaluation was conducted by Duncan’s test to discriminate among the mean values. All statistical analyses were performed using the software, Statistica (StatSoft, Tulsa, OK, USA).
Author Contributions
Conceptualization, R.D., S.M.A. and C.N.; methodology, C.N. and R.A.; formal analysis, A.F. and C.N.; investigation, R.D., S.M.A., R.A. and C.N.; resources, L.D.B.; data curation, C.N. and A.L.; writing—original draft preparation, A.F., R.A. and A.L.; writing—review and editing: L.D.B., F.B. and C.M.; supervision, L.D.B., F.B. and C.M.; funding acquisition, A.L. and L.D.B. All authors have read and agreed to the published version of the manuscript.
Funding
This research was partially funded by the Italian Ministry of Education, University and Research, Programma Operativo Nazionale FSE-FESR Ricerca e Innovazione 2014–2020, Asse I “Investimenti in Capitale Umano”, Azione I.1 “Dottorati Innovativi con caratterizzazione industriale”.
Data Availability Statement
Not applicable. All results are included in the article.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Dillon, H.; Nelson, E.C. 166. Tulbaghia leucantha: Alliaceae. Kew Mag. 1991, 8, 12–15. Available online: http://www.jstor.org/stable/45067180 (accessed on 12 August 2022).
- Lyantagaye, S.L. Ethnopharmacological and phytochemical review of Allium species (sweet garlic) and Tulbaghia species (wild garlic) from Southern Africa. Tanzan. J. Sci. 2011, 37, 58–72. Available online: https://www.ajol.info/index.php/tjs/article/view/73615 (accessed on 12 August 2022).
- Ranglová, K.; Krejčová, P.; Kubec, R. The effect of storage and processing on antimicrobial activity of Tulbaghia violacea. S. Afr. J. Bot. 2015, 97, 159–164. [Google Scholar] [CrossRef]
- Kubec, R.; Velísek, J.; Musah, R.A. The amino acid precursors and odor formation in society garlic (Tulbaghia violacea Harv.). Phytochemistry 2002, 60, 21–25. [Google Scholar] [CrossRef]
- Kubec, R.; Krejčová, P.; Mansur, L.; García, N. Flavor Precursors and Sensory-Active Sulfur Compounds in Alliaceae Species Native to South Africa and South America. J. Agric. Food Chem. 2013, 61, 1335–1342. [Google Scholar] [CrossRef]
- Ameye, M.; Allmann, S.; Verwaeren, J.; Smagghe, G.; Haesaert, G.; Schuurink, R.C.; Audenaert, K. Green leaf volatile production by plants: A meta-analysis. New Phytol. 2018, 220, 666–683. [Google Scholar] [CrossRef]
- Engelberth, J.; Alborn, H.T.; Schmelz, E.A.; Tumlinson, J.H. Airborne signals prime plants against insect herbivore attack. Proc. Natl. Acad. Sci. USA 2004, 101, 1781–1785. [Google Scholar] [CrossRef]
- Li, T.; Cofer, T.; Engelberth, M.; Engelberth, J. Defense Priming and Jasmonates: A Role for Free Fatty Acids in Insect Elicitor-Induced Long Distance Signaling. Plants 2016, 5, 5. [Google Scholar] [CrossRef]
- Meents, A.K.; Mithöfer, A. Plant-Plant Communication: Is There a Role for Volatile Damage-Associated Molecular Patterns? Front. Plant Sci. 2020, 11, 583275. [Google Scholar] [CrossRef]
- Laohakunjit, N.; Kerdchoechuen, O.; Matta, F.B.; Silva, J.L.; Holmes, W.E. Postharvest Survey of Volatile Compounds in Five Tropical Fruits Using Headspace-solid Phase Microextraction (HS-SPME). HortScience 2007, 42, 309–314. [Google Scholar] [CrossRef]
- Mostafa, S.; Wang, Y.; Zeng, W.; Jin, B. Floral Scents and Fruit Aromas: Functions, Compositions, Biosynthesis, and Regulation. Front. Plant Sci. 2022, 13, 860157. [Google Scholar] [CrossRef]
- Zhang, W.; Zhu, G.; Zhu, G. The imitation and creation of a mango flavor. Food Sci. Technol. 2022, 42, e34622. [Google Scholar] [CrossRef]
- Chehab, E.; Kaspi, R.; Savchenko, T.; Dehesh, K. Hexenyl acetate mediates indirect plant defense responses. Proc. ANAS (Biol. Sci.) 2010, 65, 145–151. [Google Scholar]
- Griffiths, G. Biosynthesis and analysis of plant oxylipins. Free Radic. Res. 2015, 49, 565–582. [Google Scholar] [CrossRef] [PubMed]
- Wasternack, C.; Feussner, I. The Oxylipin Pathways: Biochemistry and Function. Annu. Rev. Plant Biol. 2018, 69, 363–386. [Google Scholar] [CrossRef] [PubMed]
- Devoto, A.; Turner, J.G. Regulation of jasmonate-mediated plant responses in Arabidopsis. Ann. Bot. 2003, 92, 329–337. [Google Scholar] [CrossRef]
- Griffiths, G. Jasmonates: Biosynthesis, perception and signal transduction. Essays Biochem. 2020, 64, 501–512. [Google Scholar] [CrossRef] [PubMed]
- Sohn, S.-I.; Pandian, S.; Rakkammal, K.; Largia, M.J.V.; Thamilarasan, S.K.; Balaji, S.; Zoclanclounon, Y.A.B.; Shilpha, J.; Ramesh, M. Jasmonates in plant growth and development and elicitation of secondary metabolites: An updated overview. Front. Plant Sci. 2022, 13, 942789. [Google Scholar] [CrossRef]
- Pino, J.A.; Quijano-Celís, C.E.; Fuentes, V. Volatile compounds of Tulbaghia violacea harv. J. Essent. Oil-Bear. Plants 2008, 11, 203–207. [Google Scholar] [CrossRef]
- Richter, J.M.; Prinsloo, G.J.; Van Der Linde, T.C.D.K. The Response of Alate Diuraphis noxia (Kurdjumov) (Hemiptera: Aphididae) to Volatile Substances from Four Non-Host Plant Extracts Under Laboratory Conditions. Afr. Entomol. 2014, 22, 783–789. [Google Scholar] [CrossRef]
- Xavier, L.M.S.; Laumann, R.A.; Borges, M.; Magalhães, D.M.; Vilela, E.F.; Blassioli-Moraes, M.C. Trichogramma pretiosum attraction due to the Elasmopalpus lignosellus damage in maize. Pesq. Agropec. Bras. 2011, 46, 578–585. [Google Scholar] [CrossRef]
- Espadas-Pinachoa, K.; López-Guillénb, L.; Gómez-Ruiza, J.; Cruz-Lópeza, L. Induced volatiles in the interaction between soybean (Glycine max) and the Mexican soybean weevil (Rhyssomatus nigerrimus). Braz. J. Biol. 2021, 81, 611–620. [Google Scholar] [CrossRef] [PubMed]
- Arimura, G.; Ozawa, R.; Shimoda, T.; Nishioka, T.; Boland, W.; Takabayashi, J. Herbivory-induced volatiles elicit defense genes in lima bean leaves. Nature 2000, 406, 512–515. [Google Scholar] [CrossRef] [PubMed]
- Ozawa, R.; Shimoda, T.; Kawaguchi, M.; Arimura, G.; Horiuchi, J.; Nishioka, T.; Takabayashi, J. Lotus japonicus infested with herbivorous mites emits volatile compounds that attract predatory mites. J. Plant Res. 2000, 113, 427–433. [Google Scholar] [CrossRef]
- Hegde, M.; Oliveira, J.N.; da Costa, J.G.; Bleicher, E.; Goulart Santana, A.E.; Bruce, T.J.A.; Caulfield, J.; Dewhirst, S.Y.; Woodcock, C.M.; Pickett, J.A.; et al. Identification of semiochemicals released by cotton, Gossypium hirsutum, upon infestation by the cotton aphid, Aphis gossypii. J. Chem. Ecol. 2011, 37, 741–750. [Google Scholar] [CrossRef] [PubMed]
- Qiao, F.; Cai, Z.-P.; Su, J.-W. Herbivore-induced wheat volatiles facilitate biocontrol of the cereal aphid by the multicolored Asian ladybeetle. Chin. J. Entomol. 2020, 57, 189–195. [Google Scholar] [CrossRef]
- Sun, Y.-L.; Dong, J.-F.; Huang, L.-Q.; Wang, C.-Z. The cotton bollworm endoparasitoid Campoletis chlorideae is attracted by cis-jasmone or cis-3-hexenyl acetate but not by their mixtures. Arthropod Plant Interact. 2020, 14, 169–179. [Google Scholar] [CrossRef]
- Sun, Y.-L.; Dong, J.-F.; Song, Y.-Q.; Wang, S.-L. GOBP1 from the Variegated Cutworm Peridroma saucia (Hübner) (Lepidoptera: Noctuidae) Displays High Binding Affinities to the Behavioral Attractant (Z)-3-Hexenyl acetate. Insects 2021, 12, 939. [Google Scholar] [CrossRef]
- Wei, H.-s.; Qin, J.-h.; Cao, Y.-z.; Li, K.-b.; Yin, J. Two classic OBPs modulate the responses of female Holotrichia oblita to three major ester host plant volatiles. Insect Mol. Biol. 2021, 30, 390–399. [Google Scholar] [CrossRef]
- Li, P.; Zhu, J.; Qin, Y.J. Enhanced Attraction of Plutella xylostella (Lepidoptera: Plutellidae) to Pheromone-Baited Traps With the Addition of Green Leaf Volatiles. Econ. Entomol. 2012, 105, 1149–1156. [Google Scholar] [CrossRef]
- Yu, H.; Feng, J.; Zhang, Q.; Xu, H. (Z)-3-hexenyl acetate and 1-undecanol increase male attraction to sex pheromone trap in Grapholita molesta (Busck) (Lepidoptera: Tortricidae). Int. J. Pest Manag. 2014, 61, 30–35. [Google Scholar] [CrossRef]
- Vuts, J.; Furlan, L.; Tóth, M. Female Responses to Synthetic Pheromone and Plant Compounds in Agriotes brevis Candeze (Coleoptera: Elateridae). J. Insect Behav. 2018, 31, 106–117. [Google Scholar] [CrossRef]
- Ju, Q.; Li, X.; Guo, X.-Q.; Du, L.; Shi, C.-R.; Qu, M.-J. Two Odorant-Binding Proteins of the Dark Black Chafer (Holotrichia parallela) Display Preferential Binding to Biologically Active Host Plant Volatiles. Front. Physiol. 2018, 9, 769. [Google Scholar] [CrossRef] [PubMed]
- Xin, Z.-J.; Li, X.-W.; Bian, L.; Sun, X.-L. Tea green leafhopper, Empoasca vitis, chooses suitable host plants by detecting the emission level of (3Z)-hexenyl acetate. Bull. Entomol. Res. 2017, 107, 77–84. [Google Scholar] [CrossRef]
- Chen, P.; Dai, C.; Liu, H.; Hou, M. Identification of Key Headspace Volatile Compounds Signaling Preference for Rice over Corn in Adult Females of the Rice Leaf Folder Cnaphalocrocis medinalis. J. Agric. Food Chem. 2022, 70, 9826–9833. [Google Scholar] [CrossRef]
- Najdabbasi, N.; Mirmajlessi, S.M.; Dewitte, K.; Ameye, M.; Mänd, M.; Audenaert, K.; Landschoot, S.; Haesaert, G. Green Leaf Volatile Confers Management of Late Blight Disease: A Green Vaccination in Potato. J. Fungi 2021, 7, 312. [Google Scholar] [CrossRef] [PubMed]
- Ameye, M.; Audenaert, K.; De Zutter, N.; Steppe, K.; Van Meulebroek, L.; Vanhaecke, L.; De Vleesschauwer, D.; Haesaert, G.; Smagghe, G. Priming of Wheat with the Green Leaf Volatile Z-3-Hexenyl Acetate Enhances Defense against Fusarium graminearum But Boosts Deoxynivalenol Production. Plant Physiol. 2015, 167, 1671–1684. [Google Scholar] [CrossRef]
- McCall, P.J.; Turlings, T.C.J.; Loughrin, J.; Proveaux, A.T.; Tumlinson, J.H. Herbivore-induced volatile emissions from cotton (Gossypium hirsutum L.) seedlings. J. Chem. Ecol. 1994, 20, 12. [Google Scholar] [CrossRef]
- Negro, C.; Dimita, R.; Min Allah, S.; Miceli, A.; Luvisi, A.; Blando, F.; De Bellis, L.; Accogli, R. Phytochemicals and Volatiles in Developing Pelargonium ‘Endsleigh’ Flowers. Horticulturae 2021, 7, 419. [Google Scholar] [CrossRef]
- Dimita, R.; Min Allah, S.; Luvisi, A.; Greco, D.; De Bellis, L.; Accogli, R.; Mininni, C.; Negro, C. Volatile Compounds and Total Phenolic Content of Perilla frutescens at Microgreens and Mature Stages. Horticulturae 2022, 8, 71. [Google Scholar] [CrossRef]
- NIST (National Institute of Standards and Technology) Computational Chemistry Comparison and Benchmark Database NIST Standard Reference Database Number 101. [CrossRef]
- Zhao, Y.Z.; Li, Z.G.; Tian, W.L.; Fang, X.M.; Su, S.K.; Peng, W.J. Differential Volatile Organic Compounds in Royal Jelly Associated with Different Nectar Plants. J. Integr. Agric. 2016, 15, 1157–1165. [Google Scholar] [CrossRef]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).