

Rapid Quantification Method for Yield, Calorimetric Energy and Chlorophyll a Fluorescence Parameters in Nicotiana tabacum L. Using Vis-NIR-SWIR Hyperspectroscopy



Abstract
:1. Introduction
2. Results
2.1. Descriptive Analysis
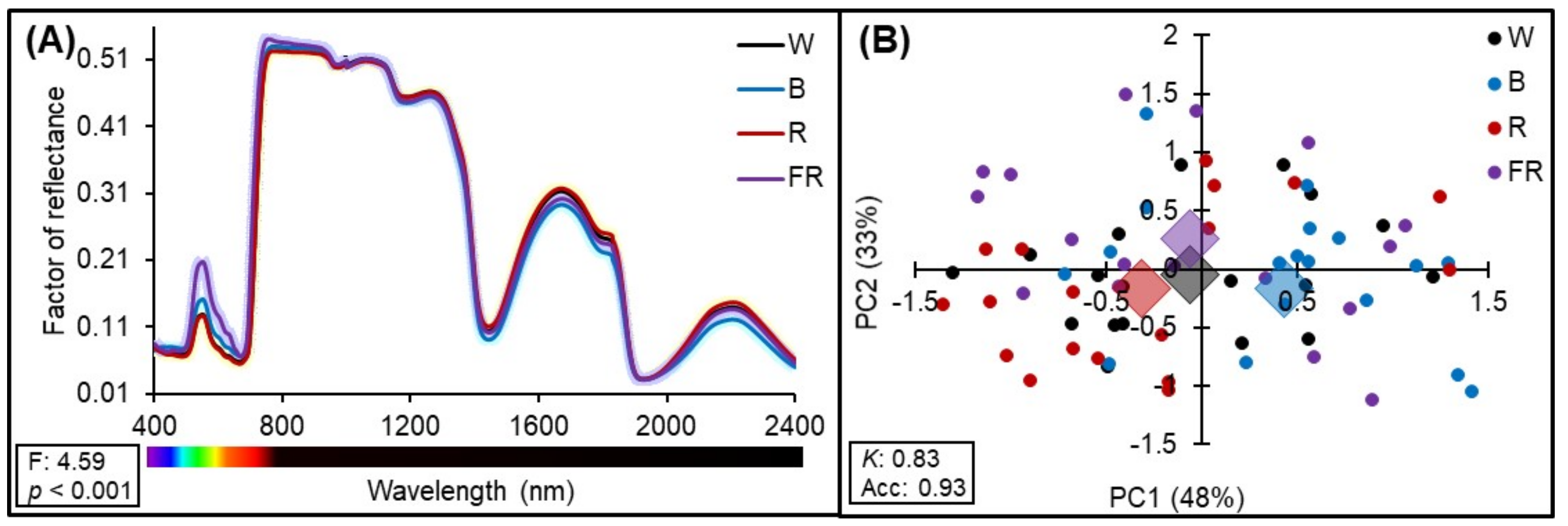
2.2. Hyperspectral Reflectance Analysis
2.3. Principal Component Analysis (PCA)
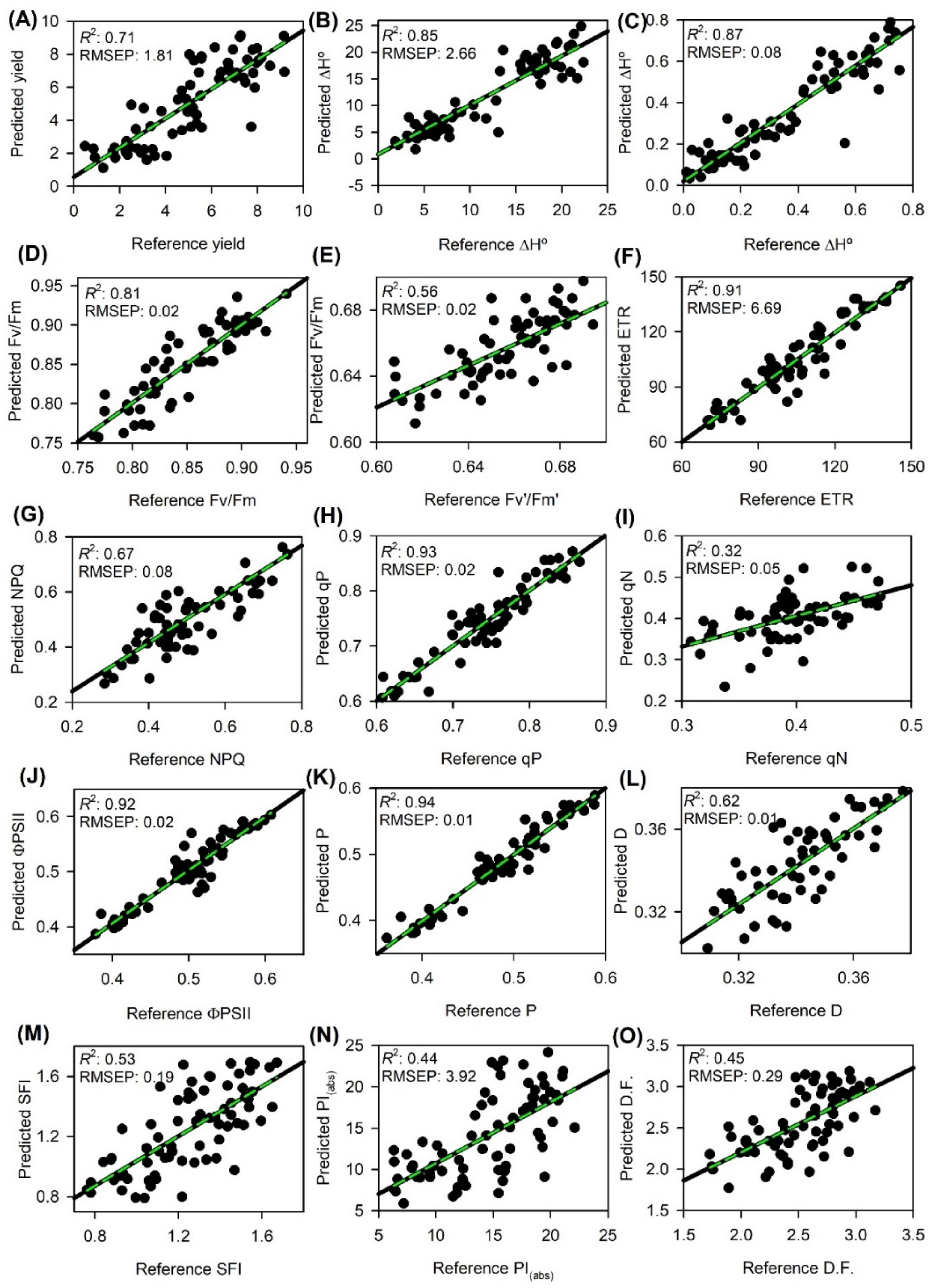
2.4. Prediction of Yield, Energy Calorific and ChlF Parameters
2.5. Regression Coefficients (RCs) and Variable Importance in Projection (VIP)
3. Discussion
3.1. Descriptive Analysis
3.2. Analysis of the Reflectance Spectrum
3.3. Partial Least Squares Regression (PLSR)
3.4. Regression Coefficients
3.5. Benefits and Limitations of Using Vis-NIR-SWIR Spectroscopy for Monitoring ChlF
4. Materials and Methods
4.1. Plant Material, Growth Conditions and Experimental Design
4.2. Growth and Yield Analysis
4.3. Calorimetric Analysis
4.4. Infrared Gas Exchange and Chlorophyll A Fluorescence Analyses Parameters
4.5. OJIP Chlorophyll a Fluorescence Transient
4.6. Hyperspectral Optical Leaf Properties
4.7. Statistical Analyses
4.7.1. Descriptive Analysis
4.7.2. Statistical Analyses of the Leaf Spectral Signature
4.7.3. Principal Component Analysis (PCA)
4.7.4. Partial Least Squares Regression (PLSR) Analysis of Reflectance Data
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations List
References
- Casal, J.J. Shade Avoidance. Arab. Book 2012, 10, e0157. [Google Scholar] [CrossRef] [PubMed]
- Falcioni, R.; Moriwaki, T.; Bonato, C.M.; Souza, L.A.; Nanni, M.R.; Antunes, W.C. Distinct Growth Light and Gibberellin Regimes Alter Leaf Anatomy and Reveal Their in Fluence on Leaf Optical Properties. Environ. Exp. Bot. 2017, 140, 86–95. [Google Scholar] [CrossRef]
- Braga, P.; Crusiol, L.G.T.; Nanni, M.R.; Caranhato, A.L.H.; Fuhrmann, M.B.; Nepomuceno, A.L.; Neumaier, N.; Farias, J.R.B.; Koltun, A.; Gonçalves, L.S.A.; et al. Vegetation Indices and NIR-SWIR Spectral Bands as a Phenotyping Tool for Water Status Determination in Soybean. Precis. Agric. 2021, 22, 249–266. [Google Scholar] [CrossRef]
- Falcioni, R.; Moriwaki, T.; Pattaro, M.; Furlanetto, R.H.; Nanni, M.R.; Antunes, W.C. High Resolution Leaf Spectral Signature as a Tool for Foliar Pigment Estimation Displaying Potential for Species Differentiation. J. Plant Physiol. 2020, 249, 153161. [Google Scholar] [CrossRef] [PubMed]
- Furlanetto, R.H.; Moriwaki, T.; Falcioni, R.; Pattaro, M.; Vollmann, A.; Sturion Junior, A.C.; Antunes, W.C.; Nanni, M.R. Hyperspectral Reflectance Imaging to Classify Lettuce Varieties by Optimum Selected Wavelengths and Linear Discriminant Analysis. Remote Sens. Appl. Soc. Environ. 2020, 20, 100400. [Google Scholar] [CrossRef]
- Fraser, D.P.; Hayes, S.; Franklin, K.A. Photoreceptor Crosstalk in Shade Avoidance. Curr. Opin. Plant Biol. 2016, 33, 1–7. [Google Scholar] [CrossRef]
- Cotrozzi, L.; Lorenzini, G.; Nali, C.; Pellegrini, E.; Saponaro, V.; Hoshika, Y.; Arab, L.; Rennenberg, H.; Paoletti, E. Hyperspectral Reflectance of Light-Adapted Leaves Can Predict Both Dark- and Light-Adapted Chl Fluorescence Parameters, and the Effects of Chronic Ozone Exposure on Date Palm (Phoenix dactylifera). Int. J. Mol. Sci. 2020, 21, 641. [Google Scholar] [CrossRef]
- Kume, A. Importance of the Green Color, Absorption Gradient, and Spectral Absorption of Chloroplasts for the Radiative Energy Balance of Leaves. J. Plant Res. 2018, 131, 501–514. [Google Scholar] [CrossRef]
- Murchie, E.H.; Lawson, T. Chlorophyll Fluorescence Analysis: A Guide to Good Practice and Understanding Some New Applications. J. Exp. Bot. 2013, 64, 3983–3998. [Google Scholar] [CrossRef]
- Natali, A.; Gruber, J.M.; Dietzel, L.; Stuart, M.C.A.; Van Grondelle, R.; Croce, R. Light-Harvesting Complexes (LHCs) Cluster Spontaneously in Membrane Environment Leading to Shortening of Their Excited State Lifetimes. J. Biol. Chem. 2016, 291, 16730–16739. [Google Scholar] [CrossRef] [Green Version]
- Jia, M.; Li, D.; Colombo, R.; Wang, Y.; Wang, X.; Cheng, T.; Zhu, Y.; Yao, X.; Xu, C.; Ouer, G.; et al. Quantifying Chlorophyll Fluorescence Parameters from Hyperspectral Reflectance at the Leaf Scale under Various Nitrogen Treatment Regimes in Winter Wheat. Remote Sens. 2019, 11, 2838. [Google Scholar] [CrossRef]
- Lysenko, V. Fluorescence Kinetic Parameters and Cyclic Electron Transport in Guard Cell Chloroplasts of Chlorophyll-Deficient Leaf Tissues from Variegated Weeping Fig (Ficus benjamina L.). Planta 2012, 235, 1023–1033. [Google Scholar] [CrossRef] [PubMed]
- Shurygin, B.; Chivkunova, O.; Solovchenko, O.; Solovchenko, A.; Dorokhov, A.; Smirnov, I.; Astashev, M.E.; Khort, D. Comparison of the Non-Invasive Monitoring of Fresh-Cut Lettuce Condition with Imaging Reflectance Hyperspectrometer and Imaging PAM-Fluorimeter. Photonics 2021, 8, 425. [Google Scholar] [CrossRef]
- Sukhova, E.; Sukhov, V. Relation of Photochemical Reflectance Indices Based on Differentwavelengths to the Parameters of Light Reactions in Photosystems I and II in Pea Plants. Remote Sens. 2020, 12, 1312. [Google Scholar] [CrossRef]
- Gupta, R. Tissue Specific Disruption of Photosynthetic Electron Transport Rate in Pigeonpea (Cajanus cajan L.) under Elevated Temperature. Plant Signal. Behav. 2019, 14, 1601952. [Google Scholar] [CrossRef]
- Kakuszi, A.; Sárvári, É.; Solti, Á.; Czégény, G.; Hideg, É.; Hunyadi-Gulyás, É.; Bóka, K.; Böddi, B. Light Piping Driven Photosynthesis in the Soil: Low-Light Adapted Active Photosynthetic Apparatus in the under-Soil Hypocotyl Segments of Bean (Phaseolus vulgaris). J. Photochem. Photobiol. B Biol. 2016, 161, 422–429. [Google Scholar] [CrossRef] [PubMed]
- Kalaji, H.M.; Račková, L.; Paganová, V.; Swoczyna, T.; Rusinowski, S.; Sitko, K. Can Chlorophyll-a Fluorescence Parameters Be Used as Bio-Indicators to Distinguish between Drought and Salinity Stress in Tilia cordata Mill? Environ. Exp. Bot. 2018, 152, 149–157. [Google Scholar] [CrossRef]
- Maxwell, K.; Johnson, G.N. Chlorophyll Fluorescence—A Practical Guide. J. Exp. Bot. 2000, 51, 659–668. [Google Scholar] [CrossRef]
- Bag, P.; Chukhutsina, V.; Zhang, Z.; Paul, S.; Ivanov, A.G.; Shutova, T.; Croce, R.; Holzwarth, A.R.; Jansson, S. Direct Energy Transfer from Photosystem II to Photosystem I Confers Winter Sustainability in Scots Pine. Nat. Commun. 2020, 11, 6388. [Google Scholar] [CrossRef]
- Bellasio, C.; Beerling, D.J.; Griffiths, H. An Excel Tool for Deriving Key Photosynthetic Parameters from Combined Gas Exchange and Chlorophyll FLuorescence: Theory and Practice. Plant Cell Environ. 2016, 39, 1164–1179. [Google Scholar] [CrossRef] [Green Version]
- Ripoll, J.; Bertin, N.; Bidel, L.P.R.; Urban, L. A User’s View of the Parameters Derived from the Induction Curves of Maximal Chlorophyll a Fluorescence: Perspectives for Analyzing Stress. Front. Plant Sci. 2016, 7, 1679. [Google Scholar] [CrossRef] [PubMed]
- Boshkovski, B.; Doupis, G.; Zapolska, A.; Kalaitzidis, C.; Koubouris, G. Hyperspectral Imagery Detects Water Deficit and Salinity Effects on Photosynthesis and Antioxidant Enzyme Activity of Three Greek Olive Varieties. Sustainability 2022, 14, 1432. [Google Scholar] [CrossRef]
- Guidi, L.; Lo Piccolo, E.; Landi, M. Chlorophyll Fluorescence, Photoinhibition and Abiotic Stress: Does It Make Any Difference the Fact to Be a C3 or C4 Species? Front. Plant Sci. 2019, 10, 174. [Google Scholar] [CrossRef] [PubMed]
- Lysenko, V.; Guo, Y.; Kosolapov, A.; Usova, E.; Varduny, T.; Krasnov, V. Polychromatic Fourier-PAM Fluorometry and Hyperspectral Analysis of Chlorophyll Fluorescence from Phaseolus vulgaris Leaves: Effects of Green Light. Inf. Process. Agric. 2019, 7, 204–211. [Google Scholar] [CrossRef]
- Kalaji, H.M.; Jajoo, A.; Oukarroum, A.; Brestic, M.; Zivcak, M.; Samborska, I.A.; Cetner, M.D.; Łukasik, I.; Goltsev, V.; Ladle, R.J.; et al. The Use of Chlorophyll Fluorescence Kinetics Analysis to Study the Performance of Photosynthetic Machinery in Plants; Academic Press: Cambridge, MA, USA, 2014; Volume 2. [Google Scholar]
- Jin, J.; Pratama, B.A.; Wang, Q. Tracing Leaf Photosynthetic Parameters Using Hyperspectral Indices in an Alpine Deciduous Forest. Remote Sens. 2020, 12, 1124. [Google Scholar] [CrossRef]
- Strasser, R.J.; Michael, M.T. Analysis of the Fluorescence Transient Alaka Srivastava Summary II. The Theoretical Background; Springer: New York, NY, USA, 2005. [Google Scholar]
- El-Hendawy, S.; Al-Suhaibani, N.; Mubushar, M.; Tahir, M.U.; Marey, S.; Refay, Y.; Tola, E. Combining Hyperspectral Reflectance and Multivariate Regression Models to Estimate Plant Biomass of Advanced Spring Wheat Lines in Diverse Phenological Stages under Salinity Conditions. Appl. Sci. 2022, 12, 1983. [Google Scholar] [CrossRef]
- Gururani, M.A.; Venkatesh, J.; Ghosh, R.; Strasser, R.J.; Ponpandian, L.N.; Bae, H. Chlorophyll-a Fluorescence Evaluation of PEG-Induced Osmotic Stress on PSII Activity in Arabidopsis Plants Expressing SIP1. Plant Biosyst. 2018, 152, 945–952. [Google Scholar] [CrossRef]
- Xiao, W.; Wang, H.; Liu, W.; Wang, X.; Guo, Y.; Strasser, R.J.; Qiang, S.; Chen, S.; Hu, Z. Action of Alamethicin in Photosystem II Probed by the Fast Chlorophyll Fluorescence Rise Kinetics and the JIP-Test. Photosynthetica 2020, 58, 358–368. [Google Scholar] [CrossRef]
- Strasser, R.J.; Srivastava, A.; Tsimilli-Michael, M. The Fluorescence Transient as a Tool to Characterize and Screen Photosynthetic Samples. In Probing Photosynthesis: Mechanism, Regulation and Adaptation; CRC Press: Boca Raton, FL, USA, 2000; pp. 443–480. [Google Scholar]
- Lazár, D.; Nauš, J. Statistical Properties of Chlorophyll Fluorescence Induction Parameters. Photosynthetica 1998, 35, 121–127. [Google Scholar] [CrossRef]
- Ralbovsky, N.M.; Smith, J.P. Multivariate Curve Resolution for Analysis of Raman Hyperspectral Imaging Data Sets for Enzyme Immobilization. Chem. Data Collect. 2022, 38, 100835. [Google Scholar] [CrossRef]
- Sexton, T.; Sankaran, S.; Cousins, A.B. Predicting Photosynthetic Capacity in Tobacco Using Shortwave Infrared Spectral Reflectance. J. Exp. Bot. 2021, 72, 4373–4383. [Google Scholar] [CrossRef] [PubMed]
- Zheng, W.; Lu, X.; Li, Y.; Li, S.; Zhang, Y. Hyperspectral Identification of Chlorophyll Fluorescence Parameters of Suaeda Salsa in Coastal Wetlands. Remote Sens. 2021, 13, 2066. [Google Scholar] [CrossRef]
- Rodrigues, M.; Berti de Oliveira, R.; Leboso Alemparte Abrantes dos Santos, G.; Mayara de Oliveira, K.; Silveira Reis, A.; Herrig Furlanetto, R.; Antônio Yanes Bernardo Júnior, L.; Silva Coelho, F.; Rafael Nanni, M. Rapid Quantification of Alkaloids, Sugar and Yield of Tobacco (Nicotiana tabacum L.) Varieties by Using Vis–NIR–SWIR Spectroradiometry. Spectrochim. Acta Part A Mol. Biomol. 2022, 274, 121082. [Google Scholar] [CrossRef] [PubMed]
- Da Silva Junior, C.A.; Nanni, M.R.; Shakir, M.; Teodoro, P.E.; de Oliveira-Júnior, J.F.; Cezar, E.; de Gois, G.; Lima, M.; Wojciechowski, J.C.; Shiratsuchi, L.S. Soybean Varieties Discrimination Using Non-Imaging Hyperspectral Sensor. Infrared Phys. Technol. 2018, 89, 338–350. [Google Scholar] [CrossRef]
- Shorten, P.R.; Leath, S.R.; Schmidt, J.; Ghamkhar, K. Predicting the Quality of Ryegrass Using Hyperspectral Imaging. Plant Methods 2019, 15, 63. [Google Scholar] [CrossRef]
- Zimmer, G.F.; Santos, R.O.; Teixeira, I.D.; Schneider, R.d.C.d.S.; Helfer, G.A.; da Costa, A. Ben Rapid Quantification of Constituents in Tobacco by NIR Fiber-Optic Probe. J. Chemom. 2020, 34, e3303. [Google Scholar] [CrossRef]
- Kováč, D.; Veselovská, P.; Klem, K.; Večeřová, K.; Ač, A.; Peñuelas, J.; Urban, O. Potential of Photochemical Reflectance Index for Indicating Photochemistry and Light Use Efficiency in Leaves of European Beech and Norway Spruce Trees. Remote Sens. 2018, 10, 1202. [Google Scholar] [CrossRef]
- Pimentel-Gomes, F. Statistics Course Experimental, 1st ed.; FEALQ: Piracicaba, Brazil, 2009. [Google Scholar]
- Coast, O.; Shah, S.; Ivakov, A.; Gaju, O.; Wilson, P.B.; Posch, B.C.; Bryant, C.J.; Negrini, A.C.A.; Evans, J.R.; Condon, A.G.; et al. Predicting Dark Respiration Rates of Wheat Leaves from Hyperspectral Reflectance. Plant Cell Environ. 2019, 42, 2133–2150. [Google Scholar] [CrossRef]
- Nogales-Bueno, J.; Baca-Bocanegra, B.; Rooney, A.; Hernández-Hierro, J.M.; Heredia, F.J.; Byrne, H.J. Linking ATR-FTIR and Raman Features to Phenolic Extractability and Other Attributes in Grape Skin. Talanta 2017, 167, 44–50. [Google Scholar] [CrossRef]
- Schansker, G.; Tóth, S.Z.; Strasser, R.J. Dark Recovery of the Chl a Fluorescence Transient (OJIP) after Light Adaptation: The QT-Component of Non-Photochemical Quenching Is Related to an Activated Photosystem I Acceptor Side. Biochim. Biophys. Acta Bioenerg. 2006, 1757, 787–797. [Google Scholar] [CrossRef] [Green Version]
- Calviño-Cancela, M.; Martín-Herrero, J. Spectral Discrimination of Vegetation Classes in Ice-Free Areas of Antarctica. Remote Sens. 2016, 8, 856. [Google Scholar] [CrossRef]
- Horikawa, Y.; Mizuno-Tazuru, S.; Sugiyama, J. Near-Infrared Spectroscopy as a Potential Method for Identification of Anatomically Similar Japanese Diploxylons. J. Wood Sci. 2015, 61, 251–261. [Google Scholar] [CrossRef]
- Locascio, A.; Blázquez, M.A.; Alabadí, D. Dynamic Regulation of Cortical Microtubule Organization through Prefoldin-DELLA Interaction. Curr. Biol. 2013, 23, 804–809. [Google Scholar] [CrossRef] [PubMed]
- Zhong, R.; Burk, D.; Morrison, W., III; Ye, Z.-H. A Kinesin-like Protein Is Essential for Oriented Deposition of Cellulose Microfibrils and Cell Wall Strength. Plant Cell 2002, 14, 3101–3117. [Google Scholar] [CrossRef]
- Baranović, G.; Šegota, S. Infrared Spectroscopy of Flavones and Flavonols. Reexamination of the Hydroxyl and Carbonyl Vibrations in Relation to the Interactions of Flavonoids with Membrane Lipids. Spectrochim. Acta Part A Mol. Biomol. Spectrosc. 2018, 192, 473–486. [Google Scholar] [CrossRef] [PubMed]
- Sobejano-Paz, V.; Mikkelsen, T.N.; Baum, A.; Mo, X.; Liu, S.; Köppl, C.J.; Johnson, M.S.; Gulyas, L.; García, M. Hyperspectral and Thermal Sensing of Stomatal Conductance, Transpiration, and Photosynthesis for Soybean and Maize under Drought. Remote Sens. 2020, 12, 3182. [Google Scholar] [CrossRef]
- Barnes, M.L.; Breshears, D.D.; Law, D.J.; van Leeuwen, W.J.D.; Monson, R.K.; Fojtik, A.C.; Barron-Gafford, G.A.; Moore, D.J.P. Beyond Greenness: Detecting Temporal Changes in Photosynthetic Capacity with Hyperspectral Reflectance Data. PLoS ONE 2017, 12, e0189539. [Google Scholar] [CrossRef]
- Jin, J.; Wang, Q. Selection of Informative Spectral Bands for PLS Models to Estimate Foliar Chlorophyll Content Using Hyperspectral Reflectance. IEEE Trans. Geosci. Remote Sens. 2019, 57, 3064–3072. [Google Scholar] [CrossRef]
- Fernandes, A.M.; Fortini, E.A.; Müller, L.A.d.C.; Batista, D.S.; Vieira, L.M.; Silva, P.O.; do Amaral, C.H.; Poethig, R.S.; Otoni, W.C. Leaf Development Stages and Ontogenetic Changes in Passionfruit (Passiflora edulis Sims.) Are Detected by Narrowband Spectral Signal. J. Photochem. Photobiol. B Biol. 2020, 209, 111931. [Google Scholar] [CrossRef]
- Asner, G.P.; Jones, M.O.; Martin, R.E.; Knapp, D.E.; Hughes, R.F. Remote Sensing of Native and Invasive Species in Hawaiian Forests. Remote Sens. Environ. 2008, 112, 1912–1926. [Google Scholar] [CrossRef]
- Zar, J.H. Biostatistical Analysis, 5th ed.; Pearson Education: Upper Saddle River, NJ, USA, 2010; ISBN 0-13-100846-3. [Google Scholar]
- Samborska, I.A.; Kalaji, H.M.; Sieczko, L.; Borucki, W.; Mazur, R.; Kouzmanova, M.; Goltsev, V. Can Just One-Second Measurement of Chlorophyll a Fluorescence Be Used to Predict Sulphur Deficiency in Radish (Raphanus sativus L. Sativus) Plants? Curr. Plant Biol. 2019, 19, 100096. [Google Scholar] [CrossRef]
- Moriwaki, T.; Falcioni, R.; Tanaka, F.A.O.; Cardoso, K.A.K.; Souza, L.A.; Benedito, E.; Nanni, M.R.; Bonato, C.M.; Antunes, W.C. Nitrogen-Improved Photosynthesis Quantum Yield Is Driven by Increased Thylakoid Density, Enhancing Green Light Absorption. Plant Sci. 2019, 278, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Falcioni, R.; Moriwaki, T.; Perez-Llorca, M.; Munné-Bosch, S.; Gibin, M.S.M.S.; Sato, F.; Pelozo, A.; Pattaro, M.C.M.C.; Giacomelli, M.E.; Rüggeberg, M.; et al. Cell Wall Structure and Composition Is Affected by Light Quality in Tomato Seedlings. J. Photochem. Photobiol. B Biol. 2020, 203, 111745. [Google Scholar] [CrossRef] [PubMed]
- Falcioni, R.; Moriwaki, T.; Furlanetto, R.H.; Nanni, M.R.; Antunes, W.C. Simple, Fast and Efficient Methods for Analysing the Structural, Ultrastructural and Cellular Components of the Cell Wall. Plants 2022, 11, 995. [Google Scholar] [CrossRef] [PubMed]
- Chicati, M.S.; Nanni, M.R.; Chicati, M.L.; Furlanetto, R.H.; Cezar, E.; De Oliveira, R.B. Hyperspectral Remote Detection as an Alternative to Correlate Data of Soil Constituents. Remote Sens. Appl. Soc. Environ. 2019, 16, 100270. [Google Scholar] [CrossRef]
- Falcioni, R.; Moriwaki, T.; Benedito, E.; Bonato, C.M.; de Souza, L.A.; Antunes, W.C. Increased Gibberellin Levels Enhance Light Capture Efficiency in Tobacco Plants and Promote Dry Matter Accumulation. Theor. Exp. Plant Physiol. 2018, 30, 235–250. [Google Scholar] [CrossRef]
- Ge, Y.; Atefi, A.; Zhang, H.; Miao, C.; Ramamurthy, R.K. High-Throughput Analysis of Leaf Physiological and Chemical Traits with VIS–NIR–SWIR Spectroscopy: A Case Study with a Maize Diversity Panel. Plant Methods 2019, 15, 66. [Google Scholar] [CrossRef]
- Thornley, R.H.; Gerard, F.F.; White, K.; Verhoef, A. Intra-Annual Taxonomic and Phenological Drivers of Spectral Variance in Grasslands. Remote Sens. Environ. 2022, 271, 112908. [Google Scholar] [CrossRef]
- Thornley, R.H.; Verhoef, A.; Gerard, F.F.; White, K. The Feasibility of Leaf Reflectance-Based Taxonomic Inventories and Diversity Assessments of Species-Rich Grasslands: A Cross-Seasonal Evaluation Using Waveband Selection. Remote Sens. 2022, 14, 2310. [Google Scholar] [CrossRef]
- Prats-Mateu, B.; Felhofer, M.; de Juan, A.; Gierlinger, N. Multivariate Unmixing Approaches on Raman Images of Plant Cell Walls: New Insights or Overinterpretation of Results? Plant Methods 2018, 14, 52. [Google Scholar] [CrossRef] [Green Version]
- Nanni, M.R.; Cezar, E.; da Silva Junior, C.A.; Silva, G.F.C.; da Silva Gualberto, A.A. Partial Least Squares Regression (PLSR) Associated with Spectral Response to Predict Soil Attributes in Transitional Lithologies. Arch. Agron. Soil Sci. 2018, 64, 682–695. [Google Scholar] [CrossRef]
- Hoffmann, W.A.; Poorter, H. Avoiding Bias in Calculations of Relative Growth Rate. Ann. Bot. 2002, 90, 37–42. [Google Scholar] [CrossRef]
- Baker, N.R. Chlorophyll Fluorescence: A Probe of Photosynthesis in Vivo. Annu. Rev. Plant Biol. 2008, 59, 89–113. [Google Scholar] [CrossRef] [PubMed]
- Licor, Inc. Using the LI-6800 Portable Photosynthesis System, 4th ed.; Licor, Inc.: Lincoln, NE, USA, 2019. [Google Scholar]
- Sitko, K.; Rusinowski, S.; Kalaji, H.M.; Szopiński, M.; Małkowski, E. Photosynthetic Efficiency as Bioindicator of Environmental Pressure in A. Halleri. Plant Physiol. 2017, 175, 290–302. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Strasser, R.J.; Srivastava, A.; Tsimilli-Michael, M. Analysis of the Fluorescence Transiet as a Tool to Characterize and Screen Photosynthetic Samples. Chlorophyll Fluoresc. Signat. Photosynth. 2004, 443–480. [Google Scholar]
- Minasny, B.; McBratney, A.B. Digital Mapping of Soil Carbon. Adv. Agron. 2013, 3, 4. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | Count (n) | Mean | Median | Minimum | Maximum | CV (%) |
|---|---|---|---|---|---|---|
| Yield (g plant−1) | 260 | 4.98 | 4.86 | 1.12 | 9.13 | 48.77 |
| ΔH° (kcal g−1) | 260 | 11.27 | 8.03 | 1.80 | 24.88 | 58.98 |
| ΔH° (kcal m−2) | 260 | 0.36 | 0.27 | 0.03 | 0.84 | 65.04 |
| Fv/Fm | 260 | 0.86 | 0.86 | 0.76 | 0.95 | 6.00 |
| Fv’/Fm’ | 260 | 0.66 | 0.66 | 0.59 | 0.71 | 3.70 |
| ETR | 260 | 104.54 | 99.29 | 69.23 | 145.21 | 19.43 |
| NPQ | 260 | 0.50 | 0.49 | 0.20 | 0.85 | 30.17 |
| qP | 260 | 0.74 | 0.74 | 0.60 | 0.87 | 9.90 |
| qN | 260 | 0.40 | 0.40 | 0.23 | 0.55 | 15.77 |
| ΦPSII | 260 | 0.51 | 0.50 | 0.39 | 0.60 | 11.53 |
| P | 260 | 0.49 | 0.48 | 0.37 | 0.59 | 12.03 |
| D | 260 | 0.34 | 0.34 | 0.29 | 0.41 | 7.03 |
| SFI | 260 | 1.24 | 1.28 | 0.79 | 1.69 | 21.87 |
| PI(abs) | 260 | 14.00 | 12.80 | 5.89 | 24.16 | 35.79 |
| D.F. | 260 | 2.57 | 2.55 | 1.77 | 3.18 | 14.34 |
| Parameter | Maximum Factors PLS | Calibration | Cross-Validation | ||||||
|---|---|---|---|---|---|---|---|---|---|
| R2 | Offset | RMSE | RPD | R2 | Offset | RMSE | RPD | ||
| Yield (g plant−1) | 5 | 0.85 | 0.75 | 0.93 | 2.61 | 0.84 | 0.82 | 1.04 | 2.5 |
| ΔH° (kcal g−1) | 11 | 0.91 | 0.98 | 1.97 | 3.37 | 0.91 | 1.11 | 2.20 | 3.3 |
| ΔH° (kcal m−2) | 10 | 0.93 | 0.03 | 0.06 | 3.81 | 0.92 | 0.03 | 0.08 | 3.5 |
| Fv/Fm | 7 | 0.82 | 0.16 | 0.02 | 2.36 | 0.76 | 0.21 | 0.03 | 2.0 |
| Fv’/Fm’ | 8 | 0.84 | 0.12 | 0.01 | 2.50 | 0.70 | 0.20 | 0.01 | 1.8 |
| ETR | 8 | 0.91 | 10.55 | 6.33 | 3.33 | 0.86 | 14.47 | 8.06 | 2.7 |
| NPQ | 2 | 0.62 | 0.18 | 0.08 | 1.63 | 0.61 | 0.19 | 0.09 | 1.6 |
| qP | 7 | 0.93 | 0.07 | 0.02 | 3.78 | 0.91 | 0.09 | 0.03 | 3.3 |
| qN | 3 | 0.29 | 0.29 | 0.05 | 1.19 | 0.22 | 0.32 | 0.05 | 1.1 |
| ΦPSII | 8 | 0.91 | 0.04 | 0.02 | 3.39 | 0.91 | 0.06 | 0.02 | 3.3 |
| P | 8 | 0.94 | 0.03 | 0.01 | 4.12 | 0.91 | 0.04 | 0.02 | 3.4 |
| D | 6 | 0.72 | 0.10 | 0.01 | 1.89 | 0.64 | 0.12 | 0.01 | 1.7 |
| SFI | 6 | 0.70 | 0.38 | 0.15 | 1.81 | 0.64 | 0.45 | 0.17 | 1.7 |
| PI(abs) | 6 | 0.61 | 5.72 | 3.27 | 1.59 | 0.54 | 6.61 | 3.72 | 1.5 |
| D.F. | 5 | 0.65 | 0.92 | 0.21 | 1.68 | 0.57 | 1.12 | 0.25 | 1.5 |
| Parameter | Maximum Factors PLS | Predicted | ||||
|---|---|---|---|---|---|---|
| R2 | Offset | SEP | RPD | Linear Equation Prediction to Calibration Model (R2P) | ||
| Yield (g plant−1) | 5 | 0.71 | 0.80 | 1.33 | 1.86 | Ŷ = 0.8875x + 0.5701 |
| ΔH° (kcal g−1) | 11 | 0.85 | 0.95 | 2.66 | 2.58 | Ŷ = 0.8224x + 0.8697 |
| ΔH° (kcal m−2) | 10 | 0.87 | 0.03 | 0.08 | 2.77 | Ŷ = 0.9344x + 0.0194 |
| Fv/Fm | 7 | 0.81 | 0.17 | 0.03 | 2.29 | Ŷ = 0.9922x + 0.0075 |
| Fv’/Fm’ | 8 | 0.56 | 0.07 | 0.01 | 1.51 | Ŷ = 0.6338x + 0.2408 |
| ETR | 8 | 0.91 | 11.38 | 6.74 | 3.33 | Ŷ = 0.9915x + 0.6146 |
| NPQ | 2 | 0.67 | 0.12 | 0.07 | 1.74 | Ŷ = 0.8813x + 0.0631 |
| qP | 7 | 0.93 | 0.07 | 0.02 | 3.78 | Ŷ = 1.0043x − 0.0021 |
| qN | 3 | 0.32 | 0.22 | 0.04 | 1.21 | Ŷ = 0.7419x + 0.1093 |
| ΦPSII | 8 | 0.92 | 0.04 | 0.02 | 3.54 | Ŷ = 0.9657x + 0.0202 |
| P | 8 | 0.94 | 0.03 | 0.01 | 4.08 | Ŷ = 1.0065x − 0.0308 |
| D | 6 | 0.62 | 0.11 | 0.01 | 1.62 | Ŷ = 0.9154x + 0.0308 |
| SFI | 6 | 0.53 | 0.45 | 0.19 | 1.46 | Ŷ = 0.8253x + 0.2124 |
| PI(abs) | 6 | 0.44 | 6.14 | 3.93 | 1.34 | Ŷ = 0.7410x + 3.3400 |
| D.F. | 5 | 0.45 | 0.86 | 0.29 | 1.35 | Ŷ = 0.6797x + 0.8462 |
| Parameter | Selection | Most Responsive VIP by Wavelength (nm) |
|---|---|---|
| Yield (g plant−1) | 8 | 440, 550, 672, 702, 935, 1404, 1590, 1922 |
| ΔH° (kcal g−1) | 17 | 401, 454, 518, 548, 682, 702, 742, 865, 998, 1114, 1331, 1387, 1430, 1530, 1712, 1927, 2048 |
| ΔH° (kcal m−2) | 12 | 401, 544, 663, 684, 703, 743, 998, 1391, 1532, 1679, 1885, 2031 |
| Fv/Fm | 11 | 405, 482, 544, 680, 705, 752, 1143, 1406, 1594, 1894, 2197 |
| Fv’/Fm’ | 9 | 405, 484, 550, 676, 702, 730, 1419, 1440, 2192 |
| ETR | 9 | 400, 483, 534, 664, 698, 724, 1332, 1440, 1873 |
| NPQ | 5 | 402, 545, 712, 770, 1287 |
| qP | 7 | 440, 527, 668, 702, 1343, 1395, 1873 |
| qN | 5 | 550, 713, 1095, 1297, 1433 |
| ΦPSII | 6 | 405, 481, 663, 702, 723, 1327 |
| P | 8 | 403, 478, 668, 697, 730, 1332, 1874, 1920 |
| D | 7 | 405, 515, 550, 630, 698, 752, 2156 |
| SFI | 11 | 405, 445, 524, 552, 675, 701, 730, 1446, 1610, 1929, 2200 |
| PI(abs) | 9 | 405, 435, 515, 550, 668, 700, 730, 1586, 1930 |
| D.F. | 10 | 400, 405, 440, 521, 674, 701, 730, 1332, 1569, 1927 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Falcioni, R.; Moriwaki, T.; Antunes, W.C.; Nanni, M.R. Rapid Quantification Method for Yield, Calorimetric Energy and Chlorophyll a Fluorescence Parameters in Nicotiana tabacum L. Using Vis-NIR-SWIR Hyperspectroscopy. Plants 2022, 11, 2406. https://doi.org/10.3390/plants11182406
Falcioni R, Moriwaki T, Antunes WC, Nanni MR. Rapid Quantification Method for Yield, Calorimetric Energy and Chlorophyll a Fluorescence Parameters in Nicotiana tabacum L. Using Vis-NIR-SWIR Hyperspectroscopy. Plants. 2022; 11(18):2406. https://doi.org/10.3390/plants11182406
Chicago/Turabian StyleFalcioni, Renan, Thaise Moriwaki, Werner Camargos Antunes, and Marcos Rafael Nanni. 2022. "Rapid Quantification Method for Yield, Calorimetric Energy and Chlorophyll a Fluorescence Parameters in Nicotiana tabacum L. Using Vis-NIR-SWIR Hyperspectroscopy" Plants 11, no. 18: 2406. https://doi.org/10.3390/plants11182406
APA StyleFalcioni, R., Moriwaki, T., Antunes, W. C., & Nanni, M. R. (2022). Rapid Quantification Method for Yield, Calorimetric Energy and Chlorophyll a Fluorescence Parameters in Nicotiana tabacum L. Using Vis-NIR-SWIR Hyperspectroscopy. Plants, 11(18), 2406. https://doi.org/10.3390/plants11182406