
Phenotypic Variability of Root System Architecture Traits for Drought Tolerance among Accessions of Citron Watermelon (Citrullus lanatus var. citroides (L.H. Bailey)

 ,
,  ,
,  , and
, and
Abstract
1. Introduction
2. Results
2.1. Gravimetric Water
2.2. Root Growth of Citron Watermelon Accessions under Non-Stress and Water Stress Conditions
2.3. ANOVA Showing Accession, Water Regime and Their Interactions on Root and Shoot Traits of Citron Watermelon
2.4. Pearson Correlation Analysis Showing Associations of RSA Traits among Citron Watermelon Accessions under Non-Stressed and Water-Stressed Conditions
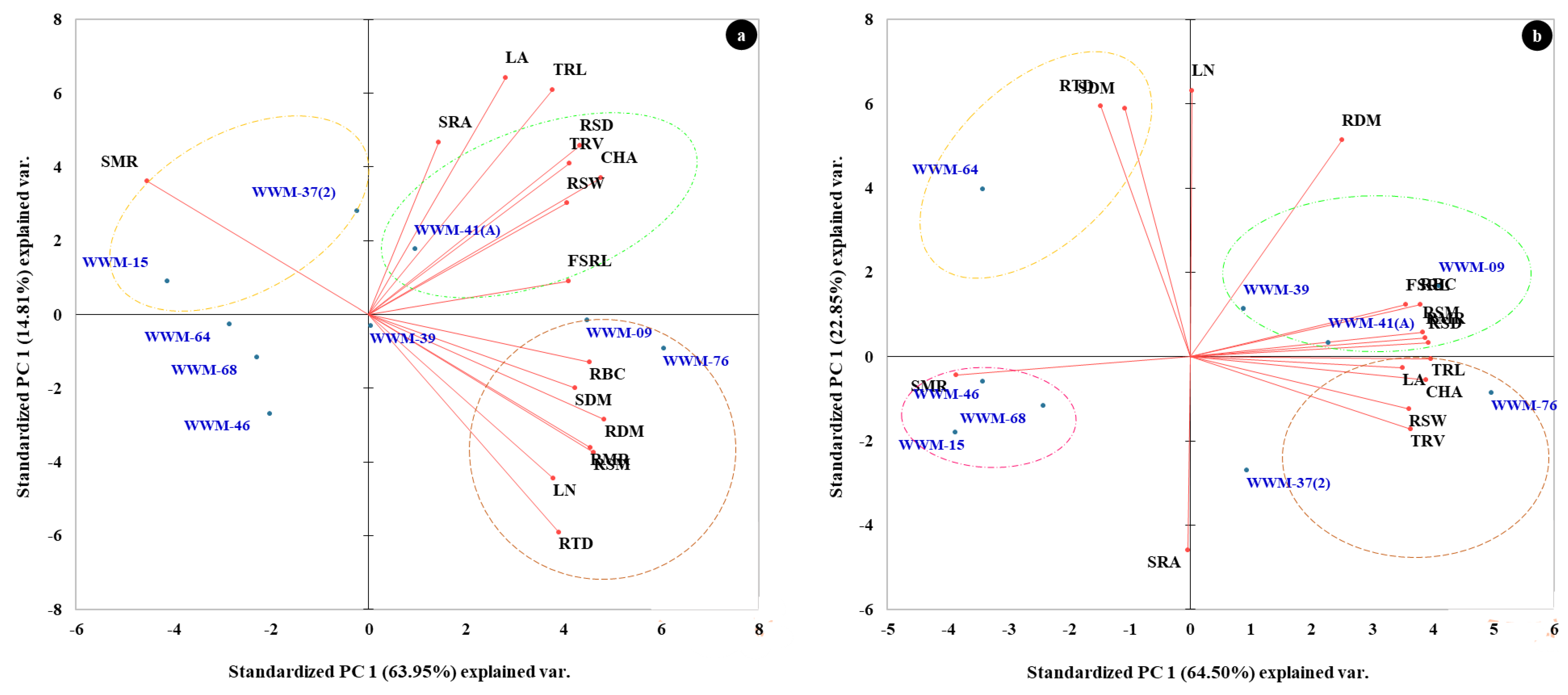
2.5. Principal Component Analysis (PCA) for Root System Architecture of Citron Watermelon Accessions Evaluated under Non-Stressed and Water-Stressed Conditions
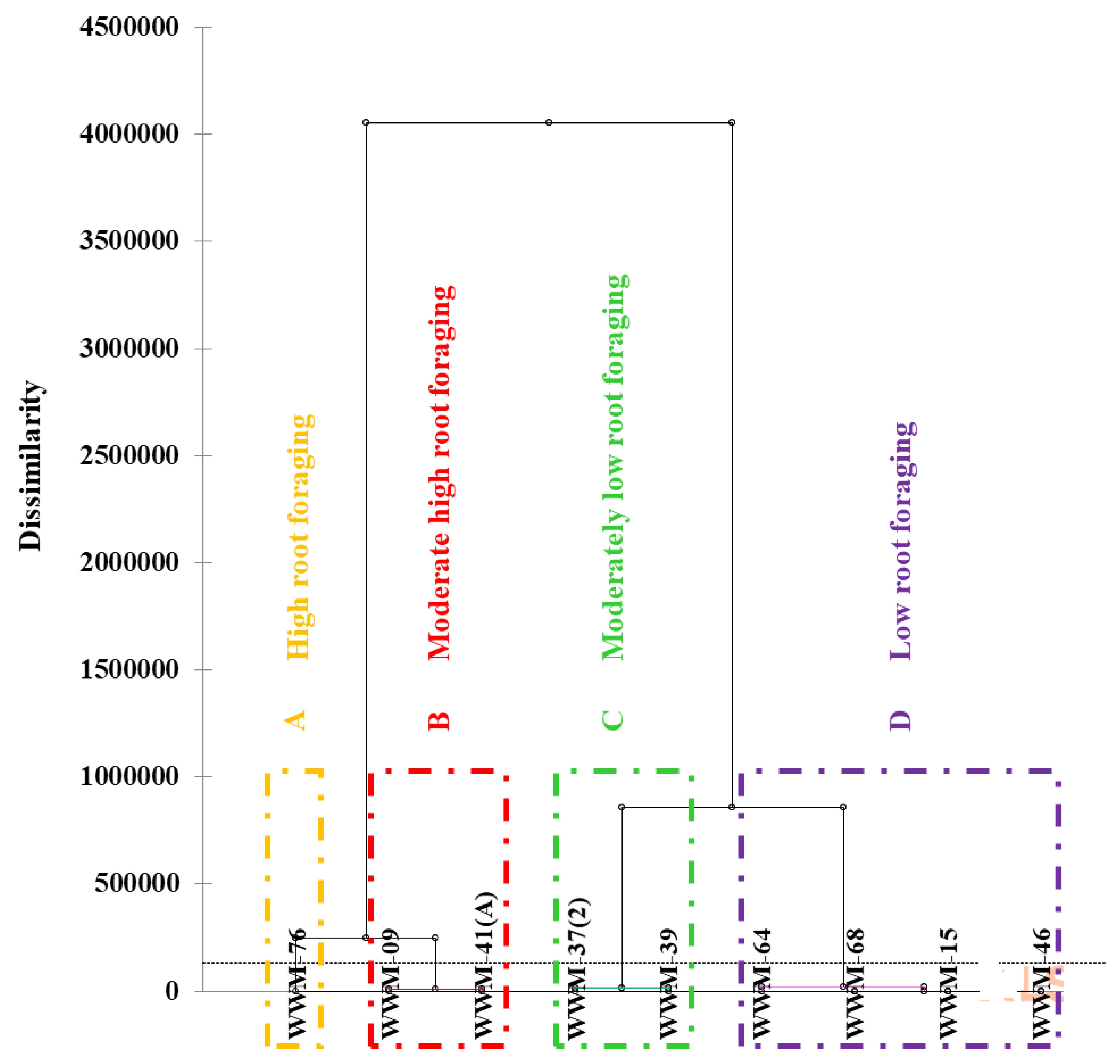
2.6. Root Vigour (Foraging Capacity) of Citron Watermelon Accessions under Water Stress Conditions
3. Discussion
4. Materials and Methods
4.1. Plant Material
4.2. Fabrication Rhizotron Prototype
4.3. Growth Substrate Selection
4.4. Experimental Design and Growth Conditions
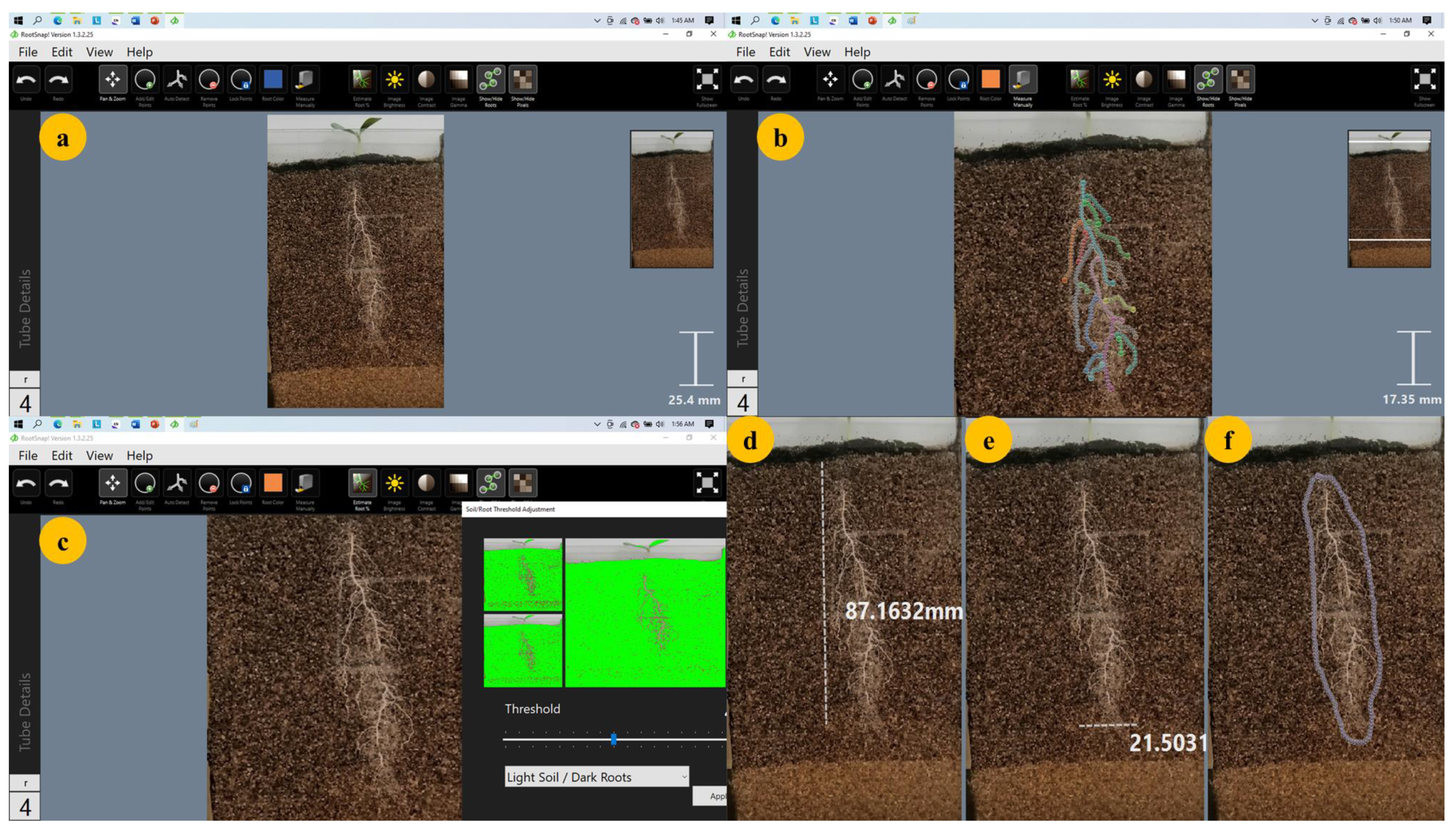
4.5. Image Acquisition
4.6. Image Analysis and Data Collection
4.7. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Mandizvo, T.; Odindo, A.O.; Mashilo, J. Nutrient composition and physical properties of citron watermelon (Citrullus lanatus var. citroides (L.H. Bailey) Mansf. ex Greb.) seeds are related to seed coat visual appearance. S. Afr. J. Bot. 2022, 145, 405–419. [Google Scholar] [CrossRef]
- Nkoana, D.K.; Mashilo, J.; Shimelis, H.; Ngwepe, R.M. Nutritional, phytochemical compositions and natural therapeutic values of citron watermelon (Citrullus lanatus var. citroides): A Review. S. Afr. J. Bot. 2021, 145, 65–77. [Google Scholar] [CrossRef]
- Mashilo, J.; Shimelis, H.; Ngwepe, R.M. Genetic resources of bottle gourd (Lagenaria siceraria (Molina) Standl.] and citron watermelon (Citrullus lanatus var. citroides (L.H. Bailey) Mansf. ex Greb.): Implications for genetic improvement, product development and commercialization: A review. S. Afr. J. Bot. 2021, 145, 28–47. [Google Scholar] [CrossRef]
- Mandizvo, T.; Odindo, A.O.; Mashilo, J. Citron Watermelon Potential to Improve Crop Diversification and Reduce Negative Impacts of Climate Change. Sustainability 2021, 13, 2269. [Google Scholar] [CrossRef]
- Hong, T.; Cai, Z.; Li, R.; Liu, J.; Li, J.; Wang, Z.; Zhang, Z. Effects of water and nitrogen coupling on watermelon growth, photosynthesis and yield under CO2 enrichment. Agric. Water Manag. 2022, 259, 107229. [Google Scholar] [CrossRef]
- Yavuz, D.; Seymen, M.; Süheri, S.; Yavuz, N.; Türkmen, Ö.; Kurtar, E.S. How do rootstocks of citron watermelon (Citrullus lanatus var. citroides) affect the yield and quality of watermelon under deficit irrigation? Agric. Water Manag. 2020, 241, 106351. [Google Scholar] [CrossRef]
- Katuuramu, D.; Branham, S.; Levi, A.; Wechter, P. Genome-wide association analysis of resistance to Pseudoperonospora cubensis in citron watermelon. Plant Dis. 2021, 106, 1952–1958. [Google Scholar] [CrossRef]
- Ngwepe, R.M.; Mashilo, J.; Shimelis, H. Progress in genetic improvement of citron watermelon (Citrullus lanatus var. citroides): A review. Genet. Resour. Crop Evol. 2019, 66, 735–758. [Google Scholar] [CrossRef]
- Maja, D.; Mavengahama, S.; Mashilo, J. Cucurbitacin biosynthesis in cucurbit crops, their pharmaceutical value and agricultural application for management of biotic and abiotic stress: A review. S. Afr. J. Bot. 2021, 145, 3–12. [Google Scholar] [CrossRef]
- Katuuramu, D.N.; Wechter, W.P.; Washington, M.L.; Horry, M.; Cutulle, M.A.; Jarret, R.L.; Levi, A. Phenotypic diversity for root traits and identification of superior germplasm for root breeding in watermelon. Hortscience 2020, 55, 1272–1279. [Google Scholar] [CrossRef]
- Schwarz, D.; Rouphael, Y.; Colla, G.; Venema, J.H. Grafting as a tool to improve tolerance of vegetables to abiotic stresses: Thermal stress, water stress and organic pollutants. Sci. Hortic. 2010, 127, 162–171. [Google Scholar] [CrossRef]
- Chomicki, G.; Renner, S.S. Watermelon origin solved with molecular phylogenetics including Linnaean material: Another example of museomics. New Phytol. 2015, 205, 526–532. [Google Scholar] [CrossRef] [PubMed]
- Gloaguen, R.M.; Brym, Z.T.; Peeples, J.; Xu, W.; Chun, H.-C.; Rowland, D.L. The plasticity of early root development in Sesamum indicum L. as influenced by genotype, water, and nutrient availability. Rhizosphere 2022, 21, 100457. [Google Scholar] [CrossRef]
- Koevoets, I.T.; Venema, J.H.; Elzenga, J.T.; Testerink, C. Roots Withstanding their Environment: Exploiting Root System Architecture Responses to Abiotic Stress to Improve Crop Tolerance. Front. Plant. Sci. 2016, 7, 1335. [Google Scholar] [CrossRef] [PubMed]
- de Kroon, H.; Huber, H.; Stuefer, J.F.; van Groenendael, J.M. A modular concept of phenotypic plasticity in plants. New Phytol. 2005, 166, 73–82. [Google Scholar] [CrossRef]
- Uygur, V.; Yetisir, H. Effects of Rootstocks on Some Growth Parameters, Phosphorous and Nitrogen Uptake Watermelon under Salt Stress. J. Plant Nutr. 2009, 32, 629–643. [Google Scholar] [CrossRef]
- Thies, J.A.; Levi, A. Characterization of Watermelon (Citrullus lanatus var. citroides) Germplasm for Resistance to Root-knot Nematodes. Hortscience 2007, 42, 1530–1533. [Google Scholar] [CrossRef]
- Omirou, M.; Ioannides, I.M.; Ehaliotis, C. Mycorrhizal inoculation affects arbuscular mycorrhizal diversity in watermelon roots, but leads to improved colonization and plant response under water stress only. Appl. Soil Ecol. 2013, 63, 112–119. [Google Scholar] [CrossRef]
- Rentería-Martínez, M.E.; Guerra-Camacho, M.Á.; Ochoa-Meza, A.; Moreno-Salazar, S.F.; Varela-Romero, A.; Gutiérrez-Millán, L.E.; Meza-Moller, A.D.C. Análisis filogenético multilocus del complejo fúngico asociado a pudrición radicular de sandía en Sonora, México. Rev. Mex. Fitopatol. Mex. J. Phytopathol. 2018, 36, 233–255. [Google Scholar] [CrossRef]
- Roy, A.; Bal, S.S.; Fergany, M.; Kaur, S.; Singh, H.; Malik, A.A.; Singh, J.; Monforte, A.J.; Dhillon, N.P.S. Wild melon diversity in India (Punjab State). Genet. Resour. Crop Evol. 2012, 59, 755–767. [Google Scholar] [CrossRef]
- Crosby, K.M.; Jifon, J.L.; Leskovar, D.I. Genetic Improvement of Early Root Vigor in Melon (Cucumis melo L.) to Enhance Stand Establishment. Acta Hortic. 2008, 273–278. [Google Scholar] [CrossRef]
- Fita, A.; Picó, B.; Nuez, F. Melon roots under stress: Melon vine decline. Plant Stress 2007, 1, 104. [Google Scholar]
- Mashilo, J.; Shimelis, H.; Odindo, A.; Amelework, B. Simple Sequence Repeat Markers Reveal Genetic Diversity Within and among Landrace Collections of Citron and Dessert Watermelon from South Africa. J. Am. Soc. Hortic. Sci. 2016, 141, 598–608. [Google Scholar] [CrossRef]
- Mashilo, J.; Shimelis, H.; Odindo, A.; Amelework, B. Assessment of the genetic diversity of dessert watermelon (Citrullus lanatus var. lanatus) landrace collections of South Africa using SSR markers. Aust. J. Crop Sci. 2017, 11, 1392–1398. [Google Scholar] [CrossRef]
- Mashilo, J.; Shimelis, H.; Odindo, A.O.; Amelework, B. Genetic diversity and differentiation in citron watermelon [Citrullus lanatus var. citroides] landraces assessed by simple sequence repeat markers. Sci. Hortic. 2017, 214, 99–106. [Google Scholar] [CrossRef]
- Mandizvo, T.; Odindo, A.O.; Mashilo, J.; Magwaza, L.S. Drought tolerance assessment of citron watermelon (Citrullus lanatus var. citroides (L.H. Bailey) Mansf. ex Greb.) accessions based on morphological and physiological traits. Plant Physiol. Biochem. 2022, 180, 106–123. [Google Scholar] [CrossRef]
- Cavalcante, V.S.; Prado, R.d.M.; Vasconcelos, R.d.L.; Júnior de Almeida, H.; Ramos da Silva, T. Growth and Nutritional Efficiency of Watermelon Plants Grown under Macronutrient Deficiencies. Hortscience 2019, 54, 738–742. [Google Scholar] [CrossRef]
- Narayanan, S.; Mohan, A.; Gill, K.S.; Prasad, P.V. Variability of root traits in spring wheat germplasm. PLoS ONE 2014, 9, e100317. [Google Scholar] [CrossRef]
- Qin, K.; Leskovar, D.I. Humic Substances Improve Vegetable Seedling Quality and Post-Transplant Yield Performance under Stress Conditions. Agriculture 2020, 10, 254. [Google Scholar] [CrossRef]
- Tomlinson, K.W.; Sterck, F.J.; Bongers, F.; da Silva, D.A.; Barbosa, E.R.M.; Ward, D.; Bakker, F.T.; van Kaauwen, M.; Prins, H.H.T.; de Bie, S.; et al. Biomass partitioning and root morphology of savanna trees across a water gradient. J. Ecol. 2012, 100, 1113–1121. [Google Scholar] [CrossRef]
- Schenk, H.J.; Jackson, R.B. Rooting Depths, Lateral Root Spreads and Below-Ground/Above-Ground Allometries of Plants in Water-Limited Ecosystems. J. Ecol. 2002, 90, 480–494. [Google Scholar] [CrossRef]
- Guzzon, F.; Müller, J.V.; do Nascimento Araujo, M.; Cauzzi, P.; Orsenigo, S.; Mondoni, A.; Abeli, T. Drought avoidance adaptive traits in seed germination and seedling growth of Citrullus amarus landraces. S. Afr. J. Bot. 2017, 113, 382–388. [Google Scholar] [CrossRef]
- Shipley, B.; Meziane, D. The balanced-growth hypothesis and the allometry of leaf and root biomass allocation. Funct. Ecol. 2002, 16, 326–331. [Google Scholar] [CrossRef]
- Glynn, C.; Herms, D.A.; Orians, C.M.; Hansen, R.C.; Larsson, S. Testing the growth-differentiation balance hypothesis: Dynamic responses of willows to nutrient availability. New Phytol. 2007, 176, 623–634. [Google Scholar] [CrossRef] [PubMed]
- Akmal, M.; Hirasawa, T. Growth responses of seminal roots of wheat seedlings to a reduction in the water potential of vermiculite. Plant Soil 2004, 267, 319–328. [Google Scholar] [CrossRef]
- Mandizvo, T.; Odindo, A.O. Seed mineral reserves and vigour of Bambara groundnut (Vigna subterranea L.) landraces differing in seed coat colour. Heliyon 2019, 5, e01635. [Google Scholar] [CrossRef]
- Maskova, T.; Herben, T. Root: Shoot ratio in developing seedlings: How seedlings change their allocation in response to seed mass and ambient nutrient supply. Ecol. Evol. 2018, 8, 7143–7150. [Google Scholar] [CrossRef]
- Seymen, M.; Yavuz, D.; Ercan, M.; Akbulut, M.; Çoklar, H.; Kurtar, E.S.; Yavuz, N.; Süheri, S.; Türkmen, Ö. Effect of wild watermelon rootstocks and water stress on chemical properties of watermelon fruit. Hortic. Environ. Biotechnol. 2021, 62, 411–422. [Google Scholar] [CrossRef]
- Thies, J.A.; Ariss, J.J.; Hassell, R.L.; Buckner, S.; Levi, A. Accessions of Citrullus lanatus var. citroides are valuable rootstocks for grafted watermelon in fields infested with root-knot nematodes. Hortscience 2015, 50, 4–8. [Google Scholar] [CrossRef]
- Wiese, A.H.; Riemenschneider, D.E.; Ronald, S., Jr. An inexpensive rhizotron design for two-dimensional, horizontal root growth measurements. Tree Plant. Notes 2005, 51, 40–46. [Google Scholar]
- Haney, R.L.; Haney, E.B. Simple and Rapid Laboratory Method for Rewetting Dry Soil for Incubations. Commun. Soil Sci. Plant Anal. 2010, 41, 1493–1501. [Google Scholar] [CrossRef]
- Easlon, H.M.; Bloom, A.J. Easy Leaf Area: Automated digital image analysis for rapid and accurate measurement of leaf area. Appl. Plant Sci. 2014, 2, 1400033. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| RSW (cm day−1) | RSD (cm day−1) | CHA (cm2 day−1) | TRL (cm day−1) | RBC (branch/day) | LA (cm2 day−1) | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Accession | NS | WS | NS | WS | NS | WS | NS | WS | NS | WS | NS | WS |
| WWM-09 | 0.285 c | 0.314 d | 1.016 b | 1.278 a | 66.743 ab | 78.593 b | 2.192 bc | 2.325 b | 1.746 f | 1.800 e | 2.578 a | 0.762 b |
| WWM-15 | 0.191 h | 0.284 g | 0.859 c | 0.889 d | 39.172 e | 40.728 e | 1.670 e | 1.779 d | 1.263 b | 1.298 b | 1.909 d | 0.278 e |
| WWM-37(2) | 0.273 e | 0.325 c | 0.909 c | 1.102 c | 58.729 c | 68.341 c | 2.499 a | 2.207 c | 1.293 bc | 1.432 c | 2.379 b | 0.788 b |
| WWM-39 | 0.236 g | 0.298 e | 1.047 ab | 1.141 bc | 48.044 d | 60.276 d | 2.207 bc | 2.144 c | 1.475 de | 1.482 c | 1.731 e | 0.667 c |
| WWM-41(A) | 0.338 b | 0.430 a | 1.045 ab | 1.152 bc | 60.933 bc | 79.814 b | 2.228 b | 2.362 ab | 1.507 e | 1.677 d | 1.987 c | 0.896 a |
| WWM-46 | 0.189 h | 0.296 e | 0.693 d | 0.875 d | 29.129 f | 40.474 e | 1.557 f | 1.724 d | 1.085 a | 1.197 a | 1.745 e | 0.277 e |
| WWM-64 | 0.252 f | 0.221 h | 0.908 c | 0.896 d | 27.663 f | 37.138 e | 1.783 d | 1.745 d | 1.322 c | 1.317 b | 1.950 cd | 0.403 d |
| WWM-68 | 0.280 d | 0.292 f | 0.845 c | 0.877 d | 31.715 f | 41.477 e | 1.553 f | 1.779 d | 1.453 d | 1.319 b | 1.268 f | 0.804 b |
| WWM-76 | 0.386 a | 0.403 b | 1.120 a | 1.250 ab | 71.403 a | 91.609 a | 2.139 c | 2.463 a | 1.832 g | 1.615 d | 1.783 e | 0.894 a |
| Mean | 0.270 | 0.318 | 0.938 | 1.051 | 48.170 | 59.828 | 1.981 | 2.057 | 1.442 | 1.460 | 1.926 | 0.641 |
| LSD | 3.431 × 10−3 | 0.003 | 0.086 | 0.120 | 6.124 | 7.376 | 0.086 | 0.100 | 0.051 | 0.069 | 0.067 | 0.051 |
| CV (%) | 0.700 | 0.600 | 5.300 | 6.700 | 7.400 | 7.200 | 2.500 | 2.900 | 2.100 | 2.700 | 2.100 | 4.700 |
| p-value | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 |
| Source of Variation | d.f | RSW | RSD | CHA | TRL | RBC | TRV | LA | LN |
|---|---|---|---|---|---|---|---|---|---|
| Accession (A) | 8 | 25.763 ** | 117.558 ** | 1.640 × 106 ** | 578.406 ** | 183.560 ** | 1.060 ** | 4.383 × 102 ** | 6.292 |
| Water Condition (WC) | 1 | 6.001 ** | 48.964 ** | 8.952 × 105 ** | 106.145 ** | 150 ** | 2.212 ** | 1.665 × 104 ** | 280.167 * |
| A × WC | 8 | 1.545 ** | 5.107 ** | 3.152 × 104 ** | 29.877 ** | 12.625 ** | 0.163 ** | 1.255 × 102 ** | 3.792 * |
| Residual | 36 | 0.025 | 0.051 | 131.900 | 0.018 | 0.093 | 0.023 | 4.138 × 10−2 | 1.847 |
| Source of Variation | d.f | FSRL | SRA | RDM | SDM | RSM | RMR | SMR | RTD |
| Accession (A) | 8 | 1.892 ** | 141.800 ns | 2.875 ** | 0.296 ** | 0.693 ** | 0.015 ** | 0.015 ** | 0.520 ** |
| Water Condition (WC) | 1 | 0.056 | 197.100 ns | 22.970 ** | 27.549 ** | 5.771 ** | 0.135 ** | 0.135 ** | 11.447 ** |
| A × WC | 8 | 0.048 ** | 130.400 * | 1.443 ** | 0.330 ** | 0.073 * | 8.489 × 10−4 | 8.489 × 10−4 ** | 0.388 ** |
| Residual | 36 | 0.007 | 379.700 | 0.056 | 0.016 | 0.027 | 5.185 × 10−4 | 5.185 × 10−4 | 0.045 |
| Below Ground | Above Ground | Allometry | ||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Accession | RSW | RSD | CHA | TRL | RBC | TRV | FSRL | SRA | LA | LN | RDM | SDM | RSM | RMR | SMR | RTD |
| WWM-09 | 10.940 c | 38.410 a | 2102 c | 75.230 a | 48 a | 2.423 b | 2.618 b | 58.120 a | 28.600 d | 6.000 ab | 2.097 a | 0.822 cd | 2.558 a | 0.718 a | 0.281 e | 0.866 bc |
| WWM-15 | 8.350 g | 27.050 f | 1129 f | 53.250 g | 35 g | 1.450 d | 1.163 f | 58.470 a | 14.170 h | 4.000 c | 0.989 e | 0.711 de | 1.393 d | 0.582 e | 0.418 a | 0.683 cde |
| WWM-37(2) | 10.400 d | 33.030 e | 1718 d | 66.510 d | 38 e | 2.697 ab | 2.285 c | 71.700 a | 30.740 c | 4.000 c | 1.355 d | 0.747 de | 1.827 bc | 0.645 bcd | 0.355 bcd | 0.504 e |
| WWM-39 | 9.450 e | 34.010 d | 1606 e | 64.720 e | 42 d | 1.983 c | 2.483 b | 61.900 a | 24.690 e | 5.000 bc | 1.916 b | 0.960 b | 2.005 b | 0.666 b | 0.334 d | 0.975 b |
| WWM-41(A) | 12.600 b | 34.820 c | 2193 b | 71.730 c | 43 c | 2.401 b | 1.899 d | 55.670 a | 35.430 a | 5.000 bc | 1.747 c | 0.900 bc | 1.976 b | 0.663 bc | 0.338 cd | 0.733 cd |
| WWM-46 | 8.720 f | 25.820 h | 1126 f | 51.180 h | 33 h | 1.315 d | 1.531 e | 65.670 a | 11.360 i | 5.000 bc | 1.258 d | 0.771 cde | 1.648 bcd | 0.620 cde | 0.380 abc | 0.969 b |
| WWM-64 | 7.620 h | 26.620 g | 1014 g | 53.030 g | 35 g | 1.493 d | 1.849 d | 55.670 a | 18.600 g | 7.000 a | 1.888 b | 1.293 a | 1.466 cd | 0.594 e | 0.407 a | 1.279 a |
| WWM-68 | 8.270 g | 26.950 f | 1114 f | 55.070 f | 36 f | 1.928 c | 1.570 e | 63.030 a | 21.070 f | 5.000 bc | 1.233 d | 0.780 cde | 1.585 cd | 0.613 de | 0.387 ab | 0.641 de |
| WWM-76 | 13.940 a | 36.990 b | 2579 a | 75.800 a | 45 b | 2.912 a | 2.888 a | 61.900 a | 31.730 b | 5.000 bc | 1.709 c | 0.681 e | 2.535 a | 0.716 a | 0.285 e | 0.589 de |
| l.s.d | 0.120 | 0.236 | 22.280 | 0.278 | 0.572 | 0.292 | 0.137 | 33.130 | 0.100 | 1.715 | 0.122 | 0.120 | 0.354 | 0.041 | 0.041 | 0.180 |
| CV (%) | 0.700 | 0.400 | 0.800 | 0.300 | 0.800 | 8.200 | 3.900 | 31.500 | 0.200 | 19.600 | 4.500 | 8.200 | 10.900 | 3.700 | 6.700 | 13.100 |
| p-value | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | 0.984 | <0.001 | 0.045 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 |
| Below Ground | Above Ground | Allometry | ||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Accession | RSW | RSD | CHA | TRL | RBC | TRV | FSRL | SRA | LA | LN | RDM | SDM | RSM | RMR | SMR | RTD |
| WWM-09 | 9.875 c | 33.620 b | 1928 b | 70.990 b | 50 b | 1.753 bc | 2.458 bc | 65.470 a | 76.410 a | 11.000 ab | 4.489 b | 2.884 a | 1.553 ab | 0.608 ab | 0.392 ef | 2.584 a |
| WWM-15 | 6.411 g | 25.860 f | 1020 f | 49.970 g | 39 f | 1.541 cd | 1.106 f | 60.600 a | 57.880 e | 7.000 e | 1.852 e | 2.042 bc | 0.905 g | 0.475 f | 0.525 a | 1.219 de |
| WWM-37(2) | 9.889 c | 31.260 d | 1594 d | 71.720 a | 36 i | 1.837 b | 2.611 b | 62.430 a | 71.820 b | 9.000 cd | 2.061 de | 1.800 c | 1.142 def | 0.533 de | 0.467 bc | 1.137 e |
| WWM-39 | 8.448 e | 31.330 d | 1337 e | 66.480 d | 45 d | 1.596 bcd | 2.353 bc | 45.470 a | 55.240 g | 9.000 de | 2.735 c | 2.172 b | 1.266 cd | 0.558 cd | 0.442 cd | 1.717 cd |
| WWM-41(A) | 12.132 b | 32.350 c | 1702 c | 67.520 c | 46 c | 1.809 b | 1.724 d | 66.600 a | 63.470 c | 10.000 bc | 2.695 c | 2.291 b | 1.169 de | 0.538 de | 0.462 bc | 1.502 cde |
| WWM-46 | 7.053 f | 23.040 g | 872 h | 48.060 i | 37 h | 1.265 e | 1.793 d | 53.030 a | 50.330 h | 10.000 bc | 2.508 cd | 1.844 c | 1.370 bc | 0.577 bc | 0.423 de | 1.992 bc |
| WWM-64 | 8.958 d | 28.770 e | 812 h | 52.140 f | 38 g | 1.369 de | 1.884 d | 45.300 a | 56.730 f | 9.000 cd | 2.129 cde | 2.247 b | 0.950 fg | 0.486 f | 0.514 a | 1.555 cde |
| WWM-68 | 8.644 de | 25.420 f | 920 g | 48.750 h | 41 e | 1.596 bcd | 1.520 e | 61.870 a | 39.870 i | 11.000 ab | 2.343 cde | 2.224 b | 1.050 efg | 0.512 ef | 0.488 ab | 1.466 cde |
| WWM-76 | 12.879 a | 34.910 a | 2078 a | 65.660 e | 54 a | 2.193 a | 2.837 a | 56.970 a | 60.720 d | 12.000 a | 5.118 a | 3.007 a | 1.704 a | 0.630 a | 0.370 f | 2.355 ab |
| l.s.d | 0.367 | 0.496 | 16.730 | 0.161 | 0.467 | 0.228 | 0.155 | 33.710 | 0.483 | 1.715 | 0.560 | 0.288 | 0.185 | 0.037 | 0.037 | 0.481 |
| CV (%) | 2.300 | 1.000 | 0.700 | 0.200 | 0.600 | 8.000 | 4.500 | 34.200 | 0.500 | 10.300 | 11.300 | 7.400 | 8.700 | 4.000 | 4.800 | 16.300 |
| p-value | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | 0.842 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 |
| Traits | RSW | RSD | CHA | TRL | RBC | TRV | FSRL | SRA | LA | LN | RDM | SDM | RSM | RMR | SMR | RTD |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| RSW | 1 | 0.83 ** | 0.98 ** | 0.90 ** | 0.78 ** | 0.85 ** | 0.69 * | −0.02 ns | 0.82 ** | −0.16 ns | 0.37 ns | −0.39 ns | 0.82 ** | 0.83 ** | −0.82 ** | −0.49 ns |
| RSD | 0.83 ** | 1 | 0.93 ** | 0.99 ** | 0.97 ** | 0.86 ** | 0.87 ** | −0.06 ns | 0.85 ** | 0.01 ns | 0.66 * | −0.22 ns | 0.93 ** | 0.94 ** | −0.94 ** | −0.31 ns |
| CHA | 0.80 ** | 0.90 ** | 1 | 0.97 ** | 0.89 ** | 0.89 ** | 0.79 ** | −0.06 ns | 0.86 ** | −0.08 ns | 0.51 * | −0.33 ns | 0.90 ** | 0.91 ** | −0.90 ** | −0.43 ns |
| TRL | 0.67 * | 0.89 ** | 0.87 ** | 1 | 0.95 ** | 0.91 ** | 0.84 ** | −0.06 ns | 0.91 ** | −0.02 ns | 0.61 * | −0.25 ns | 0.92 ** | 0.93 ** | −0.93 ** | −0.39 ns |
| RBC | 0.70 * | 0.75 ** | 0.79 ** | 0.53 * | 1 | 0.77 ** | 0.82 ** | −0.25 ns | 0.79 ** | 0.14 ns | 0.72 * | −0.15 ns | 0.93 ** | 0.93 ** | −0.93 ** | −0.21 ns |
| TRV | 0.83 ** | 0.82 ** | 0.89 ** | 0.70 * | 0.73 * | 1 | 0.79 ** | 0.23 ns | 0.92 ** | −0.19 ns | 0.38 ns | −0.36 ns | 0.78 ** | 0.80 ** | −0.80 ** | −0.62 * |
| FSRL | 0.62 * | 0.76 ** | 0.72 * | 0.76 ** | 0.51 * | 0.63 * | 1 | 0.12 ns | 0.71* | 0.20 ns | 0.73 * | −0.04 ns | 0.89 ** | 0.90 ** | −0.90 ** | −0.11 ns |
| SRA | 0.32 ns | 0.20 ns | 0.47 ns | 0.31 ns | 0.21 ns | 0.46 ns | −0.09 ns | 1 | −0.02 ns | −0.58 * | −0.43 ns | −0.50 * | −0.03 ns | 0.01 ns | −0.02 ns | −0.48 ns |
| LA | 0.41 ns | 0.70 * | 0.71 * | 0.80 ** | 0.30 ns | 0.46 ns | 0.54 * | 0.38 ns | 1 | −0.05 ns | 0.53 * | −0.10 ns | 0.70 * | 0.74 * | −0.73 * | −0.44 ns |
| LN | −0.07 ns | 0.22 ns | 0.06 ns | 0.28 ns | 0.23 ns | −0.02 ns | 0.30 ns | −0.54 * | −0.14 ns | 1 | 0.70 * | 0.79 ns | 0.12 ns | 0.09 ns | −0.09 ns | 0.80 ** |
| RDM | 0.64 * | 0.68 * | 0.79 ** | 0.49 ns | 0.91 ** | 0.68 * | 0.67 * | 0.18 ns | 0.38 ns | 0.07 ns | 1 | 0.53 * | 0.65 * | 0.66 * | −0.65 * | 0.46 ns |
| SDM | 0.64 * | 0.69 * | 0.68 * | 0.39 ns | 0.92 ** | 0.63 * | 0.47 ns | 0.18 ns | 0.31 ns | 0.01 ns | 0.91 ** | 1 | −0.28 ns | −0.28 ns | 0.28 ns | 0.84 ** |
| RSM | 0.54 * | 0.55 * | 0.73 * | 0.50 * | 0.76 ** | 0.56 * | 0.74 * | 0.11 ns | 0.35 ns | 0.16 ns | 0.92 ** | 0.69 * | 1 | 0.99 ** | −0.99 ** | −0.24 ns |
| RMR | 0.53 * | 0.54 * | 0.72 * | 0.52 * | 0.73 * | 0.54 * | 0.75 ** | 0.12 ns | 0.34 ns | 0.20 ns | 0.88 ** | 0.63 * | 0.99 ** | 1 | −1.00 ** | −0.25 ns |
| SMR | −0.53 * | −0.54 * | −0.72 * | −0.52 * | −0.73 * | −0.54 * | −0.75 ** | −0.12 ns | −0.34 ns | −0.20 ns | −0.88 ** | −0.63 * | −0.99 ** | 0.23 ns | 1 | 0.25 ns |
| RTD | 0.34 ns | 0.41 ns | 0.51 * | 0.26 ns | 0.73 * | 0.27 ns | 0.52 * | −0.01 ns | 0.26 ns | 0.10 ns | 0.89 ** | 0.78 ** | 0.88 ** | 0.86 ** | −0.86 ** | 1 |
| Non-Stress | Water-Stress | |||||
|---|---|---|---|---|---|---|
| Traits | PC 1 | PC 2 | KMO | PC 1 | PC 2 | KMO |
| RSW | 0.792 | 0.283 | 0.725 | 0.898 | −0.184 | 0.870 |
| RSD | 0.842 | 0.429 | 0.715 | 0.979 | 0.049 | 0.557 |
| CHA | 0.927 | 0.347 | 0.699 | 0.968 | −0.082 | 0.575 |
| TRL | 0.735 | 0.570 | 0.621 | 0.991 | −0.007 | 0.551 |
| RBC | 0.883 | −0.121 | 0.757 | 0.946 | 0.184 | 0.658 |
| TRV | 0.802 | 0.384 | 0.720 | 0.906 | −0.255 | 0.633 |
| FSRL | 0.798 | 0.084 | 0.667 | 0.888 | 0.182 | 0.753 |
| SRA | 0.278 | 0.438 | 0.499 | −0.011 | −0.682 | 0.243 |
| LA | 0.548 | 0.602 | 0.623 | 0.872 | −0.040 | 0.530 |
| LN | 0.738 | −0.416 | 0.621 | 0.008 | 0.936 | 0.316 |
| RDM | 0.942 | −0.267 | 0.746 | 0.622 | 0.762 | 0.441 |
| SDM | 0.824 | −0.187 | 0.633 | −0.270 | 0.874 | 0.542 |
| RSM | 0.897 | −0.351 | 0.630 | 0.955 | 0.085 | 0.676 |
| RMR | 0.885 | −0.338 | 0.645 | 0.966 | 0.066 | 0.551 |
| SMR | −0.885 | 0.338 | 0.645 | −0.965 | −0.063 | 0.556 |
| RTD | 0.761 | −0.553 | 0.823 | −0.369 | 0.882 | 0.453 |
| Eigenvalue | 10.233 | 2.369 | − | 10.319 | 3.656 | − |
| Variability (%) | 63.953 | 14.807 | − | 64.497 | 22.848 | − |
| Cumulative (%) | 63.953 | 78.760 | − | 64.497 | 87.345 | − |
| Accession | District | Village | Latitude and Longitude | STI | Seed Coat Colour |
|---|---|---|---|---|---|
| WWM-09 | Capricorn | Moletjie-Moshate | 23°36′55.9″ S 29°16′03.7″ E | 0.452 | nut brown |
| WWM-15 | Capricorn | Turfloop | 23°53′12.2″ S 29°44′52.2″ E | 0.417 | tomato red |
| WWM-37(2) | Capricorn | Ga-Molepo | 24°01′11.1″ S 29°47′05.0″ E | 0.392 | purple violet |
| WWM-39 | Capricorn | Ga-Mphela | 23°43′19.2″ S 29°12′01.4″ E | 0.431 | ruby red |
| WWM-41(A) | Sekhukhune | Nebo | 24°54′09.1″ S 29°46′15.8″ E | 0.434 | purple red |
| WWM-46 | Sekhukhune | Nebo | 24°54′07.2″ S 29°46′13.2″ E | 0.459 | signal red |
| WWM-64 | Capricorn | Ga-Mphela | 23°39′46.0″ S 29°19′16.4″ E | 0.438 | golden yellow |
| WWM-68 | Capricorn | Ga-Manamela | 23°43′01.7″ S 29°14′04.7″ E | 0.468 | brown-olive |
| WWM-76 | Capricorn | Ga-Manamela | 23°43′05.1″ S 29°14′01.3″ E | 0.546 | cream |
| Trait(s) | Description | Unit(s) |
|---|---|---|
| Root system width (RSW) | Maximal horizontal distribution of a root system | cm |
| Root system depth (RSD) | Maximal vertical depth of a root system | cm |
| Convex hull area (CHA) | Area of the convex hull that encompasses the root system | cm2 |
| Total root length (TRL) | Total sum of seminal and lateral root length | cm |
| Root branch count (RBC) | Number of lateral roots emerging from the primary root | - |
| Total root volume (TRV) | Total volume of the root system | cm3 |
| Leaf area (LA) | Area of the leaf | cm2 |
| Leaf number (LN) | Number of leaves | - |
| First seminal root length (FSRL) | Length of radicle (measured one day after germination) | cm |
| Seminal root angle (SRA) | Angle between the outermost left and right seminal roots | ◦ |
| Root dry mass (RDM) | Total dry mass of roots per plant | g |
| Shoot dry mass (SDM) | Total dry mass of shoots per plant | g |
| Root–shoot mass ratio (RSM) | Total root dry mass divided by shoot dry mass | - |
| Root mass ratio (RMR) | Dry mass of root divided by the total dry mass of entire plant | g g−1 |
| Shoot mass ratio (SMR) | Dry mass of shoot divided by the total dry mass of entire plant | g g−1 |
| Root tissue density (RTD) | Total root dry mass divided by root volume | g cm−3 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mandizvo, T.; Odindo, A.O.; Mashilo, J.; Sibiya, J.; Beck-Pay, S.L. Phenotypic Variability of Root System Architecture Traits for Drought Tolerance among Accessions of Citron Watermelon (Citrullus lanatus var. citroides (L.H. Bailey). Plants 2022, 11, 2522. https://doi.org/10.3390/plants11192522
Mandizvo T, Odindo AO, Mashilo J, Sibiya J, Beck-Pay SL. Phenotypic Variability of Root System Architecture Traits for Drought Tolerance among Accessions of Citron Watermelon (Citrullus lanatus var. citroides (L.H. Bailey). Plants. 2022; 11(19):2522. https://doi.org/10.3390/plants11192522
Chicago/Turabian StyleMandizvo, Takudzwa, Alfred Oduor Odindo, Jacob Mashilo, Julia Sibiya, and Sascha Lynn Beck-Pay. 2022. "Phenotypic Variability of Root System Architecture Traits for Drought Tolerance among Accessions of Citron Watermelon (Citrullus lanatus var. citroides (L.H. Bailey)" Plants 11, no. 19: 2522. https://doi.org/10.3390/plants11192522
APA StyleMandizvo, T., Odindo, A. O., Mashilo, J., Sibiya, J., & Beck-Pay, S. L. (2022). Phenotypic Variability of Root System Architecture Traits for Drought Tolerance among Accessions of Citron Watermelon (Citrullus lanatus var. citroides (L.H. Bailey). Plants, 11(19), 2522. https://doi.org/10.3390/plants11192522