Abstract
Rice production needs to be sustained in the coming decades, as the changeable climatic conditions are becoming more conducive to disease outbreaks. The majority of rice diseases cause enormous economic damage and yield instability. Among them, rice blast caused by Magnaportheoryzae is a serious fungal disease and is considered one of the major threats to world rice production. This pathogen can infect the above-ground tissues of rice plants at any growth stage and causes complete crop failure under favorable conditions. Therefore, management of blast disease is essentially required to sustain global food production. When looking at the drawback of chemical management strategy, the development of durable, resistant varieties is one of the most sustainable, economic, and environment-friendly approaches to counter the outbreaks of rice blasts. Interestingly, several blast-resistant rice cultivars have been developed with the help of breeding and biotechnological methods. In addition, 146 R genes have been identified, and 37 among them have been molecularly characterized to date. Further, more than 500 loci have been identified for blast resistance which enhances the resources for developing blast resistance through marker-assisted selection (MAS), marker-assisted backcross breeding (MABB), and genome editing tools. Apart from these, a better understanding of rice blast pathogens, the infection process of the pathogen, and the genetics of the immune response of the host plant are very important for the effective management of the blast disease. Further, high throughput phenotyping and disease screening protocols have played significant roles in easy comprehension of the mechanism of disease spread. The present review critically emphasizes the pathogenesis, pathogenomics, screening techniques, traditional and molecular breeding approaches, and transgenic and genome editing tools to develop a broad spectrum and durable resistance against blast disease in rice. The updated and comprehensive information presented in this review would be definitely helpful for the researchers, breeders, and students in the planning and execution of a resistance breeding program in rice against this pathogen.
1. Introduction
Rice (Oryza sativa L.) is the primary staple food and main source of nutrition for 2.5 to 3.5 billion people in the World, especially those living in fast-growing low-income countries [1,2,3]. Moreover, it is the main source of income and employment for more than 200 million populations across the world [2,4]. Over time, rice production has been augmented significantly; though, it is insufficient to fulfill the demand of the increasing global population [5,6], which needs to be increased by 38% by 2030 with the limited arable lands and severity of biotic and abiotic factors [1,7,8]. It is well understood that rice production is severely constrained by various biotic factors (pests, weeds, diseases, etc.) and abiotic factors (drought, cold, acidity, heat, salinity, etc.) [7]. However, biotic factors, especially diseases, have a great impact on rice production, which restricts rice breeders from crossing the yield barriers. It has been estimated that among the various diseases in rice, fungal diseases can decrease the global annual rice production by 14% [1,9,10]. Factually, out of the various fungal diseases, rice blast caused by Magnaporthe oryzae is one of the most devastating and recurring diseases causing enormous yield losses up to 70% to 80% within a short span of time [11,12,13]. Based on the scientific and economic importance of this fungal pathogen, it has been placed in the top ten fungal pathogens in the world [9,14]. Interestingly, the fungus Magnoporthe oryzae obtains a wide spectrum in affecting the rice plant, right from the seedling to late vegetative stages involving leaves, nodes, collar, panicles, panicle neck, and roots. The pathogen produces eye-shaped lesions on leaves (leaf blast), leaf collars (collar blast), culms, culm nodes, panicle neck nodes (neck rot), and panicles (panicle blast), which vary in color and shape depending on varietal resistance, environmental conditions, and age of plants [12,14].
When looking at the destructive nature of the disease, utmost emphasis should be given to managing the infestation of blast disease with the help of various innovative and durable techniques and strategies such as the development of resistant cultivars having durable resistance, use of fungicides, optimum fertilizer doses and appropriate planting time, etc. Moreover, among these, breeding for disease-resistant rice varieties possessing a major resistance gene (R-gene) is the most efficient and sustainable approach to counter disease outbreaks due to its economic and environmental advantage [2,4,15]. Several strategies, viz., conventional breeding, mutation breeding, marker-assisted breeding, transgenic approaches, genome editing tools, etc., have been adopted so far for the development of disease-resistant cultivars [16], of which conventional breeding methods, including the pedigree method, backcross breeding, introduction and acclimatization, multiline breeding, pureline selection, recurrent selection, etc., are the robust and mostly used for developing novel genetic variants for blast resistance [17,18,19]. International Rice Research Institute (IRRI), Manila, Philipines, has developed many blast-resistant lines/donors for blast resistance through conventional breeding, which have made a significant impact on reducing the prevalence of blasts throughout the World [12,20]. Genetically diverse rice landraces with broad-spectrum resistance are valuable sources for the introgression of the resistance genes into rice cultivars for the control of blast, but the number is limited. Therefore, artificially induced mutants are an important resource for identifying new broad-spectrum resistant (R) genes/loci. In this aspect, physical/chemical mutagenesis is a way to develop new R genes/alleles, which can generate desirable resistant mutations that are free from the association of undesirable traits or linkage drag [21]. However, due to the quantitative nature of this resistance, conventional breeding takes a long time and requires many generations of crossing and screening to test the resistance, which can be overcome by adopting advanced molecular breeding methods and biotechnological tools [16].
With the advancement in plant genomics, breeders now have a wide spectrum of biotechnological tools to further strengthen the process of screening and developing blast-resistant rice varieties. Identification and mapping of a specific R gene for differential blast races in diverse elite germplasm/mapping populations with the help of DNA markers through association mapping/quantitative trail loci (QTL) mapping is a crucial step in ensuring the accuracy of their utilization in marker-assisted breeding (MAB) [15,16,22]. Until now, about 146 R-genes for rice blast resistance have been identified and mapped from both indica and japonica subspecies of rice, and about 36 genes have been molecularly characterized and cloned so far, which have boosted the breeding strategies for blast resistance in rice [15,23,24]. Marker-assisted gene pyramiding (MAGP) is one of the most appropriate approaches to confer horizontal resistance in the genotype for different races of blast pathogen by introgression of more than one resistance (R) gene [25,26]. Similarly, marker-assisted backcross breeding (MABB) is also an important method to transfer blast resistance in otherwise high-yielding rice varieties keeping all the characters of the same recipient parent. Marker-based selection methods are more accurate, reliable, and time-saving, do not require appropriate disease-favoring environmental conditions, and select the resistant genotypes even without inoculation of the pathogen [22,23,24]. Hence, these methods have been widely used by many plant breeders to develop blast-resistant varieties Worldwide [27].
Furthermore, genetic engineering techniques have enabled plant breeders to transfer blast-resistant genes from one organism to the background of other elite cultivars for developing resistant varieties [28,29,30,31]. However, the product of this method has not been widely accepted due to various safety and regulatory issues [29]. Recently genome editing technologies offer expanded potential for crop improvement as they allow specific alterations in DNA sequences that can be performed in vivo. These editing tools precisely manipulate specific sequences in the genome, which allows the insertion, deletion or substitution of nucleotides in specific genes or sequences [32,33]. Several genomic editing techniques, such as meganucleases (MNs), Zinc-finger nucleases (ZFNs), Transcription Activator–like Effector Nucleases (TALENs), and Clustered Regularly Interspaced Short Palindrome Repeats/CRISPR-associated protein 9 (CRISPR/Cas9), are used to promote specific genetic modifications. The development of the CRISPR/Cas9 technique has opened up a wide range of applications and could be explored in improving plant resistance to pathogens [34].
Breeding for disease resistance in rice is a critical component for increasing rice production and ensuring food security. Long-lasting and durable resistance for rice blast from a single gene is feasible but not often durable, as the pathogen can rapidly mutate and attack resistant cultivars. Broad host range, continuous genetic variation, evolution, and host shifts are the main reasons for the evolution of different types of races of Magnaporthe. It has made the development of resistant cultivars a daunting task. Therefore, the rice Magnaporthe interaction pathosystem has emerged as a model system to study host–pathogen interaction, genetics of rice blast resistance, host plant resistance, favorable conditions for disease occurrence, screening protocols, and conventional and modern breeding strategies to develop blast-resistant cultivars.
2. Economic Impacts of Rice Blast
Rice blast is one of the extensively dispersed plant diseases of a major food crop with huge destructive nature in Asian and African countries. It gradually becomes more troublesome to rice-growing farmers and threatens food security worldwide [35]. Therefore, significant efforts should be made to manage this disease to sustain the food security and economy of the World. The disease was first reported as “rice fever” in China by Soong Ying-shin in 1637, and later, it was reported in Japan by Imochi-byo in 1704 [36]. In India, it was first reported in Tamil Nadu in 1913 [37], which became more prevalent and devastating after the introduction of semidwarf and high-yielding varieties during the green revolution [38,39]. The first shattering epidemic was reported in 1919 in the Tanjore delta of south India [40].
Outbreaks of rice blast are the major persistent problem in more than 85 rice-growing countries of the World, especially in the South Asian and African countries [18], with annual yield losses ranging from 10 to 80% [1,11] depending upon the various factors such as varietal susceptibility, the degree of infection, the timing of fungicide application, high humidity, drought, heavy dew, high mean temperatures, high plant density and excessive nitrogen fertilizer [41]. Various studies stated that a total of USD 203.49 million is lost annually in terms of yield loss and management cost of rice blast disease, which alone could feed 60 million people around the World [1,15,42,43]. It has been estimated that the cost of management of blast diseases through chemical fungicides has exceeded USD 70/ha/year [43]. Interestingly, about 157 million tonnes of rice were lost globally between 1975 and 1990 by the severity of rice blast disease, which was more than 30% of global rice production. In India, when the disease accomplishes an epidemic form, yield losses due to rice blasts could be as high as 50%. During natural epidemics of blasts in the wet season, disease incidence ranged from 14 to 27% (above the economic threshold), resulting in yield loss of about 27–35 percent [23,42]. Severe epidemics of the blast occurred between 1980 and 1987 in Himachal Pradesh, Andhra Pradesh, Tamil Nadu, and Haryana, resulting in huge financial losses. According to an estimate, the extent of annual yield reduction caused by rice blast disease is adequate to feed around 60 million people each year [44]. In the United States of America, Arkansas, Louisiana, and Mississippi are the most affected states by the rice blast, where 6% to 50% yield loss has been recorded, and on average, a total of USD 69.34 million is lost annually due to blasts [43]. Moreover, rice blast is also a major issue in European countries such as Italy, Spain, Portugal, Greece, and France, where a 20% to 50% reduction in the milling yield has been observed [1,15,42,43].
3. Pathogenesis of the Causal Fungus Magnoporthe oryzae
The blast disease of rice is caused by a filamentous fungal pathogen, Magnaporthe oryzae (previously Magnaporthe grisae). It is a hemibiotrophic fungus. In the early stage of pathogenesis, Magnaporthe behaves like a biotrophic pathogen and suppresses the plant immune system, and later it switches to a necrotrophic lifestyle that promotes necrosis of host cells [45]. Pathogenesis is a chain of events in a sequential manner that leads to the development of diseases such as attachment, pre-penetration activities, penetration, host recognition, infection, invasion, colonization, reproduction of pathogen, symptom development, dissemination, and survival of the pathogen [46]. Various steps involved in the process of pathogenesis are briefly described in subsequent paragraphs and also presented in Figure 1.
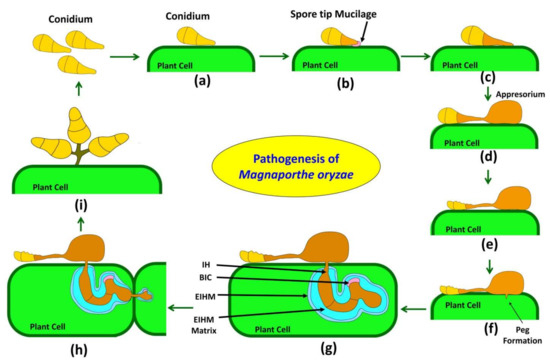
Figure 1.
Pathogenesis of the rice blast fungus Magnaporthe oryzae. (a). Infection of the rice blast fungus starts when a three-celled conidium lands on the rice leaf surface. (b). Spore tip mucilage (STM) present on conidium helps the spore to attach to hydrophobic cuticle of rice plant. (c). Conidium germinate using food reserve and produce a narrow germ tube. (d). Germ tube elongates and give rise to appressorium. (e). Autophagy occurs in three-celled conidium and it dies in a programmed process. (f). In appressorium turgor pressure increases with the help of melanin layer on cell wall and synthesized glycerol inside. Then penetration peg forms at the base, punctured the cuticle of rice and allows entry into the epidermis of plant. (g). Plant tissue invasion occurs by means of bulbous, invasive hyphae (IH) that invaginate the rice plasma membrane and invade epidermal cells. Penetration peg develops into two primary hyphae and separated from rice cytoplasm by extra-invasive hyphal membrane (EIHM). Primary hyphae develop into invasive hyphae. At the tip of primary IH, a new structure known as biotrophic interfacial complex (BIC) develops which is present within EIHM. (h). Invasive hyphae moves from one cell to another by plasmodesmata. (i). Disease lesions occur on plant and sporulation starts under humid conditions, Spores develops on conidiophores with sympodially manners.
3.1. Attachment and Germination
Magnaporthe oryzae pathogenesis starts from a three-celled conidium (Figure 1a), which attaches itself tightly to a hydrophobic, waxy leaf cuticle of rice by an adhesive called spore tip mucilage (STM). STM exists in the periplasmic regions of the conidial tip cell before attachment (Figure 1b) [47]. Upon attachment, three celled conidia germinate and form unbranched germination tubes (also called germ tubes) from the apical cell and grow across the surface of the cuticle. Out of three cells of the spore, only basal and/or apical cells give rise to the germ tube; the middle cell seldom germinates and may function as a reservoir of energy for fungal growth on the host surface [48,49].
3.2. Germ Tube Elongation and Recognition of Host
Various chemical and physical cues, such as primary alcohols, cutin monomers, plant surface waxes, and hard and hydrophobic surfaces, trigger appressorium formation [50]. When the germ tube notices physical cues, such as hydrophobic and hard surface, the tip of the germ tube develops a dome-shaped appressorium which helps in pressure generation to penetrate the host surface. For surface recognition of host, pathogen implies several signal transduction pathways such as PMK1 mitogen-activated protein kinase (MAPK), cyclic adenosine monophosphate dependent protein kinase A (cAMP/PKA), and HOG1 signaling pathways, which also help in appressorium formation, infection peg formation, osmoregulation and cell wall integrity (Figure 1c) [51].
The pth11 gene encodes transmembrane protein Pth11, which is a G-protein coupled receptor (GPCRs). GPCRs are transmembrane receptors confined to the cell membrane of fungal spores and are involved in signal transduction from the outside environment to inside the cell, which helps pathogens to synchronize cell metabolism, cell transport, and growth [52]. Mutational disruption of pth11 triggered a failure in the maturation of appressorium, although the growth of germ tube hooks was not affected in the Δpth11 mutant and confirmed the role of Pth11 in pathogenesis [53]. GPCRs protein is composed of three G alpha (MagA, MagB, and MagC), one G beta (Mgb1), and one G gamma (Mgg1) subunits. Several mutational studies confirmed the role of subunits in pathogenesis [54]. Host cues and signals bind to the GPCR and activate downstream signaling cascade via G-protein, cAMP-PKA, and MAPK signaling pathways that influence metabolism, cellular growth, and morphogenesis of appressorium. A total of eight G protein signaling (RGS) proteins were identified (MoRgs1 through MoRgs8) in Magnoporthe oryzae. Surface hydrophobicity, conidiation, and mating are positively regulated by MoRgs1 and MoRgs4. For germ tube growth and appressorium formation, MoRgs1, MoRgs2, MoRgs3, MoRgs4, MoRgs6, and MoRgs7 are crucial. Although all RGS proteins are involved in the regulation of intracellular cAMP levels, only MoRgs1, MoRgs3, MoRgs4, and MoRgs7 are indispensable for complete virulence [55].
3.3. Appressorium Formation and Maturation
Appressorium formation is regulated by a cascade of the gene in Magnoporthe oryzae. When a three-celled conidia land on the host surface, it attaches itself to the hydrophobic surface of the cuticle with the help of mucilage [56]. On germination, it forms a narrow germ tube in which a nucleus migrates and undergoes mitosis 4–6 h after germination. Various receptors present on the cell membrane of the germ tube recognize the host and initiate dome-shaped appressorium formation after the termination of the tip growth of the germ tube. Following two mitotic divisions, a daughter nucleus moves to develop appressorium, and the rest of the three nucleus returns to conidium and is degraded together with other spore contents, leaving a single nucleus in appressorium (Figure 1d) [57]. Turgor pressure generation through glycerol and other polyols synthesis inside appressorium and the formation of melanin layer between the cell membrane and the cell wall is known as appressorium maturation (Figure 1e) [45,58]. In Magnoporthe oryzae, cargo-independent autophagy and cell death were reported in three-celled conidium just before appressorium maturation to help turgor pressure generation in appressoria [59]. A total of sixteen genes were identified in non-selective macroautophagy; the mutation in any one gene leads to loss of pathogenicity in Magnoporthe oryzae [60]. Yin et al. [61] revealed the role of the autophagy-related gene (MoAtg1) of Magnaporthe, which encodes for kinase protein and possibly phosphorylates MoMkk1 to respond to endoplasmic reticulum stress during plant infection. Increased glycerol inside appressorium causes an influx of more water and builds tremendous pressure on the cell wall up to 8 MPa, and the melanin layer on the cell wall act as a barrier to the efflux of solute and helps to create structural rigidity to maintain increasing pressure [45]. Glycoprotein-rich mucilage adhesives glue melanized appressoria to host surfaces. Mucilage secreted around the base of the appressorium from the appressorial pore helps build up pressure on the host cuticle. Appressorium maturation and penetration are regulated by the MAPK signaling pathway mediated by an Mst11-Mst7-Pmk1 cascade [62]. The MAPK signaling cascade Mck1-Mkk1-Mps1 was reported to be involved in appressorium penetration, maintenance of cell wall integrity, and invasive growth of Magnoporthe oryzae [63].
3.4. Penetration Peg Formation and Invasion
Turgor pressure generated by glycerol and melanized wall of appressorium is focused onto an unmelanized, thin-walled appressorium base where penetration peg starts to develop and generate tremendous pressure on host cuticle and rupture the rice cell cuticle (Figure 1f) [64]. A small amount of cell wall degrading enzymes (CWDEs) such as cutinase, Poly-galactorunase (PGs) is secreted by growing pathogen cells to weaken hard host surfaces [65]. The CUT2 gene of Magnoporthe oryzae produces a cutinase enzyme to degrade cutin present in the cuticle of rice [66]. Two homologs of PGs were identified in the genome of Magnoporthe oryzae, namely endo-PG (MGG_08938.6) and exo-PG (MGG_08752.6). Then, Magnoporthe oryzae obtains entry forcefully to the host cell by translating turgor force into physical force [67]. Ultra-structural analysis reveals that the appressorium pore of Magnoporthe oryzae is different from the rest of the appressorium, with the absence of melanin and a much thinner cell wall [68,69]. A turgor-sensing protein, Sln1, triggers downstream pathways when turgor pressure reaches a threshold, and the septin ring develops around the pore and acts as a barrier of lateral diffusion and control appressorium repolarization [70].
After penetration peg formation, rapid membrane biogenesis and F-actin polymerization occur at the penetration site [68,71]. Recent studies revealed the role of reactive oxygen species (ROS) burst in the re-modeling of cytoskeletal and rapid polymerization of F-actin. ROS burst is catalyzed by two genes, NOX1 and NOX2, that code for NADPH oxidase [71]. NOX1 gene plays a role in the maintenance of the polarized growth and organization of the toroidal F actin network and NOX2required for septin ring formation at the base of the appressorium during penetration peg formation [71,72]. Later appressorium nucleus migrates into the penetration peg, where it undergoes further rounds of mitosis; later, the penetration peg differentiates into primary infective hyphae, then globular invasive hyphae (IH) (Figure 1g). Fungal cells inside the host are confined by the plant-derived extra-invasive hyphal membrane (EIHM) to protect the host defense mechanism. This creates an enclosed apoplastic space between the pathogens IH and the cytoplasm of rice [72]. Further, the EIHM matrix contains several proteins, such as BAS4, which help plants generate defense mechanisms against the pathogen [72,73]. First invaded host cell filled by growth of IH before pathogen spreads into the neighboring host cell. IH switches back to primary IH and moves into uninfected neighboring cells through the plasmodesmata (Figure 1h) [72]. Later it again converted to IH in the newly infected adjacent cell. At the tip of primary IH, a new structure known as biotrophic interfacial complex (BIC) develops, which is present within EIHM. As the fungus multiplies within the first infected cell, BIC remains behind the bulbous IH and again reappears at the tip of the primary IH that will move into neighboring cells (Figure 1i) [73,74]. The mode of action of the focal BIC is unknown, but several effectors accumulate in the BIC to suppress the host immune responses and the virulence of Magnoporthe oryzae [72,74].
3.5. Invasion and Defense Suppression
Magnoporthe oryzae secretes a variety of effector proteins into the host cell to evade the immune response, manipulates host metabolism, and avoids recognition to take advantage during pathogenesis [74]. To date, many effector proteins identified in Magnoporthe oryzae interfere or interact with different target sites of rice. Most effector proteins are produced during the biotrophic phase [73]. Based on the secretion of the effector, there are two kinds of effectors in Magnoporthe oryzae. Apoplastic effectors are secreted into space between IH and EIHM, while Cytoplasmic effectors are secreted into BIC and then translocated into the host cytoplasm with the help of the exocyst complex and t-SNAREs. However, apoplastic effectors are secreted by the conserved ER (endoplasmic reticulum) to the Golgi secretory pathway [75]. Proved by treatment with Brefeldin A that interferes with Golgi-dependent secretion inhibited the secretion of apoplastic effectors such as Bas4 and Slp1 but did not affect the localization of cytoplasmic effectors Pwl2, Bas1, and Bas107 to the BIC [73].
Based on the recognition of effectors by a host protein, effectors are divided into two categories; the first category is Avr effectors, encoded by the avirulence (AVR) genes, which could be recognized by the corresponding resistance (R) gene of rice [76]. Another category is Non-AVR Effectors, which could not be recognized by the host R gene [76]. Effector proteins could be detected in up to four adjacent cells before the hyphal invasion to prepare the host cell for invasion [73]. There is a vast diversity in effectors’ structure; hence, the function of a few effectors is known to date. The biochemical function of Avr-Piz-t is known, which could suppress rice pathogen-associated molecular pattern (PAMP)-triggered immunity by obstructing the ubiquitin ligase activity of the rice RING E3 ubiquitin ligase APIP6 [77]. Well-characterized non-Avr effector Slp1 is a secreted LysM protein that accumulates in between IH and EIHM [73]. However, Slp1 is not required for appressorium penetration but is indispensable for hyphal growth in planta. Slp1 bind with the host component chitin elicitor binding protein (CEBiP) and defeat chitin-induced immune responses, as well as prevent the generation of ROS and defense-related gene expression in rice [73]. Over the past decades, developments in the functional identification of secreted effector proteins from Magnoporthe oryzae have remarkably enhanced our understanding of the molecular mechanisms involved in rice Magnoporthe oryzae interactions. More than 43 secreted proteins have been functionally identified in Magnoporthe oryzae, including 10 Avr effector proteins, PWL1, PWL2 [78], AvrPi-ta [79], AvrPiz-t [80], Avr-Pia, Avr-Pii, Avr-Pik/km/kp [81], Avr-CO39 [82], AvrPi9 [83], and AvrPib [84,85], four biotrophy-associated secreted proteins, BAS1 to BAS4 [86], five pathogenicity related secreted proteins, MPG1 [87], EMP1 [88], MHP1 [89], Slp1 [90] and MC69 [91]; 12 suppressors of plant cell death, IUG6, IUG9, NUP1, NUP2 and NUP3 [92], MoHEG13 [93], and SPD2, SPD4, SPD7, SPD8, SPD9 and SPD10 [94] and 20 plant cell death-inducing effectors proteins, MoHrip1 [95], MoCDIP1 to MoCDIP5 [96], MoHrip2 [96], MSP1 [97], MoNLP1, MoNLP2 and [98], MoSM1 [99] and MoCDIP6 to MoCDIP13 [100]. Recent efforts to understand fungal effector function have revealed that 50% of the Magnoporthe oryzae avirulence effectors and other fungal effectors belong to a new family of structurally conserved MAX effectors (Magnaporthe Avrs and ToxB) [101]. Interestingly, the vast majority of the Magnoporthe oryzae MAX effectors are expressed during the biotrophic stage of infection [101].
4. Pathogenomics of Magnaporthe oryzae
Magnaporthe oryzae is a hemibiotrophic fungus that is responsible for developing blast disease in rice. The genus Magnaporthe was created and accommodated in order Diaporthales [102]. It belongs to the phylum ascomycetes, which develop sexual spores known as ascospores for infecting host plants. Magnoporthe oryzae is known to have hundreds of pathotypes (races) that infect paddy. It is capable of causing damage in almost all stages of paddy, starting from the nursery stage up to grain formation [103]. In order to gain a better understanding of the genomics of this fungus, genome sequencing will play a major role in the future in developing varieties that will be resistant to multiple strains over different geographical regions [104]. Moreover, with the help of various advanced techniques such as genome editing and biotechnological innovations, any of the steps in the biological system of a pathogen can be disrupted or terminated to achieve resistance against blast disease [105]. Host-specific strains can be silenced using targeted mutations or RNAi techniques to create sustainable and strong resistance against the attack of Magnoporthe oryzae.
Interestingly, the genome of Magnoporthe oryzae has been sequenced firstly amongst various phytopathogenic fungi and is being utilized commonly as a model system to understand the mechanism of pathogenicity of pathogen and host–pathogen interactions. Magnoporthe oryzae showed huge genome instability due to the availability of recurrent repetitive sequences in its genome [42,106]. This genome instability plays an important role in the genome variation and the fast evolution of a new race of pathogens within the population. Until now, genome sequencing of more than 74 races of Magnoporthe oryzae has been completed. Among them, each strain contained isolated specific genes and genomic regions, which determine their racial evolution, environmental adaptation, chromosomal variability, variation in repeat element distribution, and host range specificity [96,107]. Consequently, Magnoporthe oryzae has a genome size of 40.12 Mb and contains 12,684 genes in the genome [42]. Further, one of the first strains to be completely sequenced was 70-15 using a whole-genome sequencing shotgun approach. It was discovered that this pathotype contained three MPAK pathways, which were associated with virulence [106]. Two strains of Magnoporthe oryzae, P131 and Y34, were sequenced using Sanger (2-fold) and 454 sequencing technologies [104]. Both of these races had approximately 13% unique DNA when compared with the previously sequenced laboratory strain 70-15. The interesting thing to note was that the deletion of a few genes would cause a decrease in the virulence capacity of the pathogen. For example, deletion of P131_scaffold00208-2 from P131 and Y34_scaffold00875-3 from Y34 would lead to a reduction in virulence and conidiation, respectively [104]. Paired-end libraries of FJ81278 and HN19311 strains of Magnoporthe oryzae were generated through Illumina sequencing. This helped in identifying many virulent genes which were different from 70-15, and genome variation was found at both the basic nucleotide level and chromosome level [96]. The whole genome assembly of another pathotype (RMg-Dl) was completed using PacBio Single-molecule and IlluminaHiSeq 2500 techniques. Strain RMg-Dl was isolated from the Swarna variety cultivated in the Bihar region of India [105].
5. Genetics of Blast Disease Resistance in Rice
Understanding the genetics of the defense system generated by rice plants against the Magnoporthe oryzae is essential to designing a breeding program for developing the disease-resistant variety. A schematic representation of the ice defense system or immunity against blast pathogens is presented in Figure 2. Plants developed two layers of defense; in the first layer, pathogen-associated molecular patterns (PAMPs) are recognized by pattern recognition receptors (PRRs) present on an extracellular membrane or on a trans-membrane [108]. If PRRs of a plant can recognize PAMPs molecules of the pathogen, it induces a relatively weak basic immune response, known as PAMP triggered immunity (PTI), that obstructs the establishment of invading pathogen [109]. Further, the pathogen secretes effector protein to avoid or defeat triggered defense response, which is known as effector-triggered susceptibility (ETS) [110]. The second layer of plant defense is governed by resistance (R) proteins that recognize avirulence (Avr) effectors of the pathogen by direct or indirect binding and induce a wide array of defense responses; this response is known as effector-triggered immunity (ETI). A specific R protein binds to a specific AVR; hence, it is race-specific immunity [111].
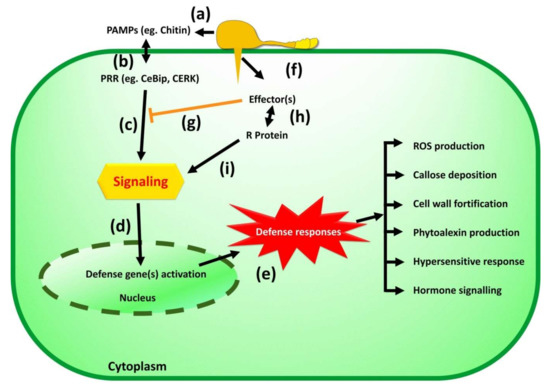
Figure 2.
Overview of rice defense system against blast pathogen Magnoporthe oryzae. (a). PAMPs molecules present on membrane of Magnaporthe spore. (b). PRR presents on the rice cell membrane help to recognize PAMP molecules. (c). A successful recognition by PRR triggers PTI (PAMP-triggered immunity) and activates resistance signaling cascade. (d). Resistance signaling activates defense gene in nucleus of rice. (e). Defense responses includes ROS production, Callose deposition, Cell wall fortification, Phytoalexin production, Hypersensitive response, Hormone signaling etc. (f). In order to avoid recognition of PAMP molecules by PRR, Magnaporthe secretes effectors molecules. (g). Effectors molecules inhibit PTI responses which known as effector triggered susceptibility (ETS). (h). Plant resistant gene recognize effectors of pathogen which known as effector triggered immunity (ETI). (i). Successful recognition of effectors molecules by R gene activates resistance signaling cascade.
PAMP molecule chitin present on the cell wall of Magnoporthe oryzae could be recognized by rice CEBiP (a chitin elicitor binding protein), a lysin motif (LysM) containing plasma membrane proteins, LYP4 and LYP6 [112]. In order to overcome this PTI response in rice, Magnoporthe oryzae secretes an effector protein, Secreted LysM Protein1 (Slp1), during the pathogenesis of new rice cells. Effector Slp1 accumulates in space between the rice plasma membrane and fungal cell wall and competes with CEBiP for binding of chitin molecules, and defeats plant defense gene expression and generation of reactive oxygen species [90].
R-proteins are multidomain proteins generally containing a nucleotide-binding (NB) and a leucine-rich repeat (LRR) domain. To date, 146 R genes, >500 QTLs against rice blast, have been identified, and 36 have been molecularly cloned and characterized [24,113,114,115]. Based on their structure, R genes can be categorized into different classes, i.e., CC-NBS-LRR, LRR, NBS NBS-LRR, Proline-containing protein, and Receptor kinase [116]. Very few pathogen AVR and rice R protein interactions are known, such as AVR1-CO39 to Pi-CO39, AVR-Pita to Pi-ta, ACE1 to Pi33, AVR-Pia to Pia, AVR-Pii to Pii, AVR-Pik/km/kp to Pik/Pik-m/Pik-p, AvrPiz-t to Piz-t, AVR-Pi9 to Pi9, AVRPib to Pib, and AVR-Pi54 to Pi54 [117].
The first studied interaction between AVR and R proteins in the Magnoporthe oryzae–rice pathosystem was AVR-Pita and Pi-ta. AVR-Pita is the first identified avirulence gene in Magnoporthe oryzae, encoding a predicted secreted protein that interacts with Pi-ta and triggers resistance [118]. Pi-ta codes a constitutively expressed 928 amino acidcytoplasmic NLR receptor which is an NBS-LRR class of R protein [119]. Effector AVR-Pita binds to the leucine-rich domain (LRD) of Pi-ta protein directly. A recent study reveals that in the absence of Pi-ta in rice, Avr-Pita targets the rice mitochondria and interacts with the OsCOX11 (Oryzae sativa cytochrome c-oxidase) assembly protein. OsCOX11 participates in mitochondrial reactive oxygen species (ROS) metabolism in rice. Avr-Pita enhances COX activity and decreases ROS accumulation in the host cell, and suppresses host innate immunity by perturbing ROS metabolism in the mitochondria [120].
Pi54 (earlier Pi-kh) is a dominant R gene that encodes ~43 kD protein and has a unique Zinc finger domain that overlaps with the leucine-rich repeat regions and belongs to the NBS-LRR family of R protein [121]. Unlike Pita, Pi54 is induced only in response to pathogen attack [121,122]. In Magnaporthe oryzae AVR-Pi54 gene encodes a predicted secreted protein with a signal peptide (SP) at the N-terminal region. The molecular docking study revealed that AVR-Pi54 protein physically interacts with Pi54 protein through novel non-LRR domains such as STI1 and RhoGEF. The STI1 and GEF domains that interact with AVR-Pi54 are also components of the rice defense complex [28,123]. Microarray analysis in transgenic rice performed at 72 h post-inoculation of the Magnoporthe oryzae revealed that many defense-related genes, such as PAL, laccase, callose, peroxidase, and enzymatic activities of defense response enzymes viz., phenylalanine ammonia-lyase, polyphenol oxidase, b-1,3-glucanase, peroxidase, chitinase, and b-glucosidase, were significantly Up-regulated [124].
Pik is a major R gene located on the long arm of chromosome 11 of rice and requires two NRL receptors, Pik-1 and Pik-2, to trigger cell death upon binding to the AVR-PikD [125]. Effector AVR-PikD interacts with specific rice HMA domain-containing heavy metal-associated isoprenylated plant proteins (OsHIPP19) and heavy metal-associated plant proteins (HPPs) [125]. Both Pik-1 and Pik-2 belong to the coiled-coil nucleotide-binding site leucine-rich repeat (CC-NBS-LRR) class of R proteins [126]. AVR-Pik encodes a secreted protein with a signal peptide at the N-terminus. Pikh-2 initiates host defense response and also physically interacts with the CC domain of Pikh-1 directly. AVRPik-D and Pikh-2 both bind the CC domain of Pikh-1 and form a complex AVR-Pik-Pikh-1-Pikh-2, then a specific signal is transferred from AVR-Pik to Pikh-2 and mediates resistance responses in Rice [83,127].
Pia gene encodes R protein present on chromosome 11 of O. sativa. Pia is composed of two adjacent NLR protein genes, RGA4 and RGA5, and is required for Pia and AVR-Pia interaction [82,128,129]. AVR-Pia encodes a predicted secreted protein with an SP at the N-terminus [81,130]. RGA5 transcripts generate two isoforms by alternative splicing, RGA5-A and RGA5-B. Only RGA5-A is required for Pia-mediated resistance. RGA4 acts as a constitutively active cell death inducer and is inhibited by RGA5 in rice plants without pathogen infection. However, RGA5 is an Avr receptor and has no role in cell death induction. RGA5 and RGA4 form hetero-complexes, and when AVR-Pia or AVR1-CO39 physically binds to the C terminal, non- LRR domain of RGA5, the interaction releases RGA4 and induces hypersensitive cell death in O. sativa [82,129].
AVR-Pii encodes a secreted protein belonging to the pex33 protein family, having four homologs [81]. AVR-Pii first accumulates in the BIC and is then translocated into the host cytoplasm [131]. Avr-Pii form complex with OsExo70-F2 and OsExo70-F3. OsExo70-F2 and OsExo70-F3 are presumably involved in exocytosis, and these proteins stably form homo- and hetero-dimers that incorporate AVR-Pii. R gene Pii encodes a 1025-amino acid protein predicted to be an NLR protein [132]. R protein Pii and AVR-Pii interact indirectly in the host cell [133]. In the absence of R protein Pii, AVR-Pii directly binds to Os-NADP-ME2 (Os nicotinamide adenine dinucleotide phosphate-malic enzyme), inhibits their activity, and thereby leads to the reduction of the PAMP-triggered ROS burst and successful biotrophy [131]. However, in the presence of Pii, OsExo70 acts as a decoy or helper in Pii/AVR-Pii interactions and activates defense response against Magnoporthe oryzae [133].
AvrPiz-t codes for a predicted secreted protein [80] and is first secreted into the BIC before translocation into the rice cell [77]. Effector AvrPiz-t targets 12 APIPs (AvrPiz-t interacting proteins) of rice. Among the twelve APIPs, APIP6 and APIP10 are functional ring E3 ubiquitin ligases, APIP5 is a bZip transcription factor, and APIP12 is a nucleoporin2 domain (Pfam 04096) containing protein [77,134,135]. Piz-t and AvrPiz-t interact indirectly [77,134,135]. Effector AvrPiz-t participates in both PTI and ETI of rice to Magnoporthe oryzae and targets the rice RING E3 ubiquitin ligases APIP6 (AvrPiz-t Interacting Protein 6) and APIP10 by promoting their degradation to suppress PTI in rice [77,136]. APIP10 promotes the degradation of Piz-t via ubiquitination. AvrPiz-t can remove negative regulation of Piz-t by degradation of APIP10 through the AvrPiz-t/APIP10 protein–protein interaction [136]. At the necrotrophic stage, a functional bZip transcription factor, APIP5 interacts with AvrPiz-t directly. APIP5 form homo-dimers and then interact with AvrPiz-t through its bZip DNA-binding domain at the N-terminus. The interaction suppresses APIP5 transcriptional activity and protein accumulation, leading to cell death. Thus, AvrPiz-t promotes effector-triggered necrosis (ETN) in the absence of Piz-t. When Piz-t is present in the rice cell, the N-terminus of Piz-t also interacts with the N-terminus of APIP5 and stabilizes APIP5 accumulation and activity to prevent rice cell necrosis. At the same time, APIP5 promotes the accumulation of Piz-t to maintain its basal level for providing resistance [134].
6. Disease Screening Protocols for Blast Resistance in Rice
The first step in a resistance breeding program is to rapidly screen all the available genetic stocks, including the local land races, improved cultivars, and exotic germplasms, using empirical techniques in glass houses or by field tests. Efficient, accurate, and reliable methods for screening disease resistance/susceptibility in crop plants are very important in developing resistant crop varieties in a relatively short period of time in a sustainable manner [137]. Based on the accuracy of the screening methods, resistant or susceptible genotypes might be identified for further breeding programs. Standard screening protocols of rice varieties for susceptibility to rice blast are usually carried out by spraying the plant with conidial suspensions under greenhouse and field conditions using local isolates [138]. Details of individual techniques are briefly described hereunder.
6.1. Field Screening Technique
A high-throughput and reliable field screening protocol for rice blast resistance are essential for the identification of resistant germplasm/varieties and resistant genes for further breeding programs [139]. In field conditions, artificial leaf blast disease screening usually takes place in a Uniform Blast Nursery (UBN) [138,140,141], which has a 10 m length and 1 m width (Figure 3). Test entries (30 plants/test entry) are planted in a nursery bed at a spacing of 10 cm plant to plant and 50 cm row to row. Moreover, a mixture of susceptible check entries is planted after every 10 lines of test entry as a spreader line and also planted throughout the border of UBN as a border line to facilitate the even spread of the blast disease. One or more susceptible entries could be used as border/spreader lines. The soil in the UBN is enriched with farm yard manures (FYM) and recommended doses of fertilizers. However, applying an excess rate of nitrogen fertilizer (150 kg N/ha) makes rice more vulnerable to spreading blast infection [138]. Simultaneously, isolation, maintenance, and multiplication of local and highly virulent blast cultures (fungal conidia) should be performed according to the method suggested by Vasudevan et al. [141], Prasad et al. [142], and Chhallagulla et al. [143]. Moreover, artificial inoculation is performed with a local and highly virulent blast race (fungal conidial suspension at a concentration of 1 × 105 spores/mL) by spraying on UBN beds at 25–30 days after sowing (DAS). Later, the nursery beds are water sprayed 3–4 times per day and are covered with polythene sheets during the night to maintain a high humidity until disease development and progression are observed in border lines and spreader lines [138,141,143]. The observations on disease resistance or susceptibility are taken from each entry 10–15 days after the artificial inoculation and taken 2–3 times at 5 days intervals using the Standard Evaluation System 2002 (SES) of International Rice Research Institute, Manila, Philippines [144]. In this system, the disease score has ranged from 0–9 based on the severity of blast infestation in the leaves of the plant. The lines with disease scores of 6–9 are considered susceptible lines, 4–5 as moderately resistant, and 0–3 as highly resistant [144].
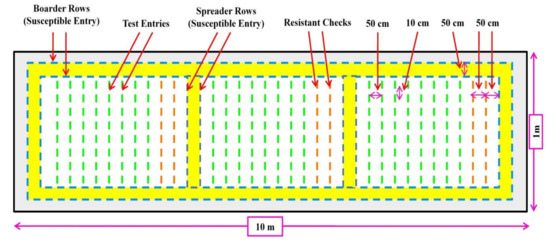
Figure 3.
Schematic representation of Uniform Blast Nursery for screening of rice genotypes for blast disease.
Several studies have been reported for using standard blast screening techniques and identified a number of novel resistance sources and genes. Vasudevan et al. [141] conducted a large-scale screening of 4246 geographically diverse rice accessions originating from 13 major rice-growing countries to identify a new resistance source for blast resistance. These accessions were selected from over 120,000 accessions based on their annotated rice blast resistance information in the international rice gene bank. The rice lines were screened using a two-step screening protocol which includes natural infection in a rice uniform blast nursery followed by artificial infections with five single rice blast isolates. Systematic screening for rice blast resistance was performed both under field and controlled environmental conditions, and rice cultivars IR72 and CO39 were used as susceptible control lines. Among the lines screened, 289 accessions showed broad-spectrum resistance (BSR) against all five single rice blast isolates. The accessions showing BSR were genotyped for the presence of the Pi2 resistance gene for the identification of promising accessions for the isolation of allelic variants of the resistance gene. Blast monogenic resistant lines for Pi54 (IRBLkh-K3), Piz-t (IRBLzt-T), Pi9 (IRBL9-W), Pita (IRBLta-CP1), and Pi2 (IRBLz5-CA), Pib (IRBLb-B) were used as control lines to isolate specific rice blast resistance genes.
Similarly, Qin et al. [139] developed a high-throughput and reliable blast resistance evaluation system at the field level for the breeding of resistant varieties. This method consists of the following steps (i) pretreatment of diseased straw; (ii) sowing of seeds (iii) initiating seedling blast of the first batch of spreader population (iv) Sowing of seeds (v) and inducing seedling blast of the second batch of spreader population and test materials (vi) induction of seedling blast. Based on this protocol, a total of 730 indica hybrid rice were screened. This procedure enables uniform and consistent infection, which facilitates efficient and accurate assessment of seedling blast resistance for diverse rice materials.
6.2. Screening Techniques under Greenhouse/Polyhouse/Controlled Conditions
Under a greenhouse/polyhouse screening procedure, the test entries 15 plants/test entry) are sown in plastic trays (10 rows × 2 columns per tray) in 4–5 batches for inoculation with different individual blast isolates (The number of batches can be increased/decreased based on the availability of individual isolates of the pathogen for screening). Plants are allowed to grow in normal conditions for 10–15 days. Rice blast pathogen cultured on culture medium may be taken for preparation of conidial suspension at 1 × 105 conidia/mL of water. All the plants are then inoculated with 50 mL of spore suspension solution with a concentration of 1 × 105 spores/mL with 1% tween-20 per tray after 10–15 DAS [141,145].
After inoculation, the plants need to be kept in a moist chamber at 26–28 °C for 24 h to maintain temperature and humidity. Plants should be transferred to the incubation chamber at 25 °C ± 2 for 1 week, and water is sprayed three to four times during day time to maintain high humidity (humidity should be near 100% for the initial 72 h to favor disease initiation [141,145]. Then the disease reaction should be assessed after nine days of inoculation and scored on a zero to nine rating scale as per the Standard Evaluation System 2002 (SES) of the International Rice Research Institute. Manila, Philippines [144].
6.3. Molecular-Marker-Based Screening of Rice Genotypes for Blast Resistance
In addition to the phenotypic screening for the identification of blast resistance source, molecular markers linked to the blast resistance genes/genes-specific primers are being utilized to identify the presence of resistance genes in the rice germplasm/line [146]. Tightly linked molecular markers, viz., simple sequence repeats (SSRs), single nucleotide polymorphisms (SNPs), and CAPSs (Cleaved Amplified Polymorphic Sequences), have been widely used for screening rice genotypes for blast-resistant genes [22,147,148,149,150,151,152,153]. Molecular-marker-based techniques not only allow the identification of resistant lines in a non-destructive manner but also help in the identification of broad-spectrum resistance genes and alleles of different genes and a combination of resistance genes in the germplasm [149]. This method gives fast and accurate results within a short period of time. With the advent of molecular markers paved the path for changing the paradigm of rice breeding by identification of genomic regions controlling several economic traits and their deployment in the elite rice lines using MAS and MABB approaches.
Singh et al. [149] screened rice accessions with allele-specific SSR markers to identify 10 major blast resistance genes (Piz-5, Pi-9, Pitp(t), Pi-1, Pi-5(t), Pi-33, Pi-b, Pi27(t), Pi-kh, and Pi-ta) in 192 rice germplasm accessions from different ecological regions. They found genetic frequencies of the 10 major rice blast resistance genes varied from 19.79% to 54.69%. Interestingly, they found 17 rice accessions that harbored seven to eight major blast resistance genes indicating their exploitation in further breeding programs for developing blast-resistant cultivars. Furthermore, Imam et al. [150], Shikari et al. [151], Yan et al. [152], and Teerasan et al. [153] also used the linked SSR markers for screening of blast resistance in rice genotypes. Kim et al. [147] employed eight SNP markers (tightly linked with six major genes, Piz, Piz-t, Pik, Pik-m, Pik-p, and Pit) to determine the genetic diversities of blast resistance (R) genes from 86 accessions of aromatic rice and found four accessions of indica type carrying the six major genes. Moreover, Kim et al. [148] used SSR and CAPS markers for blast resistance screening in rice genotypes.
7. Strategies to Develop Durable Resistance in Rice Genotypes against the Blast Disease
Many agronomical management practices, biological control, chemical control, disease forecasting, etc., have been adopted by the farmers to escape the hazards and reduce the losses caused by blast disease, but none of them has proven to be 100% efficient. Furthermore, the application of excessive fungicides to control the disease may cause severe loss of biodiversity, a threat to ecology, and produce hazardous food material [1,9,42]. Therefore, the development of broad-spectrum and durable resistance in rice varieties for blast disease is the only hope to combat the infestation of Magnoporthe oryzae. Thus, rice breeders should focus on developing durable, resistant rice varieties as resistance genes provide a worthwhile and environmentally safe option for the management of blast disease [19]. Many approaches, viz., traditional breeding methods, molecular-marker-based breeding approaches, transgenic breeding, genome editing-based methods, etc., are being deployed as major weapons to develop durable resistant varieties and are ecologically as well as economically sustainable. Advancement in genomic approaches and bioinformatic tools has led to the foundation for developing blast-resistant rice varieties with more accuracy and precision in a limited time period. A brief account of various breeding and biotechnological tools which can be used for the development of resistant rice varieties are briefly described in subsequent paragraphs and also presented in Figure 4.
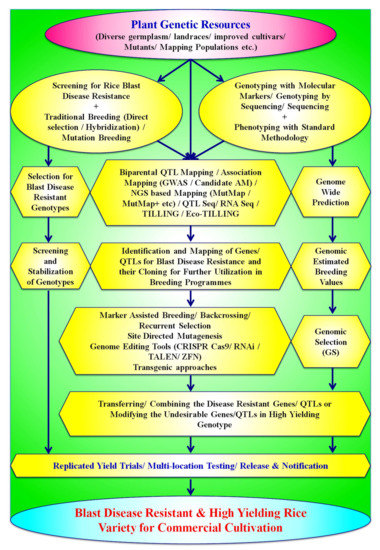
Figure 4.
Schematic representation of various breeding biotechnological approaches used for development of blast-resistant rice varieties.
7.1. Conventional Breeding Strategies for Developing Resistance against Rice Blast
The traditional breeding approaches are robust and oldest methods for developing novel genetic variants for blast resistance. Breeders have developed many blast-resistant rice varieties through traditional breeding methods, viz., pedigree method, backcross breeding, introduction and acclimatization, multiline breeding, pureline selection, recurrent selection, and mutation breeding. Interestingly, conventional breeding has enabled the International Rice Research Institute, Manila, Philippines, to generate elite cultivars with an enormous range of disease-resistant genes [12]. This approach is appropriate for developing durable and sustainable resistant rice varieties; however, its long and time-consuming breeding cycle and laborious nature are the major drawbacks of this method [19].
Introduction and acclimatization are the important, easiest, fastest, and most economically efficient breeding methods used to develop resistant varieties by introducing promising entries into new areas or regions where they have not been cultivated before. Most of the disease-resistant lines developed by IRRI, Manila, Philippines, have been disseminated to various rice-growing countries through the introduction and acclimatization process and used as breeding material for developing new disease-resistant varieties either through pureline selection or through the hybridization method [20]. Rice varieties IR36 and IR64 are the best examples of introduction and utilization in the development of new varieties. IR36 and IR64 contain the Pita gene, and IR64 also has another closely linked gene, Pi20, conferring resistance to blast disease. In addition to these known Pi genes, these varieties have accumulated several defense genes with their complex lineages [154,155], which make them durably resistant to blast disease in most locations. These varieties are used extensively in breeding programs in the southern states of India, where the blast is recognized as a potential threat to increased productivity [156].
Moreover, scientists from Indonesian Center for Rice Research (ICRR), Indonesia, have used more than 30 blast-resistant varieties of traditional and introduced materials for hybridization-based breeding programs. They have conducted the selection for blast resistance in a greenhouse through artificial inoculation of 19 pathogen races available in the ICRR’s collection. Interestingly, a total six promising blast-resistant lines, viz., TB490C-TB-1-2-1, TB361B-30-6-2, BP1976B-2-3-7-TB-1-1, TB356BTB-18-3, IR30176, and IR60080-23, with different patterns of resistance to the pathogen races were selected through participatory varietal selection and tested under farmers field [157].
Furthermore, multiline varieties or a mixture of several resistant near-isogenic lines can be used to reduce the outbreaks of blast disease. These lines carry resistance to different races of the same pathogen, which helps in developing durable resistance against the pathogen. Several scientists confirmed the use of multiline varieties for the control of the severity of blast disease [158,159,160,161,162]. The multiline variety ‘‘Sasanishiki’’ has been developed at the Furukawa Agricultural Experiment Research Station, Japan, in 1995 and commercially cultivated in 5,800 hectares of farmers’ fields in Miyagi Prefecture of northern Japan in 1997 as a blast-resistant rice variety. It consisted of seven different lines from BL1 to BL7, which carried seven different resistant genes against blast. These lines were developed by continuous backcrossing of Sasanishiki (recurrent parent) with blast-resistant land races of cultivars (Donor parent) [163]. Moreover, Zhu et al. [164] suggested using a cultivar mixture consisting of 80–90% resistant plants and 10–20% susceptible plants of similar varietal background to reduce the rapid evolution and emergence of new virulent Magnoporthe oryzae [160,161].
Simultaneously, the concept of shuttle breeding was also deployed during the 1970s and 1980s for developing blast-resistant rice varieties. Over 3000 rice germplasms were evaluated in 31 countries at 126 test sites from 1975 to 1992 under the International Network for Genetic Evaluation of Rice (INGER) [165,166]. A total of 522 resistant entries of the International Rice Blast Nursery (IRBN) were utilized in the hybridization program across 18 countries from 1984 to 1992 [166].
Furthermore, a few promising donors such as Ram Tulsi, Oryza nivara, Dawn, Tetep, Carreom, Zenith, Gam pai 15, Pankhari 203, and a number of improved plant types resulted from this study and were used on a regular basis as parents in blast-resistant breeding programs by various countries [165,166].
The pedigree method is the most efficient and extensively used breeding method for handling segregating generations from crosses, and a large number of varieties have been developed in crops such as rice, wheat, barley, sorghum, pulses, oilseeds, etc. It is the most appropriate method for improving the disease resistance ability of existing cultivars within a short span of time if the resistance is governed by major genes [20]. It is possible to combine genes for resistance to six or seven major diseases and insects in a short period through the pedigree method [12,39]. Most of the IRRI bred lines viz., IR24, IR34, IR36, IR60, IR56, IR64, IR46, and IR74 having blast-resistant genes have been used extensively in a breeding program for developing the blast-resistant rice varieties through the pedigree method [20,154]. Moreover, the TN-1 rice variety has also been used in breeding for blast resistance as a susceptible parent. Martínez et al. [167] have developed many blast-resistant genotypes from the segregating lines of Fanny (highly susceptible to blast) and 11 cultivars differing in blast resistance through the pedigree method. However, maintenance of accurate pedigree records and long breeding cycles are the major limiting factors for deploying this method. Moreover, this method will not be fruitful when blast resistance is governed by polygenes [12,19]. Backcross breeding is another widely used and common technique in rice breeding for transferring genes and chromosomes from one variety to another and from related species [20,168]. It has been extensively used for transferring disease resistance to popular and widely adapted varieties. For instance, a backcross was made between IR68835-98-2-B-2-1-1 (a broad-spectrum blast resistance variety) and KDML105 (a susceptible variety), and 83 lines from BC3F2 generation were evaluated for resistance against 12 different strains of the blast. All the BILs (Backcross Introgression Lines) displayed a low level of disease score. These lines can be further utilized in breeding programs for developing blast-resistant varieties [169]. Moreover, backcross breeding was integrated with molecular marker-based techniques to make the breeding process more clear, accurate, and authentic and also to reduce the exhaustive breeding exercise of maintaining a huge population.
In addition to the above-mentioned methods, recurrent selection has also been deployed in rice breeding programs for the development of blast-resistant varieties [170]. Recurrent selection is characterized by being a cyclical method in which gains for the trait or traits under selection occur gradually and continuously. Interestingly, this method requires shorter breeding cycles and provides better genetic gains with wider genetic diversity in breeding lines for blast resistance. The durable blast-resistant rice variety CG-91 was developed through recurrent selection [171].
Conventional breeding methods have played a significant role in sustaining food production for burgeoning populations. However, conventional breeding takes more time and effort, which can be reduced nowadays due to advancements in genomic technologies. Conventional breeding and phenotyping are inefficient when several lineages of the pathogen are present, and resistances to multiple lineages are warranted [156]. Moreover, the success in resistance breeding is generally affected by linkage drag, due to which undesired traits closely linked with resistance genes are also transferred in new selections. Therefore, molecular-marker-based breeding approaches are now preferable in resistance breeding towards increasing its efficiency.
7.2. Mutation Breeding for Blast Resistance in Rice
A mutation is a rare event, reversible and recessive in nature, and a primary source of all genetic variations existing in any organism, including plants. Mutation breeding in rice is used to complement conventional breeding since this technique is very effective for improving one or a few traits, such as agronomic traits and resistance to pests and diseases, without altering the unique properties of improved variety/germplasm to make them easy acceptable among the farmers [172]. By the year 2022, about 3402 mutant varieties in more than 225 crops have been developed through induced mutagenesis by different countries and registered in the FAO/IAEA Mutant Variety Database (MVD), International Atomic Energy Agency (IAEA), Vienna, Austria [173] which has made significant contributions for food and nutritional security.
Furthermore, many attempts have been made to develop disease resistance in rice against blast through mutation breeding. According to FAO/IAEA MVD [173], a total of 151 rice mutants having blast resistance have been developed across the World and registered in FAO/IAEA MVD. Interestingly, 72 rice mutants were developed through directly induced mutagenesis (Table 1), and 79 varieties were developed by crossing with mutant variety (Supplementary Table S1). Among the 72 rice mutants, four varieties were developed by chemical mutagenesis, whereas 68 were developed by the use of physical mutagens [173]. China, Japan, and India are the top three countries that have developed 56, 54, and 15 rice mutants, respectively [173]. The mutant variety Xiongyue 613 was the first officially approved blast-resistant rice variety, which was developed through mutation breeding (200 Gy of gamma rays) in 1965. The main improved attributes of Xiongyue 613 are moderate resistance to blast, higher yield, and good quality FAO/IAEA MVD, 2022). Interestingly, with the advancement in technologies, China has developed five rice mutants viz., Zhe 101, Hangtian 36, Huahang-simiao, Liangyouhang 2, and Neiyouhang 148 through the use of cosmic rays (treatment of seeds in aerospace). In India, a total of 15 rice mutant varieties that have blast resistance have been developed and registered in FAO/IAEA MVD, among which some mutants were developed through radiation-induced (X-rays, gamma rays) mutation breeding, while others were the results of hybridization with mutant variety. In India, ‘Jagannath’ is the first rice mutant produced in 1969 by X-rays irradiation of the popular tall variety T141, which was found to be resistant to blast [173]. Recently, a high-yielding and blast-resistant variety Vikram-TCR has been developed through radiation-induced mutation breeding under the joint collaboration of Indira Gandhi Krishi Vishwavidyalaya, Raipur, India, and Bhabha Atomic Research Centre, Mumbai, India. This variety has been developed with the help of gamma radiation (300 Gy) from the Safri-17 landrace, which is susceptible to blast disease [174,175]. When looking at the outcomes of the mutation breeding, it can be said that it is playing a significant role in combating the impact of blast disease in rice.

Table 1.
List of blast-resistant rice mutant varieties developed across the World.
7.3. Molecular-Marker-Based Approaches for Resistance to Rice Blast Disease
Conventional breeding methods are slow and time-consuming, with uncertain results due to environmental impacts. With the advent in molecular techniques, a combination of traditional methods which are aided by new molecular techniques such as gene pyramiding, marker-assisted selection (MAS), marker-assisted back cross (MABC), QTL mapping, genome-wide association studies (GWAS), etc., can help accelerate the process of selection, screening, and development of blast-resistant rice varieties. Consequently, the identification and mapping of these R genes/alleles through advanced genomic approaches will be helpful in modern plant breeding for developing durable, resistant varieties [22,26,176]. Mining and characterization of disease-resistant genes/QTLs and their further deployment for developing resistant cultivars are the most preferred strategies by plant breeders. About 100 years ago, Sasaki [177] had, for the first time, reported the resistant varieties for rice blast fungus, Magnoporthe oryzae, in Japan, and Kiyosawa [178] identified the first rice blast gene Pi-a from a japonica rice variety “Aichi Asahi”. Interestingly, until now, about 146 R genes for rice blast resistance have been identified and mapped from both indica and japonica subspecies of rice by various scientists (Table 2). Among the 146 identified R genes, scientists have molecularly characterized and cloned 36 genes so far (Table 3) [15,24,42].
Furthermore, out of the 146 mapped R-genes, about 72% (105) are located on chromosomes 2, 6, 11, and 12 containing 13, 26, 38, and 28 genes, respectively. In addition, scientists have identified and mapped more than 500 QTLs for blast resistance in rice through linkage-based QTL mapping and Genome Wide Association Mapping (GWAS), from which 23 blast resistance loci viz., PiGD-1(t), Pi25(t), Pi26(t), Pi27(t), Pitq1, Pitq5, Pitq6, Pizh, Pi24(t), Pi25(t), Pi28(t), Pi29(t), Pi30(t), Pi31(t), Pi32(t), PiGD-3(t), Pi35(t), PiGD-2(t), Pilm2, Pi7(t), Pi34, and Pi21 have been isolated [15,24,176,179]. Among the 36 cloned R genes, the Pik locus is very important as it harbors a number of blast R genes (Pik, Pikm, Pikp, Piks, Pikh, and Pi1) which are being utilized regularly in rice breeding for developing durable resistance against the blast disease [176,180,181]. Several studies reported that R genes from the Pi2/9 locus of chromosome 6 had been extensively used in the breeding for blast resistance [182,183,184]. Similarly, the cultivar IR 64 contains 11 R-genes for blast resistance which are regularly utilized as donor parents in breeding for blast-resistant varieties. Interestingly, some of the major R-genes such as Pikh, Pi-1, Pi9, Pi20, Pi27, Pi39, Pi40, and Pit have broad-spectrum resistance (BSR) against the pathogen Magnoporthe oryzae [24,185].
Table 2.
List of 146 blast-resistant genes identified and mapped in rice.
Table 2.
List of 146 blast-resistant genes identified and mapped in rice.
| S. No. | Name of Blast-Resistant Genes | Name of Source Genotype | Reporting Year | Chromosome Number | Genomic Position (Mb) | Name of Linked Molecular Marker | Name of Country | References |
|---|---|---|---|---|---|---|---|---|
| 1 * | Pb2 | Jiangnanwan | 2022 | 11 | 1.47 | SNP | China | Yu et al. [15] |
| 2 | Pi67 | Tetep | 2019 | 12 | 12.09 | SSR | India | Joshi et al. [186] |
| 3 | Pi57(t) | IL-E1454 | 2017 | 12 | 10.8 | SSR, STS | Myanmar | Dong et al. [187] |
| 4 | Pi65(t) | Gangyu 129 | 2016 | 11 | 28.22 | SNP, InDel | – | Zheng et al. [114] |
| 5 | Pi-jnw1 | Jiangnanwan | 2016 | 11 | 27.36 | SSR, InDel | – | Wang et al. [188] |
| 6 | Pi66(t) | AS20-1 | 2016 | 3 | 26.78 | SSR | Australia | Liang et al. [189] |
| 7 | Pita3(t) | IRBLta2-Re | 2015 | 12 | 9.89 | SSR | – | Chen et al. [190] |
| 8 * | Pik-e | Xiangzao 143 | 2015 | 11 | 28 | SSR, InDel | China | Chen et al. [190] |
| 9 | Pi-h2(t) | HR4 | 2015 | 1 | 7.9 | SSR | India | Xiao et al. [191] |
| 10 | Pih3(t) | HR4 | 2015 | 12 | 12.95 | SSR | India | Xiao et al. [191] |
| 11 | Pi-h1(t) | HR4 | 2015 | 11 | 28.11 | SSR, InDel | India | Xiao et al. [191] |
| 12 * | Pi64 | Yangmaogu | 2015 | 1 | 32.31 | SSR, InDel | Japan | Ma et al. [192] |
| 13 | Pitb | Zixuan | 2013 | 12 | 9.37 | SSR, InDel | – | Sun et al. [193] |
| 14 | Pi61(t) | 93-11 | 2013 | 12 | 9.98 | InDel, SSR | China | Lei et al. [194] |
| 15 | Pi60(t) | 93-11 | 2013 | 11 | 6.62 | SSR, InDel | China | Lei et al. [194] |
| 16 | Pi58(t) | Haoru | 2013 | 12 | 10.42 | SSR | Myanmar | Koide et al. [195] |
| 17 | Pi51(t) | D69 | 2012 | 6 | 10.38 | InDel, SSR | – | Xiao et al. [196] |
| 18 * | Pi50(t) | EBZ, EBZ × LTH F2 and (EBZ × LTH) × LTH, BC1F2 | 2012 | 6 | 10.41 | SSR, CAPS | – | Zhu et al. [197] and Jiang et al. [198] |
| 19 | Pi-hk1 | Heikezijing | 2012 | 11 | 27.66 | SSR | – | Wu et al. [199] and Liu et al. [200] |
| 20 | Pihk2 | Heikezijing | 2012 | 9 | 10.17 | SSR, InDel | – | He et al. [201] |
| 21 | Pias(t) | Asominori | 2012 | 4 | 31.26 | SSR, CAPS | China | Endo et al. [202] |
| 22 | Pi51(t) | Tianjingyeshengdao | 2012 | 12 | 11.95 | SSR, SFP | China | Wang et al. [203] |
| 23 | pi55(t) | Yuejingsimiao 2 | 2012 | 8 | 25.58 | SSR, STS | China | He et al. [201] |
| 24 | Pi46(t) | H4 | 2011 | 11 | 27.74 | SSR, InDel | – | Xiao et al. [204] |
| 25 | Pi-48 | Xiangzi 3150 | 2011 | 12 | 11.95 | SSR | China | Huang et al. [205] |
| 26 * | Pi-a | Aichi Asahi | 2011 | 11 | 6.49 | SSR, InDel | Japan | Zeng et al. [206] |
| 27 | Pi-45(t) | Moroberekan | 2011 | 4 | 31.49 | SSR | Japan | Kim et al. [148] |
| 28 | Pi-42(t) | DHR9 | 2010 | 12 | 10.62 | RAPD, SSR, STS | India | Kumar et al. [207] |
| 29 | Pi43(t) | Zhe733 | 2009 | 11 | 27.67 | SSR | – | Lee et al. [208] |
| 30 | Pi-41 | 93-11 | 2009 | 12 | 16.74 | SSR, STS | China | Yang et al. [209] |
| 31 * | Pid3 | Digu | 2009 | 6 | 13.05 | STS | China | Shang et al. [210] |
| 32 * | Pik-p | K60, HR22 | 2009 | 11 | 28.05 | SSR, CAPS | China | Wang et al. [211] |
| 33 | Pi2-2 | Jefferson | 2008 | 6 | 10.2 | SSR | – | Jiang et al. [198] and Ballini et al. [179] |
| 34 * | Pikahei- 1(t) | Kahei | 2008 | 4 | 31.67 | SSR, SNP | – | Xu et al. [212] |
| 35 * | Pik-h | IRBLkh-K3, HP2216, and Tetep | 2008 | 11 | 24.99 | SNP | India | Xu et al. [213] |
| 36 | Pir2-3(t) | IR64 | 2008 | 2 | – | SSR | Indonesia | Dwinita et al. [214] |
| 37 | Pirf2-1(t) | O. rufipogon | 2008 | 2 | – | SSR | Indonesia | Dwinita et al. [214] |
| 38 | Pi39(t) | Chubu 111, Q15 | 2007 | 4, 12 | – | SSR | China | Liu et al. [215] |
| 39 | Pi-39(t) | Mineasahi and Chubu 111 | 2007 | 4 | 32.68 | SSR | China | Liu et al. [215] |
| 40 | Pi-39 | Q-15 and Chubu 111 | 2007 | 12 | 10.61 | SSR | China | Liu et al. [215] |
| 41 | Pi-34 | Chubu-32 | 2007 | 11 | 19.96 | SSR | Japan | Zenbayashi et al. [216] |
| 42 | Pi-40(t) | IR65482, Co39, and O. australiensis (W) | 2007 | 6 | 9.86 | STS, SSR | Philippines | Jeung et al. [217] |
| 43 | Piz-5 | C101A51_CO39 | 2006 | 6 | – | – | – | Deng et al. [218] |
| 44 * | Pi9 | Cultivar TP309 | 2006 | 6 | 10.39 | – | – | Qu et al. [219] and Koide et al. [195] |
| 45 * | Pid2 | Digu | 2006 | 6 | 17.16 | CAPS | China | Chen et al. [220] |
| 46 * | Pigm(t) | Gumei 4 | 2006 | 6 | 10.36 | CAPS, InDel | China | Deng et al. [218] |
| 47 | Pi51(t) | Tianjingyeshengdao | 2006 | 12 | – | – | China | Qu et al. [219] |
| 48 | Pi2-1 | Tianjingyeshengdao | 2006 | 6 | 10.08 | SSR, SFP | China | Wang et al. [203] and Qu et al. [219] |
| 49 | Pi24(t) | Azuenca | 2006 | 1 | 5.24 | SSR | France | Nguyen et al. [221] |
| 50 | Pi-38 | Tadukan | 2006 | 11 | 22.48 | SSR, AFLP | India | Gowda et al. [222] |
| 51 * | Pi35(t) | Hokkai 188 | 2006 | 1 | 32.1 | SSR | Japan | Nguyen et al. [221] |
| 52 * | Pi-b | Tohoku, Koshihikari | 2006 | 2 | 35.1 | SNP | Japan | Hayashi et al. [223] |
| 53 * | Piz-t | Toride No. 1 | 2006 | 6 | 10.39 | STS | Japan | Zhou et al. [224] |
| 54 | Pi59(t) | Haoru_US-2 | 2006 | 6 | 10.82 | SSR | Myanmar | Koide et al. [195] and Zhou et al. [224] |
| 55 | Pi-9(t) | IR31917 | 2006 | 6 | 10.38 | STS | Philippines | Qu et al. [219] |
| 56 | Pi-Da(t) | Dacca 6 | 2005 | 2 | 2.21 | SSR | – | Lei et al. [225] |
| 57 * | Pi 37(t) | Cultivar St. No. 1 | 2005 | 1 | 33.1 | SSR | China | Chen et al. [226] |
| 58 | Pi26(t) | Gumei 2 | 2005 | 6 | 11.06 | RFLP, SSR | China | Wu et al. [227] |
| 59 * | Pi-36(t) | Q61 | 2005 | 8 | 2.87 | SSR, CRG | China | Liu et al. [228] |
| 60 * | Pi54 | Tetep | 2005 | 11 | 25.26 | SSR | India | Sharma et al. [121] |
| 61 | PiGD-2(t) | Sanhuangzhan 2 | 2004 | 10 | – | SSR, RFLP, RGA | – | Liu et al. [229] |
| 62 | Pi-d1(t) | Digu | 2004 | 2 | 34.94 | SSR, RFLP | China | Chen et al. [230] |
| 63 | Pi-dt(2) | Digu | 2004 | 6 | 17.16 | SSR, RGA | China | Chen et al. [230] |
| 64 | Pig(t) | Guangchangzhan | 2004 | 2 | 34.34 | SSR | China | Zhou et al. [231] |
| 65 | Pi15 | Q61 and GA25 | 2004 | 9 | 9.61 | SSR, CRG | China | Liu et al. [229] |
| 66 | PiGD-1(t) | Sanhuangzhan 2 | 2004 | 8 | 16.37 | SSR, RFLP, RGA | China | Liu et al. [229] and He et al. [201] |
| 67 | PiGD-3(t) | Sanhuangzhan 2 | 2004 | 12 | 14.45 | SSR, RFLP, RGA | China | Liu et al. [229] |
| 68 | Pi-y2(t) | Yanxian No. 1 | 2004 | 2 | 35.03 | SSR | China | Fukuta [232] and Lei et al. [225] |
| 69 | Pi-y1(t) | Yanxian No. 1 | 2004 | 2 | 35.03 | SSR | China | Fukuta [232] and Lei et al. [225] |
| 70 | Pi27(t) | Q14 and Q61 | 2004 | 1 | 5.55 | SSR | France | Zhu et al. [233] |
| 71 * | Pi-tp(t) | CO39 and Tetep | 2004 | 1 | 25.13 | SSR | India | Barman et al. [234] |
| 72 | Pi-sh | Akihikari and Shin 2 | 2004 | 1 | 33.3 | SSR | Japan | Fukuta [232] |
| 73 * | Pik-s | Shin 2 | 2004 | 11 | 27.31 | SSR | Japan | Fjellstrom et al. [235] |
| 74 | Pi28(t) | Azucena, IR64 | 2003 | 10 | 21.04 | RFLP, RAPD | – | Sallaud et al. [236] |
| 75 * | Pi56(t) | SHZ-2 | 2003 | 9 | 9.77 | SSR, CRG, SNP | – | Jeon et al. [237] |
| 76 | Pizh | Zhai-Ya-Quing8 | 2003 | 8 | 4.38 | – | China | Sallaud et al. [236] |
| 77 | Pi-25(t) | IR64 | 2003 | 2 | 34.36 | QTL | France | Sallaud et al. [236] and Nguyen et al. [221] |
| 78 | Pi27(t) | IR64 | 2003 | 6 | 6.92 | RFLP | France | Sallaud et al. [236] |
| 79 | Pi26(t) | IR64 | 2003 | 5 | 2.78 | RFLP, RAPD | France | Sallaud et al. [236] |
| 80 | Pi-32(t) | IR64 | 2003 | 12 | 21.24 | RFLP, RAPD | France | Sallaud et al. [236] |
| 81 | Pi-31(t) | IR64 | 2003 | 12 | 11.93 | RFLP, RAPD, | France | Sallaud et al. [236] |
| 82 | Pi-29(t) | IR64 | 2003 | 8 | 13.93 | RFLP, RAPD, Isozyme | France | Sallaud et al. [236] and Nguyen et al. [221] |
| 83 | Pi-30(t) | IR64 | 2003 | 11 | 4.41 | RFLP, RAPD, Isozyme | France | Sallaud et al. [236] and Nguyen et al. [221] |
| 84 | Pi-33 | IR64, Bala | 2003 | 8 | 7.56 | SSR, RFLP | France | Berruyer et al. [238] and Sallaud et al. [236] |
| 85 | Pii2 | Ishikari Shiroke | 2003 | 9 | 1.03 | – | Japan | Pan et al. [239], Kinoshita and Kiyosawa [240] |
| 86 * | Pi-5(t) | RIL249, Moroberekan | 2003 | 9 | 9.77 | AFLP, RFLP, CAPS | Philippines | Jeon et al. [237] |
| 87 * | Pi2 | 5173, C101A51 | 2002 | 6 | 10.39 | SSR, STS, RFLP | – | Jiang and Wang [241] and Zhou et al. [224] |
| 88 | Pi-24(t) | Zhong 156 | 2002 | 12 | 10.6 | RFLP, RAPD, RGA | – | Zhuang et al. [242] |
| 89 * | Pi-CO39(t) | Co39 | 2002 | 11 | 6.66 | SSR, RFLP | USA | Chauhan et al. [243] and Huang et al. [205] |
| 90 * | Pi25 | Gumei 2 | 2001 | 6 | 18.09 | – | China | Zhuang et al. [242] |
| 91 | Pi-25(t) | Gumei 2 | 2001 | 6 | 12.33 | RFLP, RGA, SSR | China | Wu et al. [199] and Zhuang et al. [242] |
| 92 | PBR | St. No. 1 | 2001 | 11 | – | RFLP, SSR | Japan | Fukuoka and Okuno [244] |
| 93 | Pi-47 | Xiangzi 3150 | 2000 | 11 | 27.67 | SSR | China | Huang et al. [205] and Ahn et al. [245] |
| 94 | Pi18 | Suweon365 | 2000 | 11 | 28.93 | RFLP | Korea | Ahn et al. [245] |
| 95 | Pi-lm2 | Lemont, Teqing | 2000 | 11 | 28.93 | RFLP | USA | Tabien et al. [246] |
| 96 | Pi-tq5 | Teqing | 2000 | 2 | 34.61 | RFLP | USA | Tabien et al. [246] Tabien et al. [247] and Zhou et al. [231] |
| 97 | Pi-tq1 | Teqing | 2000 | 6 | 29.02 | RFLP | USA | Tabien et al. [246] |
| 98 | Pi-tq6 | Teqing | 2000 | 12 | 7.73 | RFLP | USA | Tabien et al. [246] |
| 99 | Pi49 | Mowanggu | 1999 | 11 | 28.8 | SSR | – | Sun et al. [193] and Chen et al. [248] |
| 100 | Pi-16(t) | AUS373 | 1999 | 2 | 34.94 | RFLP, Isozyme | Japan | Pan et al. [249] and Zhou et al. [231] |
| 101 * | Pb1 | Modan | 1999 | 11 | 21.71 | – | Japan | Fujii et al. [250] and Hayashi et al. [251] |
| 102 | Pi-44(t) | Moroberekan | 1999 | 11 | 28.93 | RFLP, STS, AFLP | USA | Chen et al. [248] and Chauhan et al. [243] |
| 103 | Pi12 | Hong Jiao Zhan K80-R-Hang Jiao-Zhan | 1998 | 12 | 7.73 | RFLP | Japan | Zhuang et al. [252] |
| 104 | Pi-19(t) | IRBL19-A and Aichi Asahi | 1998 | 12 | 10.73 | SSR | Japan | Koide et al. [195] and Hayashi et al. [253] |
| 105 | Pi-14(t) | Maowangu | 1998 | 2 | 34.94 | RFLP, Isozyme | Japan | Pan et al. [254] and Zhou et al. [231] |
| 106 * | Pi3(t) | Pai-kan-tao | 1997 | 9 | 7.8 | – | – | Kinoshita and Kiyosawa [240] |
| 107 * | pi-21 | Owarihatamochi | 1997 | 4 | 19.81 | RFLP, SSR | Japan | Fukuoka and Okuno [244], Ahn et al. [255], and Pan et al. [254] |
| 108 | Pita-2 | Yashiromochi, Pi No. 4 | 1997 | 12 | 10.6 | RFLP, RAPD, SNP | Japan | Hayashi et al. [223] |
| 109 | Pi22 | Suweon 365 | 1997 | 6 | 4.89 | RFLP | Korea | Ahn et al. [255], Terashima et al. [256] |
| 110 | Pi23 | Suweon 365 | 1997 | 5 | 10.75 | RFLP, SSR | Korea | Ahn et al. [255], Rybka et al. [257] |
| 111 | Pi-20(t) | IR64 | 1997 | 12 | 12.95 | SSR | Philippines | Li et al. [258] and Imbe et al. [259] |
| 112 * | Pita | Tadukan, Yashiromochi | 1997 | 12 | 10.6 | RFLP, RAPD, SNP | USA | Rybka et al. [257], Hayashi et al. [223] and Bryan et al. [119] |
| 113 * | Pi-k | Kusabue, Kanto 51 | 1996 | 11 | 28.01 | RFLP, InDel, SNP | China | Hayasaka et al. [260] and Hayashi et al. [223] |
| 114 * | Pik-m | Tohoku IL4, Tsuyuake | 1996 | 11 | 28 | RFLP, SSR | China | Kaji and Ogawa [261] |
| 115 | Pi157 | Moroberekan | 1996 | 12 | 12.37 | RFLP | India | Naqvi et al. [262] |
| 116 * | Pii1 | Fujisaka 5 | 1996 | 6 | 2.29 | – | Japan | Pan et al. [263] |
| 117 | Pikg | GA20 | 1996 | 11 | 27.31 | – | Japan | Pan et al. [263] |
| 118 * | Pit | K-59, Tjahaja, K-59 | 1996 | 1 | 2.27 | RFLP, SNP | Japan | Kaji and Ogawa [261] and Hayashi et al. [223] |
| 119 | Pi8 | Kasalath | 1996 | 6 | 11.36 | leucine aminopeptidase, phosphoglucose isomerase, RFLP | Japan | Pan et al. [263] |
| 120 | Pi62(t) | Yashiromochi | 1996 | 12 | 7.73 | RAPD, RFLP | Japan | Wu et al. [264] |
| 121 | Pi62(t) | Yashiromochi | 1996 | 12 | 7.73 | RAPD, RFLP | Japan | Wu et al. [264] |
| 122 | Pi-17(t) | DJ 123 | 1996 | 7 | 22.25 | leucine aminopeptidase, phosphoglucose isomerase | Philippines | Pan et al. [263] and Zhu et al. [197] |
| 123 | Pib2 | Lemont | 1996 | 11 | 26.79 | – | Philippines | Tabien et al. [265] (1996) |
| 124 | Pitq3 | Teqing | 1996 | 3 | – | USA | Tabien et al. [265] | |
| 125 | Pitq2 | Teqing | 1996 | 2 | – | USA | USA | Tabien et al. [265] |
| 126 | Pitq4 | Teqing | 1996 | 4 | USA | USA | Tabien et al. [265] | |
| 127 | Pik-l | Liziangxintuanheigu, Kusabue | 1995 | 11 | 27.69 | SSR, STS, CAPS | China | Hayasaka et al. [260] and Hayashi et al. [223] |
| 128 | Pi-10(t) | Tongil | 1995 | 5 | 14.52 | RAPD | India | Naqvi et al. [265] and Wu et al. [227] |
| 129 * | Pi-1(t) | Apura, C101LAC | 1995 | 11 | 28 | STS, RFLP, SSR, CAPS | USA | Parco [266], Yu et al. [267] |
| 130 | Pi(t) | P167 | 1994 | 4 | 2.27 | – | – | Causse et al. [268] |
| 131 | Pi-11(t) | Zhai-Ye-Quing | 1994 | 8 | 13.93 | RFLP, RAPD | China | Causse et al. [268] |
| 132 | Pi-6(t) | Apura | 1994 | 12 | 7.73 | RFLP | USA | McCouch et al. [269] |
| 133 | Pi-7(t) | RIL29 (Moroberekan) | 1994 | 11 | 18.64 | 12.37 | USA | Wang et al. [270] |
| 134 | Pi26(t) | Azucena/Gumei 2 | 1993 | 5 | 2.07 | – | France | Wu and Tanksley [271] Ahn et al. [255] and Nguyen et al. [221] |
| 135 | Pi-13 | O. minuta (W), Kasalath | 1992 | 6 | 15.83 | SSR | Philippines | Amante- Bordeos et al. [272] |
| 136 | Pi3(t) | Pai-kan-tao | 1992 | 6 | – | – | Philippines | Mackill and Bonman [273] |
| 137 | Pi1 | LAC23 | 1991 | 11 | 26.49 | RFLP | Philippines | Yu et al. [274] |
| 138 | Pikur2 | Kuroka | 1988 | 11 | 2.84 | – | Japan | Goto [275] |
| 139 * | Pish | Nipponbare | 1985 | 11 | 33.38 | – | Japan | Imbe and Matsumoto [276] |
| 140 | Mpiz | Zenith | 1976 | 11 | 4.07 | – | Japan | Goto [277] |
| 141 | Piz | Zenith, Fukunishiki, Toride 1, Tadukan | 1976 | 6 | 10.39 | STS | Japan | Goto [277] and Zhou et al. [224] |
| 142 | Pif | Chugoku 31-1 | 1971 | 11 | 24.69 | – | Japan | Shinoda et al. [278] |
| 143 | Pii | Ishikari Shiroke | 1971 | 9 | 2.29 | – | Japan | Ise [279] and Shinoda et al. [278] |
| 144 | Piis1 | Imochi Shirazu | 1970 | 11 | 2.84 | – | Japan | Goto [280] |
| 145 | Pikur 1 | Kuroka | 1970 | 4 | 24.61 | Isozyme | Japan | Fukuoka et al. [281] and Goto [280] |
| 146 | Pise | Sensho | 1970 | 11 | 5.74 | – | Japan | Goto [280] |
‘-‘ Indicates the non-availability of data, * Indicates that the particular gene has been cloned and molecularly characterized.
Table 3.
List of blast resistance genes cloned, cloning strategy and their location on rice chromosome.
Table 3.
List of blast resistance genes cloned, cloning strategy and their location on rice chromosome.
| S. No. | Name of Blast Resistance Genes | Proteins Encoded by R Genes | Donor Rice Lines/Genotypes | Chromosome No. | Year of Cloning | Cloning Approach Used for Isolation of R Genes | Reference Serial No. of Table 2 | Reference |
|---|---|---|---|---|---|---|---|---|
| 1 | Pb2 | NBS-LRR protein with NB-ARC domain and LRR domain | Jiangnanwan | 11 | 2022 | Map-based cloning | 1 | Yu et al. [15] |
| 2 | Pid3-I1 | CC-NBS-LRR | MC276 | 6 | 2019 | Gene Mapping | 31 | Inukai et al. [282] |
| 3 | Pitr | A typical protein with an armadillo repeat (Putative E3 ligase) | Katy | 12 | 2018 | Map-based cloning | 71 | Zhao et al. [283] |
| 4 | Pigm | NBS-LRR | Gumei 4 | 6 | 2017 | Map-based cloning | 46 | Deng et al. [35] |
| 5 | Pi64 | CC–NBS–LRR | Yangmaogu | 1 | 2015 | Map-based cloning | 12 | Ma et al. [192] |
| 6 | Pi50 | NBS-LRR | Er-Ba-zhan (EBZ) | 6 | 2015 | - | 18 | Su et al. [113] |
| 7 | Pik-e | CC-NBS-LRR | Xiangzao 143 | 11 | 2015 | Map-based cloning | 8 | Chen et al. [190] |
| 8 | Pi35 | NBS-LRR | Hokkai-188 | 1 | 2014 | Map-based cloning | 51 | Fukuoka et al. [284] |
| 9 | Pi63/Pikahei-1(t) | NBS-LRR | Kahei | 4 | 2014 | Map-based cloning | 34 | Xu et al. [285] |
| 10 | PiK-h | NBS-LRR | K3 | 11 | 2014 | Positional cloning | 35 | Zhai et al. [127] |
| 11 | Pi54of | NBS–LRR | Oryza officinalis (nrcpb004) | 11 | 2014 | Map-based cloning | 60 | Devanna et al. [28] |
| 12 | Pii | NBS-LRR | Hitomebore | 9 | 2013 | MutMap-Gap | 116 | Takagi et al. [132] |
| 13 | Pi-CO39 | CC-NBS-LRR | CO39 | 11 | 2013 | -- | 89 | Cesari et al. [82] |
| 14 | Pi56 | NBS–LRR | Sanhuangzhan No. 2 | 9 | 2012 | Map-based cloning | 75 | Liu et al. [200] |
| 15 | Pi1 | CC–NBS–LRR | C101LAC | 11 | 2012 | Map-based cloning | 129 | Hua et al. [286] |
| 16 | Pi54rh | NBS-LRR | Oryza rhizomatis (nrcpb 002) | 11 | 2012 | Map-based cloning | - | Das et al. [287] |
| 17 | Pi25 | CC-NBS-LRR | Gumei2 | 6 | 2011 | Map-based cloning | 10 | Chen et al. [288] |
| 18 | Pia | CC-NBS-LRR | Aichi Asahi | 11 | 2011 | MB and mutant screening | 26 | Okuyama et al. [128] |
| 19 | Pik-p | CC-NBS-LRR | K60 | 11 | 2011 | Map-based cloning | 32 | Yuan et al. [289] |
| 20 | Pik | CC-NBS-LRR | Kusabue | 11 | 2011 | Map-based cloning | 113 | Zhai et al. [180] |
| 21 | Pish | NBS–LRR | Shin-2 | 1 | 2010 | Mutant Screening | 139 | Takahashi et al. [290] |
| 22 | Pb1 | CC-NBS-LRR | Modan | 11 | 2010 | Map-based cloning | 101 | Hayashi et al. [251] |
| 23 | Pi54/Pi-kh | NBS-LRR | Tetep | 11 | 2010 | Map-based cloning | 73 | Sharma et al. [122] |
| 24 | Pit | CC-NBS-LRR | K59 | 1 | 2009 | Map-based cloning | 118 | Hayashi and Yoshida [291] |
| 25 | pi21 | Proline-rich heavy metal binding protein | Owarihatamochi | 4 | 2009 | Map-based cloning | 107 | Fukuoka et al. [281] |
| 26 | Pi-d3 | CC-NBS-LRR | Digu | 6 | 2009 | In silico analysis | 106 | Shang et al. [210] |
| 27 | Pi5 | CC-NBS-LRR | Moroberekan | 9 | 2009 | Map-based cloning | 86 | Lee et al. [292] |
| 28 | Pik-m | NBS-LRR | Tsuyuake | 11 | 2008 | Map-based cloning | 114 | Ashikawa et al. [293] |
| 29 | Pi37 | NBS-LRR | St. No. 1 | 1 | 2007 | Map-based cloning | 57 | Lin et al. [294] |
| 30 | Pi36 | CC-NBS-LRR | Q61 | 8 | 2007 | Map-based cloning | 59 | Liu et al. [215] |
| 31 | Pi-d2 | B-lectin receptor kinase | Digu | 6 | 2006 | Map-based cloning | 45 | Chen et al. [220] |
| 32 | Pi9 | NBS-LRR | 75-1-127 | 6 | 2006 | Map-based cloning | 44 | Qu et al. [219] |
| 33 | Pi-2 | NBS-LRR | C101A51 | 6 | 2006 | Map-based cloning | 87 | Zhou et al. [224] |
| 34 | Piz-t | NBS-LRR | Toride 1 | 6 | 2006 | Map-based cloning | 53 | Zhou et al. [224] |
| 35 | Pi-ta | NBS-LRR | Yashiro-mochi | 12 | 2000 | Map-based cloning | 112 | Bryan et al. [119] |
| 36 | Pib | NBS-LRR | Tohoku IL9 | 2 | 1999 | Map-based cloning | 52 | Wang et al. [295] |
‘-‘ Indicates the non-availability of data.
Identification and mapping of the blast resistance R-genes through advanced biotechnological and genomic approaches are essential for their efficient utilization in molecular breeding programs, especially in marker-assisted selection and marker-based gene pyramiding of two or more R-genes for achieving the broad-spectrum and durable resistance [15,22,24,296,297]. In addition, this also gives information about the gene-linked, gene-based, and functional markers, which can enhance the efficiency of conventional resistance breeding programs. Further, this provides an opportunity for molecular characterization and cloning of R-genes [298]. Recent mapping studies, viz., linkage-based QTL mapping and linkage-disequilibrium-based association mapping, are the most widely used methods for identification and mapping of the blast resistance R-genes and have successfully illustrated the acceleration of the breeding program. Linkage-based QTL mapping was proposed as a useful molecular breeding technique for detecting the QTLs as it utilizes the biparental mapping population developed from two contrasting genotypes. In the past few years, more than 350 QTLs and blast resistance genes have been identified and localized on rice chromosomes for blast resistance through a linkage-based QTL mapping strategy [236,244,247,299,300,301,302,303,304,305,306,307,308,309,310,311,312].
In association mapping, genome-wide association studies (GWAS) detect the genetic variation (marker) polymorphisms of multiple individuals in the whole genome to obtain the genotype associated with the observable traits. In comparison with the linkage-based QTL mapping, it has the ability to map genes with high resolution, cover rich-captured variations, and have high efficiency in locating multiple traits simultaneously. GWAS has been deployed by scientists for the last two decades for the identification and mapping of blast-resistant genes and revealed more than 230 blast-resistant loci in rice which are distributed throughout the genome [15,22,115,176,313,314,315,316,317,318,319,320,321,322,323,324,325,326]. Among them, five rice blast resistance loci, including the cloned gene Pita, were first identified by GWAS [115,313].
The identified and mapped genes/QTLs could be easily deployed in the breeding program with the help of molecular markers. Hybridization, backcrossing, marker-assisted selections, and marker-assisted backcross breeding (MABB) are the most popular methods for the introgression of resistance (R) genes into the elite cultivar for improving the rice line for disease resistance. Gene pyramiding involves transferring more than one favorable gene/QTL of traits from multiple parents into a single genotype by marker-assisted selection (MAS). A number of studies have been made to deploy R genes in rice breeding programs for blast resistance through the gene introgression or pyramiding approach [27,185,327,328,329,330,331]. A series of improved intermediate materials with various blast resistance gene combinations or improved new varieties were bred to achieve broader and more durable resistance. The detail of rice lines/varieties improved by introgression of single, multiple genes for blast resistance and introgression of genes for multiple biotic stresses and a combination of biotic and abiotic stresses is presented in Table 4.
Most studies focused on pyramiding individual resistance genes to develop single or multiple resistances [185,332,333,334,335,336,337,338], but these lines have less durability of resistance. Hence, a pyramiding of genes for a trait by combining two R-genes [182,327,339,340,341,342,343,344,345] or more than two complementary R-genes [230,343,346,347,348,349,350] could provide superior phenotypic benefits and stability compared to a single gene. Furthermore, looking at the current scenario of changing climatic conditions, breeding rice cultivars with durable multiple biotic stresses resistance through gene pyramiding will be the most effective method, and several scientists have already started to work on this aspect [27,331,349,351,352,353,354,355]. Hence, in this way, molecular markers could be very useful in breeding multiple disease resistance along with resistance in rice.
Table 4.
List of blast resistance genes deployed for improvement of rice lines using marker-assisted selection (MAS) and marker-assisted backcross breeding (MABB).
Table 4.
List of blast resistance genes deployed for improvement of rice lines using marker-assisted selection (MAS) and marker-assisted backcross breeding (MABB).
| S. No. | Gene/QTL | Trait/Resistance | Type of Molecular Marker | Technique/Approach Used | Application/Lines Developed | References |
|---|---|---|---|---|---|---|
| Single gene for blast resistance | ||||||
| 1. | Pi1 | Resistance to blast disease | ISSR and SSR | MABB | Marker-assisted backcross breeding for improvement of variety Zhenshan97 A | Liu et al. [185] |
| 2. | Piz | Resistance to blast disease | SSR | MAS | Tightly linked markers with Pi-z locus was applied for screening of germplasm for blast resistance in rice | Fjellstrom et al. [332] |
| 3. | Pita | Resistance to blast disease | Gene-specific to gene | MAS | Applied for detection of Pita gene in 141 rice germplasms and introduction of gene through advanced breeding approaches | Wang et al. [333] |
| 4. | Pi9 | Resistance to blast disease | Gene-specific | Marker aided selection | Introgressed Pi-9(t) resistance gene in the cultivar Luhui 17 | Wen and Gao [334] |
| 5. | Pi39 | Resistance to blast disease | InDel | MABB | Introgressed into Chinese cultivar Q15 | Hua et al. [335] |
| 6. | Pikh | Resistance to blast disease | SSR | MABB | Improvement of Malaysian Cultivar, MR264 by Introgression of Pikh gene | Hasan et al. [336] |
| 7. | Pi40 | Resistance to blast disease | SSR | MABB | Introgressed into elite cultivars Turkish, Halilbey and Osmancik-97 | Beser et al. [337] |
| 8. | Pi-ar | Resistance to blast disease | RAPD | MAS | Introgression of Pi-ar gene using double haploid technique | Araujo et al. [338] |
| Two genes for Blast resistance | ||||||
| 9. | Piz-5, Pi54 | Resistance to blast disease | SSR | MABB | Blast disease resistance genes transferred to develop Pusa 1602 and Pusa 1603 | Singh et al. [356] |
| 10. | Pi1, Piz | Resistance to blast disease | SSR | MABB | Pyramiding of Pi1 and Piz-5 genes into PRR78 | Gouda et al. [340] |
| 11. | Pi1, Pi2 | Resistance to blast disease | SSR | MABB | Introgressed into Intan variety and BPT5204 | Hegde et al. [344] |
| 12. | Pi46, Pita | Resistance to blast disease | SSR | MABB | Introgression of resistance genes into Hang hui 179 (HH179) | Xiao et al. [343] |
| 13. | Pi2, Pi9 | Resistance to blast disease | SNP | MABB | Introgression of blast resistance genes into R179 | Luo et al. [345] |
| 14. | Pi-b and Pik-h | Resistance to blast disease | SSR, RM 208, RM 206 | MABB | Pyramided two blast resistance genes into MR219Malaysian rice variety | Tanweer et al. [327] |
| 15. | Piz-5 and Pi54 | Resistance to blast disease | SSR | MABB | Incorporation of blast resistance into “PRR78”, an elite Basmati rice restorer line | Singh et al. [297] |
| 16. | Pi-2 and Pi-54 | Resistance to blast disease | SSR | MABB | Introgression of blast resistance genes into the genetic background of elite, bacterial blight resistant indica rice variety, Improved Samba Mahsuri | Madhavi et al. [342] |
| 17. | Pi54 and Pi1 | Resistance to blast disease | SSR | MAS | Introgression of blast resistance genes into cold tolerant variety Tellahamsa | Oddin et al. [341] |
| 18. | Pi46 and Pita | Resistance to blast disease | SSR | MABB | Blast resistance genes were introgressed into an elite restorer line Hang-Hui-179 (HH179) | Xiao et al. [343] |
| More than Two genes for Blast resistance | ||||||
| 19. | Pi1, Piz-5, Pita | Resistance to blast disease | RFLP | MAS | Pyramiding of three NILs namely (C101LAC, C101A51 and C101PKT) for blast resistance into a single cultivar CO39, each linecarrying resistance genes Pi1, Piz-5 and Pita, respectively. | Korinsak et al. [169] |
| 20. | Pi1, Pi2, Pi33 | Resistance to blast disease | SSR | MABB | Introgression of multiple blast disease resistance genes into Jin23B | Chen et al. [302] |
| 21. | Pi1, Pi2, Pi33 | Resistance to blast disease | SSR | MAS | Improvement of Russian rice varieties by pyramiding of blast disease resistance genes | Usatov et al. [357] |
| 22. | Pi9, Pizt, Pi54 | Resistance to blast disease | SNP | MABB | Introgression of Pi9, Pizt, Pi54 blast resistance genes into japonica rice 07GY31 | Xiao et al. [343] |
| 23. | Pi1,Pi2, Pi33 | Resistance to blast disease | SSR | MABB | Improving blast resistance in Indian rice variety ADT43 by pyramiding three blast resistance genes | Divya et al. [346] |
| 24. | Pid1, Pib, Pita, Pi2 | Resistance to blast disease | SSR | MAS | Pid1, Pib and Pita genes were introduced into G46B, while Pi2 was introduced into Zhenshan 97B | Chen et al. [230] |
| 25. | Pizt, Pi2, Pigm, Pi40, Pi9, Piz | Resistance to blast disease | SSR | MAS | Introgression of multiple blast resistance genes into Yangdao 6 | Wu et al. [350] |
| Multiple stress tolerance | ||||||
| 26. | Xa21, Piz | Resistance to blast and bacterial leaf blight disease | SSR | MAS | Introgression of Blast and Bacterial leaf blight disease resistance gene | Narayanan et al. [351] |
| 27. | Pi2 and Xa23 | Resistance to blast and bacterial leaf blight disease | SSR | MAS | Introgression of broad-spectrum disease resistance genes into, elite thermo-sensitive genic male-sterile rice line-GZ63-4S | Jiang et al. [182] |
| 28. | Xa21 and Pi54 | Resistance to blast and bacterial leaf blight disease | SSR | MABB | Introgression of BLB and blast resistance into DRR17B, an elite, fine-grain type maintainer line of rice | Balachiranjeevi et al. [348] |
| 29. | Pi1, Pi2, Xa23 | Resistance to blast and bacterial leaf blight disease | SSR | MABB | Introgression of bacterial blight and blast resistance into variety Rongfeng B | Fu et al. [358] |
| 30. | Pi2, Xa21, Xa33 | Resistance to blast and bacterial leaf blight disease | SSR | MABB | Introgressed bacterial blight and blast diseases resistance genes into RPHR-1005 | Kumar et al. [359] |
| 31. | Pi9Drought | Resistance to blast and drought tolerance | Gene linked markers | MAS | Pi9 has been introgressed into different genetic backgrounds of cultivated varieties, such as indica cultivar Swarna + drought | Dixit et al. [331] |
| 32. | Pi2, Pi54, xa13 and Xa21 | Resistance to blast and bacterial leaf blight disease | SSR | MABB | Introgressed of bacterial blight and blast diseases resistance genes for improving disease resistance traits in Basmati rice varieties | Ellur et al. [352] |
| 33. | Xa21,xa13 and Pi54 | Resistance to blast and bacterial leaf blight disease | Gene-specific | MABB | Pyramiding of bacterial blight and blast diseases resistance into Indian rice variety MTU1010 | ArunaKumari et al. [353] |
| 34. | Xa21, xa13 and Pi54 | Resistance to blast and bacterial leaf blight disease | SSR | MAS | Improvement of Vallabh Basmati 22 by Introgression of Xa21, xa13 genes for Bacterial Blight and Pi54 for Blast disease resistant genes | Srikanth et al. [349] |
| 35. | Xa21, Xa33, Pi2, Rf3 and Rf4 | Resistance to blast and bacterial leaf blight disease | SSR | MAS | Marker-assisted improvement of the elite restorer line of rice, RPHR-1005 for resistance against diseases | Kumar et al. [360] |
| 36. | Xa 5 and 4 blast QTLs | Resistance to blast and bacterial leaf blight disease | SSR | MAS | Introgression of bacterial leaf blight and blast resistance genes into rice cultivar RD6 | Pinta et al. [361] |
| 37. | Xa13, Xa21, Pi54, qSBR11 | Resistance to blast and sheath bight disease | SSR | MAS | Transfer of multiple disease resistance genes for bacterial blight, blast and sheath blight disease in rice | Singh et al. [297] |
| 38. | Pi54, qSBR11-1, qSBR11-2 and qSBR7-1 | Resistance to blast and sheath bight disease | SSR and QTLs | MABB | Introgression of multiple disease resistance genes into a maintainer of Basmati rice CMS line | Singh et al. [347] |
| 39. | Pi2, Pi9, Gm1, Gm4, Sub1, and Saltol | Blast disease, Gall Midge Submergence and Salinity tolereance | SSR and gene linked markers | MAS | Pyramiding of genes/QTLs to confer resistance/tolerance to blast, gall Midge, submergence, and salinity in a released rice variety CRMAS2621-7-1 as Improved Lalat | Das and Rao [362] |
| 40. | Pi9, Xa4, xa5, xa13, Xa21, Bph3, Bph17, Gm4, Gm8 and qDTY1.1 and qDTY3.1 | Blast Bacterial leaf blight Brown planthopperGall midge and QTLs for drought tolerance | Gene based/linked markers | Marker-assisted forward breeding | MAS in combining tolerance to multiple biotic and abiotic stresses in Swarna + drought recurrent parent | Dixit et al. [331] |
| 41. | Pi9, Xa21, Gm8, qDTY1.1, qDTY2.2 and qDTY4.1 | Blast, Bacterial blight (BB), Gall midge (GM) and QTLSs drought tolerance | Gene based/linked markers | Marker-assisted forward(MAFB) and back cross (MABC) breeding | Introgressed in to Indian elite rice variety, Naveen | Janaki Ramayya et al. [354] |
| 42. | BPH3, BPH24, Pi2, Pi9, Pita, Pib, Xa21Pimh, and badh2 | Brown Plant hopperblast disease, bacterial blight and Aroma | SSR and gene linked markers | MAS | Brown planthopper (BPH), blast, and bacterial leaf blight (BLB) resistance and aroma genes into elite rice maintainers and restorers | Wang et al. [27] |
| 43. | Xa21, Pi54, Pup1 | BB resistance gene, the blast resistance gene, and low soil phosphorous tolerance QTL/gene, | Gene/QTL linked markers | Marker-assisted pedigree breeding | BB resistance gene, the blast resistance gene, and low soil phosphorous tolerance QTL/gene in to MTU 1010 (CottondoraSannalu) | LaxmiPrasanna et al. [355] |
7.4. Transgenic Breeding for Rice Blast Resistance
The most significant advancement in the area of varietal development for disease resistance is the use of the techniques of genetic engineering to develop transgenic rice resistant to diseases. This approach is advantageous for introducing disease resistance into elite rice cultivars since transgenic plants can acquire a single desired trait without any alteration of the original genetic background. Several studies have been performed to confer the disease resistance in rice against the Magnoporthe oryzae [28,29,30,31,363,364,365,366,367,368,369,370], which are presented briefly in Table 5.
Table 5.
Deployment of transgenic approaches for developing blast resistance in rice.
Using Agrobacterium-mediated transformation, Nishizawa et al. [363] reintroduced the chitinase gene in Japonica rice varieties Nipponbare and Koshihikari. Two genes, Cht-2, which accumulated chitinase intracellularly, and Cht-3, which generated chitinase extracellularly, were introduced. It was found that even if either of the genes was expressed in the transgenic plants, they showed enhanced resistance to blast fungus. Studies have demonstrated the expression of the Gns1 gene, which is responsible for hydrolyzing glycosidic bonds in cell walls of the Poaceae family. Gns1 gene was introduced in the plants with CaMV35S as its promoter. The over-expression of this gene led to the development of resistance against blast disease, but it also resulted in stunted growth of the plants. The use of this gene for blast resistance is limited as its constant expression results in poor root formation, diminutive growth, and development of certain brown specks [365].
Interestingly, a different approach to rice blast resistance was undertaken by transferring regulatory and structural genes of maize which were responsible for the production of flavonoids. Flavonoids are responsible for generating various levels of stress responses in plants. In this study, anthocyanin production was increased in Tp309, a japonica rice variety, by the transfer of the C2 gene of maize [364]. Transfer of this gene might have resulted in a mutation leading to the generation of CHS protein which provided resistance against blast disease [364]. Furthermore, Coca et al. [366] developed blast-resistant transgenic rice by transferring the ER-CecA gene from the giant silk moth Hyalophora cecropia. This gene was optimized to produce Cecropin A peptides in paddy, which are a member of antimicrobial protein families. This transformation did not account for any pathogenesis-related gene expression, which is a good indicator of the direct effect of a gene on the pathogen [366].
Another promising approach to rice blast fungus resistance was exhibited by transferring an antifreeze glycopeptide gene using Agrobacterium as a medium. This gene was overexpressed in transformed paddy plants, which were able to withstand low temperatures such as −1 °C up to 24 h. Transgenic and control plants were recovered and thawed. Interestingly, transgenic plants displayed fewer symptoms and more resistance against blast fungus as compared to other fungi. This implies that such pathogenesis-related proteins can be identified and used to generate resistance against blast [367]. Similarly, another host defense antimicrobial peptide, thanatin, was transferred into the rice. The transformants displayed a significant level of resistance against blast fungus [368]. Similarly, Chen et al. [369] transferred the Pi-d2 gene with the help of three vectors into nine lines of paddy to generate blast-resistant rice varieties. This gene exhibited broad-spectrum resistance against blast as it displays resistance against 39 strains. The selection was made using the production of crude toxins by fungus. Helliwell et al. [370] suggested that ethylene plays a major role in resistance against rice blasts. They have generated transgenic plants using ACS2 (1-aminocyclopropane-1-carboxylic acid synthase), which produces ethylene.
The transformed lines showed resistance against blast with little or no difference between the agronomic traits of transgenics and wild-type plants [370]. Devanna et al. [28] isolated the Pi54 gene from a wild species, Oryza officinalis, and it was renamed Pi54, which confers resistance against blast disease. Interestingly this study demonstrated that the Pi54 of the gene was structurally more stable and provided a higher level of resistance as compared to Pi54. This gene was transferred in two susceptible rice lines, IET16310 (indica) and TP309 (japonica), which transformed them into highly resistant strains against Magnoporthe oryzae.
Moreover, Wang et al. [29] transferred MoHrip1 and MoHrip2 genes into rice through an Agrobacterium-tumefaciens-based method, which resulted in resistance to blast disease. The transgenic paddy plants constrained the growth of fungal hyphae and also had a high water-retention capacity. Furthermore, marker-free transgenic rice was generated using maize’s Ac/Ds transposon vectors carrying fluorescent protein (GFP) and red fluorescent protein (mCherry) genetic markers to generate marker-free transgenic plants. Pi21 gene was expressed in these transgenic plants to generate resistance against rice blasts. The transformed lines had good resistance against Magnoporthe oryzae [30]. Moreover, three Pi genes, viz., Pib, Pi25, and Pi54, were transferred together into two rice varieties, the indica variety Kasalath and the japonica variety Zhenghan 10. The transformed varieties exhibited a good level of resistance against blast pathogens, but this gene pyramiding came with its baggage of linkage drag and pleiotropic effects of these genes. The transgenic plants were impairing many gene transcriptions, which ultimately interrupted the normal development of the plants [31].
The genes mentioned in the above studies suggest a potential future for transgenic crops, which will stand against huge threats to the staple food of the world. The genes can be used to generate new varieties or can be used to create strong resistance barriers against major diseases such as blasts. Genes from other organisms or species can be used to achieve this goal without negatively affecting the desired agronomical traits.
7.5. Genome Editing Tools for Developing the Blast Resistance in Rice
The emergence of fatal strains of the rice blast disease is imminent, and the existing tools and available resistance genes might not be enough to cope with their pace. Genome editing tools have provided us with new pathways and have given new perspectives for developing blast-resistant rice varieties. The genes or crops can be targeted to direct their characteristics in a particular direction and to create novel variations [372]. Several methods allow insertion, deletion, mutation, or substitution of nucleotides or sequences of nucleotides in specific locations. This can be accomplished by genome editing using highly advanced genetic engineering tools [373], such as endonucleases which are divided into four categories: Zinc Finger Nucleases (ZFNs), Meganucleases (MNs), chemical nucleases [374] TALENs (Transcription Activator-Like Effector Nucleases) and CRISPR Cas9 (Clustered Regularly Interspaced Short Palindromic Repeats) are the latest addition [375].
7.5.1. CRISPR Cas9 Based Resistance for Rice Blast
In recent years, there have been a lot of changes in the environment that directly or indirectly affect the growth of important crops. Insect and disease infestations have increased drastically, and constant evolution in the strains resistant to existing defense mechanisms has forced the scientific community to develop new and innovative methods to fight against these odds. Rice blast is one such disease that causes a lot of damage to the crop and is considered among the top 10 fungal diseases that could threaten global food security [17,376]. Biotechnology helped researchers to identify genes that can confer resistance to rice blasts in resilient varieties of rice. Recently developed techniques such as CRISPR Cas9 can be very useful in developing varieties that are resistant to the attack of Magnaporthe oryzae. CRISPR is a very effective sequence-specific nuclease (SSN) that targets particular genes and carries out efficient genome editing.
Wang et al. [97] used CRISPR Cas9 to edit the OsERF922 gene in rice. They have induced mutations in the target sites using the C-ERF922 enzyme, which generated several insertions and deletions. Interestingly, these mutations were successfully inherited in consecutive generations. Six mutated lines viz., KS2-12-1-3, KS2-27-4-1, KS2-45-6-1, KS2-70-1-2, KS2-75-1-11, KS2-144-1-2 were selected, which displayed a drastically reduced number of blast lesions and were at par with the agronomic traits when compared with the wild types. Similarly, Zhou et al. [377] attempted to mutate three broad-spectrum blast-resistant genes, i.e., Bsr-d1, Pi21, and ERF922, using CRISPR-based technology in Longke638S (LK638S) rice cultivar which is a TGMS line. They observed that single and triple mutants had improved resistance against blast. Furthermore, the improved varieties also had similar kinds of desirable superior agronomic characteristics. In another experiment, CRISPR Cas9 was used to target S genes, Pi21 and Bsr-d1 in rice which was responsible for the susceptible reaction with rice blast pathogen Magnoporthe oryzae [378]. Due to gene editing, resistance was conferred by two methods, firstly by generating a loss of function mutants and secondly by generating knock of mutants [378]. Similarly, the OsSEC3A gene mutated using targeted mutagenesis, which enhanced resistance to rice blast disease [379]. Many scientists have also utilized the CRISPR Cas9-based gene editing approach to develop resistance in rice against the blast disease [64,379,380], which is presented in Table 6.
Table 6.
Potential use of CRISPR Cas9 for resistance against Magnaporthe oryzae.
7.5.2. Transcription Activator-like Effector Nucleases (TALENs)
TALENs induce site-specific double-strand breaks (DSBs) in DNA [381]. These breaks can be amended by homologous recombination (HR) or non-homologous end-joining (NHEJ). The bacteria of Xanthomonas species secrete proteins known as TALEs (Transcription activator-like effectors) which display pathogenic activity in crops such as rice, tomato, etc. [382]. Further research revealed that these proteins could recognize and bind to specific sequences in DNA [383]. This was followed by the discovery of code which recognized specific regions of DNA [384]. The one monomer nucleotide property of TALE fascinated many researchers to work with it. This technique was used to combat filamentous fungus such as Magnoporthe oryzae, and an HR detection-based PtFg TALEN plasmid was constructed. This highly efficient nuclease can improve the efficiency of HR-mediated gene editing up to 100% and can prove to be very important for developing resistant varieties against various strains of Magnoporthe oryzae [385].
7.5.3. Meganucleases (MNs)
These are also known as homing endonucleases, which are divided into five types based on their sequence and structure motifs [386], from which the LAGLIDADG family proteins have the capacity to act as endonucleases that can bind to specific sites on the DNA [387]. They can recognize exon- or intron-free regions and bind with longer DNA sequences (14 to 40 bp), and induce double-strand breaks (DSBs) in DNA [388]. MNs are not commonly used because naturally occurring endonucleases are very limited and can recognize only a few sites on the genome. Artificial MNs require a huge input of time and money which requires a lot of sustained effort and is practically not feasible. Despite having limited usage, this technique can be kept as an alternative to generate novel genetic changes to develop resistance against various strains of Magnoporthe oryzae.
7.5.4. Zinc Finger Nucleases (ZFNs)
ZFNs are counterfeit for natural endonucleases known as Fokl and are derived from Flavobacterium okeanokoites [389]. They consist of two terminal ends, the N terminal, which binds with DNA, and the C terminal, which has cleavage activity. Each ZFN is capable of identifying a sequence of 3–4 nucleotides. Different ZFNs can be combined to recognize a longer sequence and can be used to induce DSBs in DNA for genome editing [390]. Later, using NHEJ or HR repairs, the required nucleotide sequence or gene can be inserted to achieve targeted results. ZFNs can be introduced into hosts using viral or non-viral vectors and have the capacity to incorporate themselves into any genome, including that of mitochondria [391]. ZFNs have been successfully used to induce mutations or edit genomes in various crops. An experiment was conducted on rice to identify safe sites to introduce new genes using three different ZFN constructs (pZFN1, pZFN2, pZFN3), which were delivered using Agrobacterium and β-glucuronidase (GUS) as a reporter gene. Gene expression was measured using TAIL PCR which identified 28 presumed safe sites which can be used to harbor genes of interest for improving resistance against diseases such as rice blasts [392].
8. Problems Associated with Breeding for Resistance to Rice Blast
Boosting the yield potential and productivity of rice is the primary objective for the plant breeders; however, in complementation, resistance breeding for biotic and abiotic stress has been considered a major issue due to changing climatic conditions [43]. Conventional breeding methods for developing blast-resistant rice varieties are tedious and time-consuming as it takes about 8–10 years to develop one resistant variety. In parallel, Magnoporthe oryzae is mutating continuously and evolving new races against the resistant genes available in the existing cultivars within 2–3 years. In such cases, the resistant cultivar becomes susceptible, leading to discouragement to the breeders as their long efforts have been destroyed quickly. Moreover, low-yielding cultivars that have blast resistance have not been preferred by the farmers and farming community; therefore, breeding for high-yielding rice varieties with multiple durable R-genes for blast resistance is the need of the hour. Advancement in genomic approaches has enabled the breeders to accurately introgress or pyramid multiple R-genes in the high-yielding desirable genotypes within a short period with the help of molecular markers and genomic selection [126,317]. However, linkage drag is the major issue in the case of gene introgression and pyramiding. A number of undesirable changes due to linkage drag in the recipient genotype led to their poor yield performance, and, fortunately, that cannot be compromised by the farmers [19,20,347]. In the current era of genome editing, several scientists have developed blast-resistant and high-yielding rice varieties using genetic engineering and cisgenesis [12,372,378]. However, they have not been accepted worldwide due to the imposition of regulations on genetic modification in many rice-producing and rice-importing countries. Nonetheless, no genetically modified disease-resistant rice is currently commercially available for production worldwide [43]. While these techniques have been proven successful in experimental settings, they have not been made commercially available in rice due to regulatory protocols put in place by many rice-importing countries.
9. Conclusions and Future Outlooks
It is well understood that combining the multiple race-specific R genes in elite cultivars is the most effective and appropriate strategy for developing broad-spectrum and durable resistance to blast disease. However, multiple R-genes in cultivars may promote the evolution of several new races of the pathogen, and even super races could arise, which may cause severe blast epidemics by overcoming the multiple major R-genes. Therefore, it is crucial to rationally utilize the race-specific R-genes in breeding programs to sustain the blast resistance of rice cultivars which is still poorly understood. This opens many avenues for researchers to explore blast resistance in rice. A diagrammatic representation of the possible aspects for further improvement in blast resistance is represented in Figure 5. The major obstacle in managing rice blasts is the durability of the genetic resistance. Thus, the right combination of the major R gene and minor QTLs is necessary to confer broad-spectrum and durable resistance with the help of conventional integrated breeding, advanced genomic approaches, and genome editing tools.
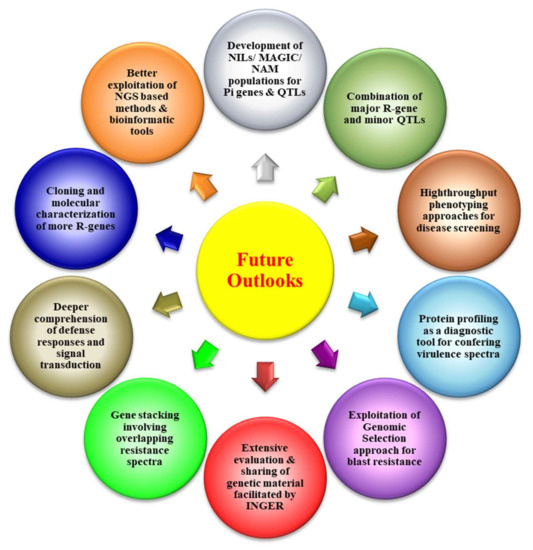
Figure 5.
Future perspectives and way forward for developing blast resistance in rice.
Interestingly, of the 36 molecularly characterized and cloned R genes, several broad-spectrum genes are available for further use in breeding programs. This indicates that along with the mining of novel genes, their cloning and characterization are also important. Furthermore, high-quality Pi genes, which are less prone to evasion by the pathogen and impose fitness penalties in mutations of the pathogen, are expected to be effective and durable. Work on these genes could be accelerated. As neck blast is more damaging to yield and grain quality, screening of neck blast resistance should be incorporated in key evaluation sites. A rice variety, ‘Mahamaya’ of Chhattisgarh, India, is very popular for flaked rice. However, it is susceptible to neck blasts; therefore, there is a need to incorporate resistant genes against the neck blast. Until now, only a few genes have been identified showing the true resistance against the panicle and leaf blast; therefore, mining the R-genes/QTLs imparting equal effectiveness against panicle blast and leaf blast diseases would enhance the resources for the breeders. Moreover, disease screening protocols can be improvised through high throughput phenotyping approaches which could help in better monitoring and management of the disease. With the available genome sequence of the blast fungus, comprehensive profiling of secreted proteins of the fungus is now possible. These proteins may confer effector functions and can act as a diagnostic tool for determining the virulence/avirulence spectrum of a given pathogen population.
Furthermore, the ability to evaluate genetic materials at multiple sites is essential to assess the spectrum of resistance of breeding lines and gain a glimpse of the potential durability. Performance records are organized by the International Network Genetic Evaluation of Rice (INGER) and are used as an indicator of the durability of resistance in varieties. This will require extensive sharing of genetic materials between countries, perhaps through the facilitation of INGER. Gene stacking involving major R genes with overlapping resistance spectra involving superior alleles would confer durable resistance. Integrating advanced breeding methods, modern molecular approaches along with next-generation sequencing (NGS) based methods, and bioinformatic tools will effectively direct in achieving rice blast resistance. High-throughput whole genome sequence (WGS) or target gene sequencing in the elite rice cultivars or core resistant germplasm will provide useful technological means for breeding selection.
In addition, genomic selection should be actively utilized in breeding for blast disease resistance as it exploits the genomic-estimated breeding values of individuals obtained from genome-wide markers to choose candidates for the next breeding cycle. The availability of a standard phenotyping approach and genome-wide high-throughput, cost-effective and flexible markers, especially with the emergence of NGS techniques, has enabled the plant breeders to exploit genomic selection (GS) for crop improvement. The NGS-based genotyping approaches, such as genotyping by sequencing, have significantly improved the prediction level of genomic-estimated breeding values in cereals and other crop species as compared to the other established marker platform and helped in deploying the GS in breeding programs. Furthermore, GS is the most suited approach for breeding for quantitative resistance conferred by minor effect genes, or a combination of minor and major genes tends to produce a more durable resistance in breeding lines (BLs) because it relies on multi-resistant alleles. In addition to increasing the accuracy of selection, GS is expected to reduce rates of inbreeding because the increased accuracy of Mendelian sampling terms in GS allows for the identification and selection of elite breeding candidates from more families, with lower co-selection of sibs. Improved statistical models that leverage genomic information to increase prediction accuracies are critical for the effectiveness of GS-enabled breeding programs.
More studies towards a deeper comprehension of defense responses and signal transduction leading to defense responses are required. NILs (Near Isogenic Lines), Multi-parent advanced generation inter-cross (MAGIC) populations, and Nested Association Mapping (NAM) are useful genetic resources for genetic analysis and, eventually, gene cloning. More NILs, MAGIC, and NAM populations for diverse and durable Pi-genes and QTLs could be developed primarily for genetic analysis and for monitoring pathogen populations. Marker-aided foreground and background selection can be used to accelerate NIL development. More expertise could be involved in performing recently developed transgenic and genome editing tools for its better exploitation to develop blast resistance in rice through specific genetic modifications. Broad spectrum resistance may arise due to the mutations in susceptibility genes in plants. Such loss of function mutations in susceptibility genes indirectly imparts disease resistance without yield penalty. Future studies must orient towards the mining of such S genes in rice in order to utilize them through genome modification techniques for developing blast-resistant varieties.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/plants11182386/s1, Table S1: List of blast disease resistant rice varieties developed through crossing with mutant varieties.
Author Contributions
Conceptualization, P.K.S. and R.S.; framework and outline: P.K.S.; writing—original draft preparation, P.K.S., R.S., D.K.C., A.T., V.K.; figure preparation, P.K.S., D.K.C., R.S.; writing—initial review and editing, P.K.S., R.S.; writing—final review and editing, S.M., D.S., B.K.D., L.J.; supervision, S.M., D.S. and B.K.D.; project administration, L.J.; funding acquisition, D.S., L.J. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
Authors are heartily thankful to the International Atomic Energy Agency (IAEA), Vienna, Austria for publication of this manuscript through IAEA CRP “D23032”, Contract No: 24661 (Identifica-tion and Mapping of Blast-resistant Genes in Rice Mutant, Vikram-TCR through NGS based MutMap Approach).
Conflicts of Interest
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
References
- Simkhada, K.; Thapa, R. Rice Blast, A Major Threat to the Rice Production and Its Various Management Techniques. Turkish J. Agric. Food Sci. Technol. 2022, 10, 147–157. [Google Scholar] [CrossRef]
- Asibi, E.A.; Chai, Q.; Coulter, J.A. Rice Blast: A Disease with Implications for Global Food Security. Agronomy 2019, 9, 451. [Google Scholar] [CrossRef]
- Muthayya, S.; Sugimoto, J.D.; Montgomery, S.; Maberly, G.F. An Overview of Global Rice Production, Supply, Trade, and Consumption. Ann. N. Y. Acad. Sci. 2014, 1324, 7–14. [Google Scholar] [CrossRef] [PubMed]
- Carrillo, M.G.C.; Martin, F.; Variar, M.; Bhatt, J.C.; Perez-Quintero, A.; Leung, H.; Leach, J.E.; Vera Cruz, C.M. Accumulating Candidate Genes for Broad-Spectrum Resistance to Rice Blast in a Drought-Tolerant Rice Cultivar. Sci. Rep. 2021, 11, 21502. [Google Scholar] [CrossRef] [PubMed]
- Sasaki, T.; Burr, B. International rice genome sequencing project: The effort to completely sequence the rice genome. Curr. Opin. Plant Biol. 2000, 3, 138–142. [Google Scholar] [CrossRef]
- Thapa, R.; Bhusal, N. Designing Rice for the 22nd Century: Towards Rice with an Enhanced Productivity and Efficient Photosynthetic Pathway. Turkish J. Agric. Food Sci. Technol. 2020, 8, 2623–2634. [Google Scholar] [CrossRef]
- Acharya, B.; Shrestha, S.M.; Manandhar, H.K.; Chaudhary, B. Screening of Local, Improved and Hybrid Rice Genotypes against Leaf Blast Disease (Pyricularia oryzae) at Banke District, Nepal. J. Agric. Nat. Resour. 2019, 2, 36–52. [Google Scholar] [CrossRef]
- Wilson, R.A.; Talbot, N.J. Under Pressure: Investigating the Biology of Plant Infection by Magnaporthe oryzae. Nat. Rev. Microbiol. 2009, 7, 185–195. [Google Scholar] [CrossRef]
- Dean, R.; Van Kan, J.A.L.; Pretorius, Z.A.; Hammond-Kosack, K.E.; DI Pietro, A.; Spanu, P.D.; Rudd, J.J.; Dickman, M.; Kahmann, R.; Ellis, J.; et al. The Top 10 Fungal Pathogens in Molecular Plant Pathology. Mol. Plant Pathol. 2012, 13, 414–430. [Google Scholar] [CrossRef]
- Agrios, G. Plant Pathology, 5th ed.; Elsevier Academic Press: Cambridge, MA, USA, 2005; p. 165. [Google Scholar]
- Nasruddin, A.; Amin, N. Effects of Cultivar, Planting Period, and Fungicide Usage on Rice Blast Infection Levels and Crop Yield. J. Agric. Sci. 2012, 5, P160. [Google Scholar] [CrossRef]
- Miah, G.; Rafii, M.Y.; Ismail, M.R.; Puteh, A.B.; Rahim, H.A.; Asfaliza, R.; Latif, M.A. Blast resistance in rice: A review of conventional breeding to molecular approaches. Mol. Biol. Rep. 2013, 40, 2369–2388. [Google Scholar] [CrossRef] [PubMed]
- Musiime, O.; Tenywa, M.M.; Majaliwa, M.J.G.; Lufafa, A.; Nanfumba, D.; Wasige, J.E.; Woomer, P.L.; Kyondha, M. Constraints to rice production in Bugiri District. Afr. Crop Sci. Conf. Proc. 2005, 7, 1495–1499. [Google Scholar]
- Kato, H. Rice blast disease. Pestic. Outlook 2001, 12, 23–25. [Google Scholar] [CrossRef]
- Yu, Y.; Ma, L.; Wang, X.; Zhao, Z.; Wang, W.; Fan, Y.; Liu, K.; Jiang, T.; Xiong, Z.; Song, Q.; et al. Genome-Wide Association Study Identifies a Rice Panicle Blast Resistance Gene, Pb2, Encoding NLR Protein. Int. J. Mol. Sci. 2022, 23, 5668. [Google Scholar] [CrossRef]
- Yadav, M.K.; Aravindan, S.; Ngangkham, U.; Raghu, S.; Prabhukarthikeyan, S.R.; Keerthana, U.; Marndi, B.C.; Adak, T.; Munda, S.; Deshmukh, R.; et al. Blast Resistance in Indian Rice Landraces: Genetic Dissection by Gene Specific Markers. PLoS ONE 2019, 14, e0211061. [Google Scholar] [CrossRef]
- Ashkani, S.; Rafii, M.Y.; Shabanimofrad, M.; Ghasemzadeh, A.; Ravanfar, S.A.; Latif, M.A. Molecular Progress on the Mapping and Cloning of Functional Genes for Blast Disease in Rice (Oryza sativa L.): Current Status and Future Considerations. Crit. Rev. Biotechnol. 2016, 36, 353–367. [Google Scholar] [CrossRef]
- Wang, G.L.; Valent, B. Durable Resistance to Rice Blast. Science 2017, 355, 906–907. [Google Scholar] [CrossRef]
- Srivastava, D.; Shamim, M.; Kumar, M.; Mishra, A.; Pandey, P.; Kumar, D.; Yadav, P.; Siddiqui, M.H.; Singh, K.N. Current Status of Conventional and Molecular Interventions for Blast Resistance in Rice. Rice Sci. 2017, 24, 299–321. [Google Scholar] [CrossRef]
- Singh, B.D. Plant Breeding: Principles and Methods; Kalyani Publishers: Ludhiana, India, 2018; Volume 11, pp. 1–907. [Google Scholar]
- Lo, K.-L.; Chen, Y.-N.; Chiang, M.-Y.; Chen, M.-C.; Panibe, J.P.; Chiu, C.-C.; Liu, L.-W.; Chen, L.-J.; Chen, C.-W.; Li, W.-H.; et al. Two Genomic Regions of a Sodium Azide Induced Rice Mutant Confer Broad-Spectrum and Durable Resistance to Blast Disease. Rice 2022, 15, 2. [Google Scholar] [CrossRef]
- Ning, X.; Yunyu, W.; Aihong, L. Strategy for Use of Rice Blast Resistance Genes in Rice Molecular Breeding. Rice Sci. 2020, 27, 263–277. [Google Scholar] [CrossRef]
- Annegowda, D.C.; Prasannakumar, M.K.; Mahesh, H.B.; Siddabasappa, C.B.; Devanna, P.; Banakar, S.N.; Manijkumar, H.B.; Prassad, S.R. Rice Blast Disease in India: Present Status and Future Challenges. In Integrative Advances in Rice Research; IntechOpen Limited: London, UK, 2022; pp. 1–42. [Google Scholar]
- Sharma, S.K.; Sharma, D.; Meena, R.P.; Yadav, M.K.; Hosahatti, R.; Dubey, A.K.; Sharma, P.; Kumar, S.; Pramesh, D.; Nabi, S.U.; et al. Recent Insights in Rice Blast Disease Resistance. In Blast Disease of Cereal Crops; Nayaka, S.C., Hosahatti, R., Prakash, G., Satyavathi, C.T., Sharma, R., Eds.; Fungal Biology; Springer Nature Switzerland AG: Cham, Switzerland, 2021; pp. 89–124. [Google Scholar]
- Mao, T.; Zhu, M.; Ahmad, S.; Ye, G.; Sheng, Z.; Hu, S.; Jiao, G.; Xie, L.; Tang, S.; Wei, X.; et al. Superior Japonica Rice Variety YJ144 with Improved Rice Blast Resistance, Yield, and Quality Achieved Using Molecular Design and Multiple Breeding Strategies. Mol. Breed. 2021, 41, 65. [Google Scholar] [CrossRef] [PubMed]
- Kumari, A.; Das, A.; Devanna, B.N.; Thakur, S.; Singh, P.K.; Singh, N.K.; Sharma, T.R. Mining of Rice Blast Resistance Gene Pi54 Shows Effect of Single Nucleotide Polymorphisms on Phenotypic Expression of the Alleles. Eur. J. Plant Pathol. 2013, 137, 55–65. [Google Scholar] [CrossRef]
- Wang, X.; Guo, X.; Ma, X.; Luo, L.; Fang, Y.; Zhao, N.; Han, Y.; Wei, Z.; Liu, F.; Qin, B.; et al. Development of New Rice (Oryza. Sativa L.) Breeding Lines through Marker-Assisted Introgression and Pyramiding of Brown Planthopper, Blast, Bacterial Leaf Blight Resistance, and Aroma Genes. Agronomy 2021, 11, 2525. [Google Scholar] [CrossRef]
- Devanna, N.B.; Vijayan, J.; Sharma, T.R. The Blast Resistance Gene Pi54of Cloned from Oryza officinalis Interacts with Avr-Pi54 through Its Novel Non-LRR Domains. PLoS ONE 2014, 9, e104840. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Han, Q.; Zi, Q.; Lv, S.; Qiu, D.; Zeng, H. Enhanced Disease Resistance and Drought Tolerance in Transgenic Rice Plants Overexpressing Protein Elicitors from Magnaporthe oryzae. PLoS ONE 2017, 12, e0175734. [Google Scholar] [CrossRef]
- Li, X.; Pan, L.; Bi, D.; Tian, X.; Li, L.; Xu, Z.; Wang, L.; Zou, X.; Gao, X.; Yang, H.; et al. Generation of Marker-Free Transgenic Rice Resistant to Rice Blast Disease Using Ac/Ds Transposon-Mediated Transgene Reintegration System. Front. Plant Sci. 2021, 12, 644437. [Google Scholar] [CrossRef]
- Peng, M.; Lin, X.; Xiang, X.; Ren, H.; Fan, X.; Chen, K. Characterization and Evaluation of Transgenic Rice Pyramided with the Pi Genes Pib, Pi25 and Pi54. Rice 2021, 14, 78. [Google Scholar] [CrossRef]
- Viana, V.E.; Pegoraro, C.; Busanello, C.; Costa de Oliveira, A. Mutagenesis in Rice: The Basis for Breeding a New Super Plant. Front. Plant Sci. 2019, 10, 1326. [Google Scholar] [CrossRef]
- Ijaz, S.; Ul Haq, I. Genome Editing Technologies for Resistance Against Phytopathogens: Principles, Applications and Future Prospects. In Plant Disease Management Strategies for Sustainable Agriculture through Traditional and Modern Approaches; Springer Nature Switzerland AG: Cham, Switzerland, 2020; pp. 237–245. [Google Scholar]
- Nizolli, V.O.; Pegoraro, C.; Oliveira, A.C. Rice blast: Strategies and challenges for improving genetic resistance. Crop. Breed. Appl. Biotechnol. 2021, 21, e387721S9. [Google Scholar] [CrossRef]
- Deng, Y.; Zhai, K.; Xie, Z.; Yang, D.; Zhu, X.; Liu, J.; Wang, X.; Qin, P.; Yang, Y.; Zhang, G.; et al. Epigenetic Regulation of Antagonistic Receptors Confers Rice Blast Resistance with Yield Balance. Science 2017, 355, 962–965. [Google Scholar] [CrossRef]
- Couch, B.C.; Fudal, I.; Lebrun, M.-H.; Tharreau, D.; Valent, B.; van Kim, P.; Nottéghem, J.-L.; Kohn, L.M. Origins of host-specific populations of the blast pathogen Magnaporthe oryzae in crop domestication with subsequent expansion of pandemic clones on rice and weeds of rice. Genetics 2005, 170, 613–630. [Google Scholar] [CrossRef]
- Padmanabhan, S.Y. Recent advances in the study of blast disease of rice. Madras Agri. J. 1965, 564–583. [Google Scholar]
- Waller, J.M. Rice Diseases. By S. H. Ou Slough, UK: Commonwealth Agricultural Bureaux (1985) 2nd Ed., pp. 380, UK £38.00, USA $70, Elsewhere £41.00. Exp. Agric. 1987, 23, 357. [Google Scholar] [CrossRef]
- Khush, G.S. Breeding methods and procedures employed at IRRI for developing rice germplasm with multiple resistance to diseases and insects. In Symposium on Methods of Crop Breeding; Tropical Agricultural Research Series; Japan International Research Center for Agricultural Sciences: Tsukuba, Japan, 1978; Volume 11, pp. 69–76. [Google Scholar]
- Padmanabhan, S.Y.; Chakravarti, N.K.; Mathur, S.C.; Veeraraghavan, J. Identification of pathogenic races of Pyricularia Oryzae in India. Phytopathology 1970, 60, 1574–1577. [Google Scholar] [CrossRef]
- Piotti, E.; Rigano, M.M.; Rodino, D.; Rodolfi, M.; Castiglione, S.; Picco, A.M.; Sala, F. Genetic Structure of Pyricularia Grisea (Cooke) Sacc. Isolates from Italian Paddy Fields. J. Phytopathol. 2005, 153, 80–86. [Google Scholar] [CrossRef]
- Devanna, B.N.; Jain, P.; Solanke, A.U.; Das, A.; Thakur, S.; Singh, P.K.; Kumari, M.; Dubey, H.; Jaswal, R.; Pawar, D.; et al. Understanding the Dynamics of Blast Resistance in Rice-Magnaporthe oryzae Interactions. J. Fungi 2022, 8, 584. [Google Scholar] [CrossRef] [PubMed]
- Nalley, L.; Tsiboe, F.; Durand-Morat, A.; Shew, A.; Thoma, G. Economic and Environmental Impact of Rice Blast Pathogen (Magnaporthe oryzae) Alleviation in the United States. PLoS ONE 2016, 11, e0167295. [Google Scholar] [CrossRef] [PubMed]
- Parker, D.; Beckmann, M.; Enot, D.P.; Overy, D.P.; Rios, Z.C.; Gilbert, M.; Talbot, N.; Draper, J. Rice Blast Infection of BrachypodiumDistachyon as a Model System to Study Dynamic Host/Pathogen Interactions. Nat. Protoc. 2008, 3, 435–445. [Google Scholar] [CrossRef]
- Howard, R.J.; Ferrari, M.A.; Roach, D.H.; Money, N.P. Penetration of hard substrates by a fungus employing enormous turgor pressures. Proc. Natl. Acad. Sci. USA 1991, 88, 11281–11284. [Google Scholar] [CrossRef] [PubMed]
- Staskawicz, B.J.; Ausubel, F.M.; Baker, B.J.; Ellis, J.G.; Jones, J.D.G. Molecular Genetics of Plant Disease Resistance. Science 1995, 268, 661–667. [Google Scholar] [CrossRef]
- Hamer, J.E.; Howard, R.J.; Chumley, F.G.; Valent, B. A mechanism for surface attachment in spores of a plant pathogenic fungus. Science 1988, 239, 288–290. [Google Scholar] [CrossRef]
- Jelitto, T.C.; Page, H.A.; Read, N.D. Role of external signals in regulating the pre-penetration phase of infection by the rice blast fungus, Magnaporthe grisea. Planta 1994, 194, 471–477. [Google Scholar] [CrossRef]
- Xiao, J.-Z.; Watanabe, T.; Kamakura, T.; Ohshima, A.; Yamaguchi, I. Studies on Cellular Differentiation of Magnaporthe grisea. Physicochemical Aspects of Substratum Surfaces in Relation to Appressorium Formation. Physiol. Mol. Plant Pathol. 1994, 44, 227–236. [Google Scholar] [CrossRef]
- Jiang, H.; Lin, L.; Tang, W.; Chen, X.; Zheng, Q.; Huang, J.; Wang, Z. Putative interaction proteins of the ubiquitin ligase Hrd1 in Magnaporthe oryzae. Evol. Bioinform. 2018, 14, 1176934318810990. [Google Scholar] [CrossRef] [PubMed]
- Li, G.; Zhou, X.; Xu, J.R. Genetic control of infection-related development in Magnaporthe oryzae. Curr. Opin. Microbiol. 2012, 15, 678–684. [Google Scholar] [CrossRef]
- Gao, J.; Xu, X.; Huang, K.; Liang, Z. Fungal G-Protein-Coupled Receptors: A Promising Mediator of the Impact of Extracellular Signals on Biosynthesis of Ochratoxin A. Front. Microbiol. 2021, 12, 631392. [Google Scholar] [CrossRef]
- DeZwaan, T.M.; Carroll, A.M.; Valent, B.; Sweigard, J.A. Magnaporthe grisea Pth11p Is a Novel Plasma Membrane Protein That Mediates Appressorium Differentiation in Response to Inductive Substrate Cues. Plant Cell 1999, 11, 2013–2030. [Google Scholar] [CrossRef]
- Kou, Y.; Tan, Y.H.; Ramanujam, R.; Naqvi, N.I. Structure–function analyses of the Pth11 receptor reveal an important role for CFEM motif and redox regulation in rice blast. New Phytol. 2017, 214, 330–342. [Google Scholar] [CrossRef]
- Zhang, H.; Tang, W.; Liu, K.; Huang, Q.; Zhang, X.; Yan, X.; Chen, Y.; Wang, J.; Qi, Z.; Wang, Z.; et al. Eight RGS and RGS-like Proteins Orchestrate Growth, Differentiation, and Pathogenicity of Magnaporthe oryzae. PLoS Pathog. 2011, 7, e1002450. [Google Scholar] [CrossRef]
- Dean, R.A. signal pathways and appressorium morphogenesis. Annu. Rev. Phytopathol. 1997, 35, 211–234. [Google Scholar] [CrossRef]
- Tucker, S.L.; Talbot, N.J. Surface Attachment and Pre-Penetration Stage Development by Plant Pathogenic Fungi. Annu. Rev. Phytopathol. 2001, 39, 385–417. [Google Scholar] [CrossRef]
- De Jong, J.C.; McCormack, B.J.; Smirnoff, N.; Talbot, N.J. Glycerol Generates Turgor in Rice Blast. Nature 1997, 389, 244. [Google Scholar] [CrossRef]
- Shen, Q.; Liang, M.; Yang, F.; Deng, Y.Z.; Naqvi, N.I. Ferroptosis Contributes to Developmental Cell Death in Rice Blast. New Phytol. 2020, 227, 1831–1846. [Google Scholar] [CrossRef]
- Talbot, N.J.; Kershaw, M.J. The Emerging Role of Autophagy in Plant Pathogen Attack and Host Defence. Curr. Opin. Plant Biol. 2009, 12, 444–450. [Google Scholar] [CrossRef]
- Yin, Z.; Feng, W.; Chen, C.; Xu, J.; Li, Y.; Yang, L.; Wang, J.; Liu, X.; Wang, W.; Gao, C.; et al. Shedding Light on Autophagy Coordinating with Cell Wall Integrity Signaling to Govern Pathogenicity of Magnaporthe oryzae. Autophagy 2020, 16, 900–916. [Google Scholar] [CrossRef] [PubMed]
- Ryder, L.S.; Talbot, N.J. Regulation of Appressorium Development in Pathogenic Fungi. Curr. Opin. Plant Biol. 2015, 26, 8–13. [Google Scholar] [CrossRef]
- Fang, Y.L.; Xia, L.M.; Wang, P.; Zhu, L.H.; Ye, J.R.; Huang, L. The MAPKKK CgMck1 is required for cell wall integrity, appressorium development, and pathogenicity in Colletotrichum gloeosporioides. Genes 2018, 9, 543. [Google Scholar] [CrossRef] [PubMed]
- Foster, A.J.; Ryder, L.S.; Kershaw, M.J.; Talbot, N.J. The Role of Glycerol in the Pathogenic Lifestyle of the Rice Blast Fungus Magnaporthe oryzae. Environ. Microbiol. 2017, 19, 1008–1016. [Google Scholar] [CrossRef] [PubMed]
- Sweigard, J.A.; Chumley, F.G.; Valent, B. Disruption of a Magnaporthe grisea Cutinase Gene. Mol. Gen. Genet. 1992, 232, 183–190. [Google Scholar] [CrossRef]
- Skamnioti, P.; Gurr, S.J. Magnaporthe grisea cutinase2 mediates appressorium differentiation and host penetration and is required for full virulence. Plant Cell 2007, 19, 2674–2689. [Google Scholar] [CrossRef] [PubMed]
- Quoc, N.B.; Bao Chau, N.N. The role of cell wall degrading enzymes in pathogenesis of Magnaporthe oryzae. Curr. Protein Pept. Sci. 2017, 18, 1019–1034. [Google Scholar] [CrossRef] [PubMed]
- Dagdas, Y.F.; Yoshino, K.; Dagdas, G.; Ryder, L.S.; Bielska, E.; Steinberg, G.; Talbot, N.J. Septin-mediated plant cell invasion by the rice blast fungus, Magnaporthe oryzae. Science 2012, 336, 1590–1595. [Google Scholar] [CrossRef]
- Kleemann, J.; Rincon-Rivera, L.J.; Takahara, H.; Neumann, U.; van Themaat, E.V.L.; van der Does, H.C.; O’Connell, R.J. Sequential delivery of host-induced virulence effectors by appressoria and intracellular hyphae of the phytopathogen Colletotrichum higginsianum. PLoS Pathog. 2012, 8, e1002643. [Google Scholar] [CrossRef]
- Ryder, L.S.; Dagdas, Y.F.; Kershaw, M.J.; Venkataraman, C.; Madzvamuse, A.; Yan, X.; Cruz-Mireles, N.; Soanes, D.M.; Oses-Ruiz, M.; Styles, V.; et al. A Sensor Kinase Controls Turgor-Driven Plant Infection by the Rice Blast Fungus. Nature 2019, 574, 423–427. [Google Scholar] [CrossRef]
- Ryder, L.S.; Dagdas, Y.F.; Mentlak, T.A.; Kershaw, M.J.; Thornton, C.R.; Schuster, M.; Chen, J.; Wang, Z.; Talbot, N.J. NADPH Oxidases Regulate Septin-Mediated Cytoskeletal Remodeling during Plant Infection by the Rice Blast Fungus. Proc. Natl. Acad. Sci. USA 2013, 110, 3179–3184. [Google Scholar] [CrossRef] [PubMed]
- Egan, M.J.; Wang, Z.Y.; Jones, M.A.; Smirnoff, N.; Talbot, N.J. Generation of reactive oxygen species by fungal NADPH oxidases is required for rice blastdisease. Proc. Natl. Acad. Sci. USA 2007, 104, 11772–11777. [Google Scholar] [CrossRef]
- Khang, C.H.; Berruyer, R.; Giraldo, M.C.; Kankanala, P.; Park, S.Y.; Czymmek, K.; Valent, B. Translocation of Magnaporthe oryzae effectors into rice cells and their subsequent cell-to-cell movement. Plant Cell 2010, 22, 1388–1403. [Google Scholar] [CrossRef] [PubMed]
- Giraldo, M.C.; Dagdas, Y.F.; Gupta, Y.K.; Mentlak, T.A.; Yi, M.; Martinez-Rocha, A.L.; Saitoh, H.; Terauchi, R.; Talbot, N.J.; Valent, B. Two Distinct Secretion Systems Facilitate Tissue Invasion by the Rice Blast Fungus Magnaporthe oryzae. Nat. Commun. 2013, 4, 1996. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Xu, J.R. Effectors and effector delivery in Magnaporthe oryzae. PLoS Pathog. 2014, 10, e1003826. [Google Scholar] [CrossRef]
- Petit-Houdenot, Y.; Fudal, I. Complex interactions between fungal avirulence genes and their corresponding plant resistance genes and consequences for disease resistance management. Front. Plant Sci. 2017, 8, 1072. [Google Scholar] [CrossRef]
- Park, C.H.; Chen, S.; Shirsekar, G.; Zhou, B.; Khang, C.H.; Songkumarn, P.; Wang, G.L. The Magnaporthe oryzae effector AvrPiz-t targets the RING E3 ubiquitin ligase APIP6 to suppress pathogen-associated molecular pattern–triggered immunity in rice. Plant Cell 2012, 24, 4748–4762. [Google Scholar] [CrossRef]
- Kang, S.; Sweigard, J.A.; Valent, B. The PWL host specificity gene family in the blast fungus Magnaporthe grisea. Mol. Plant Microbe Interact. 1995, 8, 939–948. [Google Scholar] [CrossRef]
- Orbach, M.J.; Farrall, L.; Sweigard, J.A.; Chumley, F.G.; Valent, B. A telomeric avirulence gene determines efficacy for the rice blast resistance gene Pi-ta. Plant Cell 2000, 12, 2019–2032. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Wang, B.; Wu, J.; Lu, G.; Hu, Y.; Zhang, X.; Zhou, B. The Magnaporthe oryzae avirulence gene AvrPiz-t encodes a predicted secreted protein that triggers the immunity in rice mediated by the blast resistance gene Piz-t. Mol. Plant-Microbe Interact. 2009, 22, 411–420. [Google Scholar] [CrossRef] [PubMed]
- Yoshida, K.; Saitoh, H.; Fujisawa, S.; Kanzaki, H.; Matsumura, H.; Yoshida, K.; Tosa, Y.; Chuma, I.; Takano, Y.; Win, J.; et al. Association Genetics Reveals Three Novel Avirulence Genes from the Rice Blast Fungal Pathogen Magnaporthe oryzae. Plant Cell 2009, 21, 1573–1591. [Google Scholar] [CrossRef] [PubMed]
- Cesari, S.; Thilliez, G.; Ribot, C.; Chalvon, V.; Michel, C.; Jauneau, A.; Rivas, S.; Alaux, L.; Kanzaki, H.; Okuyama, Y.; et al. The rice resistance protein pair rga4/rga5 recognizes the Magnaporthe oryzae effectors AVR-Pia and AVR1-CO39 by direct binding. Plant Cell 2013, 25, 1463–1481. [Google Scholar] [CrossRef] [PubMed]
- Wu, W.; Wang, L.; Zhang, S.; Li, Z.; Zhang, Y.; Lin, F.; Pan, Q. Stepwise arms race between AvrPik and Pik alleles in the rice blast pathosystem. Mol. Plant-Microbe Interact. 2014, 27, 759–769. [Google Scholar] [CrossRef]
- Zhang, S.; Wang, L.; Wu, W.; He, L.; Yang, X.; Pan, Q. Function and Evolution of Magnaporthe oryzae Avirulence Gene AvrPib Responding to the Rice Blast Resistance Gene Pib. Sci. Rep. 2015, 5, 11642. [Google Scholar] [CrossRef]
- Zhang, X.; He, D.; Zhao, Y.; Cheng, X.; Zhao, W.; Taylor, I.A.; Yang, J.; Liu, J.; Peng, Y.-L. A Positive-Charged Patch and Stabilized Hydrophobic Core Are Essential for Avirulence Function of AvrPib in the Rice Blast Fungus. Plant J. 2018, 96, 133–146. [Google Scholar] [CrossRef]
- Mosquera, G.; Giraldo, M.C.; Khang, C.H.; Coughlan, S.; Valent, B. Interaction transcriptome analysis identifies Magnaporthe oryzae BAS1-4 as biotrophy-associated secreted proteins in rice blast disease. Plant Cell 2009, 21, 1273–1290. [Google Scholar] [CrossRef]
- Talbot, N.J.; Ebbole, D.J.; Hamer, J.E. Identification and Characterization of MPG1, a Gene Involved in Pathogenicity from the Rice Blast Fungus Magnaporthe grisea. Plant Cell 1993, 5, 1575–1590. [Google Scholar] [CrossRef]
- Ahn, N.; Kim, S.; Choi, W.; Im, K.H.; Lee, Y.H. Extracellular matrix protein gene, EMP1, is required for appressorium formation and pathogenicity of the rice blast fungus, Magnaporthe grisea. Mol. Cells 2004, 17, 166–173. [Google Scholar]
- Kim, S.; Ahn, I.P.; Rho, H.S.; Lee, Y.H. MHP1, a Magnaporthe grisea hydrophobin gene, is required for fungal development and plant colonization. Mol. Microbiol. 2005, 57, 1224–1237. [Google Scholar] [CrossRef] [PubMed]
- Mentlak, T.A.; Kombrink, A.; Shinya, T.; Ryder, L.S.; Otomo, I.; Saitoh, H.; Talbot, N.J. Effector-mediated suppression of chitin-triggered immunity by Magnaporthe oryzae is necessary for rice blast disease. Plant Cell 2012, 24, 322–335. [Google Scholar] [CrossRef]
- Saitoh, H.; Fujisawa, S.; Mitsuoka, C.; Ito, A.; Hirabuchi, A.; Ikeda, K.; Irieda, H.; Yoshino, K.; Yoshida, K.; Matsumura, H.; et al. Large-Scale Gene Disruption in Magnaporthe oryzae Identifies MC69, a Secreted Protein Required for Infection by Monocot and Dicot Fungal Pathogens. PLoS Pathog. 2012, 8, e1002711. [Google Scholar] [CrossRef] [PubMed]
- Dong, Y.; Li, Y.; Zhao, M.; Jing, M.; Liu, X.; Liu, M.; Zhang, Z. Global genome and transcriptome analyses of Magnaporthe oryzae epidemic isolate 98-06 uncover novel effectors and pathogenicity-related genes, revealing gene gain and lose dynamics in genome evolution. PLoS Pathog. 2015, 11, e1004801. [Google Scholar] [CrossRef] [PubMed]
- Mogga, V.; Delventhal, R.; Weidenbach, D.; Langer, S.; Bertram, P.M.; Andresen, K.; Schaffrath, U. Magnaporthe oryzae effectors MoHEG13 and MoHEG16 interfere with host infection and MoHEG13 counteracts cell death caused by Magnaporthe-NLPs in tobacco. Plant Cell Rep. 2016, 35, 1169–1185. [Google Scholar] [CrossRef]
- Sharpee, W.; Oh, Y.; Yi, M.; Franck, W.; Eyre, A.; Okagaki, L.H.; Valent, B.; Dean, R.A. Identification and Characterization of Suppressors of Plant Cell Death (SPD) Effectors from Magnaporthe oryzae. Mol. Plant Pathol. 2017, 18, 850–863. [Google Scholar] [CrossRef] [PubMed]
- Chen, M.; Zeng, H.; Qiu, D.; Guo, L.; Yang, X.; Shi, H.; Zhao, J. Purification and characterization of a novel hypersensitive response-inducing elicitor from Magnaporthe oryzae that triggers defense response in rice. PLoS ONE 2012, 7, e37654. [Google Scholar] [CrossRef]
- Chen, C.; Lian, B.; Hu, J.; Zhai, H.; Wang, X.; Venu, R.; Liu, E.; Wang, Z.; Chen, M.; Wang, B.; et al. Genome comparison of two Magnaporthe oryzae field isolates reveals genome variations and potential virulence effectors. BMC Genom. 2013, 14, 887. [Google Scholar] [CrossRef]
- Wang, F.; Wang, C.; Liu, P.; Lei, C.; Hao, W.; Gao, Y.; Liu, Y.-G.; Zhao, K. Enhanced rice blast resistance by CRISPR/Cas9-Targeted Mutagenesis of the ERF Transcription Factor Gene OsERF922. PLoS ONE 2016, 11, e0154027. [Google Scholar] [CrossRef] [PubMed]
- Fang, Y.L.; Peng, Y.-L.; Fan, J. The Nep1-like Protein Family of Magnaporthe oryzae Is Dispensable for the Infection of Rice Plants. Sci. Rep. 2017, 7, 4372. [Google Scholar] [CrossRef] [PubMed]
- Hong, Y.; Yang, Y.; Zhang, H.; Huang, L.; Li, D.; Song, F. Overexpression of MoSM1, encoding for an immunity-inducing protein from Magnaporthe oryzae, in rice confers broad-spectrum resistance against fungal and bacterial diseases. Sci. rep. 2017, 7, 41037. [Google Scholar] [CrossRef] [PubMed]
- Guo, X.; Zhong, D.; Xie, W.; He, Y.; Zheng, Y.; Lin, Y.; Chen, S. Functional identification of novel cell death-inducing effector proteins from Magnaporthe oryzae. Rice 2019, 12, 59. [Google Scholar] [CrossRef]
- Guillen, K.; Ortiz-Vallejo, D.; Gracy, J.; Fournier, E.; Kroj, T.; Padilla, A. Structure Analysis Uncovers a Highly Diverse but Structurally Conserved Effector Family in Phytopathogenic Fungi. PLOS Pathog. 2015, 11, e1005228. [Google Scholar] [CrossRef] [PubMed]
- Krause, R.A.; Webster, R.K. The morphology, taxonomy, and sexuality of the rice stem rot fungus, Magnaporthesalvinii (Leptosphaeria salvinii). Mycologia 1972, 64, 103–114. [Google Scholar] [CrossRef]
- Gowrisri, N.; Kamalakannan, A.; Malathi, V.G.; Rajendran, L.; Rajesh, S. Morphological and molecular characterization of Magnaporthe oryzae B. Couch, inciting agent of rice blast disease. Madras Agric. J. 2019, 106, 255–260. [Google Scholar]
- Xue, M.; Yang, J.; Li, Z.; Hu, S.; Yao, N.; Dean, R.A.; Zhao, W.; Shen, M.; Zhang, H.; Li, C.; et al. Comparative Analysis of the Genomes of Two Field Isolates of the Rice Blast Fungus Magnaporthe oryzae. PLoS Genet. 2012, 8, e1002869. [Google Scholar] [CrossRef]
- Kumar, A.; Sheoran, N.; Prakash, G.; Ghosh, A.; Chikara, S.K.; Rajashekara, H.; Singh, U.D.; Aggarwal, R.; Jain, R.K. Genome sequence of a unique Magnaporthe oryzae RMg-Dl isolate from India that causes blast disease in diverse cereal crops, obtained using PacBio single-molecule and Illumina HiSeq2500 sequencing. Genome Announc. 2017, 5, 1570. [Google Scholar] [CrossRef]
- Dean, R.A.; Talbot, N.J.; Ebbole, D.J.; Farman, M.L.; Mitchell, T.K.; Orbach, M.J.; Thon, M.; Kulkarni, R.; Xu, J.R.; Pan, H.; et al. The genome sequence of the rice blast fungus Magnaporthe grisea. Nature 2005, 434, 980–986. [Google Scholar] [CrossRef] [PubMed]
- Gowda, M.; Shirke, M.D.; Mahesh, H.B.; Chandarana, P.; Rajamani, A.; Chattoo, B.B. Genomeanalysisofrice-blastfungus Magnaporthe oryzae field isolates from southern India. Genom. Data 2015, 5, 284–291. [Google Scholar] [CrossRef][Green Version]
- Suresh, R.; Mosser, D.M. Pattern Recognition Receptors in Innate Immunity, Host Defense, and Immunopathology. Adv. Physiol. Educ. 2013, 37, 284–291. [Google Scholar] [CrossRef] [PubMed]
- Jones, J.D.; Dangl, J.L. The plant immune system. Nature 2006, 444, 323–329. [Google Scholar] [CrossRef]
- Boller, T.; He, S.Y. Innate Immunity in Plants: An Arms Race between Pattern Recognition Receptors in Plants and Effectors in Microbial Pathogens. Science 2009, 324, 742–744. [Google Scholar] [CrossRef]
- Gassmann, W.; Bhattacharjee, S. Effector-Triggered Immunity Signaling: From Gene-for-Gene Pathways to Protein-Protein Interaction Networks. Mol. Plant-Microbe Interact. 2012, 25, 862–868. [Google Scholar] [CrossRef]
- Kaku, H.; Nishizawa, Y.; Ishii-Minami, N.; Akimoto-Tomiyama, C.; Dohmae, N.; Takio, K.; Shibuya, N. Plant cells recognize chitin fragments for defense signaling through a plasma membrane receptor. Proc. Natl. Acad. Sci. USA 2006, 103, 11086–11091. [Google Scholar] [CrossRef] [PubMed]
- Su, J.; Wang, W.; Han, J.; Chen, S.; Wang, C.; Zeng, L.; Feng, A.; Yang, J.; Zhou, B.; Zhu, X. Functional Divergence of Duplicated Genes Results in a Novel Blast Resistance Gene Pi50 at the Pi2/9 Locus. Theor. Appl. Genet. 2015, 128, 2213–2225. [Google Scholar] [CrossRef]
- Zheng, W.; Wang, Y.; Wang, L.; Ma, Z.; Zhao, J.; Wang, P.; Zhang, L.; Liu, Z.; Lu, X. Genetic Mapping and Molecular Marker Development for Pi65(t), a Novel Broad-Spectrum Resistance Gene to Rice Blast Using next-Generation Sequencing. Theor. Appl. Genet. 2016, 129, 1035–1044. [Google Scholar] [CrossRef]
- Zhu, D.; Kang, H.; Li, Z.; Liu, M.; Zhu, X.; Wang, Y.; Wang, D.; Wang, Z.; Liu, W.; Wang, G.L. A Genome-Wide Association Study of Field Resistance to Magnaporthe oryzae in Rice. Rice 2016, 9, 44. [Google Scholar] [CrossRef] [PubMed]
- Joshi, R.K.; Nayak, S. Functional characterization and signal transduction ability of nucleotide-binding site-leucine-rich repeat resistance genes in plants. Genet. Mol. Res. 2011, 10, 2637–2652. [Google Scholar] [CrossRef]
- Wang, B.H.; Ebbole, D.J.; Wang, Z.H. The arms race between Magnaporthe oryzae and rice: Diversity and interaction of Avr and R genes. J. Integr. Agric. 2017, 16, 2746–2760. [Google Scholar] [CrossRef]
- Khang, C.H.; Park, S.Y.; Lee, Y.H.; Valent, B.; Kang, S. Genome organization and evolution of the AVR-Pita avirulence gene family in the Magnaporthe grisea species complex. Mol. Plant Microbe Interact. 2008, 21, 658–670. [Google Scholar] [CrossRef] [PubMed]
- Bryan, G.T.; Wu, K.S.; Farrall, L.; Jia, Y.; Hershey, H.P.; McAdams, S.A.; Faulk, K.N.; Donaldson, G.K.; Tarchini, R.; Valent, B. A Single Amino Acid Difference Distinguishes Resistant and Susceptible Alleles of the Rice Blast Resistance Gene Pi-Ta. Plant Cell 2000, 12, 2033–2045. [Google Scholar] [CrossRef] [PubMed]
- Han, J.; Wang, X.; Wang, F.; Zhao, Z.; Li, G.; Zhu, X.; Chen, L. The fungal effector Avr-pita suppresses innate immunity by increasing COX activity in rice mitochondria. Rice 2021, 14, 12. [Google Scholar] [CrossRef] [PubMed]
- Sharma, T.R.; Madhav, M.S.; Singh, B.K.; Shanker, P.; Jana, T.K.; Dalal, V.; Pandit, A.; Singh, A.; Gaikwad, K.; Upreti, H.C.; et al. High-Resolution Mapping, Cloning and Molecular Characterization of the Pi-k h Gene of Rice, Which Confers Resistance to Magnaporthe grisea. Mol. Genet. Genomics 2005, 274, 569–578. [Google Scholar] [CrossRef] [PubMed]
- Sharma, T.R.; Rai, A.K.; Gupta, S.K.; Singh, N.K. Broad-Spectrum Blast Resistance Gene Pi-k h Cloned from Rice Line Tetep Designated as Pi54. J. Plant Biochem. Biotechnol. 2010, 19, 87–89. [Google Scholar] [CrossRef]
- Ray, S.; Singh, P.K.; Gupta, D.K.; Mahato, A.K.; Sarkar, C.; Rathour, R.; Singh, N.K.; Sharma, T.R. Analysis of Magnaporthe oryzae Genome Reveals a Fungal Effector, Which Is Able to Induce Resistance Response in Transgenic Rice Line Containing Resistance Gene, Pi54. Front. Plant Sci. 2016, 7, 1140. [Google Scholar] [CrossRef] [PubMed]
- Gupta, S.K.; Rai, A.K.; Kanwar, S.S.; Chand, D.; Singh, N.K.; Sharma, T.R. The single functional blast resistance gene Pi54 activates a complex defence mechanism in rice. J. Exp. Bot. 2012, 63, 757–772. [Google Scholar] [CrossRef]
- Maidment, J.H.R.; Franceschetti, M.; Maqbool, A.; Saitoh, H.; Jantasuriyarat, C.; Kamoun, S.; Terauchi, R.; Banfield, M.J. Multiple Variants of the Fungal Effector AVR-Pik Bind the HMA Domain of the Rice Protein OsHIPP19, Providing a Foundation to Engineer Plant Defense. J. Biol. Chem. 2021, 296, 100371. [Google Scholar] [CrossRef]
- Kanzaki, H.; Yoshida, K.; Saitoh, H.; Fujisaki, K.; Hirabuchi, A.; Alaux, L.; Terauchi, R. Arms race co-evolution of Magnaporthe oryzae AVR-Pik and rice Pik genes driven by their physical interactions. Plant J. 2012, 72, 894–907. [Google Scholar] [CrossRef]
- Zhai, C.; Zhang, Y.; Yao, N.; Lin, F.; Liu, Z.; Dong, Z.; Wang, L.; Pan, Q. Function and Interaction of the Coupled Genes Responsible for Pik-h Encoded Rice Blast Resistance. PLoS ONE 2014, 9, e98067. [Google Scholar] [CrossRef] [PubMed]
- Okuyama, Y.; Kanzaki, H.; Abe, A.; Yoshida, K.; Tamiru, M.; Saitoh, H.; Terauchi, R. A multifaceted genomics approach allows the isolation of the rice Pia-blast resistance gene consisting of two adjacent NBS-LRR protein genes. Plant J. 2011, 66, 467–479. [Google Scholar] [CrossRef]
- Cesari, S.; Bernoux, M.; Moncuquet, P.; Kroj, T.; Dodds, P.N. A novel conserved mechanism for plant NLR protein pairs: The ‘integrated decoy’ hypothesis. Front. Plant Sci. 2014, 5, 606. [Google Scholar] [CrossRef]
- Miki, S.; Matsui, K.; Kito, H.; Otsuka, K.; Ashizawa, T.; Yasuda, N.; Sone, T. Molecular cloning and characterization of the AVR-Pia locus from a Japanese field isolate of Magnaporthe oryzae. Mol. Plant Pathol. 2009, 10, 361–374. [Google Scholar] [CrossRef] [PubMed]
- Singh, R.; Dangol, S.; Chen, Y.; Choi, J.; Cho, Y.S.; Lee, J.E.; Jwa, N.S. Magnaporthe oryzae effector AVR-Pii helps to establish compatibility by inhibition of the rice NADP-malic enzyme resulting in disruption of oxidative burst and host innate immunity. Mol.Cells 2016, 39, 426. [Google Scholar]
- Takagi, H.; Uemura, A.; Yaegashi, H.; Tamiru, M.; Abe, A.; Mitsuoka, C.; Terauchi, R. MutMap-Gap: Whole-genome resequencing of mutant F2 progeny bulk combined with de novo assembly of gap regions identifies the rice blast resistance gene Pii. New Phytol. 2013, 200, 276–283. [Google Scholar] [CrossRef] [PubMed]
- Fujisaki, K.; Abe, Y.; Ito, A.; Saitoh, H.; Yoshida, K.; Kanzaki, H.; Kanzaki, E.; Utsushi, H.; Yamashita, T.; Kamoun, S.; et al. Rice Exo70 Interacts with a Fungal Effector, AVR-Pii, and Is Required for AVR-Pii-triggered Immunity. Plant J. 2015, 83, 875–887. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Wu, J.; Kim, S.G.; Tsuda, K.; Gupta, R.; Park, S.Y.; Kim, S.T.; Kang, K.Y. Magnaporthe oryzae -Secreted Protein MSP1 Induces Cell Death and Elicits Defense Responses in Rice. Mol. Plant-Microbe Interact. 2016, 29, 299–312. [Google Scholar] [CrossRef] [PubMed]
- Tang, M.; Ning, Y.; Shu, X.; Dong, B.; Zhang, H.; Wu, D.; Wang, H.; Wang, G.-L.; Zhou, B. The Nup98 Homolog APIP12 Targeted by the Effector AvrPiz-t Is Involved in Rice Basal Resistance Against Magnaporthe oryzae. Rice 2017, 10, 5. [Google Scholar] [CrossRef]
- Park, C.H.; Shirsekar, G.; Bellizzi, M.; Chen, S.; Songkumarn, P.; Xie, X.; Wang, G.L. The E3 ligase APIP10 connects the effector AvrPiz-t to the NLR receptor Piz-t in rice. PLoS Pathog. 2016, 12, e1005529. [Google Scholar] [CrossRef]
- Gupta, D.R.; Surovy, M.Z.; Mahmud, N.U.; Chakraborty, M.; Paul, S.K.; Hossain, M.S.; Bhattacharjee, P.; Mehbub, M.S.; Rani, K.; Yeasmin, R.; et al. Suitable methods for isolation, culture, storage and identification of wheat blast fungus Magnaporthe oryzae Triticum pathotype. Phytopathol. Res. 2020, 2, 30. [Google Scholar] [CrossRef]
- Jamaloddin, M.; Mahender, A.; Gokulan, G.C.; Balachiranjeevi, C.; Maliha, A.; Patel, H.K.; Ali, J. Molecular approaches for disease resistance in rice. In Rice Improvement: Physiological, Molecular Breeding and Genetic Perspectives; Ali, J., Wani, S.H., Eds.; Springer: Berlin/Heidelberg, Germany, 2021; pp. 315–378. [Google Scholar]
- Qin, P.; Hu, X.; Jiang, N.; Bai, Z.; Liu, T.; Fu, C.; Song, Y.; Wang, K.; Yang, Y. A Procedure for Inducing the Occurrence of Rice Seedling Blast in Paddy Field. Plant Pathol. J. 2021, 37, 200–203. [Google Scholar] [CrossRef] [PubMed]
- Jamaloddin, M.; Durga Rani, C.V.; Swathi, G.; Anuradha, C.; Vanisri, S.; Rajan, C.P.D.; Raju, S.K.; Bhuvaneshwari, V.; Jagadeeswar, R.; Laha, G.S.; et al. Marker assisted gene pyramiding (MAGP) for bacterial blight and blast resistance into mega rice variety “Tellahamsa”. PLoS ONE 2020, 15, e0234088. [Google Scholar] [CrossRef] [PubMed]
- Vasudevan, K.; Vera Cruz, C.M.; Gruissem, W.; Bhullar, N.K. Large Scale Germplasm Screening for Identification of Novel Rice Blast Resistance Sources. Front. Plant Sci. 2014, 5, 505. [Google Scholar] [CrossRef] [PubMed]
- Srinivas Prasad, M.; Sheshu Madhav, M.; Laha, G.S.; Ladha Lalshmi, D.; Krishnaveni, D.; Mangrauthia, S.K.; Balachandran, S.M.; Sundaram, R.M.; Arunakranthi, B.; Madhan Mohan, K.; et al. Rice Blast Disease and Its Management; Technical Bulletin No. 57; Directorate of Rice Research (ICAR): Hyderabad, India, 2011; Volume 52. [Google Scholar]
- Challagulla, V.; Bhattarai, S.; Midmore, D.J. In-vitro vs in-vivo inoculation: Screening for resistance of australian rice genotypes against blast fungus. Rice Sci. 2015, 22, 132–137. [Google Scholar] [CrossRef]
- IRRI. Standard Evaluation System (SES) 2002 for Rice; International Rice Research Institute (IRRI): Los Baños, Philippine, 2002; pp. 1–45. [Google Scholar]
- Prabhu, A.S.; Filippi, M.C.; Castro, N. Pathogenic variation among isolates of Pyricularia Oryzae affecting rice, wheat, and grasses in Brazil. Int. J. Pest Manag. 1992, 38, 367–371. [Google Scholar]
- Suprun, I.I.; Kovalyev, V.S.; Kharchenko, E.S.; Savenko, E.G. Assessment of breeding lines of rice (Oryza sativa L.) carrying the Pi-40 gene for resistance to rice blast strains from the Krasnodar Region. Vavilov J. Genet. Breed. 2016, 20, 333–336. [Google Scholar] [CrossRef]
- Kim, J.S.; Ahn, S.N.; Kim, C.K.; Shim, C.K. Screening of rice blast resistance genes from aromatic rice germplasms with SNP markers. Plant Pathol. J. 2010, 26, 70–79. [Google Scholar] [CrossRef]
- Kim, J.S.; Ahn, S.N.; Hong, S.J.; Kwon, J.H.; Kim, Y.K.; Jee, H.J.; Shim, C.K. Screening of the Dominant Rice Blast Resistance Genes with PCR-based SNP and CAPS Marker in Aromatic Rice Germplasm. Korean J. Crop Sci. 2011, 56, 329–341. [Google Scholar] [CrossRef]
- Singh, A.K.; Singh, P.K.; Arya, M.; Singh, N.K.; Singh, U.S. Molecular Screening of Blast Resistance Genes in Rice using SSR Markers. Plant Pathol. J. 2015, 31, 12–24. [Google Scholar] [CrossRef] [PubMed]
- Imam, J.; Alam, S.; Mandal, N.P.; Variar, M.; Shukla, P. Molecular screening for identification of blast resistance genes in North East and Eastern Indian rice germplasm (Oryza sativa L.) with PCR based makers. Euphytica 2014, 196, 199–211. [Google Scholar] [CrossRef]
- Shikari, A.B.; Khanna, A.; Krishnan, S.G.; Singh, U.D.; Rathour, R.; Tonapi, V.; Sharma, T.R.; Nagarajan, M.; Prabhu, K.V.; Singh, A.K. Molecular Analysis and Phenotypic Validation of Blast Resistance Genes Pita and Pita 2 in Landraces of Rice (Oryza sativa L.). Indian J. Genet. Plant Breed. 2013, 73, 131. [Google Scholar] [CrossRef]
- Yan, L.; Bai-Yuan, Y.; Yun-Liang, P.; Zhi-Juan, J.; Yu-Xiang, Z.; Han-Lin, W.; Chang-Deng, Y. Molecular Scree Ning of Blast Resistance Genes in Rice Germplasms Resistant to Magnaporthe oryzae. Rice Sci. 2017, 24, 41–47. [Google Scholar] [CrossRef]
- Teerasan, W.; Moonsap, P.; Longya, A.; Damchuay, K.; Ito, S.; Tasanasuwan, P.; Kate-Ngam, S.; Jantasuriyarat, C. Rice Blast Resistance Gene Profiling of Thai, Japanese and International Rice Varieties Using Gene-Specific Markers. Plant Genet. Resour. Charact. Util. 2022, 20, 22–28. [Google Scholar] [CrossRef]
- Imbe, T.; Tsunematsu, H.; Kato, H.; Khush, G.S. Genetic Analysis of Blast Resistance in IR Varieties and Resistant Breeding Strategy. In Advances in Rice Blast Research; Springer: Dordrecht, The Netherlands, 2000; pp. 1–8. [Google Scholar]
- Ramalingam, J.; Vera Cruz, C.M.; Kukreja, K.; Chittoor, J.M.; Wu, J.-L.; Lee, S.W.; Baraoidan, M.; George, M.L.; Cohen, M.B.; Hulbert, S.H.; et al. Candidate Defense Genes from Rice, Barley, and Maize and Their Association with Qualitative and Quantitative Resistance in Rice. Mol. Plant-Microbe Interact. 2003, 16, 14–24. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Variar, M.; Cruz, C.M.V.; Carrillo, M.G.; Bhatt, J.C.; Sangar, R.B.S. Rice Blast in India and Strategies to Develop Durably Resistant Cultivars. In Advances in Genetics, Genomics and Control of Rice Blast Disease; Springer: Dordrecht, The Netherlands, 2009; pp. 359–373. [Google Scholar]
- Suwarno; Lubis, E.; Hairmansis, A. Santoso Development of a Package of 20 Varieties for Blast Management on Upland Rice. In Advances in Genetics, Genomics and Control of Rice Blast Disease; Springer: Dordrecht, The Netherlands, 2009; pp. 347–357. [Google Scholar]
- Shindo, K.; Horino, O. Control of rice blast disease by mixed plantings of isogenic lines as multiline cultivars. Bull. Tohoku Natl. Agric. Exp. Stn. 1989, 39, 55–96. [Google Scholar]
- Ise, K. Effect of mixing planting of near isogenic lines of ‘Nipponbare’ rice to reduce blast disease. Breed Sci. 1990, 40, 288. [Google Scholar]
- Koizumi, S.; Fuji, S. Effect of mixtures of isogenic lines developed from rice cv. ‘Sasanishiki and Nipponbare’ on blast development. Res. Bull. Aichi Agric. Res. Ctr. 1994, 26, 87–97. [Google Scholar]
- Koizumi, S. Effect of field resistance on leaf blast development in mixtures of susceptible and resistant rice cultivars. Ann. Phytopathol. Soc. Jpn. 1996, 60, 585–594. [Google Scholar] [CrossRef]
- Nakajima, T.; Sonoda, R.; Yaegashi, H.; Saito, H. Factors related to suppression of leaf blast disease with a multiline ofrice cultivar Sasanishiki and its isogenic lines. Ann. Phytopathol. Soc. Jpn. 1996, 62, 360–364. [Google Scholar] [CrossRef][Green Version]
- Abe, S. Breeding of a Blast Resistant Multiline Variety of Rice, Sasanishiki BL. Jpn. Agric. Res. Q. JARQ 2004, 38, 149–154. [Google Scholar] [CrossRef]
- Zhu, Y.; Chen, H.; Fan, J.; Wang, Y.; Li, Y.; Chen, J.; Fan, J.; Yang, S.; Hu, L.; Leung, H.; et al. Genetic Diversity and Disease Control in Rice. Nature 2000, 406, 718–722. [Google Scholar] [CrossRef] [PubMed]
- Persaud, R.; McGowan, D.; Persaud, M. Managing the Imminent Danger of Rice Blast (Pyricularia oryzae Cav.) and Sheath Blight (Rhizoctonia solani Kuhn) Disease: A Critical Review Article. Am. J. Agril. Forestry 2021, 9, 409–423. [Google Scholar] [CrossRef]
- Ahn, S.W. International Collaboration on Breeding for Resistance to Rice Blast. In Rice Blast Disease; Zeigler, R.S., Leong, S.A., Teng, P.S., Eds.; CAB International: Wallingford, UK, 1994; pp. 137–153. [Google Scholar]
- Martínez, C.P.; Victoria, F.C.; Amézquita, M.C.; Tulande, E.; Lema, G.; Zeigler, R.S. Comparison of rice lines derived through anther culture and the pedigree method in relation to blast (Pyricularia grisea Sacc.) resistance. Theor. Appl. Genet. 1996, 92, 583–590. [Google Scholar] [CrossRef]
- Allard, R.W. Principles of Plant Breeding; John Wiley: New York, NY, USA, 1999; ISBN 9780471023098. [Google Scholar]
- Korinsak, S.; Sirithunya, P.; Meakwatanakarn, P.; Sarkarung, S.; Vanavichit, A.; Toojinda, T. Changing Allele Frequencies Associated with Specific Resistance Genes to Leaf Blast in Backcross Introgression Lines of Khao Dawk Mali 105 Developed from a Conventional Selection Program. Field Crops Res. 2011, 122, 32–39. [Google Scholar] [CrossRef]
- Fujimski, H. Recurrent selection by using male sterility for rice improvement. Jpn. Agric. Res. Quart. 1979, 13, 153–156. [Google Scholar]
- Guimarães, E.P.; Correa-Victoria, F.J. Utilización de la selección recurrente para desarrollar resistencia a Pyricularia grisea Sacc. en arroz. In Selección Recurrente en Arroz; Guimarães, E.P., Ed.; CIAT: Cali, Colombia, 1997; pp. 165–175. [Google Scholar]
- Hernandez-Soto, A.; Echeverría-Beirute, F.; Abdelnour-Esquivel, A.; Valdez-Melara, M.; Boch, J.; Gatica-Arias, A. Rice Breeding in the New Era: Comparison of Useful Agronomic Traits. Curr. Plant Biol. 2021, 27, 100211. [Google Scholar] [CrossRef]
- FAO/IAEA MVD. 2022. Available online: https://nucleus.iaea.org/sites/mvd/SitePages/Home.aspx (accessed on 25 June 2022).
- Anonymous. The Gazette of India (S.O. 500(E) Dated 02.02.2021); Ministry of Agriculture and Farmers Welfare, Department of Agriculture, Cooperation and Farmers Welfare, Government of India: New Delhi, India, 2021.
- Sahu, P.K.; Sao, R.; Mondal, S.; Vishwakarma, G.; Gupta, S.K.; Kumar, V.; Singh, S.; Sharma, D.; Das, B.K. Next Generation Sequencing Based Forward Genetic Approaches for Identification and Mapping of Causal Mutations in Crop Plants: A Comprehensive Review. Plants 2020, 9, 1355. [Google Scholar] [CrossRef]
- Li, C.; Wang, D.; Peng, S.; Chen, Y.; Su, P.; Chen, J.; Zheng, L.; Tan, X.; Liu, J.; Xiao, Y.; et al. Genome-wide association mapping of resistance against rice blast strains in South China and identification of a new Pik allele. Rice 2019, 12, 47. [Google Scholar] [CrossRef]
- Sasaki, R. Existence of strains in rice blast fungus. Int. J. Plant Prot. Tokyo 1922, 9, 631–644. [Google Scholar]
- Kiyosawa, S. Genetic studies on host-pathogen relationship in the rice blast disease. Tokyo Agric. Forest Fish Res. Counc. Trop. Agric. Res. Ser. 1967, 1, 137–153. [Google Scholar]
- Ballini, E.; Morel, J.-B.; Droc, G.; Price, A.; Courtois, B.; Notteghem, J.-L.; Tharreau, D. A Genome-Wide Meta-Analysis of Rice Blast Resistance Genes and Quantitative Trait Loci Provides New Insights into Partial and Complete Resistance. Mol. Plant-Microbe Interact. 2008, 21, 859–868. [Google Scholar] [CrossRef] [PubMed]
- Zhai, C.; Lin, F.; Dong, Z.; He, X.; Yuan, B.; Zeng, X.; Wang, L.; Pan, Q. The Isolation and Characterization of Pik, a Rice Blast Resistance Gene Which Emerged after Rice Domestication. New Phytol. 2011, 189, 321–334. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.D.; Liu, J.L.; Leach, J.E.; Wang, G.L. Novel insights into rice innate immunity against bacterial and fungal pathogens. Annu. Rev. Phytopathol. 2014, 52, 213–241. [Google Scholar] [CrossRef] [PubMed]
- Jiang, J.; Yang, D.; Ali, J.; Mou, T. Molecular marker-assisted pyramiding of broad-spectrum disease resistance genes, Pi2 and Xa23, into GZ63-4S, an elite thermo-sensitive genic male-sterile line in rice. Mol. Plant Breed. 2015, 35, 83–96. [Google Scholar] [CrossRef]
- Khanna, A.; Sharma, V.; Ellur, R.K.; Shikari, A.B.; Gopala Krishnan, S.; Singh, U.D.; Prakash, G.; Sharma, T.R.; Rathour, R.; Variar, M.; et al. Development and Evaluation of Near-Isogenic Lines for Major Blast Resistance Gene(s) in Basmati Rice. Theor. Appl. Genet. 2015, 128, 1243–1259. [Google Scholar] [CrossRef] [PubMed]
- Tian, H.; Cheng, H.; Hu, J.; Lei, C.; Zhu, X.; Qian, Q. Effect of introgressed Pigm gene on rice blast resistance and yield traits of Japonica rice in cold area. J Shenyang Agric. Uni. 2016, 47, 520–526. [Google Scholar]
- Liu, S.P.; Li, X.; Wang, C.Y.; Li, X.H.; He, Y.Q. Improvement of resistance to rice blast in Zhenshan 97 by molecular marker aided selection. J. Integr. Plant Biol. 2002, 45, 1346–1350. [Google Scholar]
- Joshi, S.; Dhatwalia, S.; Kaachra, A.; Sharma, K.D.; Rathour, R. Genetic and Physical Mapping of a New Rice Blast Resistance Specificity Pi-67 from a Broad Spectrum Resistant Genotype Tetep. Euphytica 2019, 215. [Google Scholar] [CrossRef]
- Dong, L.; Liu, S.; Xu, P.; Deng, W.; Li, X.; Tharreau, D.; Li, J.; Zhou, J.; Wang, Q.; Tao, D.; et al. Fine Mapping of Pi57(t) Conferring Broad Spectrum Resistance against Magnaporthe oryzae in Introgression Line IL-E1454 Derived from Oryza Longistaminata. PLoS ONE 2017, 12, e0186201. [Google Scholar] [CrossRef]
- Wang, R.; Fang, N.; Guan, C.; He, W.; Bao, Y.; Zhang, H. Characterization and Fine Mapping of a Blast Resistant Gene Pi-Jnw1 from the Japonica Rice Landrace Jiangnanwan. PLoS ONE 2016, 11, e0169417. [Google Scholar] [CrossRef] [PubMed]
- Liang, Z.; Wang, L.; Pan, Q. A New Recessive Gene Conferring Resistance against Rice Blast. Rice 2016, 9, 47. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Peng, P.; Tian, J.; He, Y.; Zhang, L.; Liu, Z.; Yin, D.; Zhang, Z. Pike, a rice blast resistance allele consisting of two adjacent NBS–LRR genes, was identified as a novel allele at the Pik Locus. Mol. Breed. 2015, 35, 117. [Google Scholar] [CrossRef]
- Xiao, W.; Yang, Q.; Sun, D.; Wang, H.; Guo, T.; Liu, Y.; Zhu, X.; Chen, Z. Identification of Three Major R Genes Responsible for Broad-Spectrum Blast Resistance in an Indica Rice Accession. Mol. Breed. 2015, 35, 49. [Google Scholar] [CrossRef]
- Ma, J.; Lei, C.; Xu, X.; Hao, K.; Wang, J.; Cheng, Z.; Ma, X.; Ma, J.; Zhou, K.; Zhang, X.; et al. Pi64, Encoding a Novel CC-NBS-LRR Protein, Confers Resistance to Leaf and Neck Blast in Rice. Mol. Plant-Microbe Interact. 2015, 28, 558–568. [Google Scholar] [CrossRef]
- Sun, P.; Liu, J.; Wang, Y.; Jiang, N.; Wang, S.; Dai, Y.; Gao, J.; Li, Z.; Pan, S.; Wang, D.; et al. Molecular Mapping of the Blast Resistance Gene Pi49 in the Durably Resistant Rice Cultivar Mowanggu. Euphytica 2012, 192, 45–54. [Google Scholar] [CrossRef]
- Lei, C.; Hao, K.; Yang, Y.; Ma, J.; Wang, S.; Wang, J.; Cheng, Z.; Zhao, S.; Zhang, X.; Guo, X.; et al. Identification and Fine Mapping of Two Blast Resistance Genes in Rice Cultivar 93-11. Crop J. 2013, 1, 2–14. [Google Scholar] [CrossRef]
- Koide, Y.; Telebanco-Yanoria, M.J.; Fukuta, Y.; Kobayashi, N. Detection of Novel Blast Resistance Genes, Pi58(t) and Pi59(t), in a Myanmar Rice Landrace Based on a Standard Differential System. Mol. Breed. 2013, 32, 241–252. [Google Scholar] [CrossRef]
- Xiao, W.; Yang, Q.; Wang, H.; Duan, J.; Guo, T.; Liu, Y.; Zhu, X.; Chen, Z. Identification and Fine Mapping of a Major R Gene to Magnaporthe oryzae in a Broad-Spectrum Resistant Germplasm in Rice. Mol. Breed. 2012, 30, 1715–1726. [Google Scholar] [CrossRef]
- Zhu, X.; Chen, S.; Yang, J.; Zhou, S.; Zeng, L.; Han, J.; Su, J.; Wang, L.; Pan, Q. The Identification of Pi50(t), a New Member of the Rice Blast Resistance Pi2/Pi9 Multigene Family. Theor. Appl. Genet. 2012, 124, 1295–1304. [Google Scholar] [CrossRef]
- Jiang, N.; Li, Z.; Wu, J.; Wang, Y.; Wu, L.; Wang, S.; Wang, D.; Wen, T.; Liang, Y.; Sun, P.; et al. Molecular Mapping of the Pi2/9 Allelic Gene Pi2-2 Conferring Broad-Spectrum Resistance to Magnaporthe oryzae in the Rice Cultivar Jefferson. Rice 2012, 5, 29. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Bao, Y.; Xie, L.; Su, Y.; Chu, R.; He, W.; Huang, J.; Wang, J.; Zhang, H. Fine Mapping and Identification of Blast Resistance Gene Pi-Hk1 in a Broad-Spectrum Resistant Japonica Rice Landrace. Phytopathology 2013, 103, 1162–1168. [Google Scholar] [CrossRef]
- Liu, Y.; Liu, B.; Zhu, X.; Yang, J.; Bordeos, A.; Wang, G.; Leach, J.E.; Leung, H. Fine-Mapping and Molecular Marker Development for Pi56(t), a NBS-LRR Gene Conferring Broad-Spectrum Resistance to Magnaporthe oryzae in Rice. Theor. Appl. Genet. 2012, 126, 985–998. [Google Scholar] [CrossRef] [PubMed]
- He, X.; Liu, X.; Wang, L.; Wang, L.; Lin, F.; Cheng, Y.; Chen, Z.; Liao, Y.; Pan, Q. Identification of the Novel Recessive Gene Pi55(t) Conferring Resistance to Magnaporthe oryzae. Sci. China Life Sci. 2012, 55, 141–149. [Google Scholar] [CrossRef][Green Version]
- Endo, T.; Yamaguchi, M.; Kaji, R.; Nakagomi, K.; Kataoka, T.; Yokogami, N.; Nakamura, T.; Ishikawa, G.; Yonemaru, J.; Nishio, T. Close Linkage of a Blast Resistance Gene, Pias(t), with a Bacterial Leaf Blight Resistance Gene, Xa1-as(t), in a Rice Cultivar ‘Asominori’. Breed. Sci. 2012, 62, 334–339. [Google Scholar] [CrossRef]
- Wang, Y.; Wang, D.; Deng, X.; Liu, J.; Sun, P.; Liu, Y.; Huang, H.; Jiang, N.; Kang, H.; Ning, Y.; et al. Molecular Mapping of the Blast Resistance Genes Pi2-1 and Pi51(t) in the Durably Resistant Rice ‘Tianjingyeshengdao’. Phytopathology 2012, 102, 779–786. [Google Scholar] [CrossRef]
- Xiao, W.; Yang, Q.; Wang, H.; Guo, T.; Liu, Y.; Zhu, X.; Chen, Z. Identification and Fine Mapping of a Resistance Gene to Magnaporthe oryzae in a Space-Induced Rice Mutant. Mol. Breed. 2010, 28, 303–312. [Google Scholar] [CrossRef]
- Huang, H.; Huang, L.; Feng, G.; Wang, S.; Wang, Y.; Liu, J.; Jiang, N.; Yan, W.; Xu, L.; Sun, P.; et al. Molecular Mapping of the New Blast Resistance Genes Pi47 and Pi48 in the Durably Resistant Local Rice Cultivar Xiangzi 3150. Phytopathology 2011, 101, 620–626. [Google Scholar] [CrossRef]
- Zeng, X.; Yang, X.; Zhao, Z.; Lin, F.; Wang, L.; Pan, Q. Characterization and Fine Mapping of the Rice Blast Resistance Gene Pia. Sci. China Life Sci. 2011, 54, 372–378. [Google Scholar] [CrossRef][Green Version]
- Kumar, P.; Pathania, S.; Katoch, P.; Sharma, T.R.; Plaha, P.; Rathour, R. Genetic and Physical Mapping of Blast Resistance Gene Pi-42(t) on the Short Arm of Rice Chromosome 12. Mol. Breed. 2009, 25, 217–228. [Google Scholar] [CrossRef]
- Lee, S.; Wamishe, Y.; Jia, Y.; Liu, G.; Jia, M.H. Identification of Two Major Resistance Genes against Race IE-1k of Magnaporthe oryzae in the Indica Rice Cultivar Zhe733. Mol. Breed. 2009, 24, 127–134. [Google Scholar] [CrossRef]
- Yang, Q.; Lin, F.; Wang, L.; Pan, Q. Identification and Mapping of Pi41, a Major Gene Conferring Resistance to Rice Blast in the Oryza sativa Subsp. Indica Reference Cultivar, 93-11. Theor. Appl. Genet. 2009, 118, 1027–1034. [Google Scholar] [CrossRef] [PubMed]
- Shang, J.; Tao, Y.; Chen, X.; Zou, Y.; Lei, C.; Wang, J.; Li, X.; Zhao, X.; Zhang, M.; Lu, Z.; et al. Identification of a New Rice Blast Resistance Gene, Pid3, by Genome wide Comparison of Paired Nucleotide-Binding Site–Leucine-Rich Repeat Genes and Their Pseudogene Alleles Between the Two Sequenced Rice Genomes. Genetics 2009, 182, 1303–1311. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Xu, X.; Lin, F.; Pan, Q. Characterization of Rice Blast Resistance Genes in the Pik Cluster and Fine Mapping of the Pik-p Locus. Phytopathology 2009, 99, 900–905. [Google Scholar] [CrossRef]
- Xu, X.; Chen, H.; Fujimura, T.; Kawasaki, S. Fine Mapping of a Strong QTL of Field Resistance against Rice Blast, Pikahei-1(t), from Upland Rice Kahei, Utilizing a Novel Resistance Evaluation System in the Greenhouse. Theor. Appl. Genet. 2008, 117, 997–1008. [Google Scholar] [CrossRef]
- Xu, X.; Hayashi, N.; Wang, C.T.; Kato, H.; Fujimura, T.; Kawasaki, S. Efficient Authentic Fine Mapping of the Rice Blast Resistance Gene Pik-h in the Pik Cluster, Using New Pik-h-Differentiating Isolates. Mol. Breed. 2008, 22, 289–299. [Google Scholar] [CrossRef]
- Dwinita, W.U.; Sugiono, M.; Hajrial, A.; Asep, S.; Ida, H. Blast resistance genes in wild rice Oryza rufipogon and rice cultivar IR64. Ind J Agric. 2008, 1, 71–76. [Google Scholar]
- Liu, X.; Yang, Q.; Lin, F.; Hua, L.; Wang, C.; Wang, L.; Pan, Q. Identification and Fine Mapping of Pi39(t), a Major Gene Conferring the Broad-Spectrum Resistance to Magnaporthe oryzae. Mol. Genet. Genom. 2007, 278, 403–410. [Google Scholar] [CrossRef]
- Zenbayashi-Sawata, K.; Fukuoka, S.; Katagiri, S.; Fujisawa, M.; Matsumoto, T.; Ashizawa, T.; Koizumi, S. Genetic and Physical Mapping of the Partial Resistance Gene, Pi34, to Blast in Rice. Phytopathology 2007, 97, 598–602. [Google Scholar] [CrossRef]
- Jeung, J.U.; Kim, B.R.; Cho, Y.C.; Han, S.S.; Moon, H.P.; Lee, Y.T.; Jena, K.K. A Novel Gene, Pi40(t), Linked to the DNA Markers Derived from NBS-LRR Motifs Confers Broad Spectrum of Blast Resistance in Rice. Theor. Appl. Genet. 2007, 115, 1163–1177. [Google Scholar] [CrossRef]
- Deng, Y.; Zhu, X.; Shen, Y.; He, Z. Genetic Characterization and Fine Mapping of the Blast Resistance Locus Pigm(t) Tightly Linked to Pi2 and Pi9 in a Broad-Spectrum Resistant Chinese Variety. Theor. Appl. Genet. 2006, 113, 705–713. [Google Scholar] [CrossRef] [PubMed]
- Qu, S.; Liu, G.; Zhou, B.; Bellizzi, M.; Zeng, L.; Dai, L.; Han, B.; Wang, G.-L. The Broad-Spectrum Blast Resistance Gene Pi9 Encodes a Nucleotide-Binding Site–Leucine-Rich Repeat Protein and Is a Member of a Multigene Family in Rice. Genetics 2006, 172, 1901–1914. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Shang, J.; Chen, D.; Lei, C.; Zou, Y.; Zhai, W.; Liu, G.; Xu, J.; Ling, Z.; Cao, G.; et al. A B-Lectin Receptor Kinase Gene Conferring Rice Blast Resistance. Plant J. 2006, 46, 794–804. [Google Scholar] [CrossRef]
- Nguyen, T.T.T.; Koizumi, S.; La, T.N.; Zenbayashi, K.S.; Ashizawa, T.; Yasuda, N.; Imazaki, I.; Miyasaka, A. Pi35(t), a New Gene Conferring Partial Resistance to Leaf Blast in the Rice Cultivar Hokkai 188. Theor. Appl. Genet. 2006, 113, 697–704. [Google Scholar] [CrossRef] [PubMed]
- Gowda, M.; Roy-Barman, S.; Chattoo, B.B. Molecular Mapping of a Novel Blast Resistance Gene Pi38 in Rice Using SSLP and AFLP Markers. Plant Breed. 2006, 125, 596–599. [Google Scholar] [CrossRef]
- Hayashi, K.; Yoshida, H.; Ashikawa, I. Development of PCR-Based Allele-Specific and InDel Marker Sets for Nine Rice Blast Resistance Genes. Theor. Appl. Genet. 2006, 113, 251–260. [Google Scholar] [CrossRef] [PubMed]
- Zhou, B.; Qu, S.; Liu, G.; Dolan, M.; Sakai, H.; Lu, G.; Bellizzi, M.; Wang, G.L. The Eight Amino-Acid Differences Within Three Leucine-Rich Repeats Between Pi2 and Piz-t Resistance Proteins Determine the Resistance Specificity to Magnaporthe grisea. Mol. Plant-Microbe Interact. 2006, 19, 1216–1228. [Google Scholar] [CrossRef]
- Lei, C.L.; Huang, D.Y.; Li, W.; Wang, L.J.; Liu, Z.L.; Wang, X.T.; Shi, K.; Cheng, Z.J.; Zhang, X.; Ling, Z.Z.; et al. Molecular mapping of a blast resistance gene in an indica rice cultivar Yanxian No. 1. Rice Genet. Newsl. 2005, 22, 76–77. [Google Scholar]
- Chen, S.; Wang, L.; Que, Z.; Pan, R.; Pan, Q. Genetic and physical mapping of Pi37(t), a new gene conferring resistance to rice blast in the famous cultivar St. No. 1. Theor. Appl. Genet. 2005, 111, 1563–1570. [Google Scholar] [CrossRef]
- Wu, J.L.; Fan, Y.-Y.; Li, D.-B.; Zheng, K.-L.; Leung, H.; Zhuang, J.-Y. Genetic Control of Rice Blast Resistance in the Durably Resistant Cultivar Gumei 2 against Multiple Isolates. Theor. Appl. Genet. 2005, 111, 50–56. [Google Scholar] [CrossRef]
- Liu, X.Q.; Wang, L.; Chen, S.; Lin, F.; Pan, Q.H. Genetic and Physical Mapping of Pi36(t), a Novel Rice Blast Resistance Gene Located on Rice Chromosome 8. Mol. Genet. Genom. 2005, 274, 394–401. [Google Scholar] [CrossRef] [PubMed]
- Liu, B.; Zhang, S.; Zhu, X.; Yang, Q.; Wu, S.; Mei, M.; Mauleon, R.; Leach, J.; Mew, T.; Leung, H. Candidate Defense Genes as Predictors of Quantitative Blast Resistance in Rice. Mol. Plant-Microbe Interact. 2004, 17, 1146–1152. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Chen, X.W.; Li, S.G.; Xu, J.C.; Zhai, W.X.; Ling, Z.Z.; Ma, B.T.; Wang, Y.P.; Wang, W.M.; Cao, G.; Ma, Y.Q.; et al. Identification of two blast resistance genes in a rice variety, Digu. J. Phytopathol. 2004, 152, 77–85. [Google Scholar] [CrossRef]
- Zhou, J.H.; Wang, J.L.; Xu, J.C.; Lei, C.L.; Ling, Z.Z. Identification and Mapping of a Rice Blast Resistance Gene Pi-g(t) in the Cultivar Guangchangzhan. Plant Pathol. 2004, 53, 191–196. [Google Scholar] [CrossRef]
- Fukuta, Y. The Pish blast resistance gene of rice (Oryza sativa L.) is located on the long arm of chromosome 1. Japan International Research Center for Agricultural Sciences (JIRCAS). Ann. Rep. 2004, 16, 46–47. [Google Scholar]
- Zhu, M.; Wang, L.; Pan, Q. Identification and Characterization of a New Blast Resistance Gene Located on Rice Chromosome 1 through Linkage and Differential Analyses. Phytopathology 2004, 94, 515–519. [Google Scholar] [CrossRef]
- Barman, S.R.; Gowda, M.; Venu, R.C.; Chattoo, B.B. Identification of a Major Blast Resistance Gene in the Rice Cultivar “Tetep”. Plant Breed. 2004, 123, 300–302. [Google Scholar] [CrossRef]
- Fjellstrom, R.; Conaway-Bormans, C.A.; McClung, A.M.; Marchetti, M.A.; Shank, A.R.; Park, W.D. Development of DNA Markers Suitable for Marker Assisted Selection of Three Pi Genes Conferring Resistance to Multiple Pyricularia grisea Pathotypes. Crop Sci. 2004, 44, 1790–1798. [Google Scholar] [CrossRef]
- Sallaud, C.; Lorieux, M.; Roumen, E.; Tharreau, D.; Berruyer, R.; Svestasrani, P.; Garsmeur, O.; Ghesquiere, A.; Notteghem, J.L. Identification of Five New Blast Resistance Genes in the Highly Blast-Resistant Rice Variety IR64 Using a QTL Mapping Strategy. Theor. Appl. Genet. 2003, 106, 794–803. [Google Scholar] [CrossRef]
- Jeon, J.S.; Chen, D.; Yi, G.-H.; Wang, G.L.; Ronald, P.C. Genetic and Physical Mapping of Pi5(t), a Locus Associated with Broad-Spectrum Resistance to Rice Blast. Mol. Genet. Genom. 2003, 269, 280–289. [Google Scholar] [CrossRef]
- Berruyer, R.; Adreit, H.; Milazzo, J.; Gaillard, S.; Berger, A.; Dioh, W.; Lebrun, M.H.; Tharreau, D. Identification and Fine Mapping of Pi33, the Rice Resistance Gene Corresponding to the Magnaporthe grisea Avirulence Gene ACE1. Theor. Appl. Genet. 2003, 107, 1139–1147. [Google Scholar] [CrossRef] [PubMed]
- Pan, Q.H.; Hu, Z.D.; Tanisaka, T.; Wang, L. Fine mapping of the blast resistance gene Pi15, linked to Pii, on rice chromosome 9. Acta Bot. Sin. 2003, 45, 871–877. [Google Scholar]
- Kinoshita, T.; Kiyosawa, S. Some considerations on linkage relationships between Pii and Piz in the blast resistance of rice. Rice Genet. Newsl. 1997, 14, 57–59. [Google Scholar]
- Jiang, J.; Wang, S. Identification of a 118-Kb DNA Fragment Containing the Locus of Blast Resistance Gene Pi-2(t) in Rice. Mol. Genet. Genom. 2002, 268, 249–252. [Google Scholar] [CrossRef]
- Zhuang, J.Y.; Wu, J.L.; Fan, Y.Y.; Rao, Z.M.; Zheng, K.L. Genetic drag between a blast resistance gene and QTL conditioning yield trait detected in a recombinant inbred line population in rice. Rice Genet Newsl. 2001, 18, 69–70. [Google Scholar]
- Chauhan, R.; Farman, M.; Zhang, H.B.; Leong, S. Genetic and Physical Mapping of a Rice Blast Resistance Locus, Pi-CO39(t), That Corresponds to the Avirulence Gene AVR1-CO39 of Magnaporthe grisea. Mol. Genet. Genom. 2002, 267, 603–612. [Google Scholar] [CrossRef]
- Fukuoka, S.; Okuno, K. QTL analysis and mapping of pi21, a recessive gene for field resistance to rice blast in Japanese upland rice. Theor. Appl. Genet. 2001, 103, 185–190. [Google Scholar] [CrossRef]
- Ahn, S.N.; Kim, Y.K.; Hong, H.C.; Han, S.S.; Kwon, S.J.; Choi, H.C.; Moon, H.P.; McCouch, S.R. Molecular mapping of a new gene for resistance to rice blast (Pyricularia grisea Sacc.). Euphytica 2000, 116, 17–22. [Google Scholar] [CrossRef]
- Tabien, R.E.; Li, Z.; Paterson, A.H.; Marchetti, M.A.; Stansel, J.W.; Pinson, S.R.M.; Park, W.D. Mapping of Four Major Rice Blast Resistance Genes from ’Lemont’ and ’Teqing’ and Evaluation of Their Combinatorial Effect for Field Resistance. Theor. Appl. Genet. 2000, 101, 1215–1225. [Google Scholar] [CrossRef]
- Tabien, R.; Li, Z.; Paterson, A.; Marchetti, M.; Stansel, J.; Pinson, S. Mapping QTLs for Field Resistance to the Rice Blast Pathogen and Evaluating Their Individual and Combined Utility in Improved Varieties. Theor. Appl. Genet. 2002, 105, 313–324. [Google Scholar] [CrossRef] [PubMed]
- Chen, D.H.; dela Viña, M.; Inukai, T.; Mackill, D.J.; Ronald, P.C.; Nelson, R.J. Molecular Mapping of the Blast Resistance Gene, Pi44(t), in a Line Derived from a Durably Resistant Rice Cultivar. Theor. Appl. Genet. 1999, 98, 1046–1053. [Google Scholar] [CrossRef]
- Wang, P.; Tanisaka, A. New Blast Resistance Gene Identified in the Indian Native Rice Cultivar Aus373 through Allelism and Linkage Tests. Plant Pathol. 1999, 48, 288–293. [Google Scholar] [CrossRef]
- Fujii, K.; Hayano-Saito, Y.; Sugiura, N.; Hayashi, N.; Saka, N.; Tooyama, T.; Izawa, T.; Shumiya, A. Gene Analysis of Panicle Blast Resistance in Rice Cultivars with Rice Stripe Resistance. Breed. Res. 1999, 1, 203–210. [Google Scholar] [CrossRef]
- Hayashi, N.; Inoue, H.; Kato, T.; Funao, T.; Shirota, M.; Shimizu, T.; Kanamori, H.; Yamane, H.; Hayano-Saito, Y.; Matsumoto, T.; et al. Durable Panicle Blast-Resistance Gene Pb1 Encodes an Atypical CC-NBS-LRR Protein and Was Generated by Acquiring a Promoter through Local Genome Duplication. Plant J. 2010, 64, 498–510. [Google Scholar] [CrossRef] [PubMed]
- Zhuang, J.Y.; Lu, J.; Qian, H.R.; Lin, H.X.; Zheng, K.L. Tagging of blast resistance gene(s) to DNA markers and marker-assisted selection (MAS) in rice improvement. In IAEA-TECDOC:1010; Int Atomic Energy Agency (IAEA): Tokyo, Japan, 1998; pp. 55–61. [Google Scholar]
- Hayashi, N.; Ando, I.; Imbe, T. Identification of a New Resistance Gene to a Chinese Blast Fungus Isolate in the Japanese Rice Cultivar Aichi Asahi. Phytopathology 1998, 88, 822–827. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Pan, Q.H.; Wang, L.; Ikehashi, H.; Yamagata, H.; Tanisaka, T. Identification of Two New Genes Conferring Resistance to Rice Blast in the Chinese Native Cultivar ‘Maowangu’. Plant Breed. 1998, 117, 27–31. [Google Scholar] [CrossRef]
- Ahn, S.N.; Kim, Y.K.; Hong, H.C.; Han, S.S.; Kwon, S.J.; Choi, H.C.; Moon, H.P.; McCouch, S.R. Molecular mapping of a gene for resistance to a Korean isolate of rice blast. Rice Genet. Newsl. 1996, 13, 74. [Google Scholar]
- Terashima, T.; Fukuoka, S.; Saka, N.; Kudo, S. Mapping of a Blast Field Resistance GenePi39(t) of Elite Rice Strain Chubu 111. Plant Breed. 2008, 127, 485–489. [Google Scholar] [CrossRef]
- Rybka, K.; Miyamoto, M.; Ando, I.; Saito, A.; Kawasaki, S. High Resolution Mapping of the Indica-Derived Rice Blast Resistance Genes II. Pi-Ta2 and Pi-Ta and a Consideration of Their Origin. Mol. Plant-Microbe Interact. 1997, 10, 517–524. [Google Scholar] [CrossRef]
- Li, W.; Lei, C.; Cheng, Z.; Jia, Y.; Huang, D.; Wang, J.; Wang, J.; Zhang, X.; Su, N.; Guo, X.; et al. Identification of SSR Markers for a Broad-Spectrum Blast Resistance Gene Pi20(t) for Marker-Assisted Breeding. Mol. Breed. 2008, 22, 141–149. [Google Scholar] [CrossRef]
- Imbe, T.; Oba, S.; Yanoria, M.J.T. Tsunematsu, H. A new gene for blast resistance in rice cultivar, IR24. Rice Genet. Newsl. 1997, 14, 60–62. [Google Scholar]
- Hayasaka, H.; Miyao, A.; Yano, M.; Matsunaga, K.; Sasaki, T. RFLP mapping of a rice blast resistance gene Pik. Breed Sci. 1996, 46, 68. [Google Scholar]
- Kaji, R.; Ogawa, T. RFLP mapping of a blast resistance gene Pikm in rice. Breed Sci. 1996, 46, 70. [Google Scholar]
- Naqvi, N.I.; Bonman, J.M.; Mackill, D.J.; Nelson, R.J.; Chattoo, B.B. Identification of RAPD Markers Linked to a Major Blast Resistance Gene in Rice. Mol. Breed. 1995, 1, 341–348. [Google Scholar] [CrossRef]
- Pan, Q. Identification of a New Blast Resistance Gene in the Indica Rice Cultivar Kasalath Using Japanese Differential Cultivars and Isozyme Markers. Phytopathology 1996, 86, 1071. [Google Scholar] [CrossRef]
- Wu, K.S.; Martinez, C.; Lentini, Z.; Tohme, J.; Chumley, F.G.; Scolnik, P.A.; Valent, B. Cloning a blast resistance gene by chromosome walking. In Proceedings of the Third International Rice Genetics Symposium, Manila, Philippines, 16–20 October 1995; Khush, G.S., Ed.; IRRI: Los Baños, Philippines, 1996; pp. 669–674. [Google Scholar]
- Tabien, R.E.; Pinson, S.R.M.; Marchetti, M.A.; Li, Z.; Park, W.D.; Paterson, A.H.; Stansel, J.W. Blast resistance genes from Teqing and Lemont. In Rice Genetics III. Proceedings of Third International Rice Genetics Symposium, Manila, Philippines, 16–20 October 1996; Khush, G.S., Ed.; IRRI: Los Baños, Philippines, 1996; pp. 451–455. [Google Scholar]
- Parco, A. STS markers for the blast resistance gene Pi-1(t). Rice Genome 1995, 4, 9. [Google Scholar]
- Yu, Z.H.; Mackill, D.J.; Bonman, J.M.; McCouch, S.R.; Guiderdoni, E.; Notteghem, J.L.; Tanksley, S.D. Molecular Mapping of Genes for Resistance to Rice Blast (Pyricularia grisea Sacc.). Theor. Appl. Genet. 1996, 93, 859–863. [Google Scholar] [CrossRef]
- Causse, M.A.; Fulton, T.M.; Cho, Y.G.; Ahn, S.N.; Chunwongse, J.; Wu, K.; Xiao, J.; Yu, Z.; Ronald, P.C.; Harrington, S.E. Saturated Molecular Map of the Rice Genome Based on an Interspecific Backcross Population. Genetics 1994, 138, 1251–1274. [Google Scholar] [CrossRef]
- McCouch, S.R.; Nelson, R.J.; Tohme, J.; Zeigler, R.S. Mapping of blast resistance genes in rice. In Rice Blast Disease; Zeigler, R.S., Leong, S.A., Teng, P.S., Eds.; C.A.B. International: Wallingford, UK, 1994; pp. 167–186. [Google Scholar]
- Wang, G.L.; Mackill, D.J.; Bonman, J.M.; McCouch, S.R.; Champoux, M.C.; Nelson, R.J. RFLP Mapping of Genes Conferring Complete and Partial Resistance to Blast in a Durably Resistant Rice Cultivar. Genetics 1994, 136, 1421–1434. [Google Scholar] [CrossRef]
- Wu, K.S.; Tanksley, S.D. Abundance, Polymorphism and Genetic Mapping of Microsatellites in Rice. Mol. Gen. Genet. 1993, 241, 225–235. [Google Scholar] [CrossRef]
- Amante-Bordeos, A.; Sitch, L.A.; Nelson, R.; Dalmacio, R.D.; Oliva, N.P.; Aswidinnoor, H.; Leung, H. Transfer of Bacterial Blight and Blast Resistance from the Tetraploid Wild Rice Oryza minuta to Cultivated Rice, Oryza sativa. Theor. Appl. Genet. 1992, 84, 345–354. [Google Scholar] [CrossRef] [PubMed]
- Mackill, D.J. Inheritance of Blast Resistance in Near-Isogenic Lines of Rice. Phytopathology 1992, 82, 746. [Google Scholar] [CrossRef]
- Yu, Z.H.; Mackill, D.J.; Bonman, J.M.; Tanksley, S.D. Tagging Genes for Blast Resistance in Rice via Linkage to RFLP Markers. Theor. Appl. Genet. 1991, 81, 471–476. [Google Scholar] [CrossRef] [PubMed]
- Goto, I. Genetic Studies on Resistance of Rice Plant to Blast Fungus. (VII). Blast Resistance Genes of Kuroka. Jpn. J. Phytopathol. 1988, 54, 460–465. [Google Scholar] [CrossRef]
- Imbe, T.; Matsumoto, S. Inheritance of Resistance of Rice Varieties to the Blast Fungus Strains Virulent to the Variety “Reiho”. Jpn. J. Breed. 1985, 35, 332–339. [Google Scholar] [CrossRef]
- Goto, I. Genetic Studies on Resistance of Rice Plant to Blast Fungus. II. Difference in Resistance to the Blast Disease between Fukunishiki and Its Parental Cultivar, Zenith. Jpn. J. Phytopathol. 1976, 42, 253–260. [Google Scholar] [CrossRef]
- Shinoda, H.; Toriyama, K.; Yunoki, T.; Ezuka, A.; Sakurai, Y. Studies on the varietal resistance of rice to blast. VI. Linkage relationship of blast resistance genes. Bull. Chugoku Nat. Agric. Exp. Stat. A 1971, 20, 1–25. [Google Scholar]
- Ise, K. Linkage analysis of some blast resistance gene in rice, Oryza sativa L. Jpn. J Breed. 1991, 42, 388–389. [Google Scholar]
- Goto, I. Genetic studies on the resistance of rice plant to the blast fungus I. Inheritance of resistance in crosses Sensho x H-79 and Imochishirazu x H-79. Ann. Phytopathol. Soc. Jpn. 1970, 36, 304–312. [Google Scholar] [CrossRef]
- Fukuoka, S.; Saka, N.; Koga, H.; Ono, K.; Shimizu, T.; Ebana, K.; Hayashi, N.; Takahashi, A.; Hirochika, H.; Okuno, K.; et al. Loss of Function of a Proline-Containing Protein Confers Durable Disease Resistance in Rice. Science 2009, 325, 998–1001. [Google Scholar] [CrossRef]
- Inukai, T.; Nagashima, S.; Kato, M. Pid3-I1 Is a Race-Specific Partial-Resistance Allele at the Pid3 Blast Resistance Locus in Rice. Theor. Appl. Genet. 2019, 132, 395–404. [Google Scholar] [CrossRef]
- Zhao, H.; Wang, X.; Jia, Y.; Minkenberg, B.; Wheatley, M.; Fan, J.; Jia, M.H.; Famoso, A.; Edwards, J.D.; Wamishe, Y.; et al. The Rice Blast Resistance Gene Ptr Encodes an Atypical Protein Required for Broad-Spectrum Disease Resistance. Nat. Commun. 2018, 9, 2039. [Google Scholar] [CrossRef]
- Fukuoka, S.; Yamamoto, S.-I.; Mizobuchi, R.; Yamanouchi, U.; Ono, K.; Kitazawa, N.; Yasuda, N.; Fujita, Y.; Thi Thanh Nguyen, T.; Koizumi, S.; et al. Multiple Functional Polymorphisms in a Single Disease Resistance Gene in Rice Enhance Durable Resistance to Blast. Sci. Rep. 2014, 4, 4550. [Google Scholar] [CrossRef]
- Xu, X.; Hayashi, N.; Wang, C.-T.; Fukuoka, S.; Kawasaki, S.; Takatsuji, H.; Jiang, C.-J. Rice Blast Resistance Gene Pikahei-1(t), a Member of a Resistance Gene Cluster on Chromosome 4, Encodes a Nucleotide-Binding Site and Leucine-Rich Repeat Protein. Mol. Breed. 2014, 34, 691–700. [Google Scholar] [CrossRef]
- Hua, L.; Wu, J.; Chen, C.; Wu, W.; He, X.; Lin, F.; Wang, L.; Ashikawa, I.; Matsumoto, T.; Wang, L.; et al. The Isolation of Pi1, an Allele at the Pik Locus Which Confers Broad Spectrum Resistance to Rice Blast. Theor. Appl. Genet. 2012, 125, 1047–1055. [Google Scholar] [CrossRef]
- Das, A.; Soubam, D.; Singh, P.K.; Thakur, S.; Singh, N.K.; Sharma, T.R. A novel blast resistance gene, Pi54rh cloned from wild species of rice, Oryza rhizomatis confers broad spectrum resistance to Magnaporthe oryzae. Funct. Integr. Genom. 2012, 12, 215–228. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Shi, Y.; Liu, W.; Chai, R.; Fu, Y.; Zhuang, J.; Wu, J. A Pid3 Allele from Rice Cultivar Gumei2 Confers Resistance to Magnaporthe oryzae. J. Genet. Genom. 2011, 38, 209–216. [Google Scholar] [CrossRef] [PubMed]
- Yuan, B.; Zhai, C.; Wang, W.; Zeng, X.; Xu, X.; Hu, H.; Lin, F.; Wang, L.; Pan, Q. The Pik-p Resistance to Magnaporthe oryzae in Rice Is Mediated by a Pair of Closely Linked CC-NBS-LRR Genes. Theor. Appl. Genet. 2011, 122, 1017–1028. [Google Scholar] [CrossRef]
- Takahashi, A.; Hayashi, N.; Miyao, A.; Hirochika, H. Unique Features of the Rice Blast Resistance Pish Locus Revealed by Large Scale Retrotransposon-Tagging. BMC Plant Biol. 2010, 10, 175. [Google Scholar] [CrossRef] [PubMed]
- Hayashi, K.; Yoshida, H. Refunctionalization of the Ancient Rice Blast Disease Resistance Gene Pit by the Recruitment of a Retrotransposon as a Promoter. Plant J. 2009, 57, 413–425. [Google Scholar] [CrossRef]
- Lee, S.K.; Song, M.Y.; Seo, Y.-S.; Kim, H.K.; Ko, S.; Cao, P.-J.; Suh, J.P.; Yi, G.; Roh, J.-H.; Lee, S.; et al. Rice Pi5-Mediated Resistance to Magnaporthe oryzae Requires the Presence of Two Coiled-Coil–Nucleotide-Binding–Leucine-Rich Repeat Genes. Genetics 2009, 181, 1627–1638. [Google Scholar] [CrossRef]
- Ashikawa, I.; Hayashi, N.; Yamane, H.; Kanamori, H.; Wu, J.; Matsumoto, T.; Ono, K.; Yano, M. Two Adjacent Nucleotide-Binding Site–Leucine-Rich Repeat Class Genes Are Required to Confer Pikm-Specific Rice Blast Resistance. Genetics 2008, 180, 2267–2276. [Google Scholar] [CrossRef]
- Lin, F.; Chen, S.; Que, Z.; Wang, L.; Liu, X.; Pan, Q. The Blast Resistance GenePi37Encodes a Nucleotide Binding Site–Leucine-Rich Repeat Protein and Is a Member of a Resistance Gene Cluster on Rice Chromosome 1. Genetics 2007, 177, 1871–1880. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.X.; Yano, M.; Yamanouchi, U.; Iwamoto, M.; Monna, L.; Hayasaka, H.; Katayose, Y.; Sasaki, T. The Pib Gene for Rice Blast Resistance Belongs to the Nucleotide Binding and Leucine-Rich Repeat Class of Plant Disease Resistance Genes. Plant J. 1999, 19, 55–64. [Google Scholar] [CrossRef] [PubMed]
- Hittalmani, S.; Parco, A.; Mew, T.V.; Zeigler, R.S.; Huang, N. Fine Mapping and DNA Marker-Assisted Pyramiding of the Three Major Genes for Blast Resistance in Rice. Theor. Appl. Genet. 2000, 100, 1121–1128. [Google Scholar] [CrossRef]
- Singh, V.K.; Singh, A.; Singh, S.P.; Ellur, R.K.; Choudhary, V.; Sarkel, S.; Singh, D.; Krishnan, S.G.; Nagarajan, M.; Vinod, K.K.; et al. Incorporation of Blast Resistance into “PRR78”, an Elite Basmati Rice Restorer Line, through Marker Assisted Backcross Breeding. Field Crop. Res. 2012, 128, 8–16. [Google Scholar] [CrossRef]
- Skamnioti, P.; Gurr, S.J. Against the Grain: Safeguarding Rice from Rice Blast Disease. Trends Biotechnol. 2009, 27, 141–150. [Google Scholar] [CrossRef] [PubMed]
- Zenbayashi, K.; Ashizawa, T.; Tani, T.; Koizumi, S. Mapping of the QTL (Quantitative Trait Locus) Conferring Partial Resistance to Leaf Blast in Rice Cultivar Chubu 32. Theor. Appl. Genet. 2002, 104, 547–552. [Google Scholar] [CrossRef]
- Sirithunya, P.; Tragoonrung, S.; Vanavichit, A.; Pa-In, N.; Vongsaprom, C.; Toojinda, T. Quantitative trait loci associated with leaf and neck blast resistance in recombinant inbred line population of rice (Oryza sativa). DNA Res. 2002, 9, 79–88. [Google Scholar] [CrossRef][Green Version]
- Talukder, Z.I.; Tharreau, D.; Price, A.H. Quantitative Trait Loci Analysis Suggests That Partial Resistance to Rice Blast Is Mostly Determined by Race–Specific Interactions. New Phytol. 2004, 162, 197–209. [Google Scholar] [CrossRef]
- Cho, Y.C.; Kwon, S.W.; Suh, J.P.; Kim, J.J.; Lee, J.H.; Roh, J.H.; Oh, M.K.; Kim, M.K.; Ahn, S.N.; Koh, H.J.; et al. QTLs identification and confirmation of field resistance to leaf blast in temperate japonica rice (Oryza sativa L.). J. Crop Sci. Biotechnol. 2008, 11, 269–276. [Google Scholar]
- Koide, Y.; Kobayashi, N.; Xu, D.; Fukuta, Y. Resistance genes and selection DNA markers for blast disease in rice (Oryza sativa L.). Jpn. Agril. Res. Q. 2009, 43, 255–280. [Google Scholar] [CrossRef]
- Jia, Y.; Liu, G. Mapping quantitative trait Loci for resistance to rice blast. Phytopathology 2011, 101, 176–181. [Google Scholar] [CrossRef] [PubMed]
- Ashkani, S.; Rafii, M.Y.; Rahim, H.A.; Latif, M.A. Genetic dissection of rice blast resistance by QTL mapping approach using an F3 population. Mol. Biol. Rep. 2013, 40, 2503–2515. [Google Scholar] [CrossRef] [PubMed]
- Ishihara, T.; Hayano-Saito, Y.; Oide, S.; Ebana, K.; La, N.T.; Hayashi, K.; Ashizawa, T.; Suzuki, F.; Koizumi, S. Quantitative trait locus analysis of resistance to panicle blast in the rice cultivar Miyazakimochi. Rice 2014, 7, 2. [Google Scholar] [CrossRef]
- Fang, N.; Wang, R.; He, W.; Yin, C.; Guan, C.; Chen, H.; Huang, J.; Wang, J.; Bao, Y.; Zhang, H. QTL mapping of panicle blast resistance in japonica landrace heikezijing and its application in rice breeding. Mol. Breed. 2016, 36, 171. [Google Scholar] [CrossRef]
- Nagaoka, I.; Sasahara, H.; Tabuchi, H.; Shigemune, A.; Matsushita, K.; Maeda, H.; Goto, A.; Fukuoka, S.; Ando, T.; Miura, K. Quantitative trait loci analysis of blast resistance in Oryza sativa L. ‘Hokuriku 193’. Breed. Sci. 2017, 67, 159–164. [Google Scholar] [CrossRef]
- Fang, N.; Wei, X.; Shen, L.; Yu, Y.; Li, M.; Yin, C.; He, W.; Guan, C.; Chen, H.; Zhang, H.; et al. Fine mapping of a panicle blast resistance gene Pb-bd1 in japonica landrace Bodao and its application in rice breeding. Rice 2019, 12, 18. [Google Scholar] [CrossRef]
- Devi, S.J.S.R.; Singh, K.; Umakanth, B.; Vishalakshi, B.; Rao, K.V.S.; Suneel, B.; Sharma, S.K.; Kadambari, G.K.M.; Prasad, M.S.; Senguttvel, P.; et al. Identification and Characterization of a Large Effect QTL from Oryza glumaepatula Revealed Pi68(t) as Putative Candidate Gene for Rice Blast Resistance. Rice 2020, 13, 17. [Google Scholar] [CrossRef]
- Korinsak, S.; Wongsaprom, C.; Jamboonsri, W.; Sriprakhon, S.; Sirithunya, K.; Vanavichit, A.; Toojinda, T. Identification of broad-spectrum resistance QTLs against rice blast fungus and their application in different rice genetic backgrounds. J. Genet. 2022, 101, 16. [Google Scholar] [CrossRef]
- Tian, T.; Chen, L.; Ai, Y.; He, H. Selection of Candidate Genes Conferring Blast Resistance and Heat Tolerance in Rice through Integration of Meta-QTLs and RNA-Seq. Genes 2022, 13, 224. [Google Scholar] [CrossRef]
- Zhao, K.; Tung, C.-W.; Eizenga, G.C.; Wright, M.H.; Ali, M.L.; Price, A.H.; Norton, G.J.; Islam, M.R.; Reynolds, A.; Mezey, J.; et al. Genome-Wide Association Mapping Reveals a Rich Genetic Architecture of Complex Traits in Oryza sativa. Nat. Commun. 2011, 2, 467. [Google Scholar] [CrossRef]
- Wang, C.; Yang, Y.; Yuan, X.; Xu, Q.; Feng, Y.; Yu, H.; Wang, Y.; Wei, X. Genome-Wide Association Study of Blast Resistance in Indica Rice. BMC Plant Biol. 2014, 14, 311. [Google Scholar] [CrossRef]
- Shinada, H.; Yamamoto, T.; Sato, H.; Yamamoto, E.; Hori, K.; Yonemaru, J.; Sato, T.; Fujino, K. Quantitative Trait Loci for Rice Blast Resistance Detected in a Local Rice Breeding Population by Genome-Wide Association Mapping. Breed. Sci. 2015, 65, 388–395. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Wang, X.; Jia, M.H.; Ghai, P.; Lee, F.N.; Jia, Y. Genome-Wide Association of Rice Blast Disease Resistance and Yield-Related Components of Rice. Mol. Plant-Microbe Interact. 2015, 28, 1383–1392. [Google Scholar] [CrossRef] [PubMed]
- Guo, L.Y.; Guo, W.; Zhao, H.W.; Wang, J.G.; Liu, H.L.; Sun, J.; Zheng, H.L.; Sha, H.J.; Zou, D.T. Association mapping and resistant alleles’ analysis for japonica rice blast resistance. Plant Breed. 2015, 134, 646–652. [Google Scholar] [CrossRef]
- Raboin, L.-M.; Ballini, E.; Tharreau, D.; Ramanantsoanirina, A.; Frouin, J.; Courtois, B.; Ahmadi, N. Association Mapping of Resistance to Rice Blast in Upland Field Conditions. Rice 2016, 9, 59. [Google Scholar] [CrossRef]
- McCouch, S.R.; Wright, M.H.; Tung, C.-W.; Maron, L.G.; McNally, K.L.; Fitzgerald, M.; Singh, N.; DeClerck, G.; Agosto-Perez, F.; Korniliev, P.; et al. Open access resources for genome wide association mapping in rice. Nat. Commun. 2016, 7, 10532. [Google Scholar] [CrossRef]
- Kang, H.; Wang, Y.; Peng, S.; Zhang, Y.; Xiao, Y.; Wang, D.; Qu, S.; Li, Z.; Yan, S.; Wang, Z.; et al. Dissection of the genetic architecture of rice resistance to the blast fungus Magnaporthe oryzae. Mol. Plant Pathol. 2016, 17, 959–972. [Google Scholar] [CrossRef]
- Mgonja, E.M.; Park, C.H.; Kang, H.; Balimponya, E.G.; Opiyo, S.; Bellizzi, M.; Mutiga, S.K.; Rotich, F.; Ganeshan, V.D.; Mabagala, R.; et al. Genotyping-by-Sequencing-Based Genetic Analysis of African Rice Cultivars and Association Mapping of Blast Resistance Genes Against Magnaporthe oryzae Populations in Africa. Phytopathology 2017, 107, 1039–1046. [Google Scholar] [CrossRef]
- Lin, H.A.; Chen, S.Y.; Chang, F.Y.; Tung, C.W.; Chen, Y.C.; Shen, W.C.; Chen, R.S.; Wu, C.W.; Chung, C.L. Genome-wide association study of rice genes and loci conferring resistance to Magnaporthe oryzae isolates from Taiwan. Bot. Stud. 2018, 59, 32. [Google Scholar] [CrossRef] [PubMed]
- Korinsaka, S.; Tangphatsornruangb, S.; Pootakhamb, W.; Wanchanab, S.; Plabplac, A.; Jantasuriyaratd, C.; Patarapuwadole, S.; Vanavichitf, S. Genome-wide association mapping of virulence gene in rice blast fungus Magnaporthe oryzae using a genotyping by sequencing approach. Genomics 2019, 111, 661–668. [Google Scholar] [CrossRef]
- Volante, A.; Tondelli, A.; Desiderio, F.; Abbruscato, P.; Menin, B.; Biselli, C.; Casella, L.; Singh, N.; McCouch, S.R.; Tharreau, D.; et al. Genome Wide Association Studies for Japonica Rice Resistance to Blast in Field and Controlled Conditions. Rice 2020, 13, 71. [Google Scholar] [CrossRef]
- Jiang, H.; Feng, Y.; Qiu, L.; Gao, G.; Zhang, Q.; He, Y. Identification of Blast Resistance QTLs Based on Two Advanced Backcross Populations in Rice. Rice 2020, 13, 1. [Google Scholar] [CrossRef]
- Frontini, M.; Boisnard, A.; Frouin, J.; Ouikene, M.; Morel, J.B.; Ballini, E. Genome-wide association of rice response to blast fungus identifies loci for robust resistance under high nitrogen. BMC Plant Biol. 2021, 21, 99. [Google Scholar] [CrossRef]
- Tanweer, F.A.; Rafii, M.Y.; Sijam, K.; Rahim, H.A.; Ahmed, F.; Ashkani, S.; Latif, M.A. Introgression of Blast Resistance Genes (Putative Pi-b and Pi-Kh) into Elite Rice Cultivar MR219 through Marker-Assisted Selection. Front. Plant Sci. 2015, 6, 1002. [Google Scholar] [CrossRef] [PubMed]
- Fuchs, M. Pyramiding resistance-conferring gene sequences in crops. Curr. Opin. Virol. 2017, 26, 36–42. [Google Scholar] [CrossRef]
- Das, G.; Patra, J.K.; Baek, K.H. Insight into MAS: A molecular tool for development of stress resistant and quality of rice through gene stacking. Front. Plant Sci. 2017, 8, 985. [Google Scholar] [CrossRef]
- Xiao, W.; Yang, Q.; Huang, M.; Guo, T.; Liu, Y.; Wang, J.; Yang, G.; Zhou, J.; Yang, J.; Zhu, X.; et al. Improvement of Rice Blast Resistance by Developing Monogenic Lines, Two-Gene Pyramids and Three-Gene Pyramid through MAS. Rice 2019, 12, 78. [Google Scholar] [CrossRef]
- Dixit, S.; Singh, U.M.; Singh, A.K.; Alam, S.; Venkateshwarlu, C.; Nachimuthu, V.V.; Yadav, S.; Abbai, R.; Selvaraj, R.; Devi, M.N.; et al. Marker Assisted Forward Breeding to Combine Multiple Biotic-Abiotic Stress Resistance/Tolerance in Rice. Rice 2020, 13, 29. [Google Scholar] [CrossRef] [PubMed]
- Fjellstrom, R.; McClung, A.M.; Shank, A.R. SSR Markers Closely Linked to the Pi-z Locus Are Useful for Selection of Blast Resistance in a Broad Array of Rice Germplasm. Mol. Breed. 2006, 17, 149–157. [Google Scholar] [CrossRef]
- Wang, Z.; Jia, Y.; Rutger, J.N.; Xia, Y. Rapid Survey for Presence of a Blast Resistance Gene Pi-Ta in Rice Cultivars Using the Dominant DNA Markers Derived from Portions of the Pi-Ta Gene. Plant Breed. 2007, 126, 36–42. [Google Scholar] [CrossRef]
- Wen, S.; Gao, B. Introgressing Blast Resistant Gene Pi-9(t) into Elite Rice Restorer Luhui17 by Marker-Assisted Selection. Rice Genom. Genet. 2012, 2, 31–36. [Google Scholar] [CrossRef][Green Version]
- Hua, L.X.; Liang, L.-Q.; He, X.-Y.; Wang, L.; Zhang, W.-S.; Liu, W.; Liu, X.-Q.; Lin, F. Development of a Marker Specific for the Rice Blast Resistance Gene Pi39 in the Chinese Cultivar Q15 and Its Use in Genetic improvement. Biotechnol. Biotechnol. Equip. 2015, 29, 448–456. [Google Scholar] [CrossRef]
- Hasan, N.; Rafii, M.; Rahim, H.; Syd Ali, N.; Mazlan, N.; Abdullah, S. Introgression of Pi-Kh Resistance Gene into a Malaysian Cultivar, MR264 Using Marker-Assisted Backcrossing (MABC). Int. J. Agric. Biol. 2015, 17, 1172–1178. [Google Scholar] [CrossRef]
- Beser, N.; Del Valle, M.M.; Kim, S.M.; Vinarao, B.R.; Surek, H.; Jena, K.K. Marker-assisted introgression of a broad-spectrum resistance gene, Pi40 improved blast resistance of two elite rice (Oryza sativa L.). Mol. Plant Breed. 2016, 7, 1–15. [Google Scholar]
- Araujo, L.; Soares, J.M.; de Filippi, M.C.C.; Rodrigues, F.Á. Cytological Aspects of Incompatible and Compatible Interactions between Rice, Wheat and the Blast Pathogen Pyricularia oryzae. Sci. Agric. 2016, 73, 177–183. [Google Scholar] [CrossRef]
- Singh, A.; Singh, V.K.; Singh, S.P.; Pandian, R.T.P.; Ellur, R.K.; Singh, D.; Bhowmick, P.K.; Gopala Krishnan, S.; Nagarajan, M.; Vinod, K.K.; et al. Molecular Breeding for the Development of Multiple Disease Resistance in Basmati Rice. AoB Plants 2012, 2012, pls029. [Google Scholar] [CrossRef]
- Gouda, P.K.; Saikumar, S.; Varma, C.M.K.; Nagesh, K.; Thippeswamy, S.; Shenoy, V.; Ramesha, M.S.; Shashidhar, H.E. Marker-Assisted Breeding of Pi-1 and Piz-5 Genes Imparting Resistance to Rice Blast in PRR78, Restorer Line of Pusa RH-10 Basmati Rice Hybrid. Plant Breed. 2013, 132, 61–69. [Google Scholar] [CrossRef]
- Oddin, J.; Durga Rani, H.V.; Swathi, G.; Anuradha, C.H.; Vanisri, S.; SheshuMadhav, M.; Sundaram, R.M.; ArunPrem Kumar, N. Introgression of blast resistance genes Pi-54 and Pi1 into cold tolerant variety Tellahamsa, by marker assisted selection. Int. J. Curr. Res. 2015, 7, 22348–22353. [Google Scholar]
- Madhavi, R.K.; Rambabu, R.; Abhilash Kumar, V.; Vijay Kumar, S.; Aruna, J.; Ramesh, S.; Sundaram, R.M.; Laha, G.S.; Sheshu Madhav, M.; Ravindra babu, V.; et al. Marker Assisted Introgression of Blast (Pi-2 and Pi-54) Genes in to the Genetic Background of Elite, Bacterial Blight Resistant Indica Rice Variety, Improved Samba Mahsuri. Euphytica 2016, 212, 331–342. [Google Scholar] [CrossRef]
- Xiao, W.; Luo, L.; Wang, H.; Guo, T.; Liu, Y.; Zhou, J.; Zhu, X.; Yang, Q.; Chen, Z. Pyramiding of Pi46 and Pita to Improve Blast Resistance and to Evaluate the Resistance Effect of the Two R Genes. J. Integr. Agric. 2016, 15, 2290–2298. [Google Scholar] [CrossRef]
- Hegde, S.S.; Prashanthi, S.K. Identification of polymorphic markers and introgression of Pi1 and Pi2 genes for blast resistance in rice. J Farm Sci. 2016, 29, 327–331. [Google Scholar]
- Luo, Y.; Ma, T.; Zhang, A.; Ong, K.H.; Li, Z.; Yang, J.; Yin, Z. Marker-assisted breeding of the rice restorer line Wanhui 6725 for disease resistance, submergence tolerance and aromatic fragrance. Rice 2016, 9, 66. [Google Scholar] [CrossRef]
- Divya, B.; Robin, S.; Rabindran, R.; Senthil, S.; Raveendran, M.; Joel, A.J. Marker Assisted Backcross Breeding Approach to Improve Blast Resistance in Indian Rice (Oryza sativa) Variety ADT43. Euphytica 2014, 200, 61–77. [Google Scholar] [CrossRef]
- Singh, A.K.; Singh, V.K.; Singh, A.; Ellur, R.K.; Pandian, R.T.P.; Gopala Krishnan, S.; Singh, U.D.; Nagarajan, M.; Vinod, K.K.; Prabhu, K.V. Introgression of Multiple Disease Resistance into a Maintainer of Basmati Rice CMS Line by Marker Assisted Backcross Breeding. Euphytica 2015, 203, 97–107. [Google Scholar] [CrossRef]
- Balachiranjeevi, C.; Bhaskar, N.S.; Abhilash, V.; Akanksha, S.; Viraktamath, B.C.; Madhav, M.S.; Hariprasad, A.S.; Laha, G.S.; Prasad, M.S.; Balachandran, S.M.; et al. Marker-Assisted Introgression of Bacterial Blight and Blast Resistance into DRR17B, an Elite, Fine-Grain Type Maintainer Line of Rice. Mol. Breed. 2015, 35, 151. [Google Scholar] [CrossRef]
- Srikanth, S.; Pandey, M.K.; BalaChiranjeevi, C.H.; Hajira, S.K.; Vijay Kumar, S.; Kousik, M.B.V.N.; Bhadana, V.P.; SheshuMadhav, M.; Suneetha, K.; Subba Rao, L.V.; et al. Introgression of major bacterial blight and blast resistant genes into Vallabh Basmati 22, an elite Basmati variety. Int. J. Dev. Res. 2016, 6, 8366–8370. [Google Scholar]
- Wu, Y.; Yu, L.; Pan, C.; Dai, Z.; Li, Y.; Xiao, N.; Zhang, X.; Ji, H.; Huang, N.; Zhao, B.; et al. Development of Near-Isogenic Lines with Different Alleles of Piz Locus and Analysis of Their Breeding Effect under Yangdao 6 Background. Mol. Breed. 2016, 36, 12. [Google Scholar] [CrossRef]
- Narayanan, N.N.; Baisakh, N.; Vera Cruz, C.M.; Gnanamanickam, S.S.; Datta, K.; Datta, S.K. Molecular Breeding for the Development of Blast and Bacterial Blight Resistance in Rice Cv. IR50. Crop Sci. 2002, 42, 2072–2079. [Google Scholar] [CrossRef]
- Ellur, R.K.; Khanna, A.; Yadav, A.; Pathania, S.; Rajashekara, H.; Singh, V.K.; Gopala Krishnan, S.; Bhowmick, P.K.; Nagarajan, M.; Vinod, K.K.; et al. Improvement of Basmati Rice Varieties for Resistance to Blast and Bacterial Blight Diseases Using Marker Assisted Backcross Breeding. Plant Sci. J. 2016, 242, 330–341. [Google Scholar] [CrossRef] [PubMed]
- Arunakumari, K.; Durgarani, C.V.; Satturu, V.; Sarikonda, K.R.; Chittoor, P.D.R.; Vutukuri, B.; Laha, G.S.; Nelli, A.P.K.; Gattu, S.; Jamal, M.; et al. Marker-Assisted Pyramiding of Genes Conferring Resistance Against Bacterial Blight and Blast Diseases into Indian Rice Variety MTU1010. Rice Sci. 2016, 23, 306–316. [Google Scholar] [CrossRef]
- Janaki Ramayya, P.; Vinukonda, V.P.; Singh, U.M.; Alam, S.; Venkateshwarlu, C.; Vipparla, A.K.; Dixit, S.; Yadav, S.; Abbai, R.; Badri, J.; et al. Marker-assisted forward and backcross breeding for improvement of elite Indian rice variety Naveen for multiple biotic and abiotic stress tolerance. PLoS ONE 2021, 16, e0256721. [Google Scholar]
- Laxmi Prasanna, B.; Dangi, K.S.; Damodar Raju, C.H.; Jagadeeshwar, R.; Rekha, G.; Sinha, P.; Aleena, D.; Harika, G.; Mastanbee, S.K.; Swapnil Ravindra, K.; et al. Marker Assisted Pedigree Breeding Based Improvement of the Indian Mega Variety of Rice MTU1010 for Resistance against Bacterial Blight and Blast and Tolerance to Low Soil Phosphorus. PLoS ONE 2022, 17, e0260535. [Google Scholar] [CrossRef]
- Singh, V.K.; Singh, A.; Singh, S.P.; Ellur, R.K.; Singh, D.; Gopala Krishnan, S.; Bhowmick, P.K.; Nagarajan, M.; Vinod, K.K.; Singh, U.D.; et al. Marker-Assisted Simultaneous but Stepwise Backcross Breeding for Pyramiding Blast Resistance Genes Piz5 and Pi54 into an Elite Basmati Rice Restorer Line ‘PRR78’. Plant Breed. 2013, 132, 486–495. [Google Scholar] [CrossRef]
- Usatov, A.V.; Kostylev, P.I.; Azarin, K.V.; Markin, N.V.; Makarenko, M.S.; Khachumov, V.A.; Bibov, M.Y. Introgression of the Rice Blast Resistance Genes Pi1, Pi2 and Pi33 into Russian Rice Varieties by Marker-Assisted Selection. Indian J. Genet. Plant Breed. 2016, 76, 18–23. [Google Scholar] [CrossRef][Green Version]
- Fu, C. Genetic Improvement of Resistance to Blast and Bacterial Blight of the Elite Maintainer Line Rongfeng B in Hybrid Rice (Oryza sativa L.) by Using Marker-Assisted Selection. Afr. J. Biotechnol. 2012, 11, 13104–13114. [Google Scholar] [CrossRef]
- Kumar, V.A.; Balachiranjeevi, C.H.; Naik, S.B.; Rambabu, R.; Rekha, G.; Madhavi, K.R.; Harika, G.; Vijay, S.; Pranathi, K.; Hajira, S.K.; et al. Marker-Assisted Introgression of the Major Bacterial Blight Resistance Gene, Xa21 and Blast Resistance Gene, Pi54 into RPHR-1005, the Restorer Line of the Popular Rice Hybrid, DRRH3. J. Plant Biochem. Biotechnol. 2016, 25, 400–409. [Google Scholar] [CrossRef]
- Kumar, V.A.; Balachiranjeevi, C.H.; Naik, S.B.; Rambabu, R.; Rekha, G.; Harika, G.; Hajira, S.K.; Pranathi, K.; Vijay, S.; Anila, M.; et al. Marker-Assisted Improvement of the Elite Restorer Line of Rice, RPHR-1005 for Resistance against Bacterial Blight and Blast Diseases. J. Genet. 2016, 95, 895–903. [Google Scholar] [CrossRef]
- Pinta, W.; Toojinda, T.; Thummabenjapone, P.; Sanitchon, J. Pyramiding of Blast and Bacterial Leaf Blight Resistance Genes into Rice Cultivar RD6 Using Marker Assisted Selection. Afr. J. Biotechnol. 2013, 12, 4432–4438. [Google Scholar] [CrossRef]
- Das, G.; Rao, G.J.N. Molecular Marker Assisted Gene Stacking for Biotic and Abiotic Stress Resistance Genes in an Elite Rice Cultivar. Front. Plant Sci. 2015, 6, 698. [Google Scholar] [CrossRef] [PubMed]
- Nishizawa, Y.; Nishio, Z.; Nakazono, K.; Soma, M.; Nakajima, E.; Ugaki, M.; Hibi, T. Enhanced Resistance to Blast (Magnaporthe grisea) in Transgenic Japonica Rice by Constitutive Expression of Rice Chitinase. Theor. Appl. Genet. 1999, 99, 383–390. [Google Scholar] [CrossRef] [PubMed]
- Gandikota, M.; De Kochko, A.; Chen, L.; Ithal, N.; Fauquet, C.; Reddy, A.R. Development of transgenic rice plants expressing maize anthocyanin genes and increased blast resistance. Mol. Plant Breed. 2001, 7, 73–83. [Google Scholar]
- Nishizawa, Y.; Saruta, M.; Nakazono, K.; Nishio, Z.; Soma, M.; Yoshida, T.; Nakajima, E.; Hibi, T. Characterization of Transgenic Rice Plants Over-Expressing the Stress-Inducible β-Glucanase Gene Gns1. Plant Mol. Biol. 2003, 51, 143–152. [Google Scholar] [CrossRef] [PubMed]
- Coca, M.; Penas, G.; Gómez, J.; Campo, S.; Bortolotti, C.; Messeguer, J.; Segundo, B.S. Enhanced Resistance to the Rice Blast Fungus Magnaporthe grisea Conferred by Expression of a Cecropin A Gene in Transgenic Rice. Planta 2006, 223, 392–406. [Google Scholar] [CrossRef]
- Zhang, S.; Wei, Y.; Pan, H. Transgenic Rice Plants Expressing a Novel Antifreeze Glycopeptide Possess Resistance to Cold and Disease. Zeitschrift für Naturforschung C 2007, 62, 583–591. [Google Scholar] [CrossRef]
- Imamura, T.; Yasuda, M.; Kusano, H.; Nakashita, H.; Ohno, Y.; Kamakura, T.; Taguchi, S.; Shimada, H. Acquired Resistance to the Rice Blast in Transgenic Rice Accumulating the Antimicrobial Peptide Thanatin. Transgenic Res. 2010, 19, 415–424. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Chen, D.; Chen, X.; Lei, C.; Ma, B.; Wang, Y.; Li, S. Rice Blast Resistance of Transgenic Rice Plants with Pi-D2 Gene. Rice Sci. 2010, 17, 179–184. [Google Scholar] [CrossRef]
- Helliwell, E.E.; Wang, Q.; Yang, Y. Transgenic Rice with Inducible Ethylene Production Exhibits Broad-Spectrum Disease Resistance to the Fungal Pathogens Magnaporthe oryzae and Rhizoctonia solani. Plant Biotechnol. J. 2013, 11, 33–42. [Google Scholar] [CrossRef]
- Feng, D.R.; Wei, J.W.; Xu, X.P.; Xu, Y.; Li, B.J. Introduction of multiple antifungal protein genes into rice and preliminary study on resistance to Pyricularia oryzae of transgenic rices. Acta Sci. Nat. Univ. Sunyatsen 1999, 38, 62–66, (In Chinese with English Abstract). [Google Scholar]
- Zhu, C.; Bortesi, L.; Baysal, C.; Twyman, R.M.; Fischer, R.; Capell, T.; Schillberg, S.; Christou, P. Characteristics of Genome Editing Mutations in Cereal Crops. Trends Plant Sci. 2017, 22, 38–52. [Google Scholar] [CrossRef] [PubMed]
- Mishra, R.; Joshi, R.K.; Zhao, K. Genome Editing in Rice: Recent Advances, Challenges, and Future Implications. Front. Plant Sci. 2018, 9, 1361. [Google Scholar] [CrossRef] [PubMed]
- Pingoud, A.; Silva, G.H. Precision Genome Surgery. Nat. Biotechnol. 2007, 25, 743–744. [Google Scholar] [CrossRef]
- Li, T.; Huang, S.; Jiang, W.Z.; Wright, D.; Spalding, M.H.; Weeks, D.P.; Yang, B. TAL Nucleases (TALNs): Hybrid Proteins Composed of TAL Effectors and FokI DNA-Cleavage Domain. Nucleic Acids Res. 2011, 39, 359–372. [Google Scholar] [CrossRef]
- Zhang, H.; Wu, Z.; Wang, C.; Li, Y.; Xu, J.-R. Germination and Infectivity of Microconidia in the Rice Blast Fungus Magnaporthe oryzae. Nat. Commun. 2014, 5, 4518. [Google Scholar] [CrossRef]
- Zhou, Y.; Xu, S.; Jiang, N.; Zhao, X.; Bai, Z.; Liu, J.; Yao, W.; Tang, Q.; Xiao, G.; Lv, C.; et al. Engineering of Rice Varieties with Enhanced Resistances to Both Blast and Bacterial Blight Diseases via CRISPR/Cas9. Plant Biotechnol. J. 2022, 20, 876–885. [Google Scholar] [CrossRef]
- Tao, H.; Shi, X.; He, F.; Wang, D.; Xiao, N.; Fang, H.; Wang, R.; Zhang, F.; Wang, M.; Li, A.; et al. Engineering Broad-spectrum Disease-resistant Rice by Editing Multiple Susceptibility Genes. J. Integr. Plant Biol. 2021, 63, 1639–1648. [Google Scholar] [CrossRef]
- Ma, J.; Chen, J.; Wang, M.; Ren, Y.; Wang, S.; Lei, C.; Cheng, Z. Sodmergen Disruption of OsSEC3A Increases the Content of Salicylic Acid and Induces Plant Defense Responses in Rice. J. Exp. Bot. 2018, 69, 1051–1064. [Google Scholar] [CrossRef]
- Xie, K.; Yang, Y. RNA-Guided Genome Editing in Plants Using a CRISPR–Cas System. Mol. Plant 2013, 6, 1975–1983. [Google Scholar] [CrossRef] [PubMed]
- Boch, J.; Scholze, H.; Schornack, S.; Landgraf, A.; Hahn, S.; Kay, S.; Lahaye, T.; Nickstadt, A.; Bonas, U. Breaking the Code of DNA Binding Specificity of TAL-Type III Effectors. Science 2009, 326, 1509–1512. [Google Scholar] [CrossRef] [PubMed]
- Nemudryi, A.A.; Valetdinova, K.R.; Medvedev, S.P.; Zakian, S.M. TALEN and CRISPR/Cas genome editing systems: Tools of discovery. Acta Nat. 2014, 6, 19–40. [Google Scholar] [CrossRef]
- Schornack, S.; Meyer, A.; Römer, P.; Jordan, T.; Lahaye, T. Gene-for-Gene-Mediated Recognition of Nuclear-Targeted AvrBs3-like Bacterial Effector Proteins. J. Plant Physiol. 2006, 163, 256–272. [Google Scholar] [CrossRef] [PubMed]
- Moscou, M.J.; Bogdanove, A.J. A Simple Cipher Governs DNA Recognition by TAL Effectors. Science 2009, 326, 1501. [Google Scholar] [CrossRef]
- Arazoe, T.; Ogawa, T.; Miyoshi, K.; Yamato, T.; Ohsato, S.; Sakuma, T.; Yamamoto, T.; Arie, T.; Kuwata, S. Tailor-Made TALEN System for Highly Efficient Targeted Gene Replacement in the Rice Blast Fungus. Biotechnol. Bioeng. 2015, 112, 1335–1342. [Google Scholar] [CrossRef] [PubMed]
- Gogarten, J.P.; Hilario, E. Inteins, Introns, and Homing Endonucleases: Recent Revelations about the Life Cycle of Parasitic Genetic Elements. BMC Evol. Biol. 2006, 6, 94. [Google Scholar] [CrossRef]
- Orlowski, J.; Boniecki, M.; Bujnicki, J.M. I-Ssp6803I: The First Homing Endonuclease from the PD-(D/E)XK Superfamily Exhibits an Unusual Mode of DNA Recognition. Bioinformatics 2007, 23, 527–530. [Google Scholar] [CrossRef]
- Maeder, M.L.; Gersbach, C.A. Genome-Editing Technologies for Gene and Cell Therapy. Mol. Ther. 2016, 24, 430–446. [Google Scholar] [CrossRef]
- Gupta, A.; Meng, X.; Zhu, L.J.; Lawson, N.D.; Wolfe, S.A. Zinc Finger Protein-Dependent and -Independent Contributions to the in Vivo off-Target Activity of Zinc Finger Nucleases. Nucleic Acids Res. 2011, 39, 381–392. [Google Scholar] [CrossRef] [PubMed]
- Paschon, D.E.; Lussier, S.; Wangzor, T.; Xia, D.F.; Li, P.W.; Hinkley, S.J.; Scarlott, N.A.; Lam, S.C.; Waite, A.J.; Truong, L.N.; et al. Diversifying the Structure of Zinc Finger Nucleases for High-Precision Genome Editing. Nat. Commun. 2019, 10, 1133. [Google Scholar] [CrossRef] [PubMed]
- Gammage, P.A.; Viscomi, C.; Simard, M.-L.; Costa, A.S.H.; Gaude, E.; Powell, C.A.; Van Haute, L.; McCann, B.J.; Rebelo-Guiomar, P.; Cerutti, R.; et al. Genome Editing in Mitochondria Corrects a Pathogenic MtDNA Mutation in Vivo. Nat. Med. 2018, 24, 1691–1695. [Google Scholar] [CrossRef] [PubMed]
- Cantos, C.; Francisco, P.; Trijatmiko, K.R.; Slamet-Loedin, I.; Chadha-Mohanty, P.K. Identification of ‘safe harbor’ Loci in Indica rice genome by harnessing the property of zinc-finger nucleases to induce DNA damage and repair. Front. Plant Sci. 2014, 5, 302. [Google Scholar] [CrossRef] [PubMed]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).