

Chemical Composition and Phytotoxic and Antibiofilm Activity of the Essential Oils of Eucalyptus bicostata, E. gigantea, E. intertexta, E. obliqua, E. pauciflora and E. tereticornis
 , ,
, ,  , , ,
, , ,  , and
, and 
Abstract
1. Introduction
2. Results
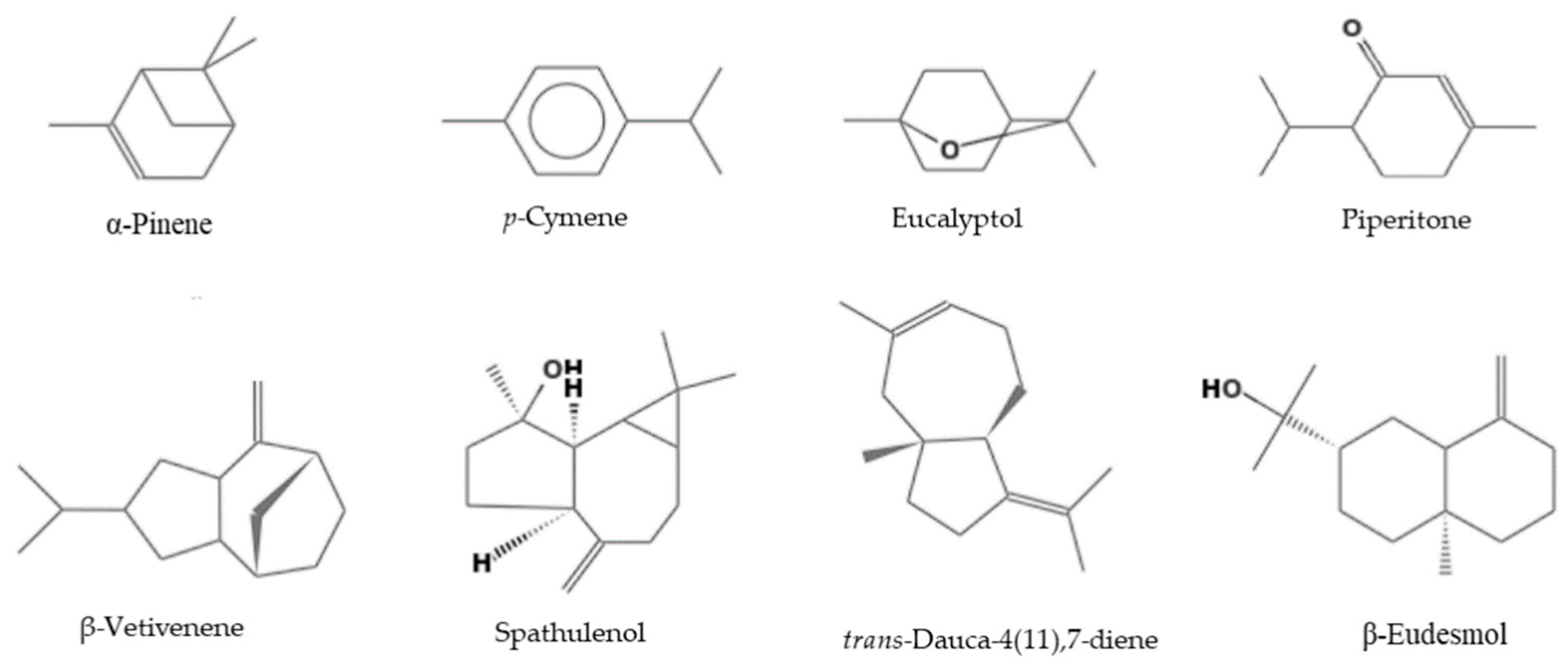
2.1. Composition of the EOs
2.2. Phytotoxic Activity
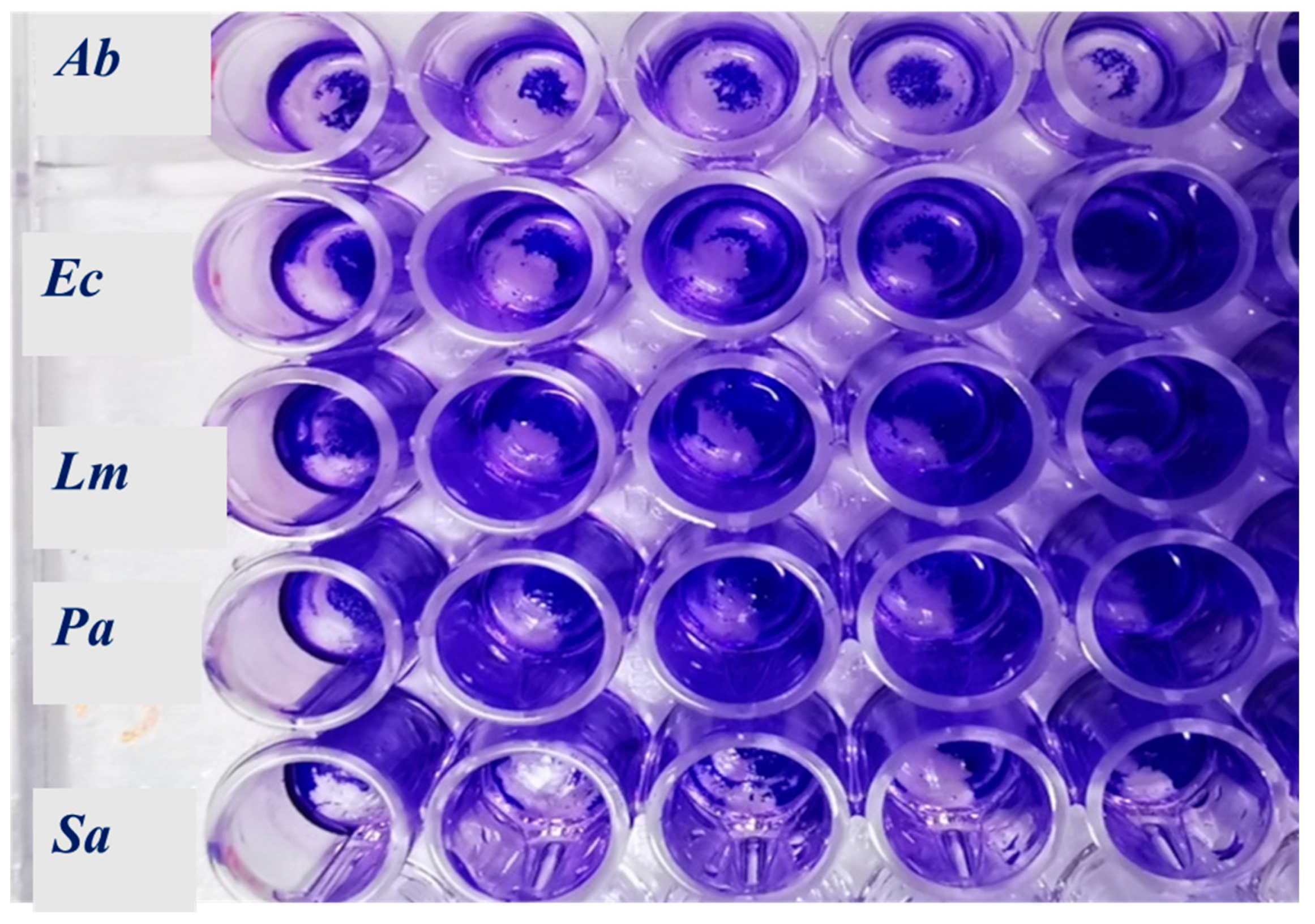
2.3. Antibacterial and Antibiofilm Activity
3. Discussion
4. Materials and Methods
4.1. Plant Material
4.2. Extraction of the Essential Oils
4.3. Analysis of the Essential Oils
4.4. Phytotoxic Activity
4.5. Antimicrobial Activity
4.5.1. Microorganisms and Culture Conditions
4.5.2. Minimal Inhibitory Concentration (MIC)
4.5.3. Biofilm Inhibitory Activity
4.5.4. Activity on Mature Bacterial Biofilm
4.5.5. Effects of EOs on Cell Metabolic Activity within the Biofilm
4.6. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Elaissi, A.; Marzouki, H.; Medini, H.; Larbi Khouja, M.; Farhat, F.; Lynene, F.; Harzallah-Skhiri, F.; Chemli, R. Variation in volatile leaf oils of 13 Eucalyptus species harvested from Souinet Arboreta (Tunisia). Chem. Biodivers. 2010, 7, 909–921. [Google Scholar] [CrossRef] [PubMed]
- Dhakad, A.K.; Pandey, V.V.; Beg, S.; Rawat, J.M.; Singh, A. Biological, medicinal and toxicological significance of Eucalyptus leaf essential oil: A review. J. Sci. Food Agric. 2018, 98, 833–848. [Google Scholar] [CrossRef] [PubMed]
- Sebei, K.; Sakouhi, F.; Herchi, W.; Khouja, M.L.; Boukhchina, S. Chemical composition and antibacterial activities of seven Eucalyptus species essential oils leaves. Biol. Res. 2015, 48, 7. [Google Scholar] [CrossRef] [PubMed]
- Pino, J.A.; Marbot, R.; Quert, R.; García, H. Study of essential oils of Eucalyptus resinifera Smith, E. tereticornis Smith and Corymbia maculata (Hook.) K.D. Hill & L.A.S. Johnson, grown in Cuba. Flavour Fragr. J. 2002, 17, 1–4. [Google Scholar]
- Toloza, A.C.; Lucia, A.; Zerba, E.; Masuh, H.; Picollo, M.I. Interspecific hybridization of Eucalyptus as a potential tool to improve the bioactivity of essential oils against permethrin-resistant head lice from Argentina. Bioresour. Technol. 2008, 99, 7341–7347. [Google Scholar] [CrossRef]
- Caputo, L.; Smeriglio, A.; Trombetta, D.; Cornara, L.; Trevena, G.; Valussi, M.; Fratianni, F.; De Feo, V.; Nazzaro, F. Chemical composition and biological activities of the essential oils of Leptospermum petersonii and Eucalyptus gunnii. Front. Microbiol. 2020, 11, 409. [Google Scholar] [CrossRef]
- Sliti, S.; Ayadi, S.; Kachouri, F.; Khouja, M.A.; Abderrabba, M.; Bouzouita, N. Leaf essential oils chemical composition, antibacterial and antioxidant activities of Eucalyptus camaldulensis and E. rudis from Korbous (Tunisia). J. Mater. Environ. Sci. 2015, 6, 743–748. [Google Scholar]
- Danna, C.; Cornara, L.; Smeriglio, A.; Trombetta, D.; Amato, G.; Aicardi, P.; De Martino, L.; De Feo, V.; Caputo, L. Eucalyptus gunnii and Eucalyptus pulverulenta ‘baby blue’essential oils as potential natural herbicides. Molecules 2021, 26, 6749. [Google Scholar] [CrossRef]
- Chu, C.; Mortimer, R.E.; Wang, H.; Wang, Y.; Liu, X.; Yu, S. Allelopathic effects of Eucalyptus on native and introduced tree species. For. Ecol. Manag. 2014, 323, 79–84. [Google Scholar] [CrossRef]
- Kaur, S.; Pal Singh, H.; Batish, D.; Kumar Kholi, R. Role of monoterpenes in Eucalyptus communities. Curr. Bioact. Compd. 2012, 8, 101–107. [Google Scholar] [CrossRef]
- Singh, D.; Kohli, R.K.; Saxena, D.B. Effect of eucalyptus oil on germination and growth of Phaseolus aureus Roxb. Plant Soil 1991, 137, 223–227. [Google Scholar] [CrossRef]
- Costerton, J.W.; Cheng, K.J.; Geesey, G.G.; Ladd, T.I.; Nickel, J.C.; Dasgupta, M.; Marrie, T.J. Bacterial biofilms in nature and disease. Annu. Rev. Microbiol. 1987, 41, 435–464. [Google Scholar] [CrossRef] [PubMed]
- Jamal, M.; Ahmad, W.; Andleeb, S.; Jalil, F.; Imran, M.; Nawaz, M.A.; Hussain, T.; Rafiq, M.; Kamil, M.A. Bacterial biofilm and associated infections. J. Chin. Med. Assoc. 2018, 81, 7–11. [Google Scholar] [CrossRef] [PubMed]
- Husain, F.M.; Ahmad, I.; Khan, M.S.; Ahmad, E.; Tahseen, Q.; Khan, M.S.; Alshabib, N.A. Sub-MICs of Mentha piperita essential oil and menthol inhibits AHL mediated quorum sensing and biofilm of Gram-negative bacteria. Front. Microbiol. 2015, 6, 420. [Google Scholar] [CrossRef]
- Elaissi, A.; Rouis, Z.; Salem, N.A.B.; Mabrouk, S.; ben Salem, Y.; Salah, K.B.H.; Aouni, M.; Farhat, F.; Chemli, R.; Harzallah-Skhiri, F.R.; et al. Chemical composition of 8 Eucalyptus species’ essential oils and the evaluation of their antibacterial, antifungal and antiviral activities. BMC Complement. Altern. Med. 2010, 12, 81. [Google Scholar] [CrossRef]
- Elaissi, A.; Salah, K.H.; Mabrouk, S.; Larbi, K.M.; Chemli, R.; Harzallah-Skhiri, F. Antibacterial activity and chemical composition of 20 Eucalyptus species’ essential oils. Food Chem. 2011, 129, 1427–1434. [Google Scholar] [CrossRef]
- Yong, W.T.L.; Ades, P.K.; Goodger, J.Q.; Bossinger, G.; Runa, F.A.; Sandhu, K.S.; Tibbits, J.F. Using essential oil composition to discriminate between myrtle rust phenotypes in Eucalyptus globulus and Eucalyptus obliqua. Ind. Crops Prod. 2019, 140, 111595. [Google Scholar] [CrossRef]
- Jucá, D.M.; da Silva, M.T.B.; Campos Junior, R.; de Lima, F.J.B.; Okoba, W.; Lahlou, S.; Brandt de Oliveira, R.; Aguiar dos Santos, A.; Magalhães, P.J.C. The essential oil of Eucalyptus tereticornis and its constituents, α-and β-pinene, show accelerative properties on rat gastrointestinal transit. Planta Med. 2011, 77, 57–59. [Google Scholar] [CrossRef]
- Kaur, S.; Singh, H.P.; Batish, D.R.; Kohli, R.K. Chemical characterization and allelopathic potential of volatile oil of Eucalyptus tereticornis against Amaranthus viridis. J. Plant Interact. 2011, 6, 297–302. [Google Scholar] [CrossRef]
- Lima, F.J.; Brito, T.S.; Freire, W.B.; Costa, R.C.; Linhares, M.I.; Sousa, F.C.; Lahlou, S.; Leal-Cardoso, J.H.; Santos, A.A.; Magalhães, P.J. The essential oil of Eucalyptus tereticornis, and its constituents α-and β-pinene potentiate acetylcholine-induced contractions in isolated rat trachea. Fitoterapia 2010, 81, 649–655. [Google Scholar] [CrossRef]
- Miguel, M.G.; Gago, C.; Antunes, M.D.; Lagoas, S.; Faleiro, M.L.; Megías, C.; Cortés-Giraldo, I.; Vioque, J.; Figueiredo, A.C. Antibacterial, antioxidant, and antiproliferative activities of Corymbia citriodora and the essential oils of eight Eucalyptus species. Medicines 2018, 5, 61. [Google Scholar] [CrossRef] [PubMed]
- Shalinder, K.; Harminder, P.S.; Daizy, R.B.; Ravinder, K.K. Chemical characterization, antioxidant and antifungal activity of essential oil from Eucalyptus tereticornis. J. Med. Plants Res. 2011, 5, 4788–4793. [Google Scholar]
- Singh, H.P.; Mittal, S.; Kaur, S.; Batish, D.R.; Kohli, R.K. Characterization and antioxidant activity of essential oils from fresh and decaying leaves of Eucalyptus tereticornis. J. Agric. Food Chem. 2009, 57, 6962–6966. [Google Scholar] [CrossRef] [PubMed]
- Vishwakarma, G.S.; Mittal, S. Bioherbicidal potential of essential oil from leaves of Eucalyptus tereticornis against Echinochloa crus-galli L. J. Biopestic. 2014, 7, 47. [Google Scholar]
- Assareh, M.H.; Jaimand, K.; Rezaee, M.B. Chemical composition of the essential oils of six Eucalyptus species (Myrtaceae) from south west of Iran. J. Essent. Oil Res. 2007, 19, 8–10. [Google Scholar] [CrossRef]
- Safaei-Ghomi, J.; Abbasi-Ahd, A.; Behpour, M.; Batooli, H. Antioxidant activity of the essential oil and metanolic extract of Eucalyptus largiflorens and Eucalyptus intertexta from central Iran. J. Essent. Oil-Bear. Plants 2010, 13, 377–384. [Google Scholar] [CrossRef]
- Sefidkon, F.; Assareh, M.H.; Abravesh, Z.; Mirza, M. Chemical composition of the essential oils of five cultivated Eucalyptus species in Iran: E. intertexta, E. platypus, E. leucoxylon, E. sargentii and E. camaldulensis. J. Essent. Oil-Bear. Plants 2006, 9, 245–250. [Google Scholar] [CrossRef]
- Sefidkon, F.; Assareh, M.H.; Abravesh, Z.; Kandi, M.N.H. Seasonal variation in the essential oil and 1, 8-cineole content of four Eucalyptus species (E. intertexta, E. platypus, E. leucoxylon and E. camaldulensis). J. Essent. Oil-Bear. Plants 2010, 13, 528–539. [Google Scholar] [CrossRef]
- Bignell, C.M.; Dunlop, P.J.; Brophy, J.J. Volatile leaf oils of some south-western and southern Australian species of the genus Eucalyptus. Part XIX. Flavour Fragr. J. 1998, 13, 131–139. [Google Scholar] [CrossRef]
- Ghazghazi, H.; Rssghaier, B.; Riguene, H.; Rigane, G.; El Aloiu, M.; Oueslati, M.A.; Ben Salem, R.; Safdi Zouauoi, N.; Naser, Z.; Laarbi Khouja, M. Phytochemical analysis, antioxidant and antimicrobial activities of Eucalyptus essential oil: A comparative study between Eucalyptus marginata L. and Eucalyptus pauciflora L. Rev. Roum. Chim. 2019, 64, 1055–1062. [Google Scholar] [CrossRef]
- Li, H.; Madden, J.L.; Potts, B.M. Variation in volatile leaf oils of the Tasmanian Eucalyptus species. Subgenus Monocalyptus. Biochem. Syst. Ecol. 1995, 23, 299–318. [Google Scholar] [CrossRef]
- Ibáñez, M.D.; Blázquez, M.A. Phytotoxicity of essential oils on selected weeds: Potential hazard on food crops. Plants 2018, 7, 79. [Google Scholar] [CrossRef] [PubMed]
- Vaughn, S.F.; Spencer, G.F. Volatile monoterpenes as potential parent structures for new herbicides. Weed Sci. 1993, 41, 114–119. [Google Scholar] [CrossRef]
- Abdelgaleil, S.A.; Abdel-Razeek, N.; Soliman, S.A. Herbicidal activity of three sesquiterpene lactones on wild oat (Avena fatua) and their possible mode of action. Weed Sci. 2009, 57, 6–9. [Google Scholar] [CrossRef]
- Kordali, S.; Cakir, A.; Sutay, S. Inhibitory effects of monoterpenes on seed germination and seedling growth. Z. Naturforsch. C 2007, 62, 207–214. [Google Scholar] [CrossRef]
- Javaid, A. Herbicidal potential of allelopathic plants and fungi against Parthenium hysterophorus—A review. Allelopathy J. 2010, 25, 331–334. [Google Scholar]
- Asplund, R.O. Monoterpenes; relation between structure and inhibition of germination. Phytochemistry 1968, 7, 1995–1997. [Google Scholar] [CrossRef]
- De Martino, L.; Mancini, E.; Rolim de Almeida, L.F.; De Feo, V. The antigerminative activity of twenty-seven monoterpenes. Molecules 2010, 15, 6630–6637. [Google Scholar] [CrossRef]
- Vokou, D.; Douvli, P.; Blionis, G.J.; Halley, J.M. Effects of monoterpenoids, acting alone or in pairs, on seed germination and subsequent seedling growth. J. Chem. Ecol. 2003, 29, 2281–2301. [Google Scholar] [CrossRef]
- Abd-ElGawad, A.M.; El Gendy, A.E.N.G.; Assaeed, A.M.; Al-Rowaily, S.L.; Alharthi, A.S.; Mohamed, T.A.; Nassar, M.I.; Dewir, Y.H.; Elshamy, A.I. Phytotoxic effects of plant essential oils: A systematic review and structure-activity relationship based on chemometric analyses. Plants 2020, 10, 36. [Google Scholar] [CrossRef]
- Singh, N.; Singh, H.P.; Batish, D.R.; Kohli, R.K.; Yadav, S.S. Chemical characterization, phytotoxic, and cytotoxic activities of essential oil of Mentha longifolia. Environ. Sci. Pollut. Res. 2020, 27, 13512–13523. [Google Scholar] [CrossRef] [PubMed]
- Hamdi, A.; Majouli, K.; Vander Heyden, Y.; Flamini, G.; Marzouk, Z. Phytotoxic activities of essential oils and hydrosols of Haplophyllum tuberculatum. Ind. Crops Prod. 2017, 97, 440–447. [Google Scholar] [CrossRef]
- Da Silva, E.R.; Igartuburu, J.M.; Overbeck, G.E.; Goncalves Soares, G.L.; Macias, F.A. Are phytotoxic effects of Eucalyptus saligna (Myrtaceae) essential oil related to its major compounds? Aust. J. Bot. 2021, 69, 174–183. [Google Scholar] [CrossRef]
- Zhou, L.; Li, J.; Kong, Q.; Luo, S.; Wang, J.; Feng, S.; Yuan, M.; Chen, T.; Yuan, S.; Ding, C. Chemical composition, antioxidant, antimicrobial, and phytotoxic potential of Eucalyptus grandis × E. urophylla leaves essential oils. Molecules 2021, 26, 1450. [Google Scholar] [CrossRef] [PubMed]
- Muller, C.H. The role of chemical inhibitors (allelopathy) in vegetational composition. Bull. Torrey Bot. Club 1966, 93, 332–351. [Google Scholar] [CrossRef]
- Romagni, J.G.; Duke, S.O.; Dayan, F.E. Inhibition of plant asparagine synthetase by monoterpene cineoles. Plant Physiol. 2000, 123, 725–732. [Google Scholar] [CrossRef][Green Version]
- Barton, F.M.; Dell, B.; Knight, A.R. Herbicide activity of cineole derivatives. J. Agric. Food Chem. 2010, 58, 10147–10155. [Google Scholar] [CrossRef]
- Li, A.; Wu, H.; Feng, Y.; Deng, S.; Hou, A.; Che, F.; Liu, Y.; Geng, Q.; Ni, H.; Wei, Y. A strategy of rapidly screening out herbicidal chemicals from Eucalyptus essential oils. Pest Manag. Sci. 2020, 76, 917–927. [Google Scholar] [CrossRef]
- Nazzaro, F.; Fratianni, F.; De Martino, L.; Coppola, R.; De Feo, V. Effect of essential oils on pathogenic bacteria. Pharmaceuticals 2013, 6, 1451–1474. [Google Scholar] [CrossRef]
- Council of Europe. European Pharmacopeia, 5th ed.; Council of Europe: Strasbourg Cedex, France, 2004. [Google Scholar]
- Adams, R.P. Identification of Essential Oil Components by Gas Chromatography/Mass Spectrometry; Allured Publishing Co.: Carol Stream, IL, USA, 2007. [Google Scholar]
- Davies, N.W. Gas chromatographic retention indices of monoterpenes and sesquiterpenes on methyl silicon and Carbowax 20M phases. J. Chromatogr. A 1990, 503, 1–24. [Google Scholar] [CrossRef]
- Jennings, W.; Shibamoto, T. Qualitative Analysis of Flavour and Fragrance Volatiles by Glass Capillary Gas Chromatography; Academic Press: New York, NY, USA, 1980. [Google Scholar]
- McLafferty, F.W. Wiley Registry of Mass Spectral Data, with NIST Spectral Data CD Rom, 7th ed.; John Wiley & Sons: New York, NY, USA, 1998. [Google Scholar]
- Smeriglio, A.; Trombetta, D.; Cornara, L.; Valussi, M.; De Feo, V.; Caputo, L. Characterization and phytotoxicity assessment of essential oils from plant byproducts. Molecules 2019, 24, 2941. [Google Scholar] [CrossRef] [PubMed]
- Bewley, J.D.; Black, M. Seeds: Physiology of Development and Germination; Plenum Press: New York, NY, USA, 1985. [Google Scholar]
- Sarker, S.D.; Nahar, L.; Kumarasamy, Y. Microtitre plate-based antibacterial assay incorporating resazurin as an indicator of cell growth, and its application in the in vitro antibacterial screening of phytochemicals. Methods 2007, 42, 321–324. [Google Scholar] [CrossRef] [PubMed]
- Fratianni, F.; d’Acierno, A.; Ombra, M.N.; Amato, G.; De Feo, V.; Ayala-Zavala, J.F.; Coppola, R.; Nazzaro, F. Fatty acid composition, antioxidant, and in vitro anti-inflammatory activity of five cold-pressed Prunus seed oils, and their anti-biofilm effect against pathogenic bacteria. Front. Nutr. 2021, 8, 775751. [Google Scholar] [CrossRef] [PubMed]
- Fratianni, F.; Ombra, M.N.; d’Acierno, A.; Caputo, L.; Amato, G.; De Feo, V.; Coppola, R.; Nazzaro, F. Polyphenols content and in vitro α-glycosidase activity of different Italian monofloral honeys, and their effect on selected pathogenic and probiotic bacteria. Microorganisms 2021, 9, 1694. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Compound Name | E. bicostata | E. gigantea | E. intertexa | E. obliqua | E. pauciflora | E. tereticornis | Ki a | Ki b | Identification c | |
|---|---|---|---|---|---|---|---|---|---|---|
| 1 | α-Pinene | 1.7 ± 0.1 | 1.6 ± 0.1 | 6.7 ± 0.3 | 13.2 ± 0.4 | 0.2 ± 0.0 | - | 867 | 1012 | 1,2,3 |
| 2 | Camphene | t | - | 0.1 ± 0.0 | 0.2 ± 0.0 | - | - | 876 | 1075 | 1,2,3 |
| 3 | β-Pinene | t | 0.3 ± 0.0 | 0.2 ± 0.1 | 1.7 ± 0.2 | 0.1 ± 0.1 | - | 902 | 1110 | 1,2,3 |
| 4 | α-Phellandrene | - | - | 0.1 ± 0.1 | 0.7 ± 0.1 | 2.2 ± 0.2 | 1.1 ± 0.2 | 930 | 1177 | 1,2,3 |
| 5 | α-Terpinene | - | 0.1 ± 0.0 | - | 0.1 ± 0.0 | 0.7 ± 0.2 | 0.2± 0.0 | 942 | 1170 | 1,2,3 |
| 6 | p-Cymene | - | - | - | 1.4 0 ± 01 | 7.6 ± 0.3 | 4.6 ± 0.3 | 952 | 1250 | 1,2,3 |
| 7 | β-Phellandrene | - | - | - | - | 2.8 ± 0.2 | - | 954 | 1189 | 1,2,3 |
| 8 | Eucalyptol | 85.5 ± 0.6 | 59.3 ± 0.5 | 65.9 ± 0.5 | 54.9 ± 0.5 | 2.0 ± 0.0 | 2.4 ± 0.1 | 958 | 1210 | 1,2,3 |
| 9 | (E)-β-Ocimene | - | 0.5 ± 0.0 | - | - | - | - | 968 | 1242 | 1,2,3 |
| 10 | (Z)-β-Ocimene | - | - | - | - | 0.1 ± 0.0 | - | 977 | - | 1,2,3 |
| 11 | p-Mentha-2,4(8)-diene | - | - | - | 0.8 ± 0.0 | - | - | 983 | - | 1,2 |
| 12 | γ-Terpinene | - | 1.3 ± 0.1- | - | - | 0.1 ± 0.0 | - | 984 | 1221 | 1,2,3 |
| 13 | cis-Sabinenehydrate | - | 0.2 ± 0.0 | - | - | - | - | 997 | 1115 | 1,2 |
| 14 | Terpinolene | - | 0.5 ± 0.1 | 0.1 ± 0.0 | 0.3 ± 0.0 | 0.3 ± 0.0 | 0.2 ± 0.0 | 1008 | 1267 | 1,2,3 |
| 15 | 6-Camphenone | - | - | 0.1 ± 0.0 | - | - | - | 1011 | - | 1,2 |
| 16 | Linalool | - | 0.5 ± 0.1 | - | - | - | - | 1024 | 1506 | 1,2,3 |
| 17 | endo-Fenchol | - | - | - | - | 4.4 ± 0.5 | - | 1025 | - | 1,2 |
| 18 | 3-Methylbutyl 3-methylbutanoate | - | - | 0.2 ± 0.0 | 0.2 ± 0.0 | - | - | 1029 | 1285 | 1,2 |
| 19 | exo-Fenchol | 0.1 | - | 0.3 ± 0.0 | 0.4 ± 0.0 | - | - | 1031 | 1591 | 1,2 |
| 20 | trans-p-Mentha-2,8-dien-1-ol | - | - | - | 0.1 ± 0.0 | - | - | 1040 | 1639 | 1,2 |
| 21 | cis-p-Menth-2-en-1-ol | - | - | - | - | 2.9 ± 0.6 | 0.2 ± 0.0 | 1041 | - | 1,2 |
| 22 | α-Campholenal | - | 0.1 ± 0.0 | 0.2 ± 0.0 | - | - | 1043 | 1485 | 1,2 | |
| 23 | allo-Ocimene | - | 1.5 ± 0.0 | 0.1 ± 0.2 | 0.3 ± 0.0 | - | - | 1051 | 1388 | 1,2,3 |
| 24 | trans-Pinocarveol | 2.5± 0.2 | 0.5± 0.1 | 4.0 ± 0.2 | 3.3 ± 0.2 | - | 0.1 ± 0.0 | 1057 | 1664 | 1,2 |
| 25 | cis-β-Terpineol | - | - | - | - | - | 0.1 ± 0.0 | 1058 | - | 1,2 |
| 26 | cis-Verbenol | - | - | - | 0.4 ± 0.0 | - | - | 1058 | 1665 | 1,2 |
| 27 | Camphor | - | - | 0.1 ± 0.0 | - | - | - | 1059 | 1491 | 1,2,3 |
| 28 | Citronellal | - | - | 0.1 ± 0.0 | - | - | - | 1063 | 1487 | 1,2 |
| 29 | Sabina ketone | - | 0.4 ± 0.0 | 0.1 ± 0.0 | 0.5 ± 0.0 | - | - | 1067 | 1651 | 1,2 |
| 30 | trans-Pinocamphone | - | - | t | 0.1 ± 0.0 | - | - | 1074 | - | 1,2 |
| 31 | Pinocarvone | 1.6 ± 0.2 | - | 1.1 ± 0.1 | 0.7 ± 0.0 | - | - | 1077 | 1586 | 1,2 |
| 32 | Borneol | 0.2 ± 0.0 | 0.4 ± 0.0 | 0.3 ± 0.0 | 0.6 ± 0.0 | - | - | 1082 | 1715 | 1,2,3 |
| 33 | p-Mentha-1,5-dien-8-ol | - | - | 0.2 ± 0.0 | - | - | - | 1087 | 1670 | 1,2 |
| 34 | Terpinen-4-ol | - | 1.3 ± 0.1 | 0.5 ± 0.0 | - | 3.1 ± 02 | 1.7 ± 0.2 | 1095 | 1590 | 1,2,3 |
| 35 | (E)-iso-Citral | - | - | - | - | 0.6 ± 0.0 | 0.9 ± 0.0 | 1099 | - | 1,2 |
| 36 | cis-Pinocarveol | - | - | 0.3 ± 0.0 | 0.2 ± 0.0 | - | - | 1099 | - | 1,2 |
| 37 | trans-Isocarveol | 0.4 ± 0.0 | - | - | - | - | - | 1099 | 1810 | 1,2 |
| 38 | cis-Dihydrocarvone | - | - | - | 0.8 ± 0.1 | - | 0.2 ± 0.0 | 1101 | - | 1,2 |
| 39 | Dihydrocarveol | 0.5 ± 0.0 | - | 0.2 ± 0.0 | 3.2 ± 0.2 | - | - | 1102 | - | 1,2 |
| 40 | Cryptone | - | 0.4 ± 0.0 | - | - | - | - | 1103 | 1659 | 1,2 |
| 41 | cis-Piperitol | - | - | - | - | 1.1 ± 0.0 | - | 1106 | 1758 | 1,2 |
| 42 | Myrtenol | 0.2 ± 0.0 | - | 0.3 ± 0.0 | - | - | - | 1107 | 1791 | 1,2 |
| 43 | α-Terpineol | 0.1 ± 0.0 | 4.5 ± 0.2 | 0.9 ± 0.1 | 1.2 ± 0.1 | 1.1 ± 0.1 | 0.3 ± 0.0 | 1110 | 1661 | 1,2,3 |
| 44 | Safranal | - | - | - | 0.3 ± 0.0 | - | - | 1117 | 1648 | 1,2 |
| 45 | trans-Piperitol | - | - | - | - | 1.5 ± 0.1 | - | 1120 | 1690 | 1,2 |
| 46 | cis-4-Caranone | - | - | - | 0.1 ± 0.0 | - | - | 1134 | - | 1,2 |
| 47 | cis-Carveol | - | - | 0.1 ± 0.0 | - | - | - | 1135 | 1848 | 1,2 |
| 48 | Verbenone | - | - | - | 0.2 ± 0.0 | - | - | 1143 | 1726 | 1,2 |
| 49 | cis-p-Mentha-1(7),8-dien-2-ol | 0.3 ± 0.0 | 0.2 ± 0.0 | 0.3 ± 0.0 | - | - | - | 1144 | 1896 | 1,2 |
| 50 | Cuminaldehyde | - | - | - | 0.4 ± 0.0 | - | - | 1149 | 1753 | 1,2 |
| 51 | Carvone | - | - | - | 0.2 ± 0.0 | - | - | 1156 | 1736 | 1,2 |
| 52 | exo-Fenchyl acetate | - | - | 0.2 ± 0.0 | - | - | - | 1158 | - | 1,2 |
| 53 | Piperitone | - | 0.9 ± 0.1 | - | 0.1 ± 0.0 | 8.8 ± 0.3 | 19.4 ± 0.5 | 1166 | 1748 | 1,2 |
| 54 | α-Terpinen-7-al | - | 0.4 ± 0.0 | - | - | - | - | 1198 | 1811 | 1,2 |
| 55 | Thymol | - | 0.3 ± 0.0 | - | 0.7 ± 0.1 | - | - | 1218 | 2172 | 1,2,3 |
| 56 | γ-Terpinen-7-al | - | - | - | 0.1 ± 0.0 | - | - | 1236 | - | 1,2 |
| 57 | δ-Elemene | - | - | - | - | 0.7 ± 0.1 | 2.2 ± 0.1 | 1237 | 1479 | 1,2,3 |
| 58 | trans-Verbenyl acetate | - | - | - | 0.2 ± 0.0 | - | - | 1244 | - | 1,2 |
| 59 | 6-camphenol acetate | - | - | 0.1 ± 0.0 | - | - | - | 1245 | - | 1,2 |
| 60 | p-Menth-1-en-9-ol | - | - | - | 0.4 ± 0.0 | - | - | 1252 | - | 1,2 |
| 61 | Copaene | - | 0.1 ± 0.0 | - | - | - | - | 1265 | 1477 | 1,2,3 |
| 62 | α-Cubebene | - | - | - | - | - | 0.1 ± 0.0 | 1270 | 1442 | 1,2 |
| 63 | β-Elemene | - | - | - | - | 0.1 ± 0.0 | 0.2 ± 0.0 | 1290 | - | 1,2,3 |
| 64 | β-Longipinene | - | - | - | - | 0.1 ± 0.0 | 0.3 ± 0.0 | 1298 | - | 1,2 |
| 65 | α-Gurjunene | - | 0.5 ± 0.0 | 1.1 ± 0.1 | - | - | - | 1300 | 1535 | 1,2 |
| 66 | α-Caryophyllene | 0.1 ± 0.0 | - | - | - | - | - | 1307 | 1617 | 1,2 |
| 67 | (Z)-Caryophyllene | - | - | 0.2 ± 0.0 | - | - | - | 1308 | 1617 | 1,2 |
| 68 | Germacrene D | - | - | - | 0.2 ± 0.0 | - | - | 1327 | 1712 | 1,2 |
| 69 | Longifolene | - | - | - | - | 0.8 ± 0.1 | 2.6 ± 0.2 | 1328 | 1574 | 1,2 |
| 70 | Aromadendrene | 0.8 ± 0.0 | - | 0.2± 0.0 | - | 0.6 ± 0.0 | 2.9 ± 0.3 | 1348 | 1631 | 1,2 |
| 71 | allo-Aromadendrene | 0.2 ± 0.0 | - | - | - | 0.1 ± 0.0 | - | 1349 | 1660 | 1,2 |
| 72 | (E)-Caryophyllene | - | 0.2 ± 0.0 | - | - | - | - | 1355 | 1612 | 1,2 |
| 73 | α-Himachalene | - | - | - | - | 0.3 ± 0.0 | - | 1366 | - | 1,2 |
| 74 | 9-epi-(E)-Caryophyllene | - | 0.7 | - | - | 0.4 ± 0.0 | 0.8 ± 0.1 | 1376 | - | 1,2 |
| 75 | cis-β-Guaiene | - | - | - | 0.3 ± 0.0 | - | - | 1383 | - | 1,2 |
| 76 | γ-Gurjunene | - | 0.1 ± 0.0 | 0.4 ± 0.0 | - | 1.2 ± 0.1 | 2.8 ± 0.2 | 1384 | - | 1,2 |
| 77 | α-Vetispirene | - | - | - | - | - | 0.4 ± 0.0 | 1401 | - | 1,2 |
| 78 | γ-Amorphene | - | - | - | - | - | 0.1 | 1408 | - | 1,2 |
| 79 | epi-Cubebol | - | 0.8 ± 0.0 | - | 0.2 ± 0.0 | - | - | 1426 | 1957 | 1,2 |
| 80 | γ-Patchoulene | - | - | - | - | 6.3 ± 0.2 | 4.2 ± 0.2 | 1438 | - | 1,2 |
| 81 | Cubebol | - | - | 1.2 ± 0.1 | - | - | - | 1441 | - | 1,2 |
| 82 | Viridiflorene | - | 0.7 ± 0.0 | 0.1 ± 0.0 | 0.2 ± 0.0 | 0.2 ± 0.0 | 1.2 ± 0.1 | 1448 | - | 1,2 |
| 83 | trans-β-Guaiene | - | - | 0.5 ± 0.0 | - | 2.8 ± 0.3 | 1.0 ± 0.0 | 1449 | - | 1,2 |
| 84 | β-Vetivenene | - | - | - | - | 8.8 | 17.3 | 1463 | - | 1,2 |
| 85 | Viridiflorol | 1.8 | - | - | 0.4 ± 0.0 | - | - | 1464 | 2110 | 1,2 |
| 86 | Globulol | 0.4 | - | 0.9 ± 0.0 | 1.2 ± 0.0 | - | - | 1466 | 2104 | 1,2 |
| 87 | Spathulenol | - | 11.8 ± 0.5 | 8.1 ± 0.3 | 3.8 ± 0.3 | - | - | 1468 | 2127 | 1,2 |
| 88 | Cubeban-11-ol | - | 0.5 ± 0.0 | 0.3 ± 0.0 | 0.2 ± 0.0 | - | - | 1476 | - | 1,2 |
| 89 | trans-Dauca-4(11),7-diene | - | - | - | - | 6.4 ± 0.1 | 17.9 ± 0.3 | 1477 | - | 1,2 |
| 90 | Guaiol | - | - | - | - | 2.5 ± 0.1 | - | 1478 | 2094 | 1,2 |
| 91 | Rosifoliol | - | - | 0.5 ± 0.0 | 0.5 ± 0.0 | - | - | 1483 | - | 1,2 |
| 92 | α-epi-7-epi-5-Eudesmol | - | 1.0 ± 0.0 | - | - | 2.7 ± 0.1 | 4.2 ± 0.1 | 1485 | - | 1,2 |
| 93 | allo-Aromadendreneepoxide | - | - | 0.2 ± 0.0 | - | - | - | 1496 | - | 1,2 |
| 94 | epi-Cedrol | 0.4 ± 0.0 | - | - | 1.5 ± 0.1 | 2.6 ± 0.1 | 1497 | - | 1,2 | |
| 95 | α-Cadinol | - | - | 0.2 ± 0.0 | - | - | - | 1502 | 2224 | 1,2 |
| 96 | γ-Eudesmol | - | 0.4 ± 0.0 | - | 0.5 ± 0.0 | 4.7 ± 0.1 | 2.0 ± 0.1 | 1508 | 2178 | 1,2 |
| 97 | 14-hydroxy-(Z)-Caryophyllene | - | - | 0.1 ± 0.0 | - | - | - | 1513 | - | 1,2 |
| 98 | cis-Cadin-4-en-7-ol | - | 0.8 ± 0.0 | - | - | 1.3 ± 0.1 | - | 1515 | - | 1,2 |
| 99 | β-Eudesmol | 0.3 | 0.8 ± 0.0 | 0.4 ± 0.0 | - | 8.1 ± 0.5 | 2.2 ± 0.1 | 1527 | 2248 | 1,2 |
| 100 | α-Eudesmol | - | 2.0 ± 0.0 | 0.3 ± 0.0 | 0.3 ± 0.0 | 6.3 ± 0.4 | - | 1530 | 2247 | 1,2 |
| 101 | 5-Hydroxy-isobornyl isobutanoate | - | 0.1 ± 0.0 | - | 0.1 ± 0.0 | - | - | 1540 | - | 1,2 |
| 102 | Vulgarone B | - | - | - | - | - | 0.4 ± 0.0 | 1543 | - | 1,2 |
| Total | 96.7 | 96.0 | 97.4 | 96.1 | 95.5 | 96.8 | ||||
| Monoterpene hydrocarbons | 1.7 | 5.8 | 7.3 | 18.7 | 14.0 | 6.1 | ||||
| Oxygenated monoterpenes | 91.4 | 68.9 | 75.0 | 68.6 | 25.5 | 25.3 | ||||
| Sesquiterpene hydrocarbons | 1.1 | 2.3 | 2.5 | 0.7 | 28.8 | 54.0 | ||||
| Oxygenated sesquiterpenes | 2.5 | 18.5 | 12.2 | 7.1 | 27.1 | 11.4 |
| Germinated Seeds | ||||||
| E. bicostata | E. gigantea | E. intertexta | E. obliqua | E.pauciflora | E. tereticornis | |
| Control (H2O) | 8.7 ± 1.2 | 9.7 ± 0.6 | 8.3 ±0.6 | 10.0 ± 0.0 | 9.3 ± 0.6 | 9.9 ± 3.5 |
| 125 µg/mL | 2.0 ± 1.0 **** | 5.7 ± 0.6 **** | 4.0 ± 1.7 * | 8.3 ± 1.2 | 6.7 ± 1.5 **** | 6.9 ± 1.5 |
| 250 µg/mL | 1.7 ± 2.1 **** | 4.7 ± 0.6 **** | 6.3 ± 1.5 | 7.7 ± 0.6 * | 4.7 ± 0.6 **** | 3.0 ± 1.0 |
| 500 µg/mL | 1.0 ± 1.0 **** | 1.3 ± 0.6 **** | 3.0 ± 2.0 *** | 7.3 ± 1.2 * | 1.3 ±0.6 **** | 3.0 ± 1.0 |
| 1000 µg/mL | 0.7 ± 0.6 **** | 0.7 ± 0.6 **** | 0.0 ± 0.0 **** | 6.0 ± 1.0 **** | 0.0 ± 0.0 **** | 0.0 ± 0.0 * |
| Radical Length (cm) | ||||||
| E. bicostata | E. gigantea | E. intertexta | E. obliqua | E.pauciflora | E. tereticornis | |
| Control (H2O) | 3.5 ± 0.4 | 3.1 ± 0.3 | 2.5 ± 1.3 | 4.8 ± 0.3 | 2.6 ± 0.1 | 1.4 ± 1.2 |
| 125 µg/mL | 2.5 ± 1.5 | 2.5 ± 0.1 | 0.9 ± 0.3 * | 3.8 ± 0.4 | 1.7 ± 0.1 **** | 1.8 ± 0.7 |
| 250 µg/mL | 1.3 ± 1.4 * | 2.1 ±0.2 ** | 0.9 ± 0.1 * | 3.8 ± 0.4 | 1.2 ± 0.1 **** | 1.4 ± 1.3 |
| 500 µg/mL | 1.1 ± 0.9 ** | 1.4 ± 0.3 **** | 0.5 ± 0.1 ** | 3.3 ± 1.2 | 1.3 ± 0.2 **** | 0.4 ± 0.3 |
| 1000 µg/mL | 0.8 ± 0.7 ** | 1.0 ± 0.9 **** | 0.0 ± 0.0 **** | 0.8 ± 0.3 | 0.0 ± 0.0 **** | 0.0 ± 0.0 * |
| Germinated Seeds | ||||||
| E. bicostata | E. gigantea | E. intertexta | E. obliqua | E.pauciflora | E. tereticornis | |
| Control (H2O) | 10.0 ± 0.0 | 9.3 ± 0.6 | 9.7 ± 0.6 | 10.0 ± 0.0 | 9.7 ± 0.6 | 9.7 ± 0.6 |
| 125 µg/mL | 8.3 ± 0.6 | 8.0 ± 1.0 | 9.0 ± 1.0 | 1.7 ± 0.6 **** | 7.7 ± 0.6 ** | 4.7 ± 1.5 *** |
| 250 µg/mL | 6.0 ± 2.6 * | 4.3 ± 1.2 **** | 4.0 ± 0.4 *** | 1.7 ± 0.6 **** | 5.0 ± 0.0 **** | 0.7 ± 0.6 **** |
| 500 µg/mL | 0.0 ± 0.0 **** | 1.0 ± 0.0 **** | 3.0 ± 2.6 **** | 0.0 ± 0.0 **** | 1.3 ± 0.6 **** | 0.0 ± 0.0 **** |
| 1000 µg/mL | 0.0 ± 0.0 **** | 0.3 ± 0.6 **** | 0.0 ± 0.0 **** | 0.0 ± 0.0 **** | 1.0 ± 0.0 **** | 0.0 ± 0.0 **** |
| Radical Length (cm) | ||||||
| E. bicostata | E. gigantea | E. intertexta | E. obliqua | E.pauciflora | E. tereticornis | |
| Control (H2O) | 2.8 ± 0.2 | 5.6 ± 0.3 | 3.3 ± 0.6 | 2.5 ± 0.1 | 3.7 ± 0.2 | 3.0 ± 0.7 |
| 125 µg/mL | 1.1 ± 0.2 | 1.1 ± 0.3 **** | 2.1 ± 0.7 | 0.7 ± 0.2 **** | 1.4 ± 0.2 **** | 0.4 ± 0.1 **** |
| 250 µg/mL | 1.0 ± 0.4 | 1.1 ± 0.2 **** | 0.5 ± 0.2 **** | 0.5 ± 0.1 **** | 1.0 ± 0.3 **** | 0.2 ± 0.1 **** |
| 500 µg/mL | 0.0 ± 0.0 *** | 0.5 ± 0.2 **** | 0.6 ± 0.5 **** | 0.0 ± 0.0 **** | 1.6 ± 0.1 **** | 0.0 ± 0.0 **** |
| 1000 µg/mL | 0.0 ± 0.0 *** | 0.1 ± 0.2 **** | 0.0 ± 0.0 **** | 0.0 ± 0.0 **** | 0.7 ± 0.1 **** | 0.0 ± 0.0 **** |
| Germinated Seeds | ||||||
| E. bicostata | E. gigantea | E. intertexta | E. obliqua | E. pauciflora | E. tereticornis | |
| Control (H2O) | 9.0 ± 1.0 | 8.3 ± 0.6 | 10.0 ± 0.0 | 10.0 ± 0.0 | 9.7 ± 0.6 | 7.7 ± 0.6 |
| 125 µg/mL | 8.7 ± 0.6 | 9.3 ± 0.6 | 8.7 ± 0.6 | 9.3 ± 0.6 | 9.7 ± 0.6 | 7.7 ± 1.2 |
| 250 µg/mL | 8.3 ± 1.5 | 9.0 ± 0.0 | 6.3 ± 1.5 * | 8.7 ± 0.6 | 9.0 ± 0.0 | 8.0 ± 1.0 |
| 500 µg/mL | 6.0 ± 1.0 | 7.3 ± 0.6 | 7.0 ± 0.0 | 7.7 ± 0.6 * | 6.0 ± 1.0 **** | 5.3 ± 1.5 |
| 1000 µg/mL | 4.7 ± 4.2 * | 6.7 ± 0.6 * | 5.0 ± 1.0 ** | 6.0 ± 3.0 **** | 0.0 ± 0.0 **** | 0.0 ± 0.0 **** |
| Radical length (cm) | ||||||
| E. bicostata | E. gigantea | E. intertexta | E. obliqua | E. pauciflora | E. tereticornis | |
| Control (H2O) | 3.6 ± 0.4 | 5.1 ± 0.5 | 4.4 ± 0.1 | 3.0 ± 0.3 | 4.8 ± 0.2 | 3.3 ± 0.6 |
| 125 µg/mL | 3.1 ± 1.0 | 2.9 ± 0.3 **** | 3.3 ± 0.4 | 1.6 ± 0.2 *** | 2.7 ± 0.3 **** | 2.2 ± 0.6 |
| 250 µg/mL | 2.1 ± 0.8 | 2.7 ± 0.3 **** | 2.8 ± 0.2 * | 1.3 ± 0.2 **** | 2.6 ± 0.2 **** | 2.3 ± 0.2 |
| 500 µg/mL | 2.0 ± 0.1 | 1.8 ± 0.1 **** | 1.8 ± 0.7 **** | 0.9 ± 0.2 **** | 1.8 ± 0.2 **** | 1.2 ± 0.4 *** |
| 1000 µg/mL | 1.4 ± 1.5 * | 0.6 ± 0.1 **** | 0.5 ± 0.1 **** | 0.4 ± 0.1 **** | 0.7 ± 0.1 **** | 0.0 ± 0.0 **** |
| EO | A. baumannii | E. coli | L. monocytogenes | P. aeruginosa | S. aureus |
|---|---|---|---|---|---|
| E. bicostata | 30 ± 2 | 25 ± 2 | 28 ± 2 * | 30 ± 2 | 28 ± 2 *** |
| E. gigantea | 25 ± 2 ** | 23 ± 1 | 25 ± 2 *** | 28 ± 2 ** | 25 ± 3 *** |
| E. intertexta | 35 ± 2 | 42 ± 1 *** | 25 ± 2 *** | 28 ± 1 ** | 28 ± 2 *** |
| E. obliqua | 33 ± 2 | 42 ± 1 *** | 28 ± 2 * | 30 ± 2 | 28 ± 3 *** |
| E. pauciflora | 25 ± 2 ** | 35 ± 3 *** | 25 ± 2 *** | 30 ± 2 | 35 ± 3 |
| E. teritcornis | 33 ± 3 | 28 ± 1 | 25 ± 2 *** | 28 ± 3 ** | 30 ± 3 *** |
| Tetracycline | 31 ± 2 | 24 ± 2 | 33 ± 1 | 34 ± 1 | 38 ± 1 |
| Time 0 | A. baumannii | E. coli | L. monocytogenes | P. aeruginosa | S. aureus |
|---|---|---|---|---|---|
| E. bicostata 10 µL/mL | 0.00 ± 0.00 | 0.00 ± 0.00 | 54.34 ± 1.25 **** | 0.00 ± 0.00 | 0.00 ± 0.00 |
| E. bicostata 20 µL/mL | 28.74 ± 1.8 **** | 79.61 ± 1.06 **** | 65.62 ± 0.31 **** | 58.74 ± 2.75 **** | 72.55 ± 0.40 **** |
| E. gigantea 10 µL/mL | 79.71 ± 0.14 **** | 79.22 ± 0.06 **** | 82.67 ± 0.10 **** | 78.76 ± 0.07 **** | 77.51 ± 0.11 **** |
| E. gigantea 20 µL/mL | 89.34 ± 0.33 **** | 85.70 ± 0.10 **** | 85.11 ± 0.16 **** | 79.69 ± 0.08 **** | 81.61 ± 0.19 **** |
| E. intertexta 10 µL/mL | 5.99 ± 0.37 **** | 0.00 ± 0.00 | 0.00 ± 0.00 | 75.97 * ± 0.17 **** | 76.20 ± 0.09 **** |
| E. intertexta 20 µL/mL | 63.45 ± 0.21 * | 0.00 ±0.00 | 80.32 ± 0.13 **** | 74.70 ± 0.35 **** | 76.47 ± 0.18 **** |
| E. obliqua 10 µL/mL | 2.96 ± 0.31 *** | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.00 ± 0.00 |
| E. obliqua 20 µL/mL | 61.50 ± 0.28 **** | 0.00 ± 0.00 | 70.02 ± 0.16 **** | 60.21 ± 0.19 **** | 70.42 ± 0.30 **** |
| E. pauciflora 10 µL/mL | 83.92 ± 0.04 **** | 20.54 ± 1.04 **** | 0.00 ± 0.00 | 0.00 ± 0.00 | 2.02 ± 0.84 **** |
| E. pauciflora 20 µL/mL | 86.73 ± 0.02 **** | 46.93 ± 0.23 **** | 79.47 ± 1.35 **** | 13.79 ± 0.83 **** | 31.08 ± 5.81 **** |
| E. tereticornis 10 µL/mL | 5.89 ± 1.46 **** | 0.00 ± 0.00 | 49.84 ± 1.00 **** | 74.17 ± 0.34 **** | 5.46 ± 2.30 **** |
| E. tereticornis 20 µL/mL | 58.73 ± 0.42 **** | 69.57 ± 0.33 **** | 83.72 ± 0.45 **** | 70.90 ± 0.48 **** | 71.80 ± 0.15 **** |
| Time 24 h | A. baumannii | E. coli | L. monocytogenes | P. aeruginosa | S. aureus |
| E. bicostata 10 µL/mL | 62.99 ± 0.91 **** | 34.13 ± 0.71 **** | 56.63 ± 0.84 **** | 51.18 ± 0.45 **** | 46.46 ± 0.79 **** |
| E. bicostata 20 µL/mL | 73.24 ± 0.29 **** | 55.13 ± 1.07 **** | 62.82 ± 0.80 **** | 85.06 ± 1.92 **** | 73.38 ± 0.33 **** |
| E. gigantea 10 µL/mL | 40.81 ± 0.66 **** | 0.00 ± 0.00 | 47.01 ± 0.45 **** | 66.55 ± 0.84 **** | 62.91 ± 0.44 **** |
| E. gigantea 20 µL/mL | 51.94 ± 1.15 **** | 28.62 ± 0.20 **** | 55.44 ± 1.34 **** | 69.79 ± 0.57 **** | 64.78 ± 0.43 **** |
| E. intertexta 10 µL/mL | 32.44 ± 0.68 **** | 40.35 ± 1.53 **** | 67.75 ± 0.52 | 56.20 ± 0.99 | 52.42 ± 0.23 **** |
| E. intertexta 20 µL/mL | 27.92 ± 1.01 **** | 19.04 ± 0.68 **** | 0.00 ± 0.00 | 36.66 ± 0.77 **** | 67.30 ± 0.25 **** |
| E. obliqua 10 µL/mL | 23.17 ± 0.96 **** | 39.82 ± 1.14 **** | 0.00 ± 0.00 | 7.94 ± 0.74 **** | 77.38 ± 0.45 **** |
| E. obliqua 20 µL/mL | 34.96 ± 0.68 **** | 42.56 ± 0.25 **** | 14.52 ± 0.94 **** | 29.79 ± 0.55 **** | 83.63 ± 0.47 **** |
| E. pauciflora 10 µL/mL | 9.71 ± 0.89 **** | 28.60 ± 0.74 **** | 0.00 ± 0.00 | 47.97 ± 1.01 **** | 49.93 ± 0.54 **** |
| E. pauciflora 20 µL/mL | 32.44 ± 1.21 **** | 40.35 ± 1.22 **** | 67.75 ± 0.32 **** | 56.20 ± 0.68 *** | 52.42 ± 0.65 **** |
| E. tereticornis 10 µL/mL | 10.99 ± 2.21 | 8.09 ± 3.84 **** | 38.86 ± 2.88 **** | 13.95 ± 1.33 **** | 0.00 ± 0.00 |
| E. tereticornis 20 µL/mL | 43.88 ± 1.45 | 33.75 ± 2.06 **** | 68.62 ± 2.36 **** | 14.10 ± 1.38 **** | 24.08 * ± 1.18 |
| Time 0 | A. baumannii | E. coli | L. monocytogenes | P. aeruginosa | S. aureus |
|---|---|---|---|---|---|
| E. bicostata 10 µL/mL | 58.01 ± 0.95 **** | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.00 ± 0.00 | 54.96 ± 0.48 **** |
| E. bicostata 20 µL/mL | 60.88 ± 0.70 **** | 0.00 ± 0.00 | 5.15 ± 0.77 ** | 34.26 ± 7.33 **** | 69.26 ± 1.24 **** |
| E. gigantea 10 µL/mL | 83.35 ± 2.78 **** | 75.29 ± 1.34 **** | 80.04 ± 1.64 **** | 79.71 ± 2.04 **** | 79.03 ± 1.56 **** |
| E. gigantea 20 µL/mL | 85.12 ± 2.88 **** | 78.23 ± 1.66 **** | 83.22 ± 1.14 **** | 81.93 ± 2.19 **** | 81.77 ± 1.16 **** |
| E. intertexta 10 µL/mL | 0.00 ± 0.00 | 0.00 ± 0.00 | 79.14 ± 2.05 **** | 75.88 ± 2.59 **** | 78.51 ± 1.58 **** |
| E. intertexta 20 µL/mL | 69.08 ± 0.35 **** | 0.00 ± 0.00 | 79.48 ± 1.70 **** | 78.81 ± 1.78 **** | 80.62 ± 0.84 **** |
| E. obliqua 10 µL/mL | 52.84 ± 1.06 **** | 20.14 ± 1.41 **** | 20.67 ± 2.98 **** | 1.06 ± 1.17 | 11.91 ± 1.37 **** |
| E. obliqua 20 µL/mL | 57.60 ± 0.98 **** | 63.66 ± 1.02 **** | 75.41 ± 1.25 **** | 74.42 ± 0.33 **** | 55.43 ± 1.41 **** |
| E. pauciflora 10 µL/mL | 41.05 ± 1.10 **** | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.00 ± 0.00 | 57.44 ± 0.81 **** |
| E. pauciflora 20 µL/mL | 55.18 ± 0.78 **** | 7.70 ± 1.36 **** | 44.69 ± 1.82 **** | 14.36 ± 2.07 **** | 72.03 ± 1.74 **** |
| E. tereticornis 10 µL/mL | 0.00 ± 0.00 | 0.00 ± 0.00 | 71.13 ± 0.32 **** | 0.00 ± 0.00 | 0.62 ± 1.68 |
| E. tereticornis 20 µL/mL | 35.63 ± 0.67 **** | 62.65 ± 0.40 **** | 88.95 ± 0.25 **** | 83.06 ± 1.60 **** | 85.38 ± 0.18 **** |
| Time 24 h | |||||
| E. bicostata 10 µL/mL | 4.66 ± 3.38 **** | 33.52 ± 0.39 **** | 29.90 ± 1.09 **** | 0.00 ± 0.00 | 30.88 ± 1.52 **** |
| E. bicostata 20 µL/mL | 38.75 ± 1.19 **** | 51.34 ± 0.92 **** | 54.04 ± 0.88 **** | 7.08 ± 0.44 **** | 33.22 ± 0.52 **** |
| E. gigantea 10 µL/mL | 0.00 ± 0.00 | 20.59 ± 0.54 **** | 0.00 ± 0.00 | 0.00 ± 0.00 | 7.95 ± 0.90 **** |
| E. gigantea 20 µL/mL | 0.00 ± 0.00 | 56.68 ± 0.38 **** | 0.00 ± 0.00 | 0.00 ± 0.00 | 21.38 ± 0.64 **** |
| E. intertexta 10 µL/mL | 0.00 ± 0.00 | 31.89 ± 1.35 **** | 0.00 ± 0.00 | 33.95 ± 0.59 **** | 11.46 ± 0.60 **** |
| E. intertexta 20 µL/mL | 12.96 ± 1.72 **** | 34.32 ± 1.03 **** | 0.00 ± 0.00 | 0.00 ± 0.00 | 18.77 ± 1.13 **** |
| E. obliqua 10 µL/mL | 0.00 ± 0.00 | 10.32 ± 0.99 **** | 0.00 ± 0.00 | 0.00 ± 0.00 | 18.43 ± 1.34 **** |
| E. obliqua 20 µL/mL | 19.87 ± 1.85 **** | 27.08 ± 1.03 **** | 0.00 ± 0.00 | 9.73 ± 0.61 **** | 30.54 ± 1.11 **** |
| E. pauciflora 10 µL/mL | 0.00 ± 0.00 | 10.32 ± 0.94 **** | 0.00 ± 0.00 | 0.00 ± 0.00 | 18.43 ± 1.11 **** |
| E. pauciflora 20 µL/mL | 88.39 ± 1.07 **** | 27.08 ± 0.82 **** | 0.00 ± 0.00 | 9.73 ± 0.71 **** | 30.54 ± 0.81 **** |
| E. tereticornis 10 µL/mL | 22.79 ± 1.12 **** | 0.00 ± 0.00 | 26.68 ± 1.03 **** | 44.15 ± 0.12 **** | 25.62 ± 0.35 **** |
| E. tereticornis 20 µL/mL | 34.69 ± 0.23 **** | 65.92 ± 0.87 **** | 29.26 ± 0.06 **** | 45.63 ± 0.28 **** | 64.24 ± 1.15 **** |
| Arboretum (Governorate) | Harvest Period | Bioclimatic Conditions | Yield (%) | |
|---|---|---|---|---|
| E. bicostata | Choucha (Bizerte) | March 2021 | Upper humid | 1.40 |
| E. gigantea | Zerniza (Bizerte) | July 2021 | Upper humid | 0.20 |
| E. intertexta | Djebel Manasour (Zaghouen) | May 2021 | Upper and middle semi-arid | 0.55 |
| E. obliqua | HenchirNaam (Siliana) | April 2021 | Upper and middle semi-arid | 3.11 |
| E. pauciflora | Zerniza (Bizerte) | July 2021 | Upper humid | 0.10 |
| E. tereticornis | Zerniza (Bizerte) | July 2021 | Upper humid | 0.03 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Polito, F.; Kouki, H.; Khedhri, S.; Hamrouni, L.; Mabrouk, Y.; Amri, I.; Nazzaro, F.; Fratianni, F.; De Feo, V. Chemical Composition and Phytotoxic and Antibiofilm Activity of the Essential Oils of Eucalyptus bicostata, E. gigantea, E. intertexta, E. obliqua, E. pauciflora and E. tereticornis. Plants 2022, 11, 3017. https://doi.org/10.3390/plants11223017
Polito F, Kouki H, Khedhri S, Hamrouni L, Mabrouk Y, Amri I, Nazzaro F, Fratianni F, De Feo V. Chemical Composition and Phytotoxic and Antibiofilm Activity of the Essential Oils of Eucalyptus bicostata, E. gigantea, E. intertexta, E. obliqua, E. pauciflora and E. tereticornis. Plants. 2022; 11(22):3017. https://doi.org/10.3390/plants11223017
Chicago/Turabian StylePolito, Flavio, Habiba Kouki, Sana Khedhri, Lamia Hamrouni, Yassine Mabrouk, Ismail Amri, Filomena Nazzaro, Florinda Fratianni, and Vincenzo De Feo. 2022. "Chemical Composition and Phytotoxic and Antibiofilm Activity of the Essential Oils of Eucalyptus bicostata, E. gigantea, E. intertexta, E. obliqua, E. pauciflora and E. tereticornis" Plants 11, no. 22: 3017. https://doi.org/10.3390/plants11223017
APA StylePolito, F., Kouki, H., Khedhri, S., Hamrouni, L., Mabrouk, Y., Amri, I., Nazzaro, F., Fratianni, F., & De Feo, V. (2022). Chemical Composition and Phytotoxic and Antibiofilm Activity of the Essential Oils of Eucalyptus bicostata, E. gigantea, E. intertexta, E. obliqua, E. pauciflora and E. tereticornis. Plants, 11(22), 3017. https://doi.org/10.3390/plants11223017