
Prospects for Using Phosphate-Solubilizing Microorganisms as Natural Fertilizers in Agriculture




Abstract
1. Introduction
2. Soil Phosphate-Solubilizing Microorganisms
3. Biochemical Properties of the Soil Related to the Bioavailability of Phosphates
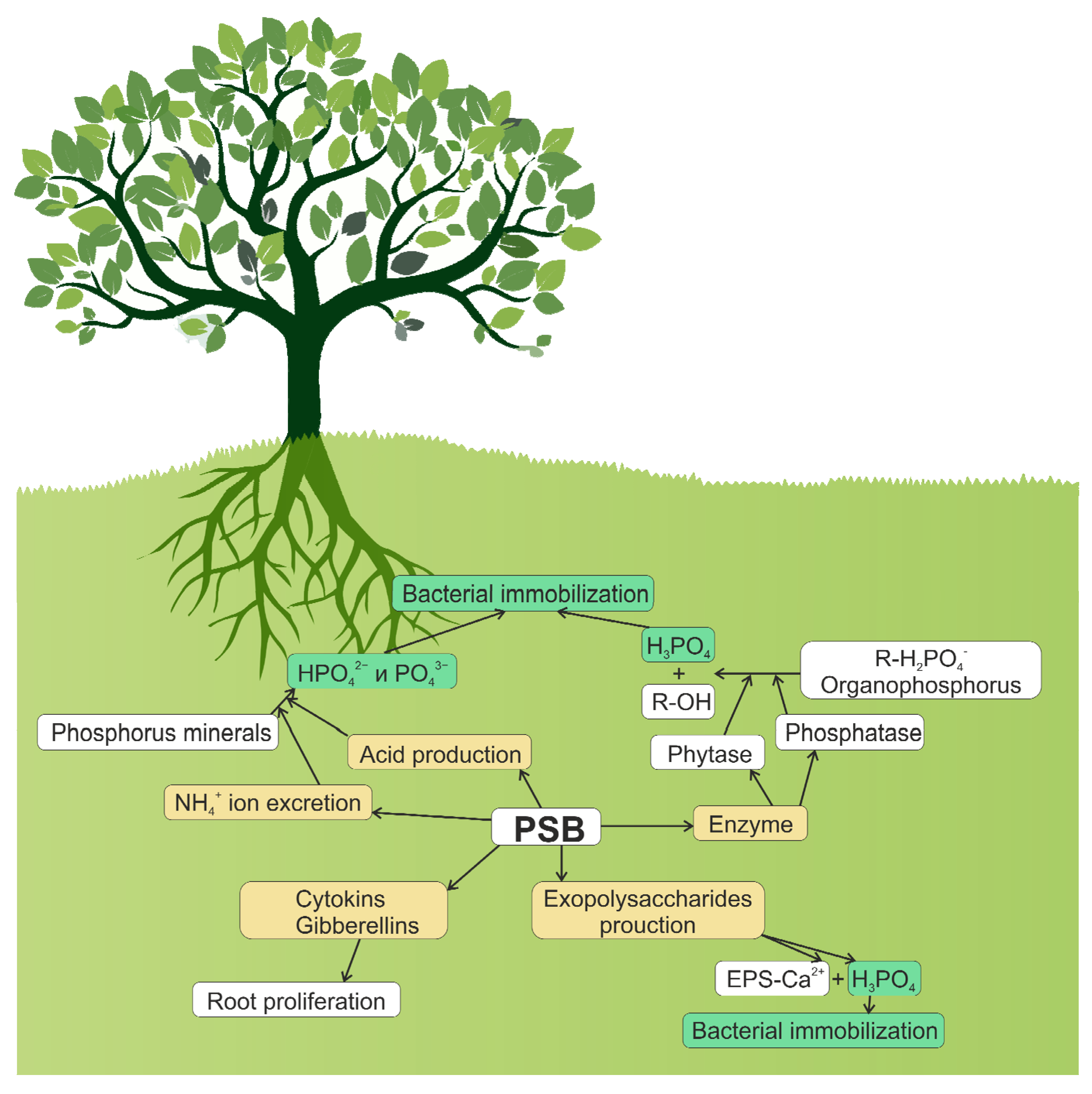
4. Mechanisms of Inorganic Phosphate Solubilization
5. Mineralization of Organic Phosphate-Containing Compounds
6. Bacterial Influence on the Root System
7. Factors Affecting the Microbial Phosphate Solubilization
8. Solubilization of Mineral Phosphates: Molecular Genetics
8.1. Mineralization of Organic Phosphate: Molecular Genetics
8.2. Regulation of Inorganic Phosphate in Bacterial Cells
8.3. Interaction of PSB with the Plant Roots
9. PSB Co-Inoculation of Plants and PSB Synergy with the Use of Phosphate Fertilizers
Symbiotic Interactions between Phosphate-Solubilizing and Nitrogen-Fixing Microorganisms
10. Perspectives
11. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Maharajan, T.; Ceasar, S.A.; Krishna, T.P.A.; Ramakrishnan, M.; Duraipandiyan, V.; Abdulla, A.-D.N.; Ignacimuthu, S. Utilization of molecular markers for improving the phosphorus efficiency in crop plants. Plant Breed. 2018, 137, 10–26. [Google Scholar] [CrossRef]
- Santana, E.B.; Marques, E.L.S.; Dias, J.C.T. Effects of phosphate-solubilizing bacteria, native microorganisms, and rock dust on Jatropha curcas L. growth. Genet. Mol. Res. 2016, 15, 1–18. [Google Scholar] [CrossRef]
- Sharma, S.B.; Sayyed, R.Z.; Trivedi, M.H.; Gobi, T.A. Phosphate solubilizing microbes: Sustainable approach for managing phosphorus deficiency in agricultural soils. Springerplus 2013, 2, 587. [Google Scholar] [CrossRef]
- Nesme, T.; Metson, G.S.; Bennett, E.M. Global phosphorus flows through agricultural trade. Glob. Environ. Chang. 2018, 50, 133–141. [Google Scholar] [CrossRef]
- Zhu, F.; Qu, L.; Hong, X.; Sun, X. Isolation and Characterization of a Phosphate-Solubilizing Halophilic Bacterium Kushneriasp. YCWA18 from Daqiao Saltern on the Coast of Yellow Sea of China. Evid. Based Complementary Altern. Med. 2011, 2011, 615032. [Google Scholar] [CrossRef]
- Ai, P.; Sun, S.; Zhao, J.; Fan, X.; Xin, W.; Guo, Q.; Yu, L.; Shen, Q.; Wu, P.; Miller, A.J.; et al. Two rice phosphate transporters, OsPht1;2 and OsPht1;6, have different functions and kinetic properties in uptake and translocation. Plant J. 2009, 57, 798–809. [Google Scholar] [CrossRef] [PubMed]
- Azziz, G.; Bajsa, N.; Haghjou, T.; Taulé, C.; Valverde, Á.; Igual, J.M.; Arias, A. Abundance, diversity and prospecting of culturable phosphate solubilizing bacteria on soils under crop–pasture rotations in a no-tillage regime in Uruguay. Appl. Soil Ecol. 2012, 61, 320–326. [Google Scholar] [CrossRef]
- Schnug, E.; Haneklaus, S.H. The Enigma of Fertilizer Phosphorus Utilization. In Phosphorus in Agriculture: 100% Zero; Springer: Dordrecht, The Netherlands, 2016; pp. 7–26. [Google Scholar]
- Chaney, R.L. Food Safety Issues for Mineral and Organic Fertilizers. Adv. Agron. 2012, 117, 51–116. [Google Scholar] [CrossRef]
- Bhattacharyya, P.N.; Jha, D.K. Plant growth-promoting rhizobacteria (PGPR): Emergence in agriculture. World J. Microbiol. Biotechnol. 2012, 28, 1327–1350. [Google Scholar] [CrossRef] [PubMed]
- Khan, M.S.; Zaidi, A.; Wani, P.A. Role of phosphate-solubilizing microorganisms in sustainable agriculture—A review. Agron. Sustain. Dev. 2007, 27, 29–43. [Google Scholar] [CrossRef]
- Owen, D.; Williams, A.P.; Griffith, G.W.; Withers, P.J.A. Use of commercial bio-inoculants to increase agricultural production through improved phosphrous acquisition. Appl. Soil Ecol. 2015, 86, 41–54. [Google Scholar] [CrossRef]
- Wani, P.; Khan, M.; Zaidi, A. Co-inoculation of nitrogen-fixing and phosphate-solubilizing bacteria to promote growth, yield and nutrient uptake in chickpea. Acta Agron. Hung. 2007, 55, 315–323. [Google Scholar] [CrossRef]
- Wang, H.-Y.; Liu, S.; Zhai, L.-M.; Zhang, J.-Z.; Ren, T.-Z.; Fan, B.-Q.; Liu, H.-B. Preparation and utilization of phosphate biofertilizers using agricultural waste. J. Integr. Agric. 2015, 14, 158–167. [Google Scholar] [CrossRef]
- Alori, E.; Fawole, O.; Afolayan, A. Characterization of Arbuscular Mycorrhizal Spores Isolated from Southern Guinea Savanna of Nigeria. J. Agric. Sci. 2012, 4, 13. [Google Scholar] [CrossRef]
- Delavaux, C.S.; Smith-Ramesh, L.M.; Kuebbing, S.E. Beyond nutrients: A meta-analysis of the diverse effects of arbuscular mycorrhizal fungi on plants and soils. Ecology 2017, 98, 2111–2119. [Google Scholar] [CrossRef]
- Browne, P.; Rice, O.; Miller, S.H.; Burke, J.; Dowling, D.N.; Morrissey, J.P.; O’Gara, F. Superior inorganic phosphate solubilization is linked to phylogeny within the Pseudomonas fluorescens complex. Appl. Soil Ecol. 2009, 43, 131–138. [Google Scholar] [CrossRef]
- Rodríguez, H.; Fraga, R. Phosphate solubilizing bacteria and their role in plant growth promotion. Biotechnol. Adv. 1999, 17, 319–339. [Google Scholar] [CrossRef]
- Jha, A.; Sharma, D.; Saxena, J. Effect of single and dual phosphate-solubilizing bacterial strain inoculations on overall growth of mung bean plants. Arch. Agron. Soil Sci. 2012, 58, 967–981. [Google Scholar] [CrossRef]
- Srinivasan, R.; Yandigeri, M.S.; Kashyap, S.; Alagawadi, A.R. Effect of salt on survival and P-solubilization potential of phosphate solubilizing microorganisms from salt affected soils. Saudi J. Biol. Sci. 2012, 19, 427–434. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Bauddh, K.; Barman, S.C.; Singh, R.P. Amendments of microbial biofertilizers and organic substances reduces requirement of urea and DAP with enhanced nutrient availability and productivity of wheat (Triticum aestivum L.). Ecol. Eng. 2014, 71, 432–437. [Google Scholar] [CrossRef]
- Jiang, Z.; Zhang, X.; Wang, Z.; Cao, B.; Deng, S.; Bi, M.; Zhang, Y. Enhanced biodegradation of atrazine by Arthrobacter sp. DNS10 during co-culture with a phosphorus solubilizing bacteria: Enterobacter sp. P1. Ecotoxicol. Environ. Saf. 2019, 172, 159–166. [Google Scholar] [CrossRef] [PubMed]
- Jahan, M.; Mahallati, M.N.; Amiri, M.B.; Ehyayi, H.R. Radiation absorption and use efficiency of sesame as affected by biofertilizers inoculation in a low input cropping system. Ind. Crop. Prod. 2013, 43, 606–611. [Google Scholar] [CrossRef]
- Zhao, L.; Zhang, Y.-Q. Effects of phosphate solubilization and phytohormone production of Trichoderma asperellum Q1 on promoting cucumber growth under salt stress. J. Integr. Agric. 2015, 14, 1588–1597. [Google Scholar] [CrossRef]
- Postma, J.; Nijhuis, E.H.; Someus, E. Selection of phosphorus solubilizing bacteria with biocontrol potential for growth in phosphorus rich animal bone charcoal. Appl. Soil Ecol. 2010, 46, 464–469. [Google Scholar] [CrossRef]
- Mamta; Rahi, P.; Pathania, V.; Gulati, A.; Singh, B.; Bhanwra, R.K.; Tewari, R. Stimulatory effect of phosphate-solubilizing bacteria on plant growth, stevioside and rebaudioside-A contents of Stevia rebaudiana Bertoni. Appl. Soil Ecol. 2010, 46, 222–229. [Google Scholar] [CrossRef]
- Chakraborty, U.; Chakraborty, B.N.; Basnet, M.; Chakraborty, A.P. Evaluation of Ochrobactrum anthropic TRS-2 and its talc based formulation for enhancement of growth of tea plants and management of brown root rot disease. J. Appl. Microbiol. 2009, 107, 625–634. [Google Scholar] [CrossRef]
- Dastager, S.G.; Deepa, C.K.; Pandey, A. Isolation and characterization of novel plant growth promoting Micrococcus sp. NII-0909 and its interaction with cowpea. Plant Physiol. Biochem. 2010, 48, 987–992. [Google Scholar] [CrossRef]
- Bidondo, L.F.; Silvani, V.; Colombo, R.; Pérgola, M.; Bompadre, J.; Godeas, A. Pre-symbiotic and symbiotic interactions between Glomus intraradices and two Paenibacillus species isolated from AM propagules. In Vitro and in vivo assays with soybean (AG043RG) as plant host. Soil Biol. Biochem. 2011, 43, 1866–1872. [Google Scholar] [CrossRef]
- Oteino, N.; Lally, R.D.; Kiwanuka, S.; Lloyd, A.; Ryan, D.; Germaine, K.J.; Dowling, D.N. Plant growth promotion induced by phosphate solubilizing endophytic Pseudomonas isolates. Front. Microbiol. 2015, 6, 745. [Google Scholar] [CrossRef]
- Paul, D.; Sinha, S.N. Isolation and characterization of phosphate solubilizing bacterium Pseudomonas aeruginosa KUPSB12 with antibacterial potential from river Ganga, India. Ann. Agrar. Sci. 2017, 15, 130–136. [Google Scholar] [CrossRef]
- Tajini, F.; Trabelsi, M.; Drevon, J.-J. Combined inoculation with Glomus intraradices and Rhizobium tropici CIAT899 increases phosphorus use efficiency for symbiotic nitrogen fixation in common bean (Phaseolus vulgaris L.). Saudi J. Biol. Sci. 2012, 19, 157–163. [Google Scholar] [CrossRef] [PubMed]
- Korir, H.; Mungai, N.W.; Thuita, M.; Hamba, Y.; Masso, C.; Tola, Y.H. Co-inoculation Effect of Rhizobia and Plant Growth Promoting Rhizobacteria on Common Bean Growth in a Low Phosphorus Soil. Front. Plant Sci. 2017, 8, 141. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Bai, T.; Dai, L.; Wang, F.; Tao, J.; Meng, S.; Hu, Y.; Wang, S.; Hu, S. A study of organic acid production in contrasts between two phosphate solubilizing fungi: Penicillium oxalicum and Aspergillus niger. Sci. Rep. 2016, 6, 25313. [Google Scholar] [CrossRef]
- Behera, B.C.; Yadav, H.; Singh, S.K.; Mishra, R.R.; Sethi, B.K.; Dutta, S.K.; Thatoi, H.N. Phosphate solubilization and acid phosphatase activity of Serratia sp. isolated from mangrove soil of Mahanadi river delta, Odisha, India. J. Genet. Eng. Biotechnol. 2017, 15, 169–178. [Google Scholar] [CrossRef] [PubMed]
- Thampi, A.; Bhai, R.S. Rhizosphere actinobacteria for combating Phytophthora capsici and Sclerotium rolfsii, the major soil borne pathogens of black pepper (Piper nigrum L.). Biol. Control 2017, 109, 1–13. [Google Scholar] [CrossRef]
- Kalayu, G. Phosphate Solubilizing Microorganisms: Promising Approach as Biofertilizers. Int. J. Agron. 2019, 2019, 4917256. [Google Scholar] [CrossRef]
- Arai, Y.; Sparks, D. Phosphate Reaction Dynamics in Soils and Soil Components: A Multiscale Approach. Adv. Agron. 2007, 94, 135–179. [Google Scholar] [CrossRef]
- Devau, N.; Le Cadre, E.; Hinsinger, P.; Jaillard, B.; Gérard, F. Soil pH controls the environmental availability of phosphorus: Experimental and mechanistic modelling approaches. Appl. Geochem. 2009, 24, 2163–2174. [Google Scholar] [CrossRef]
- Mnkeni, P.N.S.; MacKenzie, A.F. Retention of Ortho- and Polyphosphates in Some Quebec Soils as Affected by Added Organic Residues and Calcium Carbonate. Can. J. Soil Sci. 1985, 65, 575–585. [Google Scholar] [CrossRef]
- Hallama, M.; Pekrun, C.; Lambers, H.; Kandeler, E. Hidden miners—The roles of cover crops and soil microorganisms in phosphorus cycling through agroecosystems. Plant Soil 2019, 434, 7–45. [Google Scholar] [CrossRef]
- Ehlers, K.; Bakken, L.R.; Frostegård, Å.; Frossard, E.; Bünemann, E.K. Phosphorus limitation in a Ferralsol: Impact on microbial activity and cell internal P pools. Soil Biol. Biochem. 2010, 42, 558–566. [Google Scholar] [CrossRef]
- Richardson, A.E.; Simpson, R.J. Soil Microorganisms Mediating Phosphorus Availability Update on Microbial Phosphorus. Plant Physiol. 2011, 156, 989–996. [Google Scholar] [CrossRef] [PubMed]
- Kulaev, I.S.; Vagabov, V.M. Polyphosphate Metabolism in Micro-Organisms. Adv. Microb. Physiol. 1983, 24, 83–171. [Google Scholar] [CrossRef] [PubMed]
- Hirota, R.; Kuroda, A.; Kato, J.; Ohtake, H. Bacterial phosphate metabolism and its application to phosphorus recovery and industrial bioprocesses. J. Biosci. Bioeng. 2010, 109, 423–432. [Google Scholar] [CrossRef] [PubMed]
- Kuroda, A.; Murphy, H.; Cashel, M.; Kornberg, A. Guanosine Tetra- and Pentaphosphate Promote Accumulation of Inorganic Polyphosphate in Escherichia coli. J. Biol. Chem. 1997, 272, 21240–21243. [Google Scholar] [CrossRef]
- Moradali, M.F.; Rehm, B.H.A. Bacterial biopolymers: From pathogenesis to advanced materials. Nat. Rev. Microbiol. 2020, 18, 195–210. [Google Scholar] [CrossRef]
- Karl, D.M. Microbially Mediated Transformations of Phosphorus in the Sea: New Views of an Old Cycle. Annu. Rev. Mar. Sci. 2014, 6, 279–337. [Google Scholar] [CrossRef]
- Seeling, B.; Zasoski, R.J. Microbial effects in maintaining organic and inorganic solution phosphorus concentrations in a grassland topsoil. Plant Soil 1993, 148, 277–284. [Google Scholar] [CrossRef]
- Khan, K.S.; Joergensen, R.G. Changes in microbial biomass and P fractions in biogenic household waste compost amended with inorganic P fertilizers. Bioresour. Technol. 2009, 100, 303–309. [Google Scholar] [CrossRef]
- Sugito, T.; Yoshida, K.; Takebe, M.; Shinano, T.; Toyota, K. Soil microbial biomass phosphorus as an indicator of phosphorus availability in a Gleyic Andosol. Soil Sci. Plant Nutr. 2010, 56, 390–398. [Google Scholar] [CrossRef]
- Spohn, M.; Widdig, M. Turnover of carbon and phosphorus in the microbial biomass depending on phosphorus availability. Soil Biol. Biochem. 2017, 113, 53–59. [Google Scholar] [CrossRef]
- Malik, M.A. Phosphorus fractions, microbial biomass and enzyme activities in some alkaline calcareous subtropical soils. Afr. J. Biotechnol. 2012, 11, 4773–4781. [Google Scholar] [CrossRef]
- Chen, G.C.; He, Z.L. Microbial biomass phosphorus turnover in variable-charge soils in China. Commun. Soil Sci. Plant Anal. 2002, 33, 2101–2117. [Google Scholar] [CrossRef]
- Dinh, M.-V.; Guhr, A.; Spohn, M.; Matzner, E. Release of phosphorus from soil bacterial and fungal biomass following drying/rewetting. Soil Biol. Biochem. 2017, 110, 1–7. [Google Scholar] [CrossRef]
- Patel, D.K.; Archana, G.; Kumar, G.N. Variation in the Nature of Organic Acid Secretion and Mineral Phosphate Solubilization by Citrobacter sp. DHRSS in the Presence of Different Sugars. Curr. Microbiol. 2008, 56, 168–174. [Google Scholar] [CrossRef]
- Vijayakumar, A.; Rajasekharan, R. Distinct Roles of Alpha/Beta Hydrolase Domain Containing Proteins. Biochem. Mol. Biol. J. 2016, 2, 1–3. [Google Scholar] [CrossRef]
- Son, H.-J.; Park, G.-T.; Cha, M.-S.; Heo, M.-S. Solubilization of insoluble inorganic phosphates by a novel salt- and pH-tolerant Pantoea agglomerans R-42 isolated from soybean rhizosphere. Bioresour. Technol. 2006, 97, 204–210. [Google Scholar] [CrossRef]
- Zhao, K.; Penttinen, P.; Zhang, X.; Ao, X.; Liu, M.; Yu, X.; Chen, Q. Maize rhizosphere in Sichuan, China, hosts plant growth promoting Burkholderia cepacia with phosphate solubilizing and antifungal abilities. Microbiol. Res. 2014, 169, 76–82. [Google Scholar] [CrossRef]
- Asea, P.; Kucey, R.; Stewart, J. Inorganic phosphate solubilization by two Penicillium species in solution culture and soil. Soil Biol. Biochem. 1988, 20, 459–464. [Google Scholar] [CrossRef]
- Illmer, P.; Schinner, F. Solubilization of inorganic calcium phosphates—Solubilization mechanisms. Soil Biol. Biochem. 1995, 27, 257–263. [Google Scholar] [CrossRef]
- Kouas, S.; Labidi, N.; Debez, A.; Abdelly, C. Effect of P on nodule formation and N fixation in bean. Agron. Sustain. Dev. 2005, 25, 389–393. [Google Scholar] [CrossRef]
- Taha, S.M.; Mahmoud, S.A.Z.; El-Damaty, A.H.; El-Hafez, A.M. Activity of phosphate-dissolving bacteria in Egyptian soils. Plant Soil 1969, 31, 149–160. [Google Scholar] [CrossRef]
- Bajpai, P.D.; Rao, W.V.B.S. Phosphate solubilising bacteria. Soil Sci. Plant Nutr. 1971, 17, 46–53. [Google Scholar] [CrossRef]
- Banik, S.; Dey, B.K. Phosphate-Solubilizing Potentiality of the Microorganisms Capable of Utilizing Aluminium Phosphate as a Sole Phosphate Source. Zentralbl. Mikrobiol. 1983, 138, 17–23. [Google Scholar] [CrossRef]
- Singal, R.; Gupta, R.; Saxena, R.K. Rock phosphate solubilization under alkaline conditions by Aspergillus japonicus and A. foetidus. Folia Microbiol. 1994, 39, 33–36. [Google Scholar] [CrossRef] [PubMed]
- Whitelaw, M.A.; Harden, T.J.; Helyar, K.R. Phosphate solubilisation in solution culture by the soil fungus Penicillium radicum. Soil Biol. Biochem. 1999, 31, 655–665. [Google Scholar] [CrossRef]
- Kim, K.Y.; McDonald, G.A.; Jordan, D. Solubilization of hydroxyapatite by Enterobacter agglomerans and cloned Escherichia coli in culture medium. Biol. Fertil. Soils 1997, 24, 347–352. [Google Scholar] [CrossRef]
- Vazquez, P.; Holguin, G.; Puente, M.E.; Lopez-Cortes, A.; Bashan, Y. Phosphate-solubilizing microorganisms associated with the rhizosphere of mangroves in a semiarid coastal lagoon. Biol. Fertil. Soils 2000, 30, 460–468. [Google Scholar] [CrossRef]
- Reyes, I.; Baziramakenga, R.; Bernier, L.; Antoun, H. Solubilization of phosphate rocks and minerals by a wild-type strain and two UV-induced mutants of Penicillium rugulosum. Soil Biol. Biochem. 2001, 33, 1741–1747. [Google Scholar] [CrossRef]
- Kim, K.Y.; Hwangbo, H.; Park, R.D.; Kim, Y.W.; Rim, Y.S.; Park, K.H.; Kim, T.H.; Suh, J.S. 2-Ketogluconic Acid Production and Phosphate Solubilization by Enterobacter intermedium. Curr. Microbiol. 2003, 47, 87–92. [Google Scholar] [CrossRef]
- Rashid, M.; Khalil, S.; Ayub, N.; Alam, S.; Latif, F. Organic Acids Production and Phosphate Solubilization by Phosphate Solubilizing Microorganisms (PSM) Under in vitro Conditions. Pak. J. Biol. Sci. 2004, 7, 187–196. [Google Scholar] [CrossRef]
- Fankem, H.; Nwaga, D.; Deube, A.; Dieng, L.; Merbach, W.; Etoa, F. Occurrence and functioning of phosphate solubilizing microorganisms from oil palm tree (Elaeis guineensis) rhizosphere in Cameroon. Afr. J. Biotechnol. 2006, 5, 2450–2460. [Google Scholar]
- Chuang, C.-C.; Kuo, Y.-L.; Chao, C.-C.; Chao, W.-L. Solubilization of inorganic phosphates and plant growth promotion by Aspergillus niger. Biol. Fertil. Soils 2007, 43, 575–584. [Google Scholar] [CrossRef]
- Vyas, P.; Gulati, A. Organic acid production in vitro and plant growth promotion in maize under controlled environment by phosphate-solubilizing fluorescent Pseudomonas. BMC Microbiol. 2009, 9, 174. [Google Scholar] [CrossRef] [PubMed]
- Puente, M.E.; Li, C.Y.; Bashan, Y. Microbial Populations and Activities in the Rhizoplane of Rock-Weathering Desert Plants. II. Growth Promotion of Cactus Seedlings. Plant Biol. 2004, 6, 643–650. [Google Scholar] [CrossRef]
- Puente, M.E.; Li, C.Y.; Bashan, Y. Rock-degrading endophytic bacteria in cacti. Environ. Exp. Bot. 2009, 66, 389–401. [Google Scholar] [CrossRef]
- Lopez, B.R.; Bashan, Y.; Bacilio, M. Endophytic bacteria of Mammillaria fraileana, an endemic rock-colonizing cactus of the southern Sonoran Desert. Arch. Microbiol. 2011, 193, 527–541. [Google Scholar] [CrossRef]
- Oubrie, A.; Rozeboom, H.J.; Kalk, K.H.; Olsthoorn, A.J.; Duine, J.A.; Dijkstra, B.W. Structure and mechanism of soluble quinoprotein glucose dehydrogenase. EMBO J. 1999, 18, 5187–5194. [Google Scholar] [CrossRef] [PubMed]
- Lessie, T.G.; Phibbs, P.V. Alternative Pathways of Carbohydrate Utilization in Pseudomonads. Annu. Rev. Microbiol. 1984, 38, 359–388. [Google Scholar] [CrossRef]
- Thomas, L.; Hodgson, D.A.; Wentzel, A.; Nieselt, K.; Ellingsen, T.E.; Moore, J.; Morrissey, E.R.; Legaie, R.; The STREAM Consortium; Wohlleben, W.; et al. Metabolic Switches and Adaptations Deduced from the Proteomes of Streptomyces coelicolor Wild Type and phoP Mutant Grown in Batch Culture. Mol. Cell. Proteom. 2012, 11, M111.013797. [Google Scholar] [CrossRef]
- Buch, A.; Archana, G.; Kumar, G.N. Metabolic channeling of glucose towards gluconate in phosphate-solubilizing Pseudomonas aeruginosa P4 under phosphorus deficiency. Res. Microbiol. 2008, 159, 635–642. [Google Scholar] [CrossRef] [PubMed]
- Zeng, Q.; Wu, X.; Wen, X. Effects of Soluble Phosphate on Phosphate-Solubilizing Characteristics and Expression of gcd Gene in Pseudomonas frederiksbergensis JW-SD2. Curr. Microbiol. 2016, 72, 198–206. [Google Scholar] [CrossRef]
- Ben Farhat, M.; Fourati, A.; Chouayekh, H. Coexpression of the Pyrroloquinoline Quinone and Glucose Dehydrogenase Genes from Serratia marcescens CTM 50650 Conferred High Mineral Phosphate-Solubilizing Ability to Escherichia coli. Appl. Biochem. Biotechnol. 2013, 170, 1738–1750. [Google Scholar] [CrossRef]
- Liu, S.T.; Lee, L.Y.; Tai, C.Y.; Hung, C.-H.; Chang, Y.S.; Wolfram, J.H.; Rogers, R.; Goldstein, A.H. Cloning of an Erwinia herbicola gene necessary for gluconic acid production and enhanced mineral phosphate solubilization in Escherichia coli HB101: Nucleotide sequence and probable involvement in biosynthesis of the coenzyme pyrroloquinoline quinone. J. Bacteriol. 1992, 174, 5814–5819. [Google Scholar] [CrossRef][Green Version]
- Zeng, Q.; Wu, X.; Wang, J.; Ding, X. Phosphate Solubilization and Gene Expression of Phosphate-Solubilizing Bacterium Burkholderia multivorans WS-FJ9 under Different Levels of Soluble Phosphate. J. Microbiol. Biotechnol. 2017, 27, 844–855. [Google Scholar] [CrossRef] [PubMed]
- Peix, A.; Mateos, P.F.; Rodriguez-Barrueco, C.; Martínez-Molina, E.; Velazquez, E. Growth promotion of common bean (Phaseolus vulgaris L.) by a strain of Burkholderia cepacia under growth chamber conditions. Soil Biol. Biochem. 2001, 33, 1927–1935. [Google Scholar] [CrossRef]
- Nannipieri, P.; Giagnoni, L.; Landi, L.; Renella, G. Role of Phosphatase Enzymes in Soil. In Phosphorus in Action; Bünemann, E.K., Oberson, A., Frossard, E., Eds.; Springer: Berlin/Heidelberg, Germany, 2011; pp. 215–243. [Google Scholar] [CrossRef]
- Jorquera, M.A.; Crowley, D.E.; Marschner, P.; Greiner, R.; Fernández, M.T.; Romero, D.; Menezes-Blackburn, D.; De La Luz Mora, M. Identification of β-propeller phytase-encoding genes in culturable Paenibacillus and Bacillus spp. from the rhizosphere of pasture plants on volcanic soils. FEMS Microbiol. Ecol. 2011, 75, 163–172. [Google Scholar] [CrossRef] [PubMed]
- Lim, B.L.; Yeung, P.; Cheng, T.; Hill, J.E. Distribution and diversity of phytate-mineralizing bacteria. ISME J. 2007, 1, 321–330. [Google Scholar] [CrossRef]
- Turner, B.L.; Papházy, M.J.; Haygarth, P.M.; McKelvie, I.D. Inositol phosphates in the environment. Philos. Trans. R. Soc. London. Ser. B Biol. Sci. 2002, 357, 449–469. [Google Scholar] [CrossRef]
- Rao, D.E.C.S.; Rao, K.V.; Reddy, T.P.; Reddy, V.D. Molecular characterization, physicochemical properties, known and potential applications of phytases: An overview. Crit. Rev. Biotechnol. 2009, 29, 182–198. [Google Scholar] [CrossRef]
- Huang, H.; Shi, P.; Wang, Y.; Luo, H.; Shao, N.; Wang, G.; Yang, P.; Yao, B. Diversity of Beta-Propeller Phytase Genes in the Intestinal Contents of Grass Carp Provides Insight into the Release of Major Phosphorus from Phytate in Nature. Appl. Environ. Microbiol. 2009, 75, 1508–1516. [Google Scholar] [CrossRef] [PubMed]
- Kim, O.-H.; Kim, Y.-O.; Shim, J.-H.; Jung, Y.-S.; Jung, W.-J.; Choi, W.-C.; Lee, H.; Lee, S.-J.; Kim, K.-K.; Auh, J.-H.; et al. β-Propeller Phytase Hydrolyzes Insoluble Ca2+-Phytate Salts and Completely Abrogates the Ability of Phytate to Chelate Metal Ions. Biochemistry 2010, 49, 10216–10227. [Google Scholar] [CrossRef]
- Fu, S.; Sun, J.; Qian, L. Effect of Ca2+ on beta-propeller phytases. Protein Pept. Lett. 2008, 15, 39–42. [Google Scholar] [CrossRef]
- Cheng, C.; Lim, B.L. Beta-propeller phytases in the aquatic environment. Arch. Microbiol. 2006, 185, 1–13. [Google Scholar] [CrossRef]
- Kerovuo, J.; Lauraeus, M.; Nurminen, P.; Kalkkinen, N.; Apajalahti, J. Isolation, Characterization, Molecular Gene Cloning, and Sequencing of a Novel Phytase from Bacillus subtilis. Appl. Environ. Microbiol. 1998, 64, 2079–2085. [Google Scholar] [CrossRef]
- Kim, Y.-O.; Lee, J.-K.; Kim, H.-K.; Yu, J.-H.; Oh, T.-K. Cloning of the thermostable phytase gene (phy) from Bacillus sp. DS11 and its overexpression in Escherichia coli. FEMS Microbiol. Lett. 1998, 162, 185–191. [Google Scholar] [CrossRef] [PubMed]
- Tye, A.; Siu, F.; Leung, T.; Lim, B. Molecular cloning and the biochemical characterization of two novel phytases from B. subtilis 168 and B. licheniformis. Appl. Microbiol. Biotechnol. 2002, 59, 190–197. [Google Scholar] [CrossRef] [PubMed]
- Huang, H.; Shao, N.; Wang, Y.; Luo, H.; Yang, P.; Zhou, Z.; Zhan, Z.; Yao, B. A novel beta-propeller phytase from Pedobacter nyackensis MJ11 CGMCC 2503 with potential as an aquatic feed additive. Appl. Microbiol. Biotechnol. 2009, 83, 249–259. [Google Scholar] [CrossRef]
- Zhang, R.; Yang, P.; Huang, H.; Yuan, T.; Shi, P.; Meng, K.; Yao, B. Molecular and biochemical characterization of a new alkaline β-propeller phytase from the insect symbiotic bacterium Janthinobacterium sp. TN115. Appl. Microbiol. Biotechnol. 2011, 92, 317–325. [Google Scholar] [CrossRef]
- Jorquera, M.A.; Hernández, M.T.; Rengel, Z.; Marschner, P.; de la Luz Mora, M. Isolation of culturable phosphobacteria with both phytate-mineralization and phosphate-solubilization activity from the rhizosphere of plants grown in a volcanic soil. Biol. Fertil. Soils 2008, 44, 1025–1034. [Google Scholar] [CrossRef]
- Rodríguez, H.; Rossolini, G.M.; Gonzalez, T.; Li, J.P.; Glick, B.R. Isolation of a Gene from Burkholderia cepacia IS-16 Encoding a Protein That Facilitates Phosphatase Activity. Curr. Microbiol. 2000, 40, 362–366. [Google Scholar] [CrossRef] [PubMed]
- Thaller, M.C.; Berlutti, F.; Schippa, S.; Iori, P.; Passariello, C.; Rossolini, G.M. Heterogeneous Patterns of Acid Phosphatases Containing Low-Molecular-Mass Polypeptides in Members of the Family Enterobacteriaceae. Int. J. Syst. Bacteriol. 1995, 45, 255–261. [Google Scholar] [CrossRef]
- Richardson, A.E.; Hadobas, P.A. Soil isolates of Pseudomonas spp. that utilize inositol phosphates. Can. J. Microbiol. 1997, 43, 509–516. [Google Scholar] [CrossRef] [PubMed]
- Ohtake, H.; Wu, H.; Imazu, K.; Anbe, Y.; Kato, J.; Kuroda, A. Bacterial phosphonate degradation, phosphite oxidation and polyphosphate accumulation. Resour. Conserv. Recycl. 1996, 18, 125–134. [Google Scholar] [CrossRef]
- Lidbury, I.D.E.A.; Murphy, A.R.J.; Scanlan, D.J.; Bending, G.D.; Jones, A.M.E.; Moore, J.D.; Goodall, A.; Hammond, J.P.; Wellington, E.M.H. Comparative genomic, proteomic and exoproteomic analyses of three Pseudomonas strains reveals novel insights into the phosphorus scavenging capabilities of soil bacteria. Environ. Microbiol. 2016, 18, 3535–3549. [Google Scholar] [CrossRef]
- Brito, L.F.; Gil López, M.; Straube, L.; Passaglia, L.M.P.; Wendisch, V.F. Inorganic Phosphate Solubilization by Rhizosphere Bacterium Paenibacillus sonchi: Gene Expression and Physiological Functions. Front. Microbiol. 2020, 11. [Google Scholar] [CrossRef] [PubMed]
- Heuck, C.; Smolka, G.; Whalen, E.D.; Frey, S.; Gundersen, P.; Moldan, F.; Fernandez, I.J.; Spohn, M. Effects of long-term nitrogen addition on phosphorus cycling in organic soil horizons of temperate forests. Biogeochemistry 2018, 141, 167–181. [Google Scholar] [CrossRef]
- Marklein, A.R.; Houlton, B.Z. Nitrogen inputs accelerate phosphorus cycling rates across a wide variety of terrestrial ecosystems. New Phytol. 2012, 193, 696–704. [Google Scholar] [CrossRef] [PubMed]
- Contesto, C.; Desbrosses, G.; Lefoulon, C.; Bena, G.; Borel, F.; Galland, M.; Gamet, L.; Varoquaux, F.; Touraine, B. Effects of rhizobacterial ACC deaminase activity on Arabidopsis indicate that ethylene mediates local root responses to plant growth-promoting rhizobacteria. Plant Sci. 2008, 175, 178–189. [Google Scholar] [CrossRef]
- Haidar, B.; Ferdous, M.; Fatema, B.; Ferdous, A.S.; Islam, M.R.; Khan, H. Population diversity of bacterial endophytes from jute (Corchorus olitorius) and evaluation of their potential role as bioinoculants. Microbiol. Res. 2018, 208, 43–53. [Google Scholar] [CrossRef]
- Kudoyarova, G.R.; Vysotskaya, L.B.; Arkhipova, T.N.; Kuzmina, L.Y.; Galimsyanova, N.F.; Sidorova, L.V.; Gabbasova, I.M.; Melentiev, A.I.; Veselov, S.Y. Effect of auxin producing and phosphate solubilizing bacteria on mobility of soil phosphorus, growth rate, and P acquisition by wheat plants. Acta Physiol. Plant. 2017, 39, 253. [Google Scholar] [CrossRef]
- Ortíz-Castro, R.; Contreras-Cornejo, H.A.; Macías-Rodríguez, L.; López-Bucio, J. The role of microbial signals in plant growth and development. Plant Signal. Behav. 2009, 4, 701–712. [Google Scholar] [CrossRef]
- Li, Y.; Liu, X.; Hao, T.; Chen, S. Colonization and Maize Growth Promotion Induced by Phosphate Solubilizing Bacterial Isolates. Int. J. Mol. Sci. 2017, 18, 1253. [Google Scholar] [CrossRef]
- Saraf, M.; Jha, C.K.; Patel, D. The Role of ACC Deaminase Producing PGPR in Sustainable Agriculture. In Plant Growth and Health Promoting Bacteria; Springer: Berlin/Heidelberg, Germany, 2010; pp. 365–385. [Google Scholar] [CrossRef]
- Bal, H.B.; Nayak, L.; Das, S.; Adhya, T.K. Isolation of ACC deaminase producing PGPR from rice rhizosphere and evaluating their plant growth promoting activity under salt stress. Plant Soil 2012, 366, 93–105. [Google Scholar] [CrossRef]
- White, C.; Sayer, J.; Gadd, G. Microbial solubilization and immobilization of toxic metals: Key biogeochemical processes for treatment of contamination. FEMS Microbiol. Rev. 1997, 20, 503–516. [Google Scholar] [CrossRef]
- Kang, J.; Amoozegar, A.; Hesterberg, D.; Osmond, D. Phosphorus leaching in a sandy soil as affected by organic and inorganic fertilizer sources. Geoderma 2011, 161, 194–201. [Google Scholar] [CrossRef]
- Rosado, A.S.; De Azevedo, F.S.; Da Cruz, D.W.; Van Elsas, J.D.; Seldin, L. Seldin Phenotypic and genetic diversity of Paenibacillus azotofixans strains isolated from the rhizoplane or rhizosphere soil of different grasses. J. Appl. Microbiol. 1998, 84, 216–226. [Google Scholar] [CrossRef]
- Nahas, E. Factors determining rock phosphate solubilization by microorganisms isolated from soil. World J. Microbiol. Biotechnol. 1996, 12, 567–572. [Google Scholar] [CrossRef]
- Nautiyal, C.S.; Bhadauria, S.; Kumar, P.; Lal, H.; Mondal, R.; Verma, D. Stress induced phosphate solubilization in bacteria isolated from alkaline soils. FEMS Microbiol. Lett. 2000, 182, 291–296. [Google Scholar] [CrossRef]
- Johri, J.K.; Surange, S.; Nautiyal, C.S. Occurrence of Salt, pH, and Temperature-tolerant, Phosphate-solubilizing Bacteria in Alkaline Soils. Curr. Microbiol. 1999, 39, 89–93. [Google Scholar] [CrossRef]
- Seshachala, U.; Tallapragada, P. Phosphate Solubilizers from the Rhizospher of Piper nigrum L. in Karnataka, India. Chil. J. Agric. Res. 2012, 72, 397–403. [Google Scholar] [CrossRef]
- Zhang, L.; Ding, X.; Chen, S.; He, X.; Zhang, F.; Feng, G. Reducing carbon: Phosphorus ratio can enhance microbial phytin mineralization and lessen competition with maize for phosphorus. J. Plant Interactions 2014, 9, 850–856. [Google Scholar] [CrossRef]
- Babu-Khan, S.; Yeo, T.C.; Martin, W.L.; Duron, M.R.; Rogers, R.D.; Goldstein, A.H. Cloning of a mineral phosphate-solubilizing gene from Pseudomonas cepacia. Appl. Environ. Microbiol. 1995, 61, 972–978. [Google Scholar] [CrossRef] [PubMed]
- Van Schie, B.J.; Hellingwerf, K.J.; van Dijken, J.P.; Elferink, M.G.; van Dijl, J.M.; Kuenen, J.G.; Konings, W.N. Energy transduction by electron transfer via a pyrrolo-quinoline quinone-dependent glucose dehydrogenase in Escherichia coli, Pseudomonas aeruginosa, and Acinetobacter calcoaceticus (var. lwoffi). J. Bacteriol. 1985, 163, 493–499. [Google Scholar] [CrossRef]
- Xie, J.; Shi, H.; Du, Z.; Wang, T.; Liu, X.; Chen, S. Comparative genomic and functional analysis reveal conservation of plant growth promoting traits in Paenibacillus polymyxa and its closely related species. Sci. Rep. 2016, 6, 21329. [Google Scholar] [CrossRef]
- Grady, E.N.; Macdonald, J.; Liu, L.; Richman, A.; Yuan, Z.-C. Current knowledge and perspectives of Paenibacillus: A review. Microb. Cell Factories 2016, 15, 203. [Google Scholar] [CrossRef]
- Rosenberg, H. Phosphate Transport in Prokaryotes. In Ion Transport in Prokaryotes; Elsevier: Amsterdam, The Netherlands, 1987; pp. 205–248. [Google Scholar]
- Thaller, M.C.; Berlutti, F.; Schippa, S.; Lombardi, G.; Rossolini, G.M. Characterization and sequence of PhoC, the principal phosphate-irrepressible acid phosphatase of Morganella morganii. Microbiology 1994, 140, 1341–1350. [Google Scholar] [CrossRef]
- Pond, J.L.; Eddy, C.K.; Mackenzie, K.F.; Conway, T.; Borecky, D.J.; Ingram, L.O. Cloning, sequencing, and characterization of the principal acid phosphatase, the phoC+ product, from Zymomonas mobilis. J. Bacteriol. 1989, 171, 767–774. [Google Scholar] [CrossRef]
- Kier, L.D.; Weppelman, R.; Ames, B.N. Resolution and purification of three periplasmic phosphatases of Salmonella typhimurium. J. Bacteriol. 1977, 130, 399–410. [Google Scholar] [CrossRef]
- Miller, S.I.; Kukral, A.M.; Mekalanos, J.J. A two-component regulatory system (phoP phoQ) controls Salmonella typhimurium virulence. Proc. Natl. Acad. Sci. USA 1989, 86, 5054–5058. [Google Scholar] [CrossRef]
- Burini, J.-F.; Gügi, B.; Merieau, A.; Guespin-Michel, J.F. Lipase and acidic phosphatase from the psychrotrophic bacterium Pseudomonas fluorescens: Two enzymes whose synthesis is regulated by the growth temperature. FEMS Microbiol. Lett. 1994, 122, 13–18. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez, H.; Fraga, R.; Gonzalez, T.; Bashan, Y. Genetics of phosphate solubilization and its potential applications for improving plant growth-promoting bacteria. Plant Soil 2006, 287, 15–21. [Google Scholar] [CrossRef]
- Alori, E.T.; Glick, B.R.; Babalola, O.O. Microbial Phosphorus Solubilization and Its Potential for Use in Sustainable Agriculture. Front. Microbiol. 2017, 8, 971. [Google Scholar] [CrossRef]
- Passariello, C.; Schippa, S.; Iori, P.; Berlutti, F.; Thaller, M.C.; Rossolini, G.M. The molecular class C acid phosphatase of Chryseobacterium meningosepticum (OlpA) is a broad-spectrum nucleotidase with preferential activity on 5′-nucleotides. Biochim. Biophys. Acta—Proteins Proteom. 2003, 1648, 203–209. [Google Scholar] [CrossRef]
- Jiang, L.; Liu, X.; Yin, H.; Liang, Y.; Liu, H.; Miao, B.; Peng, Q.; Meng, D.; Wang, S.; Yang, J.; et al. The utilization of biomineralization technique based on microbial induced phosphate precipitation in remediation of potentially toxic ions contaminated soil: A mini review. Ecotoxicol. Environ. Saf. 2020, 191, 110009. [Google Scholar] [CrossRef]
- Xie, E.; Su, Y.; Deng, S.; Kontopyrgou, M.; Zhang, D. Significant influence of phosphorus resources on the growth and alkaline phosphatase activities of Microcystis aeruginosa. Environ. Pollut. 2020, 268, 115807. [Google Scholar] [CrossRef] [PubMed]
- Reher, M.; Bott, M.; Schönheit, P. Characterization of glycerate kinase (2-phosphoglycerate forming), a key enzyme of the nonphosphorylative Entner-Doudoroff pathway, from the thermoacidophilic euryarchaeon Picrophilus torridus. FEMS Microbiol. Lett. 2006, 259, 113–119. [Google Scholar] [CrossRef] [PubMed]
- Conway, T. The Entner-Doudoroff pathway: History, physiology and molecular biology. FEMS Microbiol. Lett. 1992, 103, 1–28. [Google Scholar] [CrossRef] [PubMed]
- Basu, A.; Phale, P.S. Inducible uptake and metabolism of glucose by the phosphorylative pathway in Pseudomonas putida CSV86. FEMS Microbiol. Lett. 2006, 259, 311–316. [Google Scholar] [CrossRef] [PubMed]
- Elferink, M.G.L.; Albers, S.-V.; Konings, W.N.; Driessen, A.J.M. Sugar transport in Sulfolobus solfataricus is mediated by two families of binding protein-dependent ABC transporters. Mol. Microbiol. 2001, 39, 1494–1503. [Google Scholar] [CrossRef]
- Lamarche, M.G.; Wanner, B.L.; Crépin, S.; Harel, J. The phosphate regulon and bacterial virulence: A regulatory network connecting phosphate homeostasis and pathogenesis. FEMS Microbiol. Rev. 2008, 32, 461–473. [Google Scholar] [CrossRef]
- Hulett, F.M.; Lee, J.; Shi, L.; Sun, G.; Chesnut, R.; Sharkova, E.; Duggan, M.F.; Kapp, N. Sequential action of two-component genetic switches regulates the PHO regulon in Bacillus subtilis. J. Bacteriol. 1994, 176, 1348–1358. [Google Scholar] [CrossRef]
- Eder, S.; Shi, L.; Jensen, K.; Yamane, K.; Hulett, F.M. A Bacillus subtilis secreted phosphodiesterase/alkaline phosphatase is the product of a Pho regulon gene, phoD. Microbiology 1996, 142, 2041–2047. [Google Scholar] [CrossRef] [PubMed]
- Maddocks, S.E.; Oyston, P.C.F. Structure and function of the LysR-type transcriptional regulator (LTTR) family proteins. Microbiology 2008, 154, 3609–3623. [Google Scholar] [CrossRef] [PubMed]
- Heller, K.J.; Kadner, R.J.; Günther, K. Suppression of the btuB451 mutation by mutations in the tonB gene suggests a direct interaction between TonB and TonB-dependent receptor proteins in the outer membrane of Escherichia coli. Gene 1988, 64, 147–153. [Google Scholar] [CrossRef]
- Braun, V.; Mahren, S.; Ogierman, M. Regulation of the FecI-type ECF sigma factor by transmembrane signalling. Curr. Opin. Microbiol. 2003, 6, 173–180. [Google Scholar] [CrossRef]
- Wanner, B.L.; Chang, B.D. The phoBR operon in Escherichia coli K-12. J. Bacteriol. 1987, 169, 5569–5574. [Google Scholar] [CrossRef]
- Santos-Beneit, F.; Rodríguez-García, A.; Franco-Domínguez, E.; Martín, J.F. Phosphate-dependent regulation of the low- and high-affinity transport systems in the model actinomycete Streptomyces coelicolor. Microbiology 2008, 154, 2356–2370. [Google Scholar] [CrossRef] [PubMed]
- Ghorbel, S.; Smirnov, A.; Chouayekh, H.; Sperandio, B.; Esnault, C.; Kormanec, J.; Virolle, M.-J. Regulation of ppk Expression and In Vivo Function of Ppk in Streptomyces lividans TK24. J. Bacteriol. 2006, 188, 6269–6276. [Google Scholar] [CrossRef]
- Liu, W.; Eder, S.; Hulett, F.M. Analysis of Bacillus subtilis tagAB and tagDEF Expression during Phosphate Starvation Identifies a Repressor Role for PhoP∼P. J. Bacteriol. 1998, 180, 753–758. [Google Scholar] [CrossRef]
- Tommassen, J.; de Geus, P.; Lugtenberg, B.; Hackett, J.; Reeves, P. Regulation of the pho regulon of Escherichia coli K-12. J. Mol. Biol. 1982, 157, 265–274. [Google Scholar] [CrossRef]
- Hulett, F.M. The signal-transduction network for Pho regulation in Bacillus subtilis. Mol. Microbiol. 1996, 19, 933–939. [Google Scholar] [CrossRef] [PubMed]
- Novak, R.; Cauwels, A.; Charpentier, E.; Tuomanen, E. Identification of a Streptococcus pneumoniae Gene Locus Encoding Proteins of an ABC Phosphate Transporter and a Two-Component Regulatory System. J. Bacteriol. 1999, 181, 1126–1133. [Google Scholar] [CrossRef] [PubMed]
- Makino, K.; Amemura, M.; Kim, S.K.; Nakata, A.; Shinagawa, H. Role of the sigma 70 subunit of RNA polymerase in transcriptional activation by activator protein PhoB in Escherichia coli. Genes Dev. 1993, 7, 149–160. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Blanco, A.G.; Solà, M.; Gomis-Ruth, F.X.; Coll, M. Tandem DNA Recognition by PhoB, a Two-Component Signal Transduction Transcriptional Activator. Structure 2002, 10, 701–713. [Google Scholar] [CrossRef]
- Yuan, Z.-C.; Zaheer, R.; Morton, R.; Finan, T.M. Genome prediction of PhoB regulated promoters in Sinorhizobium meliloti and twelve proteobacteria. Nucleic Acids Res. 2006, 34, 2686–2697. [Google Scholar] [CrossRef]
- Auesukaree, C.; Homma, T.; Tochio, H.; Shirakawa, M.; Kaneko, Y.; Harashima, S. Intracellular Phosphate Serves as a Signal for the Regulation of the PHO Pathway in Saccharomyces cerevisiae. J. Biol. Chem. 2004, 279, 17289–17294. [Google Scholar] [CrossRef]
- Carmany, D.O.; Hollingsworth, K.; McCleary, W.R. Genetic and Biochemical Studies of Phosphatase Activity of PhoR. J. Bacteriol. 2003, 185, 1112–1115. [Google Scholar] [CrossRef]
- Steed, P.M.; Wanner, B.L. Use of the rep technique for allele replacement to construct mutants with deletions of the pstSCAB-phoU operon: Evidence of a new role for the PhoU protein in the phosphate regulon. J. Bacteriol. 1993, 175, 6797–6809. [Google Scholar] [CrossRef]
- Schau, M.; Eldakak, A.; Hulett, F.M. Terminal Oxidases Are Essential to Bypass the Requirement for ResD for Full Pho Induction in Bacillus subtilis. J. Bacteriol. 2004, 186, 8424–8432. [Google Scholar] [CrossRef]
- Birkey, S.M.; Liu, W.; Zhang, X.; Duggan, M.F.; Hulett, F.M. Pho signal transduction network reveals direct transcriptional regulation of one two-component system by another two-component regulator: Bacillus subtilis PhoP directly regulates production of ResD. Mol. Microbiol. 1998, 30, 943–953. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Zhou, J.; Zhou, H.-J.; Wang, M.-M.; Liu, M.-M.; Ke, Y.-Z.; Li, P.-F.; Li, J.-N.; Du, H. Global Survey and Expressions of the Phosphate Transporter Gene Families in Brassica napus and Their Roles in Phosphorus Response. Int. J. Mol. Sci. 2020, 21, 1752. [Google Scholar] [CrossRef] [PubMed]
- Srivastava, S.; Srivastava, S. Prescience of endogenous regulation in Arabidopsis thaliana by Pseudomonas putida MTCC 5279 under phosphate starved salinity stress condition. Sci. Rep. 2020, 10, 5855. [Google Scholar] [CrossRef] [PubMed]
- Saia, S.; Rappa, V.; Ruisi, P.; Abenavoli, M.R.; Sunseri, F.; Giambalvo, D.; Frenda, A.S.; Martinelli, F. Soil inoculation with symbiotic microorganisms promotes plant growth and nutrient transporter genes expression in durum wheat. Front. Plant Sci. 2015, 6, 815. [Google Scholar] [CrossRef]
- Kadmiri, I.M.; Chaouqui, L.; Azaroual, S.E.; Sijilmassi, B.; Yaakoubi, K.; Wahby, I. Phosphate-Solubilizing and Auxin-Producing Rhizobacteria Promote Plant Growth Under Saline Conditions. Arab. J. Sci. Eng. 2018, 43, 3403–3415. [Google Scholar] [CrossRef]
- Chatterjee, S.; Sau, G.B.; Sinha, S.; Mukherjee, S.K. Effect of co-inoculation of plant growth-promoting rhizobacteria on the growth of amaranth plants. Arch. Agron. Soil Sci. 2012, 58, 1387–1397. [Google Scholar] [CrossRef]
- Bargaz, A.; Lyamlouli, K.; Chtouki, M.; Zeroual, Y.; Dhiba, D. Soil Microbial Resources for Improving Fertilizers Efficiency in an Integrated Plant Nutrient Management System. Front. Microbiol. 2018, 9, 1606. [Google Scholar] [CrossRef]
- Mahanta, D.; Rai, R.K.; Dhar, S.; Varghese, E.; Raja, A.; Purakayastha, T.J. Modification of root properties with phosphate solubilizing bacteria and arbuscular mycorrhiza to reduce rock phosphate application in soybean-wheat cropping system. Ecol. Eng. 2018, 111, 31–43. [Google Scholar] [CrossRef]
- Adnan, M.; Fahad, S.; Zamin, M.; Shah, S.; Mian, I.A.; Danish, S.; Zafar-Ul-Hye, M.; Battaglia, M.L.; Naz, R.M.M.; Saeed, B.; et al. Coupling Phosphate-Solubilizing Bacteria with Phosphorus Supplements Improve Maize Phosphorus Acquisition and Growth under Lime Induced Salinity Stress. Plants 2020, 9, 900. [Google Scholar] [CrossRef]
- Javeed, H.M.R.; Qamar, R.; Rehman, A.U.; Ali, M.; Rehman, A.; Farooq, M.; Zamir, S.I.; Nadeem, M.; Cheema, M.A.; Shehzad, M.; et al. Improvement in Soil Characteristics of Sandy Loam Soil and Grain Quality of Spring Maize by Using Phosphorus Solublizing Bacteria. Sustainability 2019, 11, 7049. [Google Scholar] [CrossRef]
- Suleman, M.; Yasmin, S.; Rasul, M.; Yahya, M.; Atta, B.M.; Mirza, M.S. Phosphate solubilizing bacteria with glucose dehydrogenase gene for phosphorus uptake and beneficial effects on wheat. PLoS ONE 2018, 13, e0204408. [Google Scholar] [CrossRef] [PubMed]
- Park, J.H.; Bolan, N.; Megharaj, M.; Naidu, R. Concomitant rock phosphate dissolution and lead immobilization by phosphate solubilizing bacteria (Enterobacter sp.). J. Environ. Manag. 2011, 92, 1115–1120. [Google Scholar] [CrossRef]
- Manzoor, M.; Abbasi, M.K.; Sultan, T. Isolation of Phosphate Solubilizing Bacteria from Maize Rhizosphere and Their Potential for Rock Phosphate Solubilization–Mineralization and Plant Growth Promotion. Geomicrobiol. J. 2017, 34, 81–95. [Google Scholar] [CrossRef]
- Elhaissoufi, W.; Khourchi, S.; Ibnyasser, A.; Ghoulam, C.; Rchiad, Z.; Zeroual, Y.; Lyamlouli, K.; Bargaz, A. Phosphate Solubilizing Rhizobacteria Could Have a Stronger Influence on Wheat Root Traits and Aboveground Physiology Than Rhizosphere P Solubilization. Front. Plant Sci. 2020, 11, 979. [Google Scholar] [CrossRef] [PubMed]
- Ben Zineb, A.; Trabelsi, D.; Ayachi, I.; Barhoumi, F.; Aroca, R.; Mhamdi, R. Inoculation with Elite Strains of Phosphate-Solubilizing Bacteria Enhances the Effectiveness of Fertilization with Rock Phosphates. Geomicrobiol. J. 2020, 37, 22–30. [Google Scholar] [CrossRef]
- Tahir, M.; Khalid, U.; Ijaz, M.; Shah, G.M.; Naeem, M.A.; Shahid, M.; Mahmood, K.; Ahmad, N.; Kareem, F. Combined application of bio-organic phosphate and phosphorus solubilizing bacteria (Bacillus strain MWT 14) improve the performance of bread wheat with low fertilizer input under an arid climate. Braz. J. Microbiol. 2018, 49, 15–24. [Google Scholar] [CrossRef]
- Jayakumar, N.; Paulraj, P.; Sajeesh, P.; Sajna, K.; Zinneera, A. Application of native phosphate solubilizing bacteria for the use of cheap organic and inorganic phosphate source in agricultural practise of Capsicum annum (Chili)—A pilot scale field study. Mater. Today Proc. 2019, 16, 1630–1639. [Google Scholar] [CrossRef]
- Bi, Q.-F.; Li, K.-J.; Zheng, B.-X.; Liu, X.-P.; Li, H.-Z.; Jin, B.-J.; Ding, K.; Yang, X.-R.; Lin, X.-Y.; Zhu, Y.-G. Partial replacement of inorganic phosphorus (P) by organic manure reshapes phosphate mobilizing bacterial community and promotes P bioavailability in a paddy soil. Sci. Total Environ. 2020, 703, 134977. [Google Scholar] [CrossRef]
- Skorupska, A.; Wielbo, J.; Kidaj, D.; Marek-Kozaczuk, M. Enhancing Rhizobium–Legume Symbiosis Using Signaling Factors. In Microbes for Legume Improvement; Springer: Vienna, Austria, 2010; pp. 27–54. [Google Scholar] [CrossRef]
- Lindström, K.; Mousavi, S.A. Effectiveness of nitrogen fixation in rhizobia. Microb. Biotechnol. 2020, 13, 1314–1335. [Google Scholar] [CrossRef]
- Lazali, M.; Zaman-Allah, M.; Amenc, L.; Ounane, G.; Abadie, J.; Drevon, J.-J. A phytase gene is overexpressed in root nodules cortex of Phaseolus vulgaris–rhizobia symbiosis under phosphorus deficiency. Planta 2013, 238, 317–324. [Google Scholar] [CrossRef]
- Arfarita, N.; Muhibuddin, A.; Imai, T. Exploration of indigenous free nitrogen-fixing bacteria from rhizosphere of Vigna radiata for agricultural land treatment. J. Degrad. Min. Lands Manag. 2019, 6, 1617–1623. [Google Scholar] [CrossRef]
- Nasto, M.K.; Alvarez-Clare, S.; Lekberg, Y.; Sullivan, B.W.; Townsend, A.R.; Cleveland, C.C. Interactions among nitrogen fixation and soil phosphorus acquisition strategies in lowland tropical rain forests. Ecol. Lett. 2014, 17, 1282–1289. [Google Scholar] [CrossRef]
- Mitran, T.; Meena, R.S.; Lal, R.; Layek, J.; Kumar, S.; Datta, R. Role of Soil Phosphorus on Legume Production. In Legumes for Soil Health and Sustainable Management; Springer: Singapore, 2018; pp. 487–510. [Google Scholar]
- Si, Z.; Yang, Q.; Liang, R.; Chen, L.; Chen, D.; Li, Y. Digalactosyldiacylglycerol Synthase Gene MtDGD1 Plays an Essential Role in Nodule Development and Nitrogen Fixation. Mol. Plant-Microbe Interact. 2019, 32, 1196–1209. [Google Scholar] [CrossRef]
- Aziz, A.; Ahiabor, B.; Opoku, A.; Abaidoo, R. Contributions of Rhizobium Inoculants and Phosphorus Fertilizer to Biological Nitrogen Fixation, Growth and Grain Yield of Three Soybean Varieties on a Fluvic Luvisol. Am. J. Exp. Agric. 2016, 10, 1–11. [Google Scholar] [CrossRef]
- Zaidi, A.; Ahemad, M.; Oves, M.; Ahmad, E.; Khan, M.S. Role of Phosphate-Solubilizing Bacteria in Legume Improvement. In Microbes for Legume Improvement; Springer: Vienna, Austria, 2010; pp. 273–292. [Google Scholar] [CrossRef]
- Sanz-Saez, A.; Morales, F.; Arrese-Igor, C.; Aranjuelo, I. P Deficiency: A Major Limiting Factor for Rhizobial Symbiosis. In Legume Nitrogen Fixation in Soils with Low Phosphorus Availability; Springer International Publishing: Cham, Switzerland, 2017; pp. 21–39. [Google Scholar]
- Van Gerven, L.P.A.; Kuiper, J.J.; Mooij, W.M.; Janse, J.H.; Paerl, H.W.; de Klein, J.J.M. Nitrogen fixation does not axiomatically lead to phosphorus limitation in aquatic ecosystems. Oikos 2019, 128, 563–570. [Google Scholar] [CrossRef]
- Tewari, S.; Pooniya, V.; Sharma, S. Next generation bioformulation prepared by amalgamating Bradyrhizobium, cell free culture supernatant, and exopolysaccharides enhances the indigenous rhizospheric rhizobial population, nodulation, and productivity of pigeon pea. Appl. Soil Ecol. 2020, 147, 103363. [Google Scholar] [CrossRef]
- Batterman, S.A.; Wurzburger, N.; Hedin, L.O. Nitrogen and phosphorus interact to control tropical symbiotic N2fixation: A test in Inga punctata. J. Ecol. 2013, 101, 1400–1408. [Google Scholar] [CrossRef]
- Singh, O.; Gupta, M.; Mittal, V.; Kiran, S.; Nayyar, H.; Gulati, A.; Tewari, R. Novel phosphate solubilizing bacteria ‘Pantoea cypripedii PS1′ along with Enterobacter aerogenes PS16 and Rhizobium ciceri enhance the growth of chickpea (Cicer arietinum L.). Plant Growth Regul. 2014, 73, 79–89. [Google Scholar] [CrossRef]
- Püschel, D.; Janoušková, M.; Voříšková, A.; Gryndlerová, H.; Vosátka, M.; Jansa, J. Arbuscular Mycorrhiza Stimulates Biological Nitrogen Fixation in Two Medicago spp. through Improved Phosphorus Acquisition. Front. Plant Sci. 2017, 8, 390. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Li, Q.; Guan, G.; Chen, S. Phosphate solubilizing bacteria stimulate wheat rhizosphere and endosphere biological nitrogen fixation by improving phosphorus content. PeerJ 2020, 8, e9062. [Google Scholar] [CrossRef] [PubMed]
- Zaidi, A.; Khan, M.S. Stimulatory effects of dual inoculation with phosphate solubilising microorganisms and arbuscular mycorrhizal fungus on chickpea. Aust. J. Exp. Agric. 2007, 47, 1016–1022. [Google Scholar] [CrossRef]
- Wani, P.A.; Khan, M.S.; Zaidi, A. Synergistic effects of the inoculation with nitrogen-fixing and phosphate-solubilizing rhizobacteria on the performance of field-grown chickpea. J. Plant Nutr. Soil Sci. 2007, 170, 283–287. [Google Scholar] [CrossRef]


{kind=link}
| Bacteria | Fungi | ||
|---|---|---|---|
| Genera | Ref. | Genera | Ref. |
| Aeromonas | [19] | Achrothcium | [3,20] |
| Agrobacterium | [18] | Alternaria | [3,20] |
| Azotobacter | [21,22] | Arthrobotrys | [3,20] |
| Bacillus | [23] | Aspergillus | [24] |
| Bradyrhizobium | [25] | Cephalosporium | [3,20] |
| Burkholderia | [19,26] | Chaetomium | [3,20] |
| Cyanobacteria | [3] | Cladosporium | [3,20] |
| Enterobacter | [22] | Cunninghamella | [3,20] |
| Erwinia | [27] | Curvularia | [3,20] |
| Kushneria | [5] | Fusarium | [3,20] |
| Micrococcus | [28] | Glomus | [3,20] |
| Paenibacillus | [29] | Helminthosporium | [3,20] |
| Pseudomonas | [30,31] | Micromonospora | [3,20] |
| Rhizobium | [32,33] | Phenomiocenspora | [3,20] |
| Rhodococcus | [25] | Phenomiocenspora | [3,20] |
| Salmonella | [25] | Phenomycylum | [34] |
| Serratia | [35] | Populospora | [3,20] |
| Sinomonas | [25] | Pythium | [3,20] |
| Thiobacillus | [25] | Rhizoctonia | [3,20] |
| Rhizopus | [3,20] | ||
| Saccharomyces | [3,20] | ||
| Schizosaccharomyces | [3,20] | ||
| Schwanniomyces | [3,20] | ||
| Sclerotium | [36] | ||
| Torula | [24] | ||
| Trichoderma | [24] | ||
| Yarrowia | [3,20] | ||
| Phosphate-Solubilizing Microorganism | Ecological Niche | Predominantly Produced Acids | Ref. |
|---|---|---|---|
| Escherichia freundii | Soil | Lactic | [62] |
| Aspergillus niger, Penicillium sp. | Soil | Citric, glycolic, succinic, gluconic, oxalic, lactic | [62] |
| Bacillus megaterium, Pseudomonas sp., Bacillus subtilus | Soil rizoshpere | Lactic, malic | [63] |
| Arthrobacter sp., Bascillus sp., Bacillus firmus B-7650 | Wheat and cowpea rhizosphere | Lactic, citric | [64] |
| Aspergillus sp., Penicillium sp., Chaetomium nigricolor | Lateritic soil | Oxalic, succinic, citric, 2-ketogluconic | [65] |
| A. japonicus, A. foetidus | Indian rock phosphate | Oxalic, citric, gluconic, succinic, tartaric | [66] |
| P. radicum | Wheat rhizosphere | Gluconic | [67] |
| Enterobacter agglomerans | Wheat rhizosphere | Oxalic, citric | [68] |
| Bacillus amyloliquefaciens, B. licheniformis, B. atrophaeus, Penibacillus macerans, Vibrio proteolyticus, Xanthobacter agilis, Enterobacter aerogenes, E. taylorae, E. asburiae, Kluyvera cryocrescens, Pseudomonas aeromonassens, Chrysler | Mangrove | Lactic, itaconic, isovaleric, isobutyric, acetic | [69] |
| Penicillium rugulosum | Venezuelan phosphate rocks | Citric, gluconic acid | [70] |
| Enterobacter intermedius | Grass rhizosphere | 2-ketogluconic | [71] |
| Aspergillus flavus, A. niger, Penicillium canescens | Wheat grains | Oxalic, citric, gluconic, succinic | [72] |
| Pseudomonas fluorescens | Oil palms rhizosphere | Citric, malic, tartaric, gluconic | [73] |
| Aspergillus niger | Tropical and subtropical soils | Gluconic, oxalic | [74] |
| P. trivialis | Rhizosphere of Hippophae rhamnoides (cold Howl and Spiti deserts, Trans-Himalayas) | Lactic, formic | [75] |
| B. pumilus var.2; B. subtilis var.2; Actinomadura oligospora; Citrobacter sp. | Giant cardon cactus (P. pringlei) | Gluconic, propionic, isovaleric, heptonic, caproic, isocaproic, formic, valeric, succinic, oxalic, oxaloacetic, malonic | [76] |
| B. pumilus CHOO8A; B. fusiformis | Opuntia cholla | gluconic, oxalic, 2-ketogluconic, lactic, succinic aid, formic, citric, malic | [76] |
| Bacillus sp. SENDO 6 и | P. pringlei | Gluconic, propionic, isovaleric, formic, succinic, lactic | [77] |
| Pseudomonas putida M5TSA, Enterobacter sakazakii M2PFe и Bacillus megaterium M1PCa | Mammillaria fraileana cactus | Gluconic, propionic, acetic, formic, succinic, lactic, oxalic | [78] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Timofeeva, A.; Galyamova, M.; Sedykh, S. Prospects for Using Phosphate-Solubilizing Microorganisms as Natural Fertilizers in Agriculture. Plants 2022, 11, 2119. https://doi.org/10.3390/plants11162119
Timofeeva A, Galyamova M, Sedykh S. Prospects for Using Phosphate-Solubilizing Microorganisms as Natural Fertilizers in Agriculture. Plants. 2022; 11(16):2119. https://doi.org/10.3390/plants11162119
Chicago/Turabian StyleTimofeeva, Anna, Maria Galyamova, and Sergey Sedykh. 2022. "Prospects for Using Phosphate-Solubilizing Microorganisms as Natural Fertilizers in Agriculture" Plants 11, no. 16: 2119. https://doi.org/10.3390/plants11162119
APA StyleTimofeeva, A., Galyamova, M., & Sedykh, S. (2022). Prospects for Using Phosphate-Solubilizing Microorganisms as Natural Fertilizers in Agriculture. Plants, 11(16), 2119. https://doi.org/10.3390/plants11162119