
Photosynthesis, Phytohormone Signaling and Sugar Catabolism in the Culm Sheaths of Phyllostachys edulis


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Site
2.2. Measurement of Gas Exchange Parameters and Sampling for Physiological Analysis
2.3. Chlorophyll Fluorescence Measurements
2.4. Measurement of Photosynthetic Pigments
2.5. Anatomical Structure
2.6. Sucrose, Glucose, and Fructose Content Determination
2.7. Transcriptome Sequencing and Analysis
2.8. qRT-PCR Verification
2.9. Statistical Analysis
3. Results
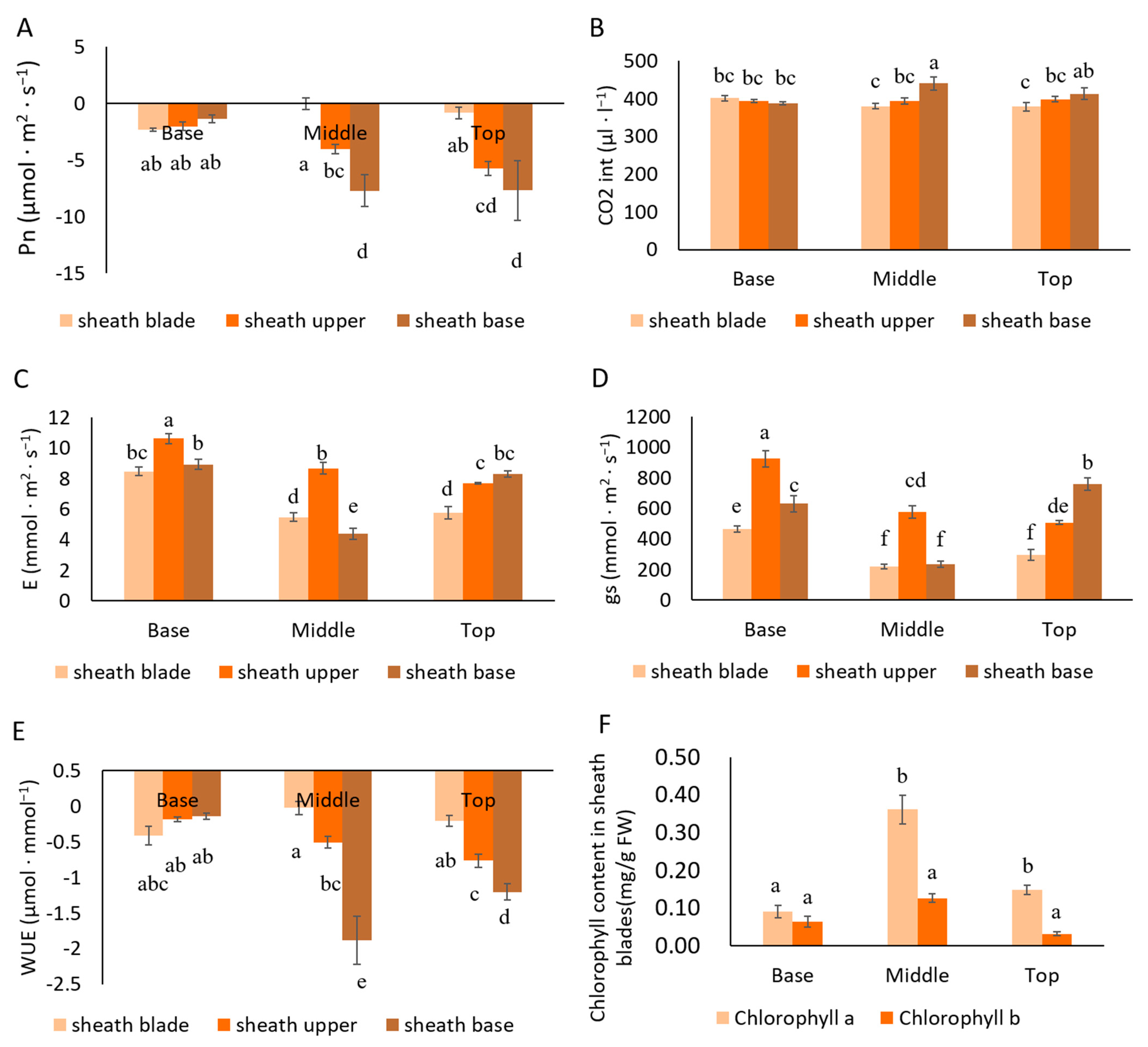
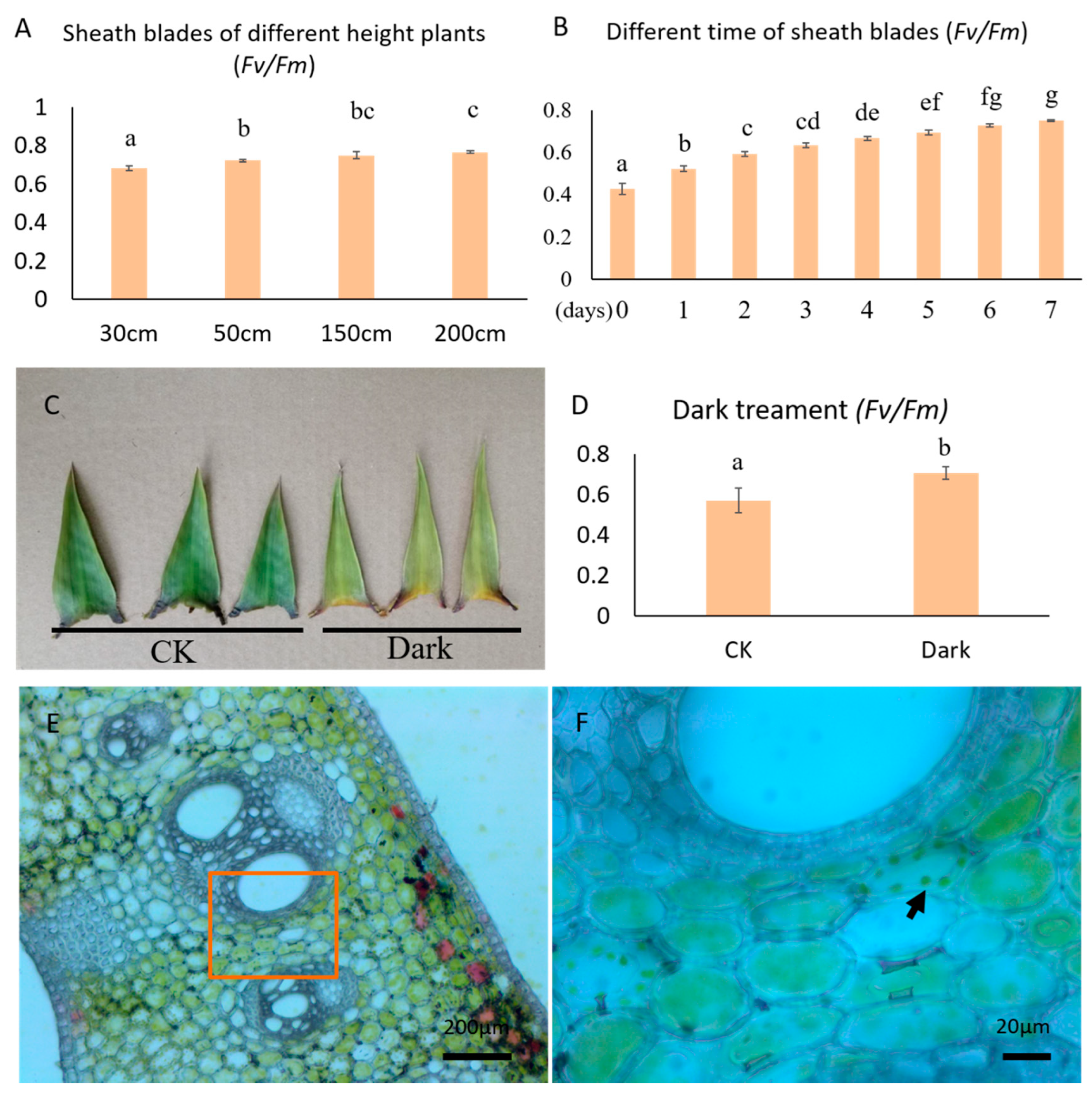
3.1. Changes in the Photosynthetic Capacity of Sheath Blades
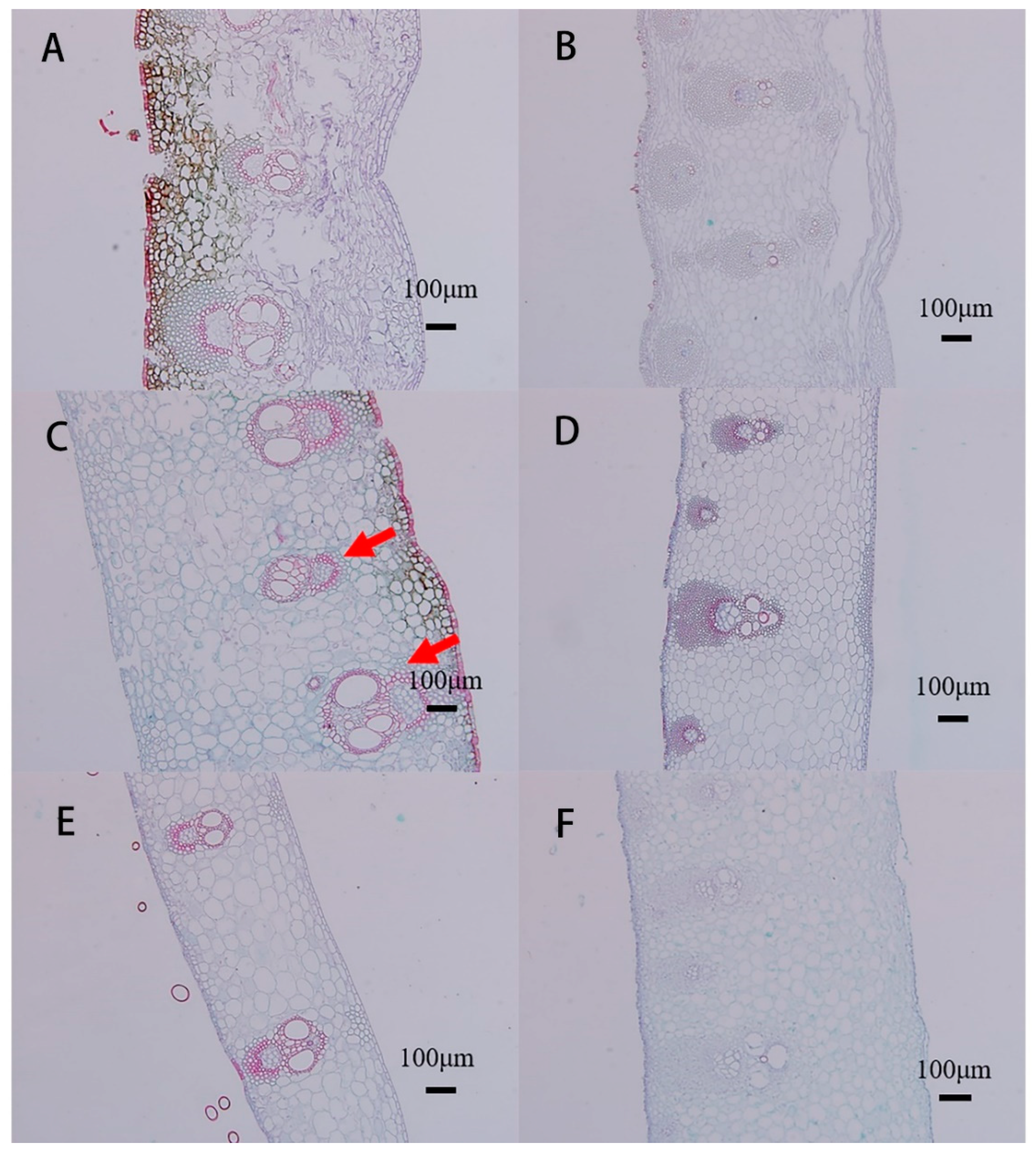
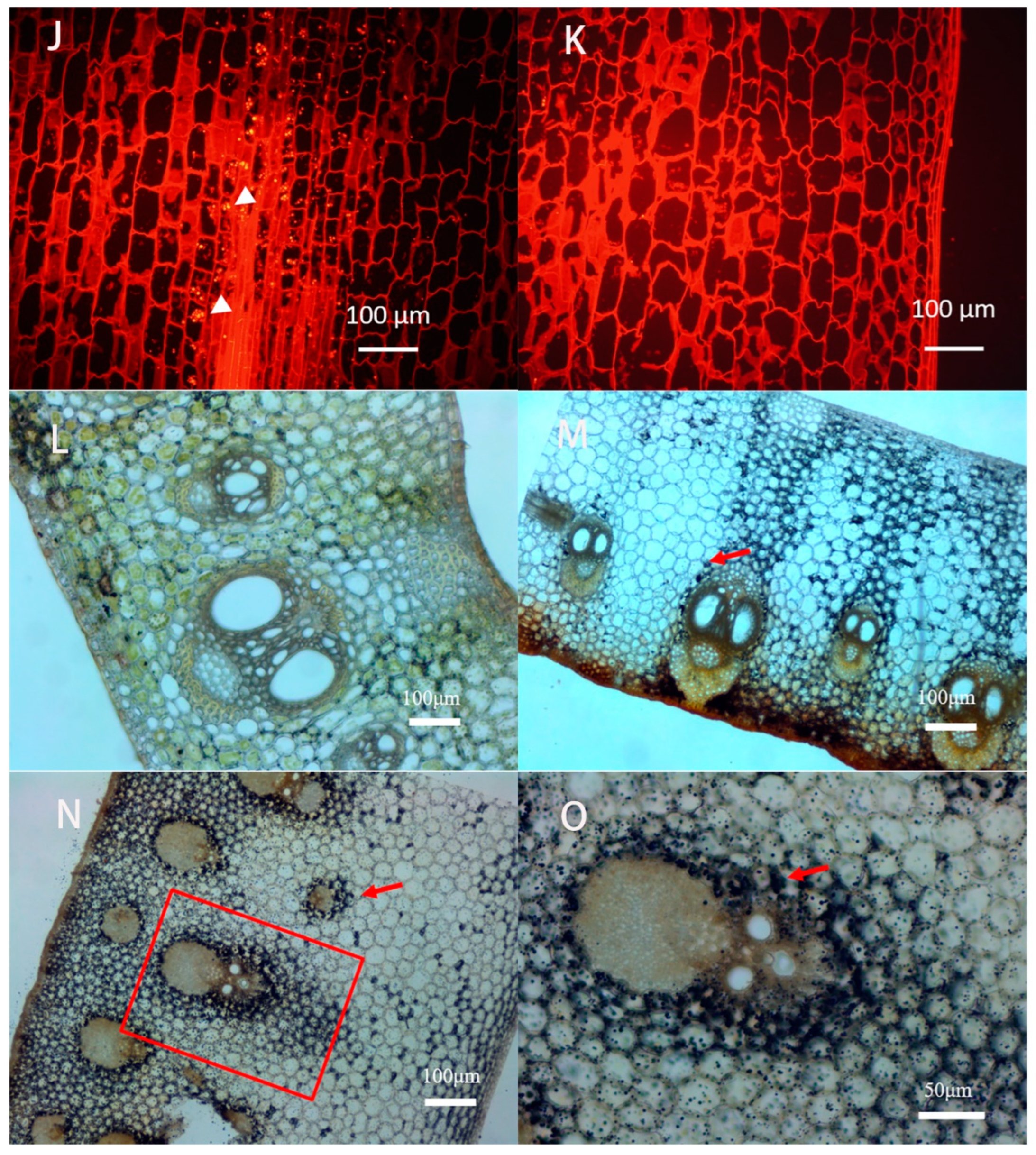
3.2. Anatomical Characteristics of Culm Sheaths
3.3. Sucrose, Glucose, and Fructose Content and Starch Distribution in Culm Sheaths
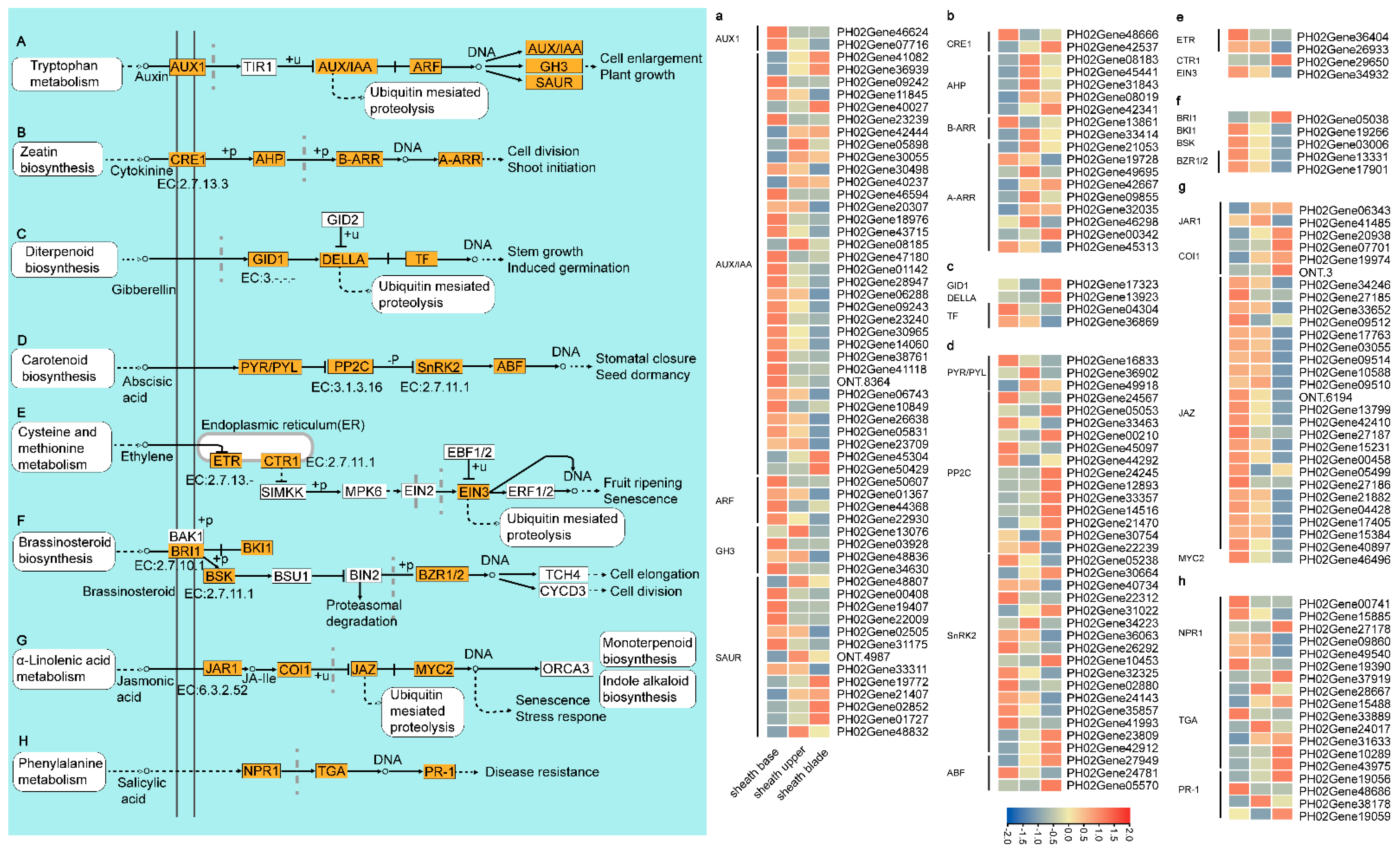
3.4. Active Phytohormone Signaling and Sugar Catabolism in the Sheath Base
4. Discussion
4.1. Sheath Blades Are Capable of Photosynthesis and This Ability Develops Gradually
4.2. Sheath Base Stores Starch and Is the Main Site of Respiration and Sugar Metabolism
4.3. Plant Hormones Play an Important Role in Culm Sheaths
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Peng, Z.; Lu, Y.; Li, L.; Zhao, Q.; Feng, Q.; Gao, Z.; Lu, H.; Hu, T.; Yao, N.; Liu, K.; et al. The draft genome of the fast-growing non-timber forest species moso bamboo (Phyllostachys heterocycla). Nat. Genet. 2013, 45, 456–461. [Google Scholar] [CrossRef] [PubMed]
- Peng, Z.; Zhang, C.; Zhang, Y.; Hu, T.; Mu, S.; Li, X.; Gao, J. Transcriptome sequencing and analysis of the fast growing shoots of moso bamboo (Phyllostachys edulis). PLoS ONE 2013, 8, e78944. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Lu, J.; Zhang, X. Spatiotemporal trend of carbon storage in China’s bamboo industry, 1993–2018. J. Environ. Manag. 2022, 314, 114989. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Cheng, Z.; Ma, Y.; Bai, Q.; Li, X.; Cao, Z.; Wu, Z.; Gao, J. The association of hormone signalling genes, transcription and changes in shoot anatomy during moso bamboo growth. Plant Biotechnol. J. 2018, 16, 72–85. [Google Scholar] [CrossRef]
- Xu, M.; Ji, H.; Zhuang, S. Carbon stock of Moso bamboo (Phyllostachys pubescens) forests along a latitude gradient in the subtropical region of China. PLoS ONE 2018, 13, e0193024. [Google Scholar] [CrossRef]
- Song, X.; Peng, C.; Zhou, G.; Gu, H.; Li, Q.; Zhang, C. Dynamic allocation and transfer of non-structural carbohydrates, a possible mechanism for the explosive growth of Moso bamboo (Phyllostachys heterocycla). Sci. Rep. 2016, 6, 25908. [Google Scholar] [CrossRef]
- Wei, Q.; Shi, P. Investigating the Shape of the Shoot Apical Meristem in Bamboo Using a Superellipse Equation. Bio-Protocol 2017, 7, e2644. [Google Scholar] [CrossRef]
- Singh, K.; Gupta, I.; Gupta, S. Classification of Bamboo Species by Fourier and Legendre Moment. Int. J. Adv. Sci. Technol. 2013, 50, 61–70. [Google Scholar]
- Wang, S.; He, W.; Zhan, H. Culm sheaths affect height growth of bamboo shoots in Fargesia yunnanensis. Braz. J. Bot. 2018, 41, 255–266. [Google Scholar] [CrossRef]
- Zheng, H.; Cai, M.; Bai, Y.; Xu, J.; Xie, Y.; Song, H.; Li, J.; Gao, J. The Effect of Guttation on the Growth of Bamboo Shoots. Forests 2021, 13, 31. [Google Scholar] [CrossRef]
- Wei, Q.; Guo, L.; Jiao, C.; Fei, Z.; Chen, M.; Cao, J.; Ding, Y.; Yuan, Q. Characterization of the developmental dynamics of the elongation of a bamboo internode during the fast growth stage. Tree Physiol. 2019, 39, 1201–1214. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Zhan, H.; Li, P.; Chu, C.; Li, J.; Wang, C. Physiological Mechanism of Internode Bending Growth after the Excision of Shoot Sheath in Fargesia yunnanensis and Its Implications for Understanding the Rapid Growth of Bamboos. Front. Plant Sci. 2020, 11, 418. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Pei, J.; Li, J.; Tang, G.; Zhao, J.; Peng, X.; Nie, S.; Ding, Y.; Wang, C. Sucrose and starch metabolism during Fargesia yunnanensis shoot growth. Physiol. Plant. 2020, 168, 188–204. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Chen, T.H.; Liu, E.U.; Liu, C.P. Accessing the nursing behaviour of Moso bamboo (Phyllostachys edilus) on carbohydrates dynamics and photosystems. Sci. Rep. 2020, 10, 1015. [Google Scholar] [CrossRef] [PubMed]
- Scofield, G.N.; Ruuska, S.A.; Aoki, N.; Lewis, D.C.; Tabe, L.M.; Jenkins, C.L. Starch storage in the stems of wheat plants: Localization and temporal changes. Ann. Bot. 2009, 103, 859–868. [Google Scholar] [CrossRef]
- Miyake, H. Starch Accumulation in the Bundle Sheaths of C3 Plants: A Possible Pre-Condition for C4 Photosynthesis. Plant Cell Physiol. 2016, 57, 890–896. [Google Scholar] [CrossRef]
- Ionuț, R.; Rozalia, K.; Diana, H.; Marius, B.; Florin, D.B.; Camelia, U.; Adina, T. Evaluation of Photosynthesis Capacity of Some Winter Wheat Genotypes in Transylvanian Plain Conditions. Bull. USAMV Ser. Agric. 2018, 75, 56–57. [Google Scholar]
- Brestic, M.; Zivcak, M.; Hauptvogel, P.; Misheva, S.; Kocheva, K.; Yang, X.; Li, X.; Allakhverdiev, S.I. Wheat plant selection for high yields entailed improvement of leaf anatomical and biochemical traits including tolerance to non-optimal temperature conditions. Photosynth. Res. 2018, 136, 245–255. [Google Scholar] [CrossRef]
- Harmut, A. Chlorophylls and carotenoids: Pigments of photosynthetic membranes. Methods Enzymol. 1987, 148, 350–383. [Google Scholar]
- Hu, W.; Loka, D.A.; Fitzsimons, T.R.; Zhou, Z.; Oosterhuis, D.M. Potassium deficiency limits reproductive success by altering carbohydrate and protein balances in cotton (Gossypium hirsutum L.). Environ. Exp. Bot. 2018, 145, 87–94. [Google Scholar] [CrossRef]
- Fan, C.; Ma, J.; Guo, Q.; Li, X.; Wang, H.; Lu, M. Selection of reference genes for quantitative real-time PCR in bamboo (Phyllostachys edulis). PLoS ONE 2013, 8, e56573. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Ribeiro, R.V.; Machado, E.C.; Santos, M.G.; Oliveira, R.F. Seasonal and diurnal changes in photosynthetic limitation of young sweet orange trees. Environ. Exp. Bot. 2009, 66, 203–211. [Google Scholar] [CrossRef]
- Zha, T.; Ryyppo, A.; Wang, K.Y.; Kellomaki, S. Effects of elevated carbon dioxide concentration and temperature on needle growth, respiration and carbohydrate status in field-grown Scots pines during the needle expansion period. Tree Physiol. 2001, 21, 1279–1287. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Xu, C.Y.; Schuster, W.S.; Griffin, K.L. Seasonal variation of temperature response of respiration in invasive Berberis thunbergii (Japanese barberry) and two co-occurring native understory shrubs in a northeastern US deciduous forest. Oecologia 2007, 153, 809–819. [Google Scholar] [CrossRef]
- Sawicki, M.; Ait Barka, E.; Clement, C.; Gilard, F.; Tcherkez, G.; Baillieul, F.; Vaillant-Gaveau, N.; Jacquard, C. Cold-night responses in grapevine inflorescences. Plant Sci. 2015, 239, 115–127. [Google Scholar] [CrossRef] [PubMed]
- Way, D.A.; Aspinwall, M.J.; Drake, J.E.; Crous, K.Y.; Campany, C.E.; Ghannoum, O.; Tissue, D.T.; Tjoelker, M.G. Responses of respiration in the light to warming in field-grown trees: A comparison of the thermal sensitivity of the Kok and Laisk methods. New Phytol. 2019, 222, 132–143. [Google Scholar] [CrossRef] [PubMed]
- Ishimaru, K.; Kosone, M.; Sasaki, H.; Kashiwagi, T. Leaf contents differ depending on the position in a rice leaf sheath during sink—Source transition. Plant Physiol. Biochem. 2004, 42, 855–860. [Google Scholar] [CrossRef] [PubMed]
- Ruan, Y.L. Sucrose metabolism: Gateway to diverse carbon use and sugar signaling. Annu. Rev. Plant Biol. 2014, 65, 33–67. [Google Scholar] [CrossRef] [PubMed]
- Ding, Y.L.; Walter, L. On the nodal structure of bamboo. J. Bamboo Res. 1995, 14, 24–32. [Google Scholar]
- Dahiya, A.; Saini, R.; Saini, H.S.; Devi, A. Sucrose metabolism: Controls the sugar sensing and generation of signalling molecules in plants. J. Pharmacogn. Phytochem. 2017, 6, 1563–1572. [Google Scholar]
- Xiong, Y.; McCormack, M.; Li, L.; Hall, Q.; Xiang, C.; Sheen, J. Glucose-TOR signalling reprograms the transcriptome and activates meristems. Nature 2013, 496, 181–186. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Cai, W.; Liu, Y.; Li, H.; Fu, L.; Liu, Z.; Xu, L.; Liu, H.; Xu, T.; Xiong, Y. Differential TOR activation and cell proliferation in Arabidopsis root and shoot apexes. Proc. Natl. Acad. Sci. USA 2017, 114, 2765–2770. [Google Scholar] [CrossRef]
- Cho, Y.H.; Yoo, S.D.; Sheen, J. Regulatory Functions of Nuclear Hexokinase1 Complex in Glucose Signaling. Cell 2006, 127, 579–589. [Google Scholar] [CrossRef] [PubMed]
- Chen, M.; Ju, Y.; Ahmad, Z.; Yin, Z.; Ding, Y.; Que, F.; Yan, J.; Chu, J.; Wei, Q. Multi-analysis of sheath senescence provides new insights into bamboo shoot development at the fast growth stage. Tree Physiol. 2020, 41, 491–507. [Google Scholar] [CrossRef] [PubMed]
- Bai, Y.; Cai, M.; Mu, C.; Cheng, W.; Zheng, H.; Cheng, Z.; Li, J.; Mu, S.; Gao, J. New Insights Into the Local Auxin Biosynthesis and Its Effects on the Rapid Growth of Moso Bamboo (Phyllostachys edulis). Front. Plant Sci. 2022, 13, 858686. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Yang, X.; Cheng, L.; Guo, Z.; Wang, H.; Wu, W.; Shin, K.; Zhu, J.; Zheng, X.; Bian, J.; et al. Physiological and transcriptomic analyses of brassinosteroid function in moso bamboo (Phyllostachys edulis) seedlings. Planta 2020, 27, 252. [Google Scholar] [CrossRef] [PubMed]
- Gao, J. The Moso Bamboo Genome; Springer Cham: Berlin/Heidelberg, Germany, 2021; p. 208. [Google Scholar]
- Hou, D.; Bai, Q.; Li, J.; Xie, L.; Li, X.; Cheng, Z.; Gao, J. The Gibberellic Acid-Stimulated Transcript Gene Family in Moso Bamboo: A Genome-Wide Survey and Expression Profiling during Development and Abiotic Stresses. J. Plant Growth Regul. 2018, 37, 1135–1147. [Google Scholar] [CrossRef]
- Retzer, K.; Weckwerth, W. The TOR-Auxin Connection Upstream of Root Hair Growth. Plants 2021, 10, 150. [Google Scholar] [CrossRef] [PubMed]
- Cui, K.; He, C.Y.; Zhang, J.G.; Duan, A.G.; Zeng, Y.F. Temporal and spatial profiling of internode elongation—Associated protein expression in rapidly growing culms of bamboo. J. Proteome Res. 2012, 11, 2492–2507. [Google Scholar] [CrossRef]








Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zheng, H.; Bai, Y.; Li, X.; Song, H.; Cai, M.; Cheng, Z.; Mu, S.; Li, J.; Gao, J. Photosynthesis, Phytohormone Signaling and Sugar Catabolism in the Culm Sheaths of Phyllostachys edulis. Plants 2022, 11, 2866. https://doi.org/10.3390/plants11212866
Zheng H, Bai Y, Li X, Song H, Cai M, Cheng Z, Mu S, Li J, Gao J. Photosynthesis, Phytohormone Signaling and Sugar Catabolism in the Culm Sheaths of Phyllostachys edulis. Plants. 2022; 11(21):2866. https://doi.org/10.3390/plants11212866
Chicago/Turabian StyleZheng, Huifang, Yucong Bai, Xiangyu Li, Huajian Song, Miaomiao Cai, Zhanchao Cheng, Shaohua Mu, Juan Li, and Jian Gao. 2022. "Photosynthesis, Phytohormone Signaling and Sugar Catabolism in the Culm Sheaths of Phyllostachys edulis" Plants 11, no. 21: 2866. https://doi.org/10.3390/plants11212866
APA StyleZheng, H., Bai, Y., Li, X., Song, H., Cai, M., Cheng, Z., Mu, S., Li, J., & Gao, J. (2022). Photosynthesis, Phytohormone Signaling and Sugar Catabolism in the Culm Sheaths of Phyllostachys edulis. Plants, 11(21), 2866. https://doi.org/10.3390/plants11212866