
Genome-Wide Analysis of Multiple Organellar RNA Editing Factor (MORF) Family in Kiwifruit (Actinidia chinensis) Reveals Its Roles in Chloroplast RNA Editing and Pathogens Stress

 and
and
Abstract
:1. Introduction
2. Results
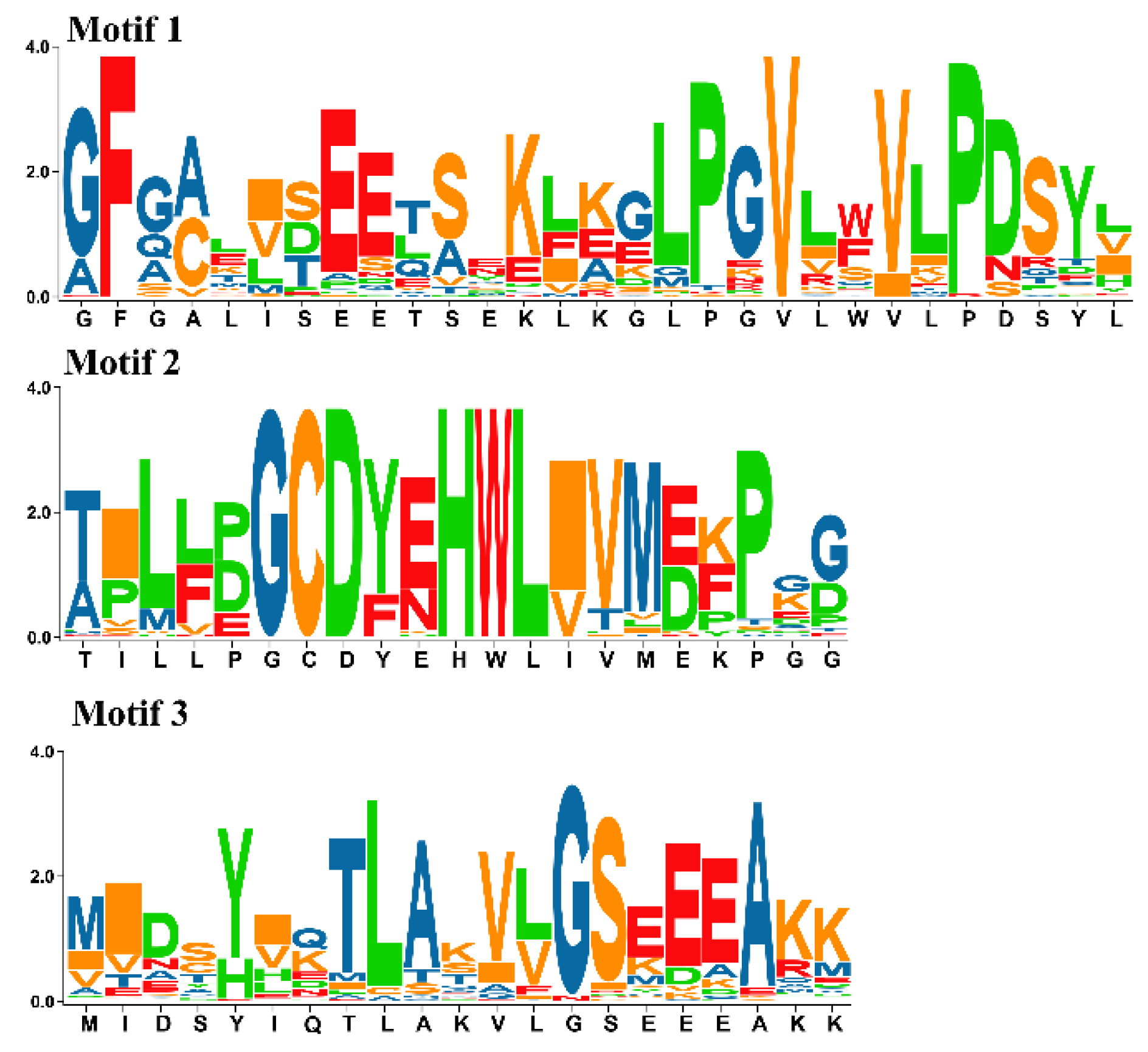
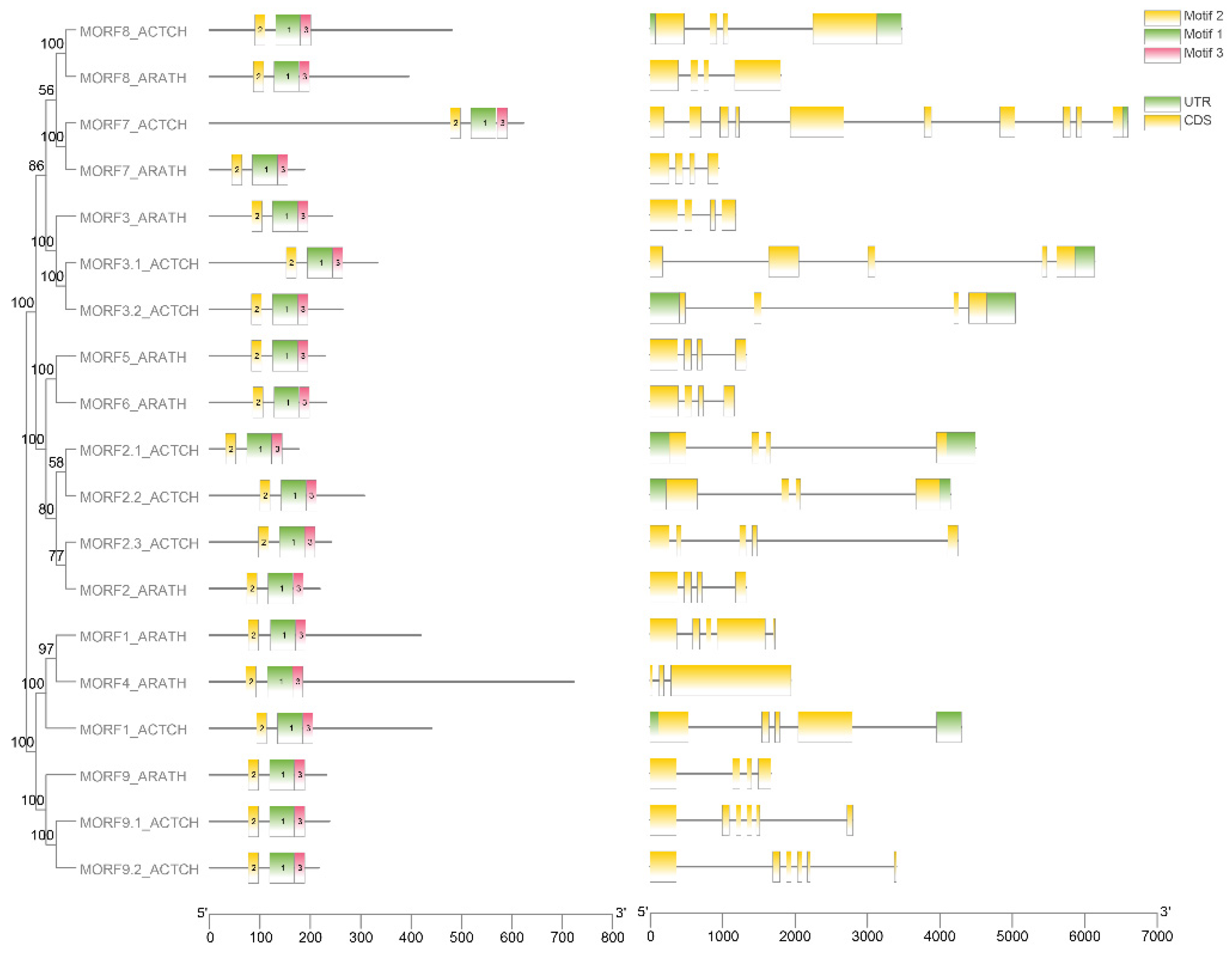
2.1. Identification of MORF Gene Family Members in Kiwifruit
2.2. Phylogenetic Analysis of MORF Genes
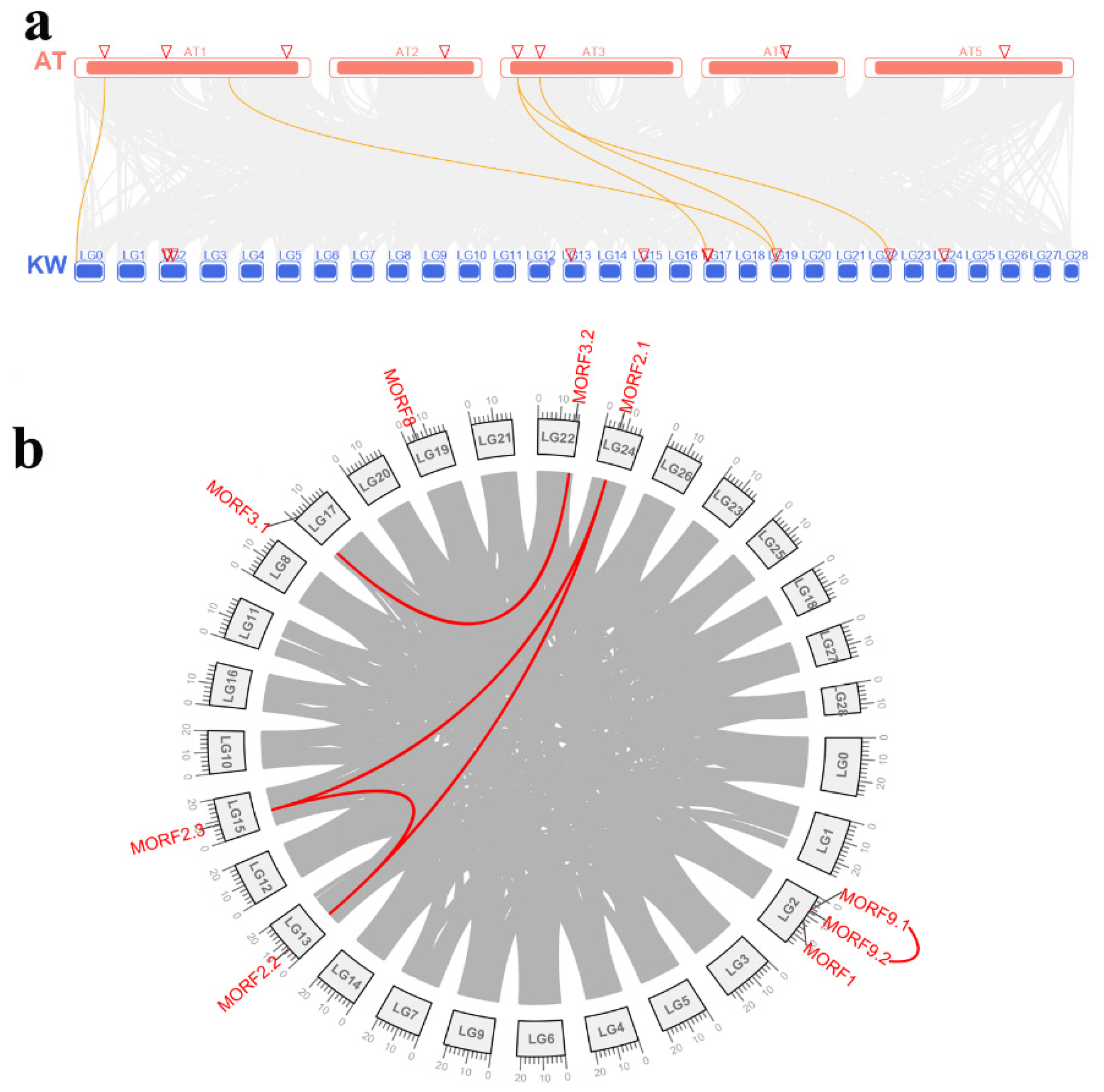
2.3. Genome Distribution and Synteny Analysis of MORF Genes in Kiwifruit
2.4. Identification of RNA Editing Sites in Kiwifruit
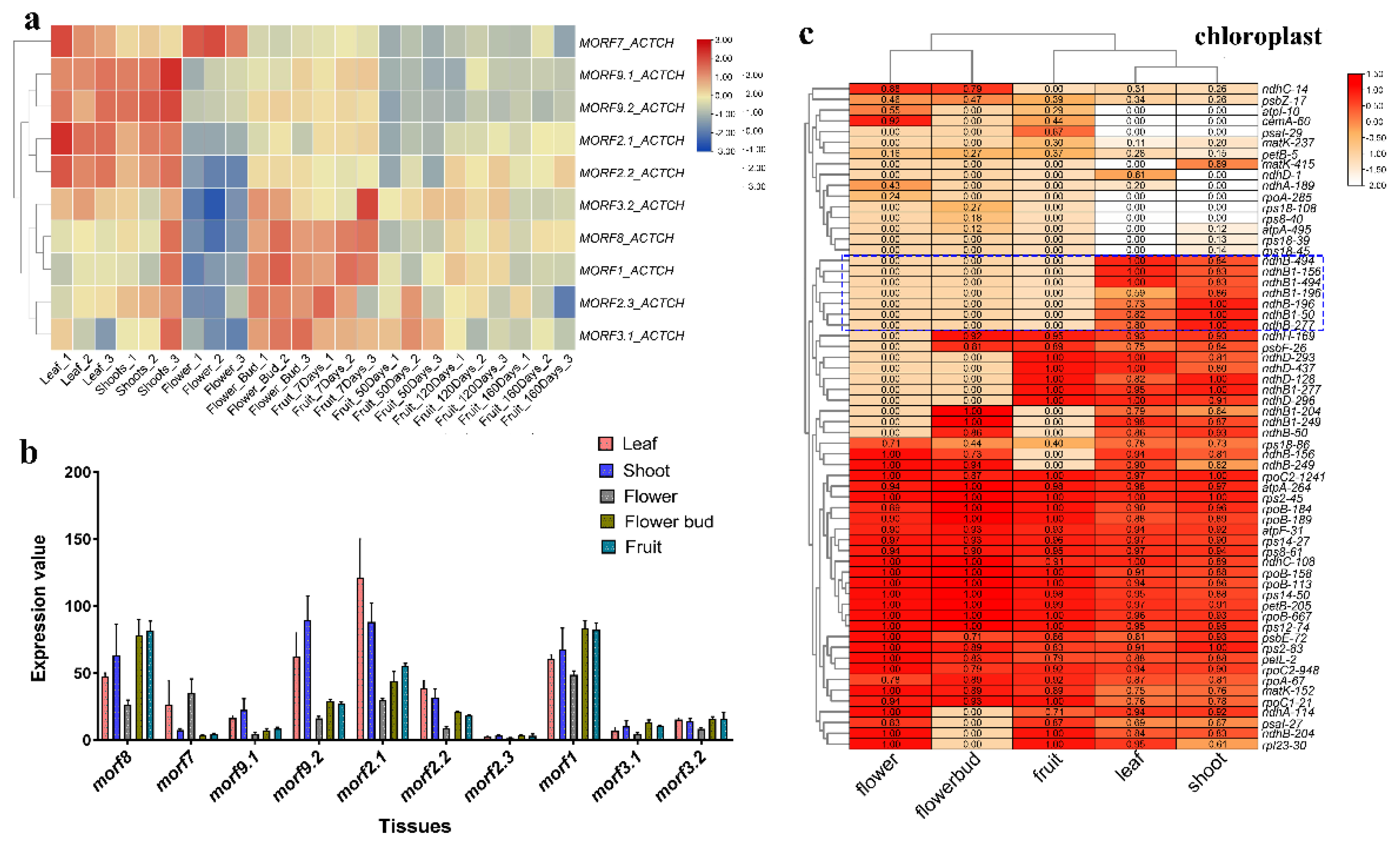
2.5. Profiles of MORF Gene Expression and RNA Editing Patterns in Different Tissues of Kiwifruit
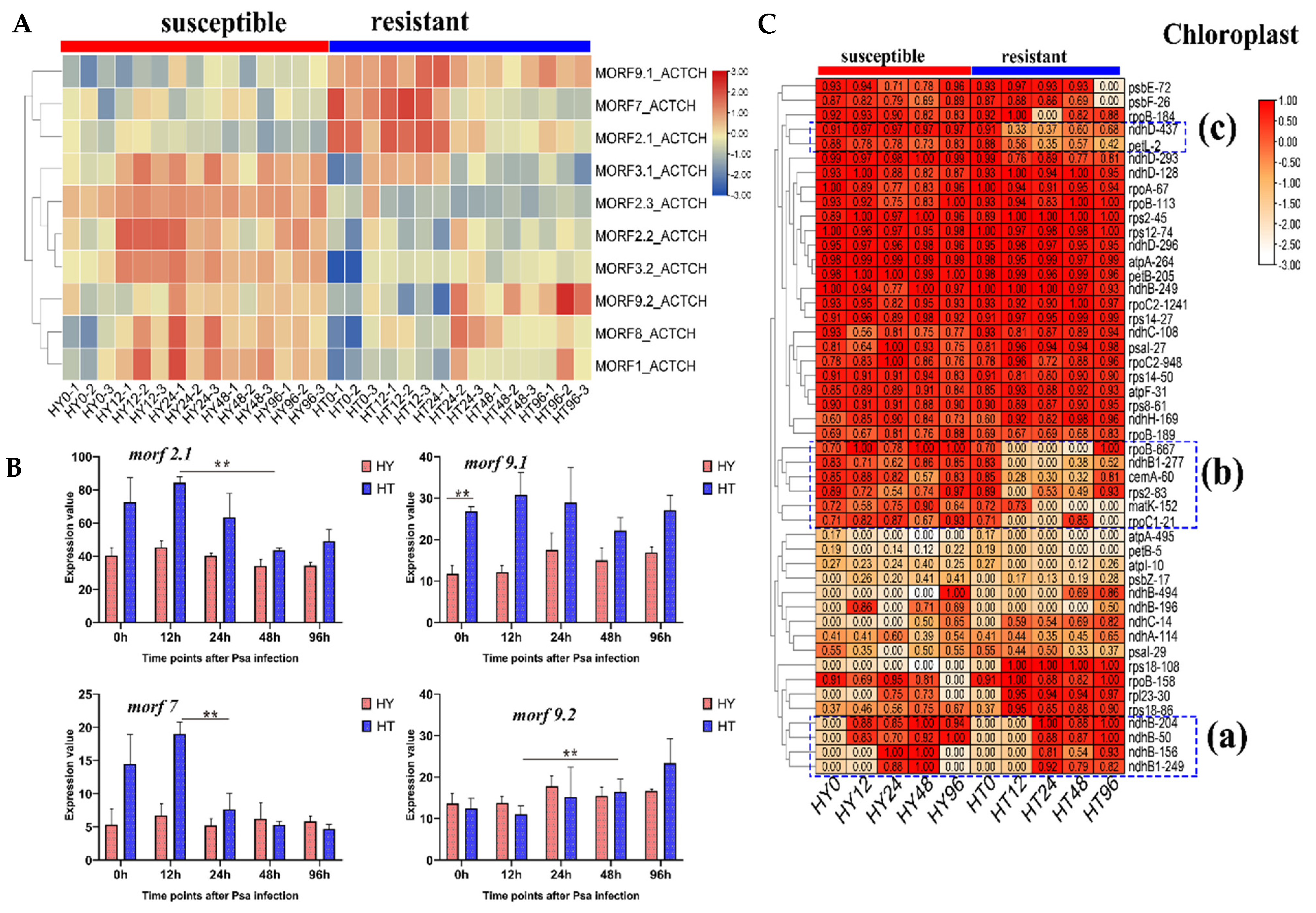
2.6. Different MORF Genes Expression in Response to Psa Infection between Resistant and Susceptible Kiwifruits
2.7. Different RNA Editing Patterns in Response to Psa Infection between Resistant and Susceptible Kiwifruits
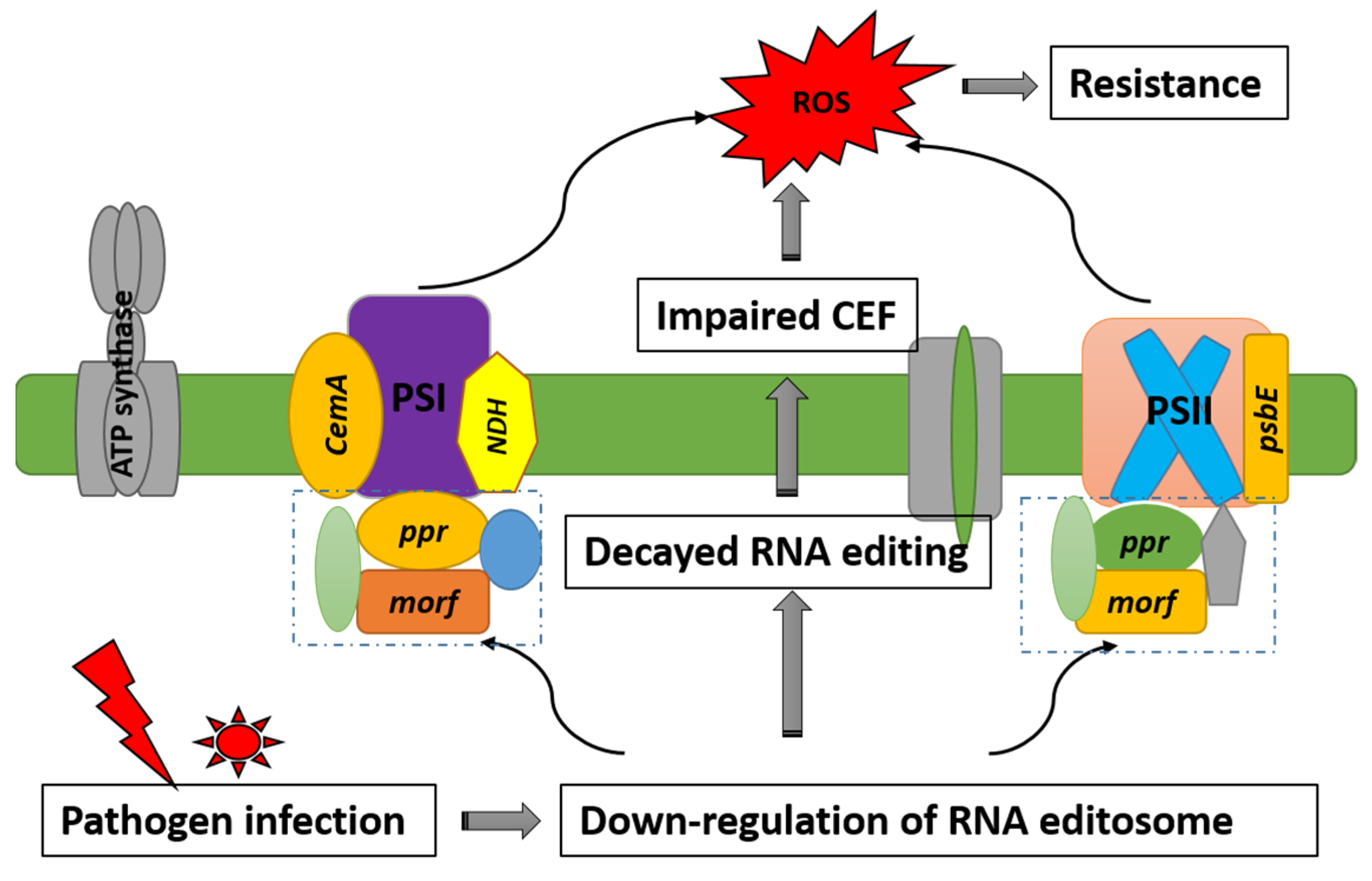
3. Discussion
4. Materials and Methods
4.1. Genome-Wide Identification of MORF Genes in Kiwifruit
4.2. Gene Structure Analysis, Subcellular and Physical Localization
4.3. Phylogenetic Tree Construction
4.4. Synteny Analysis and Detection of Tandemly/Segmentally Duplicated MORF Genes
4.5. Transcriptome Data Collection and Preprocessing
4.6. Expression Analysis of MORF Genes in Kiwifruit
4.7. Identification of RNA Editing Sites
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Wu, H.; Ma, T.; Kang, M.; Ai, F.; Zhang, J.; Dong, G.; Liu, J. A high-quality Actinidia chinensis (kiwifruit) genome. Hortic. Res. 2019, 6, 117. [Google Scholar] [CrossRef] [Green Version]
- Park, Y.S.; Im, M.H.; Ham, K.S.; Kang, S.G.; Park, Y.K.; Namiesnik, J.; Leontowicz, H.; Leontowicz, M.; Katrich, E.; Gorinstein, S. Nutritional and pharmaceutical properties of bioactive compounds in organic and conventional growing kiwifruit. Plant Foods Hum. Nutr. 2013, 68, 57–64. [Google Scholar] [CrossRef]
- Petriccione, M.; Di Cecco, I.; Arena, S.; Scaloni, A.; Scortichini, M. Proteomic changes in Actinidia chinensis shoot during systemic infection with a pandemic Pseudomonas syringae pv. actinidiae strain. J. Proteom. 2013, 78, 461–476. [Google Scholar] [CrossRef]
- Vanneste, J.L.; Yu, J.; Cornish, D.A.; Tanner, D.J.; Windner, R.; Chapman, J.R.; Taylor, R.K.; Mackay, J.F.; Dowlut, S. Identification, Virulence, and Distribution of Two Biovars of Pseudomonas syringae pv. actinidiae in New Zealand. Plant Dis. 2013, 97, 708–719. [Google Scholar] [CrossRef] [Green Version]
- Giege, P.; Brennicke, A. RNA editing in Arabidopsis mitochondria effects 441 C to U changes in ORFs. Proc. Natl. Acad. Sci. USA 1999, 96, 15324–15329. [Google Scholar] [CrossRef] [Green Version]
- Ruchika; Okudaira, C.; Sakari, M.; Tsukahara, T. Genome-Wide Identification of U-To-C RNA Editing Events for Nuclear Genes in Arabidopsis thaliana. Cells 2021, 10, 635. [Google Scholar] [CrossRef]
- Wang, D.; Meng, S.; Su, W.; Bao, Y.; Lu, Y.; Yin, W.; Liu, C.; Xia, X. Genome-Wide Analysis of Multiple Organellar RNA Editing Factor Family in Poplar Reveals Evolution and Roles in Drought Stress. Int. J. Mol. Sci. 2019, 20, 1425. [Google Scholar] [CrossRef] [Green Version]
- Chateigner-Boutin, A.L.; Small, I. Plant RNA editing. RNA Biol. 2010, 7, 213–219. [Google Scholar] [CrossRef]
- Zhang, A.; Jiang, X.; Zhang, F.; Wang, T.; Zhang, X. Dynamic response of RNA editing to temperature in grape by RNA deep sequencing. Funct. Integr. Genom. 2020, 20, 421–432. [Google Scholar] [CrossRef] [Green Version]
- Fang, J.; Jiang, X.H.; Wang, T.F.; Zhang, X.J.; Zhang, A.D. Tissue-specificity of RNA editing in plant: Analysis of transcripts from three tobacco (Nicotiana tabacum) varieties. Plant Biotechnol. Rep. 2021, 15, 471–482. [Google Scholar] [CrossRef]
- Takenaka, M.; Zehrmann, A.; Verbitskiy, D.; Kugelmann, M.; Hartel, B.; Brennicke, A. Multiple organellar RNA editing factor (MORF) family proteins are required for RNA editing in mitochondria and plastids of plants. Proc. Natl. Acad. Sci. USA 2012, 109, 5104–5109. [Google Scholar] [CrossRef] [Green Version]
- Zhang, F.; Tang, W.; Hedtke, B.; Zhong, L.; Liu, L.; Peng, L.; Lu, C.; Grimm, B.; Lin, R. Tetrapyrrole biosynthetic enzyme protoporphyrinogen IX oxidase 1 is required for plastid RNA editing. Proc. Natl. Acad. Sci. USA 2014, 111, 2023–2028. [Google Scholar] [CrossRef] [Green Version]
- Tian, F.; Yu, J.; Zhang, Y.; Xie, Y.; Wu, B.; Miao, Y. MORF9 Functions in Plastid RNA Editing with Tissue Specificity. Int. J. Mol. Sci. 2019, 20, 4635. [Google Scholar] [CrossRef] [Green Version]
- Haag, S.; Schindler, M.; Berndt, L.; Brennicke, A.; Takenaka, M.; Weber, G. Crystal structures of the Arabidopsis thaliana organellar RNA editing factors MORF1 and MORF9. Nucleic Acids Res. 2017, 45, 4915–4928. [Google Scholar] [CrossRef] [Green Version]
- Barkan, A.; Small, I. Pentatricopeptide repeat proteins in plants. Annu. Rev. Plant Biol. 2014, 65, 415–442. [Google Scholar] [CrossRef]
- Yan, J.; Zhang, Q.; Yin, P. RNA editing machinery in plant organelles. Sci. China Life Sci. 2018, 61, 162–169. [Google Scholar] [CrossRef]
- Zhang, Q.; Shen, L.; Ren, D.; Hu, J.; Chen, G.; Zhu, L.; Gao, Z.; Zhang, G.; Guo, L.; Zeng, D. Characterization, Expression, and Interaction Analyses of OsMORF Gene Family in Rice. Genes 2019, 10, 694. [Google Scholar] [CrossRef] [Green Version]
- Luo, M.; Cai, M.; Zhang, J.; Li, Y.; Zhang, R.; Song, W.; Zhang, K.; Xiao, H.; Yue, B.; Zheng, Y. Functional divergence and origin of the DAG-like gene family in plants. Sci. Rep. 2017, 7, 5688. [Google Scholar] [CrossRef] [Green Version]
- Yang, Y.; Fan, G.; Zhao, Y.; Wen, Q.; Wu, P.; Meng, Y.; Shan, W. Cytidine-to-Uridine RNA Editing Factor NbMORF8 Negatively Regulates Plant Immunity to Phytophthora Pathogens. Plant Physiol. 2020, 184, 2182–2198. [Google Scholar] [CrossRef]
- Yang, J.; Zhang, M.; Wang, X. Crystal structure of the chloroplast RNA editing factor MORF2. Biochem Biophys Res Commun 2018, 495, 2038–2043. [Google Scholar] [CrossRef]
- Bentolila, S.; Heller, W.P.; Sun, T.; Babina, A.M.; Friso, G.; van Wijk, K.J.; Hanson, M.R. RIP1, a member of an Arabidopsis protein family, interacts with the protein RARE1 and broadly affects RNA editing. Proc. Natl. Acad. Sci. USA 2012, 109, E1453–E1461. [Google Scholar] [CrossRef] [Green Version]
- Zehrmann, A.; Hartel, B.; Glass, F.; Bayer-Csaszar, E.; Obata, T.; Meyer, E.; Brennicke, A.; Takenaka, M. Selective homo- and heteromer interactions between the multiple organellar RNA editing factor (MORF) proteins in Arabidopsis thaliana. J. Biol. Chem. 2015, 290, 6445–6456. [Google Scholar] [CrossRef] [Green Version]
- Huang, C.; Li, Z.R.; Yu, Q.B.; Ye, L.S.; Cui, Y.L.; Molloy, D.P.; Yang, Z.N. MORF2 tightly associates with MORF9 to regulate chloroplast RNA editing in Arabidopsis. Plant Sci. 2019, 278, 64–69. [Google Scholar] [CrossRef]
- Zhang, Q.; Wang, Y.; Xie, W.; Chen, C.; Ren, D.; Hu, J.; Zhu, L.; Zhang, G.; Gao, Z.; Guo, L.; et al. OsMORF9 is necessary for chloroplast development and seedling survival in rice. Plant Sci. 2021, 307, 110907. [Google Scholar] [CrossRef]
- Potter, S.C.; Luciani, A.; Eddy, S.R.; Park, Y.; Lopez, R.; Finn, R.D. HMMER web server: 2018 update. Nucleic Acids Res. 2018, 46, W200–W204. [Google Scholar] [CrossRef] [Green Version]
- Bailey, T.L.; Johnson, J.; Grant, C.E.; Noble, W.S. The MEME Suite. Nucleic Acids Res. 2015, 43, W39–W49. [Google Scholar] [CrossRef] [Green Version]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis Version 7.0 for Bigger Datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [Green Version]
- Chen, C.; Chen, H.; Zhang, Y.; Thomas, H.R.; Frank, M.H.; He, Y.; Xia, R. TBtools: An Integrative Toolkit Developed for Interactive Analyses of Big Biological Data. Mol. Plant 2020, 13, 1194–1202. [Google Scholar] [CrossRef]
- Salazar, J.A.; Vergara-Pulgar, C.; Jorquera, C.; Zapata, P.; Ruiz, D.; Martinez-Gomez, P.; Infante, R.; Meneses, C. De Novo Transcriptome Sequencing in Kiwifruit (Actinidia chinensis var. deliciosa (A Chev) Liang et Ferguson) and Development of Tissue-Specific Transcriptomic Resources. Agronomy 2021, 11, 919. [Google Scholar] [CrossRef]
- Song, Y.; Sun, L.; Lin, M.; Chen, J.; Qi, X.; Hu, C.; Fang, J. Comparative transcriptome analysis of resistant and susceptible kiwifruits in response to Pseudomonas syringae pv. Actinidiae during early infection. PLoS ONE 2019, 14, e0211913. [Google Scholar] [CrossRef]
- Amirsadeghi, S.; Robson, C.A.; Vanlerberghe, G.C. The role of the mitochondrion in plant responses to biotic stress. Physiol. Plant. 2007, 129, 253–266. [Google Scholar] [CrossRef]
- Garcia-Andrade, J.; Ramirez, V.; Lopez, A.; Vera, P. Mediated plastid RNA editing in plant immunity. PLoS Pathog. 2013, 9, e1003713. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boos, F.; Muhlhaus, T.; Herrmann, J.M. Detection of Internal Matrix Targeting Signal-like Sequences (iMTS-Ls) in Mitochondrial Precursor Proteins Using the TargetP Prediction Tool. Bio Protoc. 2018, 8, e2474. [Google Scholar] [CrossRef] [PubMed]
- Hung, J.H.; Weng, Z. Sequence Alignment and Homology Search with BLAST and ClustalW. Cold Spring Harb. Protoc. 2016, 2016, 11. [Google Scholar]
- Wang, Y.; Li, J.; Paterson, A.H. MCScanX-transposed: Detecting transposed gene duplications based on multiple colinearity scans. Bioinformatics 2013, 29, 1458–1460. [Google Scholar] [CrossRef] [Green Version]
- Brown, J.; Pirrung, M.; McCue, L.A. FQC Dashboard: Integrates FastQC results into a web-based, interactive, and extensible FASTQ quality control tool. Bioinformatics 2017, 33, 3137–3139. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, D.; Langmead, B.; Salzberg, S.L. HISAT: A fast spliced aligner with low memory requirements. Nat. Methods 2015, 12, 357–360. [Google Scholar] [CrossRef] [Green Version]
- Li, H.; Handsaker, B.; Wysoker, A.; Fennell, T.; Ruan, J.; Homer, N.; Marth, G.; Abecasis, G.; Durbin, R. The Sequence Alignment/Map format and SAMtools. Bioinformatics 2009, 25, 2078–2079. [Google Scholar] [CrossRef] [Green Version]
- Danecek, P.; Bonfield, J.K.; Liddle, J.; Marshall, J.; Ohan, V.; Pollard, M.O.; Whitwham, A.; Keane, T.; McCarthy, S.A.; Davies, R.M.; et al. Twelve years of SAMtools and BCFtools. Gigascience 2021, 10, 1–4. [Google Scholar] [CrossRef]
- Pertea, M.; Pertea, G.M.; Antonescu, C.M.; Chang, T.C.; Mendell, J.T.; Salzberg, S.L. StringTie enables improved reconstruction of a transcriptome from RNA-seq reads. Nat. Biotechnol. 2015, 33, 290–295. [Google Scholar] [CrossRef] [Green Version]
- Robinson, M.D.; McCarthy, D.J.; Smyth, G.K. edgeR: A Bioconductor package for differential expression analysis of digital gene expression data. Bioinformatics 2010, 26, 139–140. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Danecek, P.; McCarthy, S.A. BCFtools/csq: Haplotype-aware variant consequences. Bioinformatics 2017, 33, 2037–2039. [Google Scholar] [CrossRef] [PubMed]
- Wu, S.; Liu, W.; Aljohi, H.A.; Alromaih, S.A.; Alanazi, I.O.; Lin, Q.; Yu, J.; Hu, S. REDO: RNA Editing Detection in Plant Organelles Based on Variant Calling Results. J. Comput. Biol. 2018, 25, 509–516. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | Gene ID | Chromosome | Length of Protein (aa 1) | Predicted Subcellular Location |
|---|---|---|---|---|
| MORF9.1_ACTCH | Actinidia35298.t1 | LG2 | 238 | chloroplast |
| MORF9.2_ACTCH | Actinidia35301.t1 | LG2 | 217 | chloroplast |
| MORF1_ACTCH | Actinidia39020.t1 | LG2 | 441 | chloroplast |
| MORF3.1_ACTCH | Actinidia18632.t1 | LG17 | 333 | mitochondrion |
| MORF3.2_ACTCH | Actinidia40300.t1 | LG22 | 272 | mitochondrion |
| MORF2.1_ACTCH | Actinidia30139.t1 | LG24 | 177 | chloroplast |
| MORF2.2_ACTCH | Actinidia37538.t1 | LG13 | 307 | chloroplast |
| MORF2.3_ACTCH | Actinidia09527.t1 | LG15 | 250 | chloroplast |
| MORF8_ACTCH | Actinidia12559.t2 | LG19 | 481 | mitochondrion |
| MORF7_ACTCH | Actinidia26821.t2 | Ctig01446 | 624 | chloroplast |
| Number of Editing Sites | Number of Edited Genes | |||
|---|---|---|---|---|
| Tissues | Chloroplast | Mitochondrion | Chloroplast | Mitochondrion |
| All | 61 | 347 | 29 | 33 |
| Flower bud | 35 | 195 | 21 | 27 |
| Flower | 37 | 151 | 24 | 24 |
| Fruit | 41 | 219 | 27 | 28 |
| Leaf | 53 | 186 | 25 | 27 |
| Shoot | 54 | 138 | 25 | 25 |
| Variety | Time Points After Psa Infection | Genes | logFC | p-Value |
|---|---|---|---|---|
| HY-HT | 0 hai | MORF9.1_ACTCH | 0.97 | 5.17 × 10−4 |
| MORF7_ACTCH | 1.21 | 2.15 × 10−3 | ||
| MORF2.1_ACTCH | 0.64 | 4.053 × 10−3 | ||
| MORF3.2_ACTCH | −0.93 | 7.06 × 10−3 | ||
| MORF1_ACTCH | −0.40 | 3.91 × 10−2 | ||
| 12 hai | MORF2.1_ACTCH | 0.84 | 1.85 × 10−8 | |
| MORF9.1_ACTCH | 1.29 | 1.35 × 10−6 | ||
| MORF7_ACTCH | 1.43 | 3.86 × 10−5 | ||
| MORF2.3_ACTCH | −3.01 | 1.8 × 10−6 | ||
| MORF2.2_ACTCH | −1.02 | 1.97 × 10−4 | ||
| MORF1_ACTCH | −0.46 | 2.08 × 10−3 | ||
| MORF3.2_ACTCH | −0.73 | 1.01 × 10−2 | ||
| HT | 0 hai−12 hai | MORF2.3_ACTCH | −1.96 | 4.75 × 10−2 |
| 12 hai−24 hai | MORF7_ACTCH | −1.23 | 2.63 × 10−4 | |
| 12 hai−48 hai | MORF2.1_ACTCH | −0.62 | 4.04 × 10−5 | |
| MORF7_ACTCH | −1.49 | 5.71 × 10−5 | ||
| MORF9.2_ACTCH | 0.88 | 5.31 × 10−3 | ||
| MORF1_ACTCH | 0.42 | 9.13 × 10−3 | ||
| MORF8_ACTCH | 0.39 | 1.34 × 10−2 | ||
| HY | 12 hai−48 hai | MORF2.2_ACTCH | −0.55 | 3.2764 × 10−2 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xiong, Y.; Fang, J.; Jiang, X.; Wang, T.; Liu, K.; Peng, H.; Zhang, X.; Zhang, A. Genome-Wide Analysis of Multiple Organellar RNA Editing Factor (MORF) Family in Kiwifruit (Actinidia chinensis) Reveals Its Roles in Chloroplast RNA Editing and Pathogens Stress. Plants 2022, 11, 146. https://doi.org/10.3390/plants11020146
Xiong Y, Fang J, Jiang X, Wang T, Liu K, Peng H, Zhang X, Zhang A. Genome-Wide Analysis of Multiple Organellar RNA Editing Factor (MORF) Family in Kiwifruit (Actinidia chinensis) Reveals Its Roles in Chloroplast RNA Editing and Pathogens Stress. Plants. 2022; 11(2):146. https://doi.org/10.3390/plants11020146
Chicago/Turabian StyleXiong, Yuhong, Jing Fang, Xiaohan Jiang, Tengfei Wang, Kangchen Liu, Huixiang Peng, Xiujun Zhang, and Aidi Zhang. 2022. "Genome-Wide Analysis of Multiple Organellar RNA Editing Factor (MORF) Family in Kiwifruit (Actinidia chinensis) Reveals Its Roles in Chloroplast RNA Editing and Pathogens Stress" Plants 11, no. 2: 146. https://doi.org/10.3390/plants11020146
APA StyleXiong, Y., Fang, J., Jiang, X., Wang, T., Liu, K., Peng, H., Zhang, X., & Zhang, A. (2022). Genome-Wide Analysis of Multiple Organellar RNA Editing Factor (MORF) Family in Kiwifruit (Actinidia chinensis) Reveals Its Roles in Chloroplast RNA Editing and Pathogens Stress. Plants, 11(2), 146. https://doi.org/10.3390/plants11020146