
Advanced Breeding for Biotic Stress Resistance in Poplar

 ,
,  ,
,
Abstract
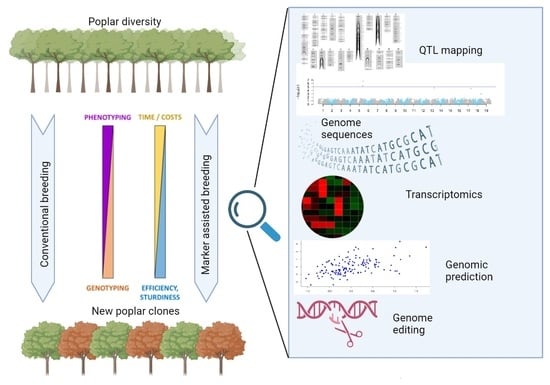
1. Introduction
2. Harnessing Poplar Diversity to Improve Pest and Disease Resistance
3. Genome Sequences Are Pivotal to Improve Poplar Breeding for Biotic Stress Resistance
4. An Overview of Known Loci Co-Segregating with Biotic Stress Resistance in Poplar
5. Transcriptomics for Disease Resistance in Poplar
6. Integration of Transcriptomic and Genomic Data
7. Breeding for Biotic Stresses in Poplar: Future Perspective
8. Conclusions: Are Current Knowledge on Poplar Sufficient to Support Advanced Breeding for Biotic Stress Resistance?
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| AFLP | Amplified Fragment Length Polymorphism |
| BAC | Artificial Bacterial Clone |
| CNV | Copy Number Variation |
| CRISPR | Cluster Regularly Interspersed Short Palindromic Repeats |
| DEG | Differentially Expressed Gene |
| eQTL | expression QTL |
| GbS | Genotyping-by-Sequencing |
| GE | Genome Editing |
| GO | Gene Onthology |
| GP | Genomic Prediction |
| GWAS | Genome Wide Association Scan |
| INDEL | insertion/deletion |
| LG | Linkage Group |
| MAB | marker-assisted breeding |
| MAS | marker-assisted selection |
| miRNA | microRNA |
| NGS | Next-Generation Sequencing |
| QTL | Quantitative Trait Locus |
| QTN | Quantitative Trait Nucleotide |
| R gene | Resistance gene |
| RNA-Seq | RNA Sequencing |
| SNP | Single Nucleotide Polymorphism |
| SSR | Simple Sequence Repeat |
| WPA | Woolly Poplar Aphid |
References
- Eckenwalder, J.E. Systematics and Evolution of of Populus. In Biology of Populus and Its Implications for Management and Conservation; Stettler, R.F., Bradshaw, H.D.J., Heilman, P.E., Hinckley, T.M., Eds.; NRC Research Press: Ottawa, ON, Canada, 1996; Chapter 1; pp. 7–32. [Google Scholar]
- Hamzeh, M.; Dayanandan, S. Phylogeny of Populus (Salicaceae) based on nucleotide sequences of chloroplast trnT-trnF region and nuclear rDNA. Am. J. Bot. 2004, 91, 1398–1408. [Google Scholar] [CrossRef] [PubMed]
- Ma, J.; Wan, D.; Duan, B.; Bai, X.; Bai, Q.; Chen, N.; Ma, T. Genome sequence and genetic transformation of a widely distributed and cultivated poplar. Plant Biotechnol. J. 2019, 17, 451–460. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.C.; Wang, J.; Delhomme, N.; Schiffthaler, B.; Sundström, G.; Zuccolo, A.; Nystedt, B.; Hvidsten, T.R.; de la Torre, A.; Cossu, R.M.; et al. Functional and evolutionary genomic inferences in Populus through genome and population sequencing of American and European aspen. Proc. Natl. Acad. Sci. USA 2018, 115, E10970–E10978. [Google Scholar] [CrossRef] [PubMed]
- Mohrdiek, O. Future possibilities for poplar breeding. Can. J. For. Res. 1983, 13, 465–471. [Google Scholar] [CrossRef]
- Tuskan, G.A.; DiFazio, S.; Jansson, S.; Bohlmann, J.; Grigoriev, I.; Hellsten, U.; Putnam, N.; Ralph, S.; Rombauts, S.; Salamov, A.; et al. The Genome of Black Cottonwood, Populus trichocarpa (Torr. & Gray). Science 2006, 313, 1596–1604. Available online: https://www.science.org/doi/abs/10.1126/science.1128691 (accessed on 7 July 2022).
- Carletti, G.; Carra, A.; Allegro, G.; Vietto, L.; Desiderio, F.; Bagnaresi, P.; Gianinetti, A.; Cattivelli, L.; Valè, G.; Nervo, G. QTLs for Woolly Poplar aphid (Phloeomyzus passerinii L.) resistance detected in an inter-specific Populus deltoides × P. nigra mapping population. PLoS ONE 2016, 11, e0152569. [Google Scholar] [CrossRef]
- Stanton, B.J.; Neale, D.B.; Li, S. Populus Breeding: From the Classical to the Genomic Approach. In Genetics and Genomics of Populus; Jansson, S., Bhalerao, R., Groover, A., Eds.; Springer: New York, NY, USA, 2010; pp. 309–348. [Google Scholar] [CrossRef]
- Pra, A.; Pettenella, D. Investment returns from hybrid poplar plantations in northern Italy between 2001 and 2016: Are we losing a bio-based segment of the primary economy? Ital. Rev. Agric. Econ. 2019, 74, 49–71. [Google Scholar] [CrossRef]
- Bergante, S.; Facciotto, G. Yields of poplar SRC and VSRC grown with different fertilization and irrigation management. In Proceedings of the 23th European Biomass Conference and Exhibition, Vienna, Austria, 1–4 June 2015; pp. 1–4. [Google Scholar]
- FAO. Poplars and Other Fast-Growing Trees—Renewable Resources for Future Green Economies. Synthesis of Country Progress Reports. In Proceedings of the 25th Session of the International Poplar Commission, Berlin, Germany, 12–16 September 2016; pp. 13–16. [Google Scholar]
- Cellerino, G. Review of Fungal Diseases in Poplar. Food and Agriculture Organization of the United Nations, 1999, AC492/E. Available online: https://www.fao.org/3/ac492e/AC492E.pdf (accessed on 7 July 2022).
- Stirling, B.; Newcombe, G.; Vrebalov, J.; Bosdet, I.; Bradshaw, J.D. Suppressed recombination around the MXC3 locus, a major gene for resistance to poplar leaf rust. Theor. Appl. Genet. 2001, 103, 1129–1137. [Google Scholar] [CrossRef]
- Duplessis, S.; Major, I.; Martin, F.; Séguin, A. Poplar and pathogen interactions: Insights from populus genome-wide analyses of resistance and defense gene families and gene expression profiling. Crit. Rev. Plant Sci. 2009, 28, 309–334. [Google Scholar] [CrossRef]
- Ostry, M.; Ramstedt, M.; Newcombe, G.; Steenackers, M. Diseases of poplars and willows. In Poplars and Willows: Trees for Society and the Environment; Isebrands, J.G., Richardson, J., Eds.; The Food and Agriculture Organization of the United Nations: Rome, Italy; CABI: Wallingford, UK, 2014; Chapter 8; pp. 443–458. [Google Scholar] [CrossRef]
- Gennaro, M.; Giorcelli, A. The biotic adversities of poplar in Italy: A reasoned analysis of factors determining the current state and future perspectives. Ann. Silvic. Res. 2019, 43, 41–51. [Google Scholar] [CrossRef]
- Newcombe, G.; Ostry, M.; Hubbes, M. Poplar diseases. In Poplar Culture in North America; Dickman, D.I., Isebrands, J.G., Eckenwalder, J.E., Richardson, J., Eds.; NRC Research Press: Ottawa, Canada, 2001; Chapter 8; pp. 249–276. [Google Scholar]
- Charles, J.G.; Nef, L.; Allegro, G.; Collins, C.M.; Delplanque, A.; Gimenez, R.; Hoglund, S.; Jiafu, H.; Larsson, S.; Luo, Y.; et al. Insect and other pests of poplars and willows. In Poplars and Willows: Trees for Society and the Environment; Isebrands, J.G., Richardson, J., Eds.; The Food and Agriculture Organization of the United Nations: Rome, Italy; CABI: Wallingford, UK, 2014; Chapter 9; pp. 459–526. [Google Scholar] [CrossRef]
- Allegro, G. The main insect pests affecting poplar plantations in Italy: Updates and perspectives. Forest 2017, 14, 74–83. [Google Scholar] [CrossRef]
- De Tillesse, V.; Nef, L.; Charles, J.; Hopkin, A.; Augustin, S. Damaging Poplar Insects, Internationally Important Species; The Food and Agriculture Organization of the United Nations: Rome, Italy, 2007; Available online: http://foris.fao.org/static/pdf/ipc/damaging_poplar_insects_eBook.pdf (accessed on 7 July 2022).
- Navratil, S. Virus and Virus-Like Diseases of Poplar: Are They Threatening Diseases? Aspen Bibliogr. 1979, 4635. Available online: https://digitalcommons.usu.edu/cgi/viewcontent.cgi?article=5634&context=aspen_bib (accessed on 7 July 2022).
- Li, A.; He, W. Molecular Aspects of an Emerging Poplar Canker Caused by Lonsdalea populi. Front. Microbiol. 2019, 10, 2496. [Google Scholar] [CrossRef] [PubMed]
- Zlatković, M.; Tenorio-Baigorria, I.; Lakatos, T.; Tóth, T.; Koltay, A.; Pap, P.; Markovic, M.; Orlović, S. Bacterial canker disease on Populus × euramericana caused by Lonsdalea populi in serbia. Forests 2020, 11, 1080. [Google Scholar] [CrossRef]
- Sturrock, R.N.; Frankel, S.J.; Brown, A.V.; Hennon, P.E.; Kliejunas, J.T.; Lewis, K.J.; Worrall, J.J.; Woods, A.J. Climate change and forest diseases. Plant Pathol. 2011, 60, 133–149. [Google Scholar] [CrossRef]
- Ji, Y.; Zhou, G.; Li, Z.; Wang, S.; Zhou, H.; Song, X. Triggers of widespread dieback and mortality of poplar (Populus spp.) plantations across northern China. J. Arid Environ. 2020, 174, 104076. [Google Scholar] [CrossRef]
- Seserman, D.M.; Pohle, I.; Veste, M.; Freese, D. Simulating climate change impacts on hybrid-poplar and black locust short rotation coppices. Forests 2018, 9, 419. [Google Scholar] [CrossRef]
- Bebber, D.P.; Ramotowski, M.A.T.; Gurr, S.J. Crop pests and pathogens move polewards in a warming world. Nat. Clim. Change 2013, 3, 985–988. [Google Scholar] [CrossRef]
- Burdon, J.J.; Zhan, J. Climate change and disease in plant communities. PLoS Biol. 2020, 18, e3000949. [Google Scholar] [CrossRef]
- Brodde, L.; Adamson, K.; Camarero, J.J.; Castaño, C.; Drenkhan, R.; Lehtijärvi, A.; Luchi, N.; Migliorini, D.; Sanchez-Miranda, A.; Stenlid, J. Diplodia tip blight on its way to the north: Drivers of disease emergence in northern Europe. Front. Plant Sci. 2019, 9, 1818. [Google Scholar] [CrossRef]
- Mcivor, I.; Jones, T. Novel Poplars and Willow Adapted to Climate Change; Final Report PFR SPTS No. 11980; The New Zealand Institute for Plant & Food Research Limited: Auckland, New Zealand, 2015. [Google Scholar]
- Allwright, M.R.; Taylor, G. Molecular Breeding for Improved Second Generation Bioenergy Crops. Trends Plant Sci. 2016, 21, 43–54. [Google Scholar] [CrossRef]
- Stanturf, J.A.; van Oosten, C. Operational poplar and willow culture. In Poplars and Willows: Trees for Society and the Environment; Isebrands, J.G., Richardson, J., Eds.; The Food and Agriculture Organization of the United Nations: Rome, Italy; CABI: Wallingford, UK, 2014; Chapter 5; pp. 200–257. [Google Scholar] [CrossRef]
- Gortari, F.; Guiamet, J.J.; Cortizo, S.C.; Graciano, C. Poplar leaf rust reduces dry mass accumulation and internal nitrogen recycling more markedly under low soil nitrogen availability, and decreases growth in the following spring. Tree Physiol. 2018, 39, 19–30. [Google Scholar] [CrossRef] [PubMed]
- Teissier, E. Breeding Strategies with Poplars in Europe Tree breeders have always favoured poplars in their programmes for various reasons: High vigour of most of the poplar species, short generations, natural ability for interspecific hybridization and great poten. Science 1984, 8, 23–39. [Google Scholar]
- Steenackers, V.; Strobl, S.; Steenackers, M. Collection and distribution of poplar species, hybrids and clones. Biomass 1990, 22, 1–20. [Google Scholar] [CrossRef]
- Bisoffi, S.; Gullberg, U. Poplar breeding and selection strategies. In Biology of Populus and Its Implications for Management and Conservation; Stettler, R.F., Bradshaw, H.D.J., Heilman, P.E., Hinckley, T.M., Eds.; NRC Research Press: Ottawa, ON, Canada, 1996; Chapter 6; pp. 139–158. [Google Scholar]
- Cagelli, L.; Lefèvre, F. The conservation of Populus nigra L. and gene flow within cultivated poplars in Europe. Bocconea 1997, 7, 63–75. [Google Scholar]
- Pinon, J.; Frey, P. Interactions Between Poplar Clones and Melampsora Populations and their implications for breeding for durable resistance. In Rust Diseses of Willow and Poplar; Pei, M.H., McCracken, A.R., Eds.; CABI Publishing: Oxfordshire, Cambridge, 2005; pp. 139–154. Available online: http://sherekashmir.informaticspublishing.com/609/1/9780851999999.pdf (accessed on 7 July 2022).
- Li, S.; Zhang, Z.; Luo, J.; He, C.; Pu, Y.; An, X. Progress and strategies in cross breeding of poplars in China. For. Stud. China 2005, 7, 54–60. [Google Scholar] [CrossRef]
- McIvor, I.R.; Douglas, G.B. Poplars and willows in hill country-stabilising soils and storing carbon. In Advanced Nutrient Management: Gains from the Past–Goals for the Future; Currie, L.D., Christensen, C.L., Eds.; Massey University: Palmerston North, New Zealand, 2012; pp. 1–11. Available online: https://www.massey.ac.nz/~flrc/workshops/12/Manuscripts/McIvor_2012.pdf (accessed on 7 July 2022).
- Tsarev, A.P.; Tsareva, R.P.; Tsarev, V.A. Poplar testing and breeding in the Central Chernozem region of Russia. IOP Conf. Ser. Earth Environ. Sci. 2019, 392, 12010. [Google Scholar] [CrossRef]
- Liesebach, M. Poplars and Other Fast Growing Tree Species in Germany: Report of the National Poplar Commission. 2016–2019; Thünen Working Paper, No. 141a; Johann Heinrich von Thünen-Institut: Braunschweig, Germany, 2020. [Google Scholar] [CrossRef]
- Dunnell, K.L.; Berguson, B.; McMahon, B.; Leboldus, J.M. Variation in resistance of Populus nigra to Sphaerulina musiva in the north-central United States. Plant Dis. 2016, 100, 287–291. [Google Scholar] [CrossRef]
- Godbout, J.; Gros-Louis, M.C.; Lamothe, M.; Isabel, N. Going with the flow: Intraspecific variation may act as a natural ally to counterbalance the impacts of global change for the riparian species Populus deltoides. Evol. Appl. 2020, 13, 176–194. [Google Scholar] [CrossRef]
- Legionnet, A.; Muranty, H.; Lefèvre, F. Genetic variation of the riparian pioneer tree species Populus nigra. II. Variation in susceptibility to the foliar rust Melampsora larici-populina. Heredity 1999, 82, 318–327. [Google Scholar] [CrossRef]
- Benetka, V.; Novotná, K.; Štochlová, P. Wild populations as a source of germplasm for black poplar (Populus nigra L.) breeding programmes. Tree Genet. Genom. 2012, 8, 1073–1084. [Google Scholar] [CrossRef]
- Dunlap, J.M.; Stettler, R.F. Genetic variation and productivity of Populus trichocarpa and its hybrids. IX. Phenology and Melampsora rust incidence of native black cottonwood clones from four river valleys in Washington. For. Ecol. Manag. 1996, 87, 233–256. [Google Scholar] [CrossRef]
- Bari, A.; Street, K.; Mackay, M.; Endresen, D.T.F.; De Pauw, E.; Amri, A. Focused identification of germplasm strategy (FIGS) detects wheat stem rust resistance linked to environmental variables. Genet. Res. Crop Evol. 2012, 59, 1465–1481. [Google Scholar] [CrossRef]
- Bisoffi, S. Poplar breeding strategies between conventional methods and new techniques. In Proceedings of the 1 Simposio del Chopo, Zamora, España, 9−11 May 2001. [Google Scholar]
- Chen, Q.; Wang, J.; Li, D.; Wang, Z.; Wang, F.; Zhang, R. Molecular characterization of the transcription factors in susceptible poplar infected with virulent Melampsora larici-populina. Int. J. Mol. Sci. 2019, 20, 4806. [Google Scholar] [CrossRef] [PubMed]
- Cervera, M.T.; Gusmão, J.; Steenackers, M.; Peleman, J.; Storme, V.; Vanden Broeck, A.; Van Montagu, M.; Boerjan, W. Identification of AFLP molecular markers for resistance against Melampsora larici-populina in Populus. Theor. Appl. Genet. 1996, 93, 733–737. [Google Scholar] [CrossRef] [PubMed]
- Newcombe, G.; Van Oosten, C. Variation in resistance to Venturia populina, the cause of poplar leaf and shoot blight in the Pacific Northwest. Can. J. For. Res. 1997, 27, 883–889. [Google Scholar] [CrossRef]
- Bisoffi, S. The development of a breeding strategy for poplars. International Poplar Commission. In Proceedings of the Ad Hoc Committee for Poplar and Willow Breeding, Buenos Aires, Argentina, 19−23 March 1990. [Google Scholar]
- Yang, W.; Wang, K.; Zhang, J.; Ma, J.; Liu, J.; Ma, T. The draft genome sequence of a desert tree Populus pruinosa. GigaScience 2017, 6, 2797. [Google Scholar] [CrossRef]
- Zhang, Z.; Chen, Y.; Zhang, J.; Ma, X.; Li, Y.; Li, M.; Wang, D.; Kang, M.; Wu, H.; Yang, Y.; et al. Improved genome assembly provides new insights into genome evolution in a desert poplar (Populus euphratica). Mol. Ecol. Resour. 2020, 20, 781–794. [Google Scholar] [CrossRef]
- Ma, T.; Wang, J.; Zhou, G.; Yue, Z.; Hu, Q.; Chen, Y.; Liu, B.; Qiu, Q.; Wang, Z.; Zhang, J.; et al. Genomic insights into salt adaptation in a desert poplar. Nat. Commun. 2013, 4, 2797. [Google Scholar] [CrossRef]
- Liu, Y.J.; Wang, X.R.; Zeng, Q.Y. De novo assembly of white poplar genome and genetic diversity of white poplar population in Irtysh River basin in China. Sci. China Life Sci. 2019, 62, 609–618. [Google Scholar] [CrossRef]
- Wu, H.; Yao, D.; Chen, Y.; Yang, W.; Zhao, W.; Gao, H.; Tong, C. De novo genome assembly of Populus simonii further supports that Populus simonii and Populus trichocarpa belong to different sections. G3 Genes Genom. Genet. 2020, 10, 455–466. [Google Scholar] [CrossRef]
- Bai, S.; Wu, H.; Zhang, J.; Pan, Z.; Zhao, W.; Li, Z.; Tong, C. Genome assembly of salicaceae populus deltoides (eastern cottonwood) i-69 based on nanopore sequencing and hi-c technologies. J. Hered. 2021, 112, 303–310. [Google Scholar] [CrossRef] [PubMed]
- Qiu, D.; Bai, S.; Ma, J.; Zhang, L.; Shao, F.; Zhang, K.; Yang, Y.; Sun, T.; Huang, J.; Zhou, Y.; et al. The genome of Populus alba × Populus tremula var. glandulosa clone 84K. DNA Res. 2019, 26, 423–431. [Google Scholar] [CrossRef] [PubMed]
- Huang, X.; Chen, S.; Peng, X.; Bae, E.K.; Dai, X.; Liu, G.; Qu, G.; Ko, J.H.; Lee, H.; Chen, S.; et al. An improved draft genome sequence of hybrid Populus alba × Populus glandulosa. J. For. Res. 2021, 32, 1663–1672. [Google Scholar] [CrossRef]
- Pinosio, S.; Giacomello, S.; Faivre-Rampant, P.; Taylor, G.; Jorge, V.; Le Paslier, M.C.; Zaina, G.; Bastien, C.; Cattonaro, F.; Marroni, F.; et al. Characterization of the Poplar Pan-Genome by Genome-Wide Identification of Structural Variation. Mol. Biol. Evol. 2016, 33, 2706–2719. [Google Scholar] [CrossRef] [PubMed]
- Filiz, E.; Vatansever, R. Genome-wide identification of mildew resistance locus O (MLO) genes in tree model poplar (Populus trichocarpa): Powdery mildew management in woody plants. Eur. J. Plant Pathol. 2018, 152, 95–109. [Google Scholar] [CrossRef]
- Jiang, Y.; Duan, Y.; Yin, J.; Ye, S.; Zhu, J.; Zhang, F.; Lu, W.; Fan, D.; Luo, K. Genome-wide identification and characterization of the Populus WRKY transcription factor family and analysis of their expression in response to biotic and abiotic stresses. J. Exp. Bot. 2014, 65, 6629–6644. [Google Scholar] [CrossRef]
- Xing, H.; Fu, X.; Yang, C.; Tang, X.; Guo, L.; Li, C.; Xu, C.; Luo, K. Genome-wide investigation of pentatricopeptide repeat gene family in poplar and their expression analysis in response to biotic and abiotic stresses. Sci. Rep. 2018, 8, 2817. [Google Scholar] [CrossRef]
- Piot, A.; Prunier, J.; Isabel, N.; Klápště, J.; El-Kassaby, Y.A.; Villarreal Aguilar, J.C.; Porth, I. Genomic Diversity Evaluation of Populus trichocarpa Germplasm for Rare Variant Genetic Association Studies. Front. Genet. 2020, 10, 1–13. [Google Scholar] [CrossRef]
- Fahrenkrog, A.M.; Neves, L.G.; Resende, M.F.R.; Dervinis, C.; Davenport, R.; Barbazuk, W.B.; Kirst, M. Population genomics of the eastern cottonwood (Populus deltoides). Ecol. Evol. 2017, 7, 9426–9440. [Google Scholar] [CrossRef]
- Geraldes, A.; Difazio, S.P.; Slavov, G.T.; Ranjan, P.; Muchero, W.; Hannemann, J.; Gunter, L.E.; Wymore, A.M.; Grassa, C.J.; Farzaneh, N.; et al. A 34K SNP genotyping array for Populus trichocarpa: Design, application to the study of natural populations and transferability to other Populus species. Mol. Ecol. Resour. 2013, 13, 306–323. [Google Scholar] [CrossRef]
- Faivre-Rampant, P.; Zaina, G.; Jorge, V.; Giacomello, S.; Segura, V.; Scalabrin, S.; Guerin, V.; De Paoli, E.; Aluome, C.; Viger, M.; et al. New resources for genetic studies in Populus nigra: Genome-wide SNP discovery and development of a 12k Infinium array. Mol. Ecol. Resour. 2016, 16, 1023–1036. [Google Scholar] [CrossRef] [PubMed]
- Lachance, J.; Tishkoff, S.A. SNP ascertainment bias in population genetic analyses: Why it is important, and how to correct it. BioEssays News Rev. Mol. Cell. Dev. Biol. 2013, 35, 780–786. [Google Scholar] [CrossRef]
- Schilling, M.P.; Wolf, P.G.; Duffy, A.M.; Rai, H.S.; Rowe, C.A.; Richardson, B.A.; Mock, K.E. Genotyping-by-sequencing for Populus population genomics: An assessment of genome sampling patterns and filtering approaches. PLoS ONE 2014, 9, e95292. [Google Scholar] [CrossRef]
- Scaglione, D.; Pinosio, S.; Marroni, F.; Centa, E.; Di Fornasiero, A.; Magris, G.; Scalabrin, S.; Cattonaro, F.; Taylor, G.; Morgante, M. Single primer enrichment technology as a tool for massive genotyping: A benchmark on black poplar and maize. Ann. Bot. 2019, 124, 543–551. [Google Scholar] [CrossRef]
- Yin, T.M.; DiFazio, S.P.; Gunter, L.E.; Jawdy, S.S.; Boerjan, W.; Tuskan, G.A. Genetic and physical mapping of Melampsora rust resistance genes in Populus and characterization of linkage disequilibrium and flanking genomic sequence. New Phytol. 2004, 164, 95–105. [Google Scholar] [CrossRef]
- DeWoody, J.; Viger, M.; Lakatos, F.; Tuba, K.; Taylor, G.; Smulders, M.J.M. Insight into the genetic components of community genetics: QTL mapping of insect association in a fast-growing forest tree. PLoS ONE 2013, 8, e79925. [Google Scholar] [CrossRef] [PubMed]
- Newcombe, G.; Muchero, W.; Busby, P.E. Resistance to an eriophyid mite in an interspecific hybrid pedigree of Populus. PLoS ONE 2018, 13, e0207839. [Google Scholar] [CrossRef] [PubMed]
- Jiang, H.; Wan, Z.; Liu, M.; Hou, J.; Yin, T. Mapping quantitative trait loci conferring resistance to Marssonina leaf spot disease in Populus deltoides. Trees-Struct. Funct. 2019, 33, 697–706. [Google Scholar] [CrossRef]
- Newcombe, G.; Bradshaw, J. Quantitative trait loci conferring resistance in hybrid poplar to Septoria populicola, the cause of leaf spot. Can. J. For. Res. 1996, 26, 1943–1950. [Google Scholar] [CrossRef]
- Newcombe, G.; Bradshaw, H.D.; Chastagner, G.A.; Stettler, R.F. A major gene for resistance to Melampsora medusae f. sp. deltoidae in a hybrid poplar pedigree. Phytopathology 1996, 86, 87–94. [Google Scholar] [CrossRef]
- Tabor, G.M.; Kubisiak, T.L.; Klopfenstein, N.B.; Hall, R.B.; McNabb, J. Bulked segregant analysis identifies molecular markers linked to Melampsora medusae resistance in Populus deltoides. Phytopathology 2000, 90, 1039–1042. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Zhang, J.; Steenackers, M.; Storme, V.; Neyrinck, S.; Van Montagu, M.; Gerats, T.; Boerjan, W. Fine mapping and identification of nucleotide binding site/leucine-rich repeat sequences at the MER locus in Populus deltoides “S9-2”. Phytopathology 2001, 91, 1069–1073. [Google Scholar] [CrossRef]
- Jorge, V.; Dowkiw, A.; Faivre-Rampant, P.; Basrtien, C. Genetic architecture of qualitative and quantitative Melampsora larici-populina leaf rust resistance in hybrid poplar: Genetic mapping and QTL detection. New Phytol. 2005, 167, 113–127. [Google Scholar] [CrossRef] [PubMed]
- Bresson, A.; Jorge, V.; Dowkiw, A.; Guerin, V.; Bourgait, I.; Tuskan, G.A.; Schmutz, J.; Chalhoub, B.; Bastien, C.; Faivre Rampant, P. Qualitative and quantitative resistances to leaf rust finely mapped within two nucleotide-binding site leucine-rich repeat (NBS-LRR)-rich genomic regions of chromosome 19 in poplar. New Phytol. 2011, 192, 151–163. [Google Scholar] [CrossRef] [PubMed]
- Wei, S.; Wu, H.; Li, X.; Chen, Y.; Yang, Y.; Dai, M.; Yin, T. Identification of genes underlying the resistance to Melampsora larici-populina in an R Gene supercluster of the Populus deltoides genome. Plant Dis. 2020, 104, 1133–1143. [Google Scholar] [CrossRef] [PubMed]
- Nvsvrot, T.; Xia, W.; Xiao, Z.; Zhan, C.; Liu, M.; Yang, X.; Zhang, Y.; Wang, N. Combining QTL mapping with genome resequencing identifies an indel in an R gene that is associated with variation in leaf rust disease resistance in poplar. Phytopathology 2020, 110, 900–906. [Google Scholar] [CrossRef] [PubMed]
- La Mantia, J.; Klápště, J.; El-Kassaby, Y.A.; Azam, S.; Guy, R.D.; Douglas, C.J.; Mansfield, S.D.; Hamelin, R. Association analysis identifies Melampsora xcolumbiana poplar leaf rust resistance SNPs. PLoS ONE 2013, 8, e78423. [Google Scholar] [CrossRef]
- Muchero, W.; Sondreli, K.L.; Chen, J.G.; Urbanowicz, B.R.; Zhang, J.; Singan, V.; Yang, Y.; Brueggeman, R.S.; Franco-Coronado, J.; Abraham, N.; et al. Association mapping, transcriptomics, and transient expression identify candidate genes mediating plant-pathogen interactions in a tree. Proc. Natl. Acad. Sci. USA 2018, 115, 11573–11578. [Google Scholar] [CrossRef]
- Zhang, J.; Yang, Y.; Zheng, K.; Xie, M.; Feng, K.; Jawdy, S.S.; Gunter, L.E.; Ranjan, P.; Singan, V.R.; Engle, N.; et al. Genome-wide association studies and expression-based quantitative trait loci analyses reveal roles of HCT2 in caffeoylquinic acid biosynthesis and its regulation by defense-responsive transcription factors in Populus. New Phytol. 2018, 220, 502–516. [Google Scholar] [CrossRef]
- Chu, Y.; Su, X.; Huang, Q.; Zhang, X. Patterns of DNA sequence variation at candidate gene loci in black poplar (Populus nigra L.) as revealed by single nucleotide polymorphisms. Genetica 2009, 137, 141–150. [Google Scholar] [CrossRef]
- Liu, X.; Lin, H.; Ding, C.; Su, X. Identification of single nucleotide polymorphisms and analysis of linkage disequilibrium in black poplar (Populus nigra L.) inbred lines using the candidate gene approach. Sci. For. 2021, 49, 1–13. [Google Scholar] [CrossRef]
- Fahrenkrog, A.M.; Neves, L.G.; Resende, M.F.R.; Vazquez, A.I.; de los Campos, G.; Dervinis, C.; Sykes, R.; Davis, M.; Davenport, R.; Barbazuk, W.B.; et al. Genome-wide association study reveals putative regulators of bioenergy traits in Populus deltoides. New Phytol. 2017, 213, 799–811. [Google Scholar] [CrossRef] [PubMed]
- Ismail, M.; Soolanayakanahally, R.Y.; Ingvarsson, P.K.; Guy, R.D.; Jansson, S.; Silim, S.N.; El-Kassaby, Y.A. Comparative nucleotide diversity across north American and European Populus species. J. Mol. Evol. 2012, 74, 257–272. [Google Scholar] [CrossRef] [PubMed]
- Jia, H.; Liu, G.; Li, J.; Zhang, J.; Sun, P.; Zhao, S.; Zhou, X.; Lu, M.; Hu, J. Genome resequencing reveals demographic history and genetic architecture of seed salinity tolerance in Populus euphratica. J. Exp. Bot. 2020, 71, 4308–4320. [Google Scholar] [CrossRef] [PubMed]
- Olson, M.S.; Robertson, A.L.; Takebayashi, N.; Silim, S.; Schroeder, W.R.; Tiffin, P. Nucleotide diversity and linkage disequilibrium in balsam poplar (Populus balsamifera). New Phytol. 2010, 186, 526–536. [Google Scholar] [CrossRef]
- Ingvarsson, P.K. Multilocus patterns of nucleotide polymorphism and the demographic history of Populus tremula. Genetics 2008, 180, 329–340. [Google Scholar] [CrossRef]
- Du, Q.; Lu, W.; Quan, M.; Xiao, L.; Song, F.; Li, P.; Zhou, D.; Xie, J.; Wang, L.; Zhang, D. Genome-wide association studies to improve wood properties: Challenges and prospects. Front. Plant Sci. 2018, 871, 1912. [Google Scholar] [CrossRef]
- Kovalchuk, A.; Keriö, S.; Oghenekaro, A.O.; Jaber, E.; Raffaello, T.; Asiegbu, F.O. Antimicrobial defenses and resistance in forest trees: Challenges and perspectives in a genomic era. Ann. Rev. Phytopathol. 2013, 51, 221–244. [Google Scholar] [CrossRef]
- Miranda, M.; Ralph, S.G.; Mellway, R.; White, R.; Heath, M.C.; Bohlmann, J.; Constabel, C.P. The transcriptional response of hybrid poplar (Populus trichocarpa × P. deltoides) to infection by Melampsora medusae leaf rust involves induction of flavonoid pathway genes leading to the accumulation of proanthocyanidins. Mol. Plant-Mic. Interact. 2007, 20, 816–831. [Google Scholar] [CrossRef]
- Rinaldi, C.; Kohler, A.; Frey, P.; Duchaussoy, F.; Ningre, N.; Couloux, A.; Wincker, P.; Le Thiec, D.; Fluch, S.; Martin, F.; et al. Transcript profiling of poplar leaves upon infection with compatible and incompatible strains of the foliar rust Melampsora larici-populina. Plant Physiol. 2007, 144, 347–366. [Google Scholar] [CrossRef] [PubMed]
- Azaiez, A.; Boyle, B.; Levée, V.; Séguin, A. Transcriptome profiling in hybrid poplar following interactions with Melampsora rust fungi. Mol. Plant-Mic. Interact. 2009, 22, 190–200. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Zhang, X.; Chen, Y.; Wang, Q.; Wang, M.; Huang, M. Function and Chromosomal Localization of Differentially Expressed Genes Induced by Marssonina brunnea f. sp. multigermtubi in Populus deltoides. J. Genet. Genom. 2007, 34, 641–648. [Google Scholar] [CrossRef]
- Wang, Z.; Gerstein, M.; Snyder, M. RNA-Seq: A revolutionary tool for transcriptomics. Nat. Rev. Genet. 2009, 10, 57–63. [Google Scholar] [CrossRef] [PubMed]
- Liang, H.; Staton, M.; Xu, Y.; Xu, T.; Leboldus, J. Comparative expression analysis of resistant and susceptible Populus clones inoculated with Septoria musiva. Plant Sci. 2014, 223, 69–78. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Tian, L.; Yan, D.H.; He, W. Genome-wide transcriptome analysis reveals the comprehensive response of two susceptible poplar sections to Marssonina brunnea infection. Genes 2018, 9, 154. [Google Scholar] [CrossRef]
- Müller, N.A.; Kersten, B.; Fladung, M.; Schroeder, H. RNA-seq of eight different poplar clones reveals conserved up-regulation of gene expression in response to insect herbivory. BMC Genom. 2019, 20, 673. [Google Scholar] [CrossRef] [PubMed]
- Chen, M.; Cao, Z. Genome-wide expression profiling of microRNAs in poplar upon infection with the foliar rust fungus Melampsora larici-populina. BMC Genom. 2015, 16, 696. [Google Scholar] [CrossRef]
- Chen, S.; Zhang, Y.; Zhao, Y.; Xu, W.; Li, Y.; Xie, J.; Zhang, D. Key genes and genetic interactions of plant-pathogen functional modules in poplar infected by Marssonina brunnea. Mol. Plant-Mic. Interact. 2020, 33, 1080–1090. [Google Scholar] [CrossRef]
- Hansen, B.G.; Halkier, B.A.; Kliebenstein, D.J. Identifying the molecular basis of QTLs: eQTLs add a new dimension. Trends Plant Sci. 2008, 13, 72–77. [Google Scholar] [CrossRef]
- Muranty, H.; Jorge, V.; Dowkiw, A.; Guerin, V.; Faivre-Rampant, P.; Bresson, A.; Labbè, J.; Tuskan, G.A.; SAnchez, L.; Bastien, C. A case study in marker-assisted breeding for disease resistance: The building of complex resistance to Melampsora larici-populina in hybrid poplar. In Proceedings of the Final Conference Noveltree Tree Breeding, Genomics and Evolutionary Biology: New Synergies to Tackle the Impact of Climate Change in the 21st Century, Helsinki, Finland, 16−17 October 2012; pp. 49–50. Available online: https://hal.archives-ouvertes.fr/index/index#page=50 (accessed on 7 July 2022).
- Naidoo, S.; Slippers, B.; Plett, J.M.; Coles, D.; Oates, C.N. The road to resistance in forest trees. Front. Plant Sci. 2019, 10, 273. [Google Scholar] [CrossRef]
- Migicovsky, Z.; Myles, S. Exploiting wild relatives for genomics-assisted breeding of perennial crops. Front. Plant Sci. 2017, 8, 460. [Google Scholar] [CrossRef] [PubMed]
- Butcher, P.; Southerton, S. Marker-assisted selection in forestry species. In Current Status and Future Perspectives in Crops, Livestock, Forestry and Fish; Guimraes, E.P., Ruane, J., Scherf, B.D., Sonnino, A., Dargie, J.D., Eds.; Food and Agriculture Organization of the United Nation (FAO): Rome, Italy, 2007; Volume 15, pp. 283–305. [Google Scholar]
- Meuwissen, T.H.E.; Hayes, B.J.; Goddard, M.E. Prediction of total genetic value using genome-wide dense marker maps. Genetics 2001, 157, 1819–1829. [Google Scholar] [CrossRef] [PubMed]
- Cros, D.; Denis, M.; Bouvet, J.M.; Sánchez, L. Long-term genomic selection for heterosis without dominance in multiplicative traits: Case study of bunch production in oil palm. BMC Genom. 2015, 16, 651. [Google Scholar] [CrossRef] [PubMed]
- Wong, C.K.; Bernardo, R. Genomewide selection in oil palm: Increasing selection gain per unit time and cost with small populations. Theor. Appl. Genet. 2008, 116, 815–824. [Google Scholar] [CrossRef]
- Thakur, A.K.; Kumar, P.; Parmar, N.; Shandil, R.K.; Aggarwal, G.; Gaur, A.; Srivastava, D.K. Achievements and prospects of genetic engineering in poplar: A review. New For. 2021, 52, 889–920. [Google Scholar] [CrossRef]
- Buyel, J.F.; Stöger, E.; Bortesi, L. Targeted genome editing of plants and plant cells for biomanufacturing. Transgenic Res. 2021, 30, 401–426. [Google Scholar] [CrossRef]
- Strauss, S.H.; Costanza, A.; Séguin, A. Genetically engineered trees: Paralysis from good intentions: Forest crises demand regulation and certification reform. Science 2015, 349, 794–795. [Google Scholar] [CrossRef]
- Doudna, J.A.; Charpentier, E. Genome editing. The new frontier of genome engineering with CRISPR-Cas9. Science 2015, 346, 1258096. [Google Scholar] [CrossRef]
- Schaeffer, S.M.; Nakata, P.A. CRISPR/Cas9-mediated genome editing and gene replacement in plants: Transitioning from lab to field. Plant Sci. 2015, 240, 130–142. [Google Scholar] [CrossRef]
- Kantor, A.; Mcclements, M.E. CRISPR-Cas9 DNA Base-Editing and Prime-Editing. Int. J. Mol. Sci. 2020, 21, 6240. [Google Scholar] [CrossRef]
- Li, H.; Yang, Y.; Hong, W.; Huang, M.; Wu, M.; Zhao, X. Applications of genome editing technology in the targeted therapy of human diseases: Mechanisms, advances and prospects. Signal Transduct. Target. 2020, 5, 1. [Google Scholar] [CrossRef] [PubMed]
- Wada, N.; Ueta, R.; Osakabe, Y.; Osakabe, K. Precision genome editing in plants: State-of-the-art in CRISPR/Cas9-based genome engineering. BMC Plant Biol. 2020, 20, 234. [Google Scholar] [CrossRef] [PubMed]
- Clifton-Brown, J.; Harfouche, A.; Casler, M.D.; Dylan Jones, H.; Macalpine, W.J.; Murphy-Bokern, D.; Smart, L.B.; Adler, A.; Ashman, C.; Awty-Carroll, D.; et al. Breeding progress and preparedness for mass-scale deployment of perennial lignocellulosic biomass crops switchgrass, miscanthus, willow and poplar. GCB Bioenergy 2019, 11, 118–151. [Google Scholar] [CrossRef] [PubMed]
- Dort, E.N.; Tanguay, P.; Hamelin, R.C. CRISPR/Cas9 Gene Editing: An Unexplored Frontier for Forest Pathology. Front. Plant Sci. 2020, 11, 1126. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Y.; Guo, L.; Ma, X.; Zhao, X.; Jiao, B.; Li, C.; Luo, K. The WRKY transcription factors PtrWRKY18 and PtrWRKY35 promote Melampsora resistance in Populus. Tree Physiol. 2017, 37, 665–675. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Ran, L.; Hou, Y.; Tian, Q.; Li, C.; Liu, R.; Fan, D.; Luo, K. The transcription factor MYB115 contributes to the regulation of proanthocyanidin biosynthesis and enhances fungal resistance in poplar. New Phytol. 2017, 215, 351–367. [Google Scholar] [CrossRef]
- Fan, D.; Wang, X.; Tang, X.; Ye, X.; Ren, S.; Wang, D.; Luo, K. Histone H3K9 demethylase JMJ25 epigenetically modulates anthocyanin biosynthesis in poplar. Plant J. 2018, 96, 1121–1136. [Google Scholar] [CrossRef]



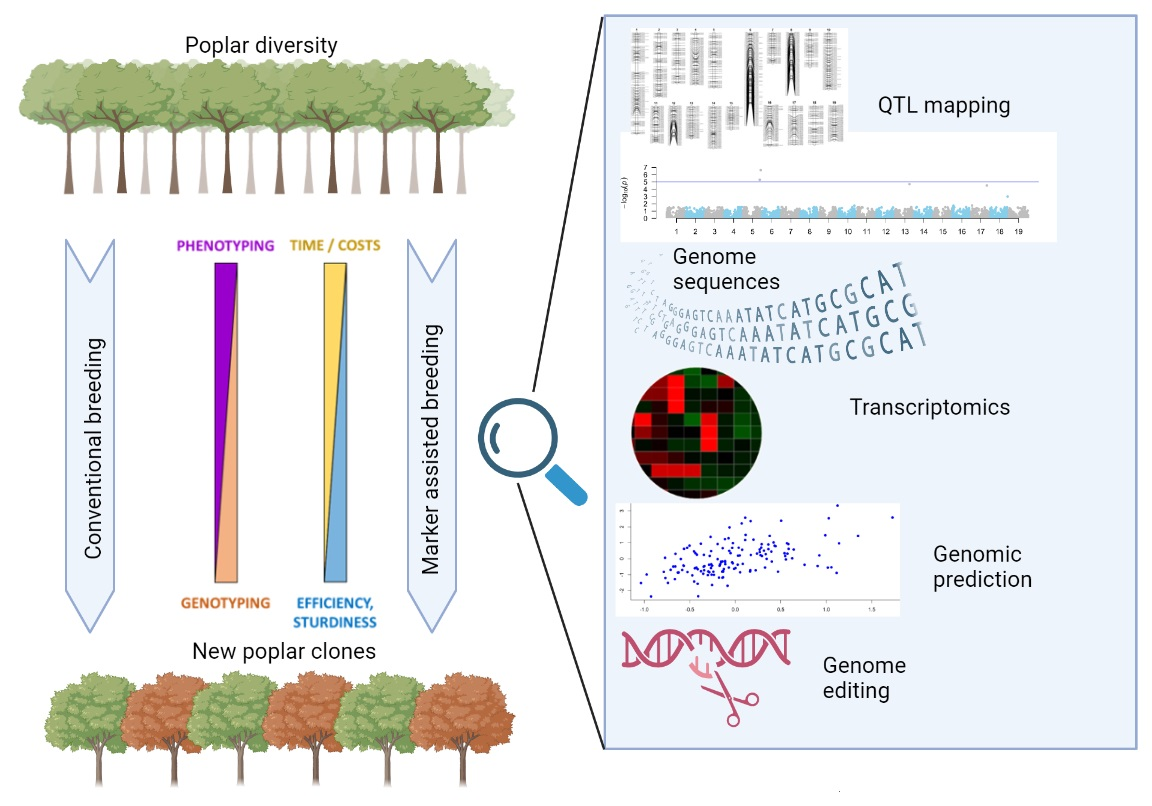{kind=link}
{kind=link}
{kind=link}
| Species | Section | Main Use |
|---|---|---|
| P. deltoides | Aigeiros | Female parent of P. ×canadiensis Parent of P. ×generosa |
| P. nigra | Aigeiros | Male parent of P. ×canadensis |
| P. alba | Populus | Parent of cross hybrids with P. tremuloides |
| P. tremula | Populus | Female parent of P. ×canescens Parent of cross hybrids with P. tremuloides |
| P. tremuloides | Populus | Parent of cross hybrids with P. tremula |
| P. trichocarpa | Tacamahaca | Parent of P. ×generosa |
| P. maximowiczii | Tacamahaca | Parent of cross hybrids with P. deltoides |
| Pathogen/Pest | Damages | Area | Incidence | Species | |
|---|---|---|---|---|---|
| Fungi | Alternaria alternata | Leaf blight | India, China | Not severe | Populus spp. |
| Apioplagiostoma populi | Bronze leaf | North America | Considerable | Populus spp. | |
| Armillaria spp. | Decline sectors of the crown, stunted vegetation | Atlantic-Mediterranean Europe, Southern Russia, Georgia, Syria, North Africa, tropical Africa, USA | Not severe | Populus spp. | |
| Botrydiplodia populea | Canker | China | Not severe | Populus spp. | |
| Ceratocystis fimbriata | Black or target canker | Alaska, USA, Quebec, Poland, India | Not severe | P. tremuloides, P. deltoides in India | |
| Cercospora populina | Leaf blotch | India | Considerable | P. deltoides | |
| Ciborinia whetzelii | Ink-spot disease | Canada, Northern USA | Not severe | P. tremuloides | |
| Cladosporium humile | Phylloptoses | India | Considerable | P. ciliata | |
| Corticium salmonicolor | Pink disease | India | Not severe | P. deltoides, P. ×euramericana, P. yunnanensis | |
| Cryptosphaeria lignyota | Snake canker and woody decay | Alaska, USA | Not severe | P. tremuloides (mainly), P. balsamifera, P. thrichocarpa, P. deltoides | |
| Cytospora spp. | Cytospora stem canker | Worldwide, mainly Central and Southern Italy, Eastern Europe, Near East, Northern India, and West-Central USA | Not severe, attacks occur under stresses or poor agronomic management | Aegiros, Tacamahaca, and Leuce sections | |
| Drechslera maydis | Leaf blight | India | Considerable | P. deltoides | |
| Diaporthe spp. | Phomosis stem canker | Germany, Italy, Argentina, Canada, Portugal, Japan, USA | Not severe | Aegeiros hybrids, P. deltoides, P. alba, P. ×euramericana, P. nigra, P. maximowiczii | |
| Diplodia tumefaciens | Bark alterations, woody galls | Northern Europe, Canada, Northern USA | Not severe | P. tremula, P. tremuloides, P. balsamifera | |
| Discosporium populeum | Canker | Worldwide, mainly Eurasia, North Africa, North America, and Argentina | Considerable | Aegiros section | |
| Dothichiza populea | Dothichiza stem canker | Europe, North America | Considerable | Populus spp. | |
| Dothiorella gregaria | Bark necrosis, blister canker, ulcer disease | China | Not severe | Populus spp. | |
| Encoelia pruinosa | Sooty-bark canker | Alaska, Western Canada, Mid-West USA, Northern Mexico, Norway | Considerable | P. tremuloides, P. balsamifera | |
| Erysiphe adunca | Powdery mildew | Italy | Not severe | P. nigra | |
| Gibberella spp. | Fusarium stem canker | Europe, North America | Not severe, limited to nurseries | Aegiros and Tacamahaca sections and intersectional hybrids, P. trichocarpa, P. ×euramericana | |
| Glomerella cingulata | Leaf and shoot blight | North-Western America, India, France | Not severe | Populus spp. | |
| Hypoxylon mammatum | Canker | North America, Europe | Considerable | P. tremuloides in North America, Leuce section in Europe | |
| Linospora spp. | Leaf blight | Eurasia, USA, Canada | Not severe | P. balsamifera, P. deltoides, P. trichocarpa ×P. deltoides (L. tetraspora), P. alba, P. tremula, P. tremuloides, P. grandidentata (L. ceuthocarpa) | |
| Marssonnina brunnea | Leaf spot | Worldwide | Considerable | Populus spp. | |
| Melampsora spp. | Leaf rust | Europe, Australia, New Zealand, Southern Africa, Argentina, North America, India, Japan, Canada | Considerable | Populus spp. | |
| Nectaria galligena | Canker | Central Europe, Rocky mountains | Not severe | P. tremuloides | |
| Phaeoramularia maculicola | Leaf spot | North America, Scandinavia, India | Not severe | Aegiros, Tacamahaca, and Leuce sections | |
| Phoma exigua | Cortical lesions | The Netherlands | Not severe | P. nigra, P. ×euramericana, P. trichocarpa | |
| Phyllactinia guttata | Powdery mildew | Southern Asia | Not severe | Euramerican poplars | |
| Phyllosticta spp. | Leaf spot | Europe, Argentina, Southern Australia, Japan, India | Not severe | Populus spp. | |
| Rhizoctonia solani | Leaf web blight | India | Considerable in nurseries and young plantations in humid conditions | Populus spp. | |
| Rhytidiella moriformis | Rough bark or cork bark | Canada | P. balsamifera, P. tremuloides | ||
| Rosellinia necatrix | Dematophora root rot | Worldwide, mainly Italy, Portugal, Southern Africa, and India | Considerable in intensive plantations in warm- temperate or sub-tropical climates | Populus spp. | |
| Septoria spp. | Canker and leaf spot | North-Central Europe, North America | Considerable | Aegiros × Tacamahaca | |
| Septotinia podophyllina | Leaf blotch | North America, France, Holland, ex-Yugoslavia, ex-Czechoslovakia, Russia, Korea, Japan | Not severe | Leuce, Aegiros, and Tacamahaca sections | |
| Sphaceloma populi | Anthracnoses | India, Europe, Japan, Argentina | Not severe | Aegiros and Tacamahaca sections | |
| Taphrina populina | Leaf blister | Worldwide | Not severe | Aegiros, Tacamahaca, and Leuce sections, P. alba | |
| Uncinula adunca | Powdery mildew | Eurasia, North America | Not severe | Leuce, Aegiros, and Tacamahaca sections | |
| Venturia spp. | Spring leaf and shoot blight | Eurasia, North America, North Africa, China | Considerable | Populus spp. | |
| Bacteria | Erwinia spp. | Bacterial twig canker | North America, Europe | Considerable | Populus spp. |
| Lonsdalea populi | Bark canker | Europe, China | Considerable | P. ×euramericana | |
| Phytophtora spp. | Root rot | Europe, Africa, USA, South America, Eastern Asia, Australia, New Zealand | Not severe | Populus spp. | |
| Pseudomonas syringae | Bacterial blight | Worldwide | Considerable | Populus spp. | |
| Sphingomonas spp. | Bark canker | Worldwide | Not severe | Populus spp. | |
| Xanthomonas populi | Canker | Europe, North America | Considerable | Aegiros and Tacamahaca sections | |
| Viruses | Arabis mosaic virus | Leaf mosaic | Europe, America, Japan, New Zealand | Considerable | P. ×euramericana |
| Poplar decline virus | Leaf spot, necrosis | America | Considerable | Populus spp. | |
| Poplar mosaic virus | Leaf mosaic | Worldwide | Considerable | P. nigra, P. deltoides, P. trichocarpa, P. candicans, P. ×euramericana | |
| Potato virus Y | Mottling or yellowing of leaflets, leaf crinkling, leaf drop | Worldwide | Not severe | P. tremuloides, Aegiros section | |
| Tobacco necrosis virus | Vein necrosis | Worldwide | Not severe | P. tremuloides | |
| Tobacco rattle virus | Mottling, chlorotic or necrotic local lesion, ringspots or line patterns, necrosis | Worldwide | Not severe | Populus spp. | |
| Tomato black ring virus | Mottling, deformation, leaf necrosis | Worldwide | Considerable | P. balsamifera | |
| Insects | Aceria parapopuli | Soap sucker, galls | North America | Can be considerable | Populus spp. |
| Agrilus suvorovi | Borer | Europe, Asia | Not severe, more considerable on one-year plants | P. tremula, P. deltoides, P. alba | |
| Altica populi | Defoliation | North America | Not severe | P. tacamahaca, P. tremuloides | |
| Anoplophora spp. | Borer | China, North America, Japan, Northern India, Pakistan | Considerable, the most important pests in China (A. nobilis and A. glabripennis), not severe in Japan (A. malasiaca) | Populus spp. | |
| Apriona spp. | Borer | Northern India, Pakistan, China, Japan | Can be considerable | Populus spp., P. ×euramericana (A. cinerea) | |
| Asymmetrasca decedens | Defoliation | Mediterranean areas, India | Not severe | Populus spp. | |
| Batocera lineolata | Borer | Japan | Considerable | Populus spp. | |
| Byctiscus populi | Defoliation | Europe | Can be considerable | P. deltoides and Euramerican hybrids | |
| Capnodis miliaris | Borer | Syria, Turkey, Iran, Iraq, Southern Italy | Considerable in drought conditions | Populus spp. | |
| Cerura spp. | Defoliation | Continental Europe, United Kingdom | Not severe | Populus spp. | |
| Choristoneura conflictana | Defoliation | Canada, Alaska, North-Eastern and Central USA | Considerable | P. tremuloides, P. deltoides, P. gradidentata | |
| Chrysomela spp. | Defoliation | Europe, North America | Considerable in young plantations and nurseries (C. populi and C. tremulae) | P. tremula × P. tremuloides, P. tremula × P. alba, P. alba | |
| Clostera spp. | Defoliation | Europe, Siberia, Japan, Korea, China, India, Pakistan | Considerable | P. tremula (mainly), P. euroamericana, P. euphratica, P. nigra | |
| Cossus cossus | Borer | Europe, North Africa | Can be considerable | Populus spp. | |
| Cryptorhynchus lapathi | Borer | Europe, China, Japan, USA, Canada | Considerable in young plantations and nurseries | Populus spp. | |
| Dasineura salicis | Galls | Europe, North America | Not severe | Populus spp. | |
| Epinotia solandriana | Defoliation | Europe, North America | Not severe in Europe, considerable in Canada for P. tremuloides | Populus spp. | |
| Gypsonoma spp. | Borer, leaf mining, galls | Europe, North Africa, North America, Pakistan | Not severe, can be considerable in young plantations and nurseries | P. deltoides (G. haimbachiana), P. euphratica (G. riparia) | |
| Halyomorpha halys | Borer | China, Japan, Taiwan, USA, Europe | Considerable | Populus spp. | |
| Hyphantria cunea | Defoliation | North America, Canada, Central and South-Eastern Europe, Japan, Korea | Considerable | Populus spp. | |
| Janus spp. | Defoliation | East USA, South Canada, Central and Southern Europe | Not severe | Populus spp. | |
| Leucoma spp. | Defoliation | Europe, Middle East, Japan, America, China | Not severe | Mainly P. alba, P. deltoides, P. nigra, P. x euramericana hybrids | |
| Megaplatypus mutatus | Tunnels in stems | South America, Europe | Can be considerable | Populus spp. | |
| Melanophila picta | Borer | Bulgaria, Spain, Southern France, Italy, Portugal, Pakistan, Turkey | Not severe, attacks occur only under water stress, more considerable on one-year plants | Populus spp., mainly P. ×euroamericana and P. euphratica in Iraq, P. nigra is less vulnerable | |
| Monosteira unicostata | Defoliation | Mediterranean areas, Turkey | Can be considerable in young plantations and nurseries | Populus spp. | |
| Mordwilkoja vagabunda | Galls | North America, Canada | Not severe | Populus spp., P. tremuloides in Canada | |
| Nematus spp. | Defoliation | Europe, South Africa, North America | Not severe | Populus spp., P. deltoides in South Africa | |
| Operophtera brumata | Defoliation | Europe, Asia, British Columbia, North America | Not severe, higher damages during drought stress | P. tremuloides, P. deltoides × P. nigra | |
| Orgyia spp. | Defoliation | Europe, North America, Japan, Korea, China, Russia | Can be severe | Populus spp. | |
| Paranthrene tabaniformis | Borer | Centre and southern Europe, North Africa, Asia (mainly China, Northern India, and Pakistan), Canada, Russia, Finland | Considerable in nurseries of one-year plants | Populus spp., mainly P. ×trichocarpa | |
| Parthenolecanium corni | Soap sucker | Europe, North America, New Zealand | Can be considerable | Populus spp. | |
| Phassus excrescens | Borer | Japan, Korea | Not severe | Populus spp. | |
| Phloeomyzus spp. | Soap sucker | Europe, North Africa, South America, China | Considerable | Populus spp., higher resistance for P. deltoides | |
| Phratora spp. | Defoliation | Europe, North America, Russia | Considerable in the event of outbreaks, especially in nurseries and young plantations | Populus spp., P. tremuloides in North America (P. purpurea purpurea) | |
| Phyllobius spp. | Defoliation | Europe, Russia, Iran, Turkey, North America | Not severe | Populus spp. | |
| Phyllocnistis spp. | Leaf mining | Europe, Canada | Not severe | P. nigra, P. deltoides × P. nigra | |
| Phyllonorycter spp. | Leaf mining | North America, Europe | Not severe | Populus spp., P. nigra (Europe) | |
| Phytobia spp. | Borer | Europe | Considerable | Populus spp. | |
| Platypus sulcatus | Borer | South America, mainly Argentina | Considerable | Populus spp. | |
| Polydrusus spp. | Defoliation | Spain, France, Italy, ex-Yugoslavia, Hungary, Eastern Canada, North-Eastern USA | Not severe | Populus spp. | |
| Popillia japonica | Defoliation | Japan, USA, Canada, China, Europe | Considerable | Populus spp. | |
| Porthetria spp. | Defoliation | Northern hemisphere | Not severe | Populus spp., mainly P. nigra (P. obfuscata) | |
| Saperda spp. | Borer | Europe, Asia, North America | Considerable, the main poplar pest in the Mediterranean basin (S. carcharias) | Populus spp. | |
| Sesia apiformis | Borer | Europe, Middle East, Asia Minor, China, North America, Canada | Not severe | Populus spp. | |
| Trichiocampus spp. | Defoliation | Europe, Middle East, North America, Japan | Can be considerable | P. deltoides, P. nigra var. italica, P. tremula, P. tremuloides | |
| Xyleborus dispar | Borer | Europe, North Africa, North America | Can be considerable | Populus spp. | |
| Yponomeuta rorrela | Defoliation | Europe | Not severe | Populus spp., mainly P. alba (Y. gigas) | |
| Zeuzera pyrina | Borer | Central Europe, Mediterranean basin, Asia, India, Japan, North America, South Africa | Can be considerable | Populus spp. | |
| Hybrid | Cross |
|---|---|
| P. ×canadiensis | P. deltoides × P. nigra |
| P. ×generosa | P. trichocarpa × P. deltoides |
| P. ×tomentosa | P. alba × P. tremula |
| P. ×wettsteinii | P. tremula × P. tremuloides |
| P.×interamericana | P. deltoides × P. trichocarpa |
| Species | Total Size (Mb) | Coverage (X) | n. Scaffolds | N50 Scaffold Size (kb) | N50 Contig Size (kb) | Chromosomes (%) | Repetitive Elements (%) | Protein- Coding Genes | Non-Coding RNAs | References |
|---|---|---|---|---|---|---|---|---|---|---|
| P. trichocarpa | 423 | 9.44 | 1446 | 19,500 | 552.8 | 84.53 | 48.07 | 42,950 | 817 tRNAs, 88 snRNAs, 427 snoRNAs, 169 miRNAs | [6] |
| P. euphratica | 496.5 | 312 | 9673 | 482 | 40.438 | 44 | 34,279 | 764 tRNAs, 706 rRNAs, 4826 snRNAs, 266 miRNAs | [56] | |
| P. euphratica | 574.35 | 246.54 | 507 | 28.59 | 900 | 98.85 | 56.95 | 36,606 | 8767 long non-coding RNAs | [55] |
| P. pruinosa | 479 | 125 | 78,960 | 698.5 | 14 | 45.47 | 35,131 | [54] | ||
| P. tremula | 390 | 98 | 216,318 | 42.844 | 21.54 | 35,984 | [4] | |||
| P. tremuloides | 378 | 86.35 | 164,504 | 15.222 | 22.09 | 36,830 | [4] | |||
| P. alba var. pyramidalis | 466 | 320 | 17,797 | 459.178 | 26.535 | 44.61 | 37,901 | 940 tRNAs, 569 rRNAs, 123 snRNAs, 1050 miRNAs | [3] | |
| P. alba | 416 | 130 | 1285 | 1180 | 45.16 | 32,963 | 764 tRNAs, 706 rRNAs, 4826 snRNAs, 266 miRNAs | [57] | ||
| P. simonii | 441 | 138 | 686 | 194 | 90.2 | 41.47 | 45,459 | 1177 tRNAs, 290 rRNAs, 618 snRNAs, 1153 miRNAs | [58] | |
| P. deltoides W94 | 446.8 | 62.94 | 1375 | 21,700 | 590.2 | 90.2 | 44,853 | |||
| P. deltoides I-69 | 429 | 273 | 934 | 21,500 | 2620 | 97.4 | 32.28 | 44,853 | [59] | |
| Poplar 84K (P. alba × P. tremula var. glandulosa) | 747.5 | 119.79 | 1384 | 19,600 | 1990 | 94.98 | 24.40 | 85,755 | 1312 tRNAs, 1140 rRNAs, 1126 snRNAs, 1983 miRNAs | [60] |
| Poplar 84K (P. alba × P. tremula var. glandulosa) | 781.36 (405.31 subgenome A; 376.05 subgenome G) | 189 | 2109 (1179 subgenome A; 930 subgenome G) | 3660 (5430 subgenome A; 2150 subgenome G) | 43.7 subgenome A; 40.5 subgenome G | 38,701 subgenome A; 38,449 subgenome G | [61] |
| Pathogen | Genotypes | QTL/Locus Name | Markers | LG/Chr./Sc. | bp | cM | Markers | Candidate Genes | Reference |
|---|---|---|---|---|---|---|---|---|---|
| Septoria populicola | F2 331 (107 individuals): P. trichocarpa (93-968, R) × P. deltoides (ILL-129, S) | FLD94 | P1064-B15_17 | LG X | RFLP, RAPD, STS-343 | [77] | |||
| FLD94 | P13292-P1043 | LG M | |||||||
| FLD95 | P1064-B15_17 | LG X | |||||||
| FLD95 | P1322-P1310 | LG A | |||||||
| Melampsora medusae | F2 331 (107 individuals): P trichocarpa (93-968, R) × P. deltoides (ILL-129, S) | Mmd1 | P222 | LG Q | 5.1 cM from P222 | RFLP, RAPD, STS-343 | [78] | ||
| Melampsora medusae | F1 C9425DD (116 individuals): P. deltoides (7300501, S) × P. deltoides (7302801, R) | Lrd1 | OPG10340 OPZ191800 | 2.6 cM from OPG10340 7.4 cM from OPZ191800 | RAPD-84 | [79] | |||
| Melampsora Xcolumbiana | F1 545 (1902 individuals): P. trichocarpa (clone 383-2499, R) × P. deltoides (clone 14-101, S) | MXC3 | CGA.TCT_01 GAC.TAC_01 | 0.68-2.05 | AFLP–19 linked | [13] | |||
| Melampsora larici-populina | F1 87001 (139 individuals) and 95001 (77 individuals): P. deltoides (S9-2, R) × P. nigra (Ghoy, S); F1 87002 (106 individuals) and 95002 (120 individuals): P. deltoides (S9-2, R) × P. trichocarpa (V24, S); backcross 95003 (70 individuals): (P. deltoides (S9-2) × P. nigra (Ghoy), R) × P. nigra (Ghoy, S) | MER | E40G37 E39G01 E44G09 E32G43 E45G29 E47G14 E39F39r E48G14 E61G36 E51G05 E43G28 | 3.4 (interval) | AFLP–11 linked | AF393736_NBS/LRR AF393738_NBS/LRR AF393739_NBS/LRR | [80] | ||
| Melampsora larici-populina | F1 13 (171 individuals): clone 52-225 (P. trichocarpa 93-968 × P. deltoides ILL-101, R) × P. deltoides (D109, S) | MXC3 | STS1_A, STS3, O_349, O_356 | LG IV; Chr. 4 | 580,744-713,007 | 4.1 | SSR, STS, AFLP-588 | NP_195325.1_thaumatin NP_173432.2_thaumatin NP_197963.1_disease-resistance protein NP_177296.1_disease-resistance protein (LRR) T10504_disease-resistance protein Cf-2.1 T10504 disease-resistance protein Cf-2.1 | [73] |
| MER | T4_3, S2_19, R_7, O_206, S1_8 | LG XIX; Chr. 19 | 13,586,903(O_206) | 33.6 | |||||
| Melampsora larici-populina | F1 (343 individuals): P. deltoides (73028-62, R) × P. trichocarpa (101-74, S) | E4M1-4 | LG TXI | 0 | AFLP, RAPD, SSR, SNP, RFLP, phenotypic markers-391 | [81] | |||
| E2M6-42 | LG DIII | 24.4 | |||||||
| E2M4-16 | LG D? | 0 | |||||||
| Rus | LG T? | 10.2 | |||||||
| Rus | LG T? | 11.1 | |||||||
| Rus | LG T? | 13.1 | |||||||
| Rus | LG T? | 15.1 | |||||||
| Rus | LG T? | 17.1 | |||||||
| Rus | LG T? | ||||||||
| E5M5-4 | LG TXII | ||||||||
| E5M5-7 | LG T? | 6.0 | |||||||
| E5M5-7 | LG T? | 8.2 | |||||||
| E5M5-7 | LG T? | 6.0 | |||||||
| E5M5-7 | LG T? | 8.0 | |||||||
| E4M4-10 | LG DVI | 158.4 | |||||||
| rE1M4-8 | LG TXII | 66.3 | |||||||
| E4M4-10 | LG DVI | 168.4 | |||||||
| rE2M4-10 | LG DIII | 141.4 | |||||||
| rORPM277 | LG DXIX | 133.0 | |||||||
| rORPM277 | LG DXIX | 137.0 | |||||||
| R1 | LG DXIX | 144.0 | |||||||
| R1 | LG DXIX | 145.0 | |||||||
| E1M2-8 | LG DXIX | 117.0 | |||||||
| E1M2-8 | LG DXIX | 125.0 | |||||||
| Melampsora larici-populina | F1 (1415 individuals): P. deltoides (73028-62, R) × P. trichocarpa (101-74, S) | R1 | G_79–I_920-3 | Chr.19 | 16,965,396–17,119,994 | 3.90–4.00 | SSR, STS, AFLP, RAPD-68 | BED-NBS-LRR TIR-NBS-LRR Serine threonine kinase | [82] |
| Rus | Is_165_1–RGAs297 | Chr.19 | 5.50–6.00 | TIR-NBS-LRR | |||||
| Melampsora larici-populina | R1 | I_1211–I_920_3 | Chr.19 | 16,965,396–17,119,994 | Fragment-based | [83] | |||
| Rus | 14N08-F–RGAs135-1 | Chr.19 | 16,441,457–16,460,757 | EVM0026813_TNL | |||||
| Mer | O_263–O_206 | Chr.19 | 13,586,903–15,058,693 | EVM0004305_CNL EVM0025825_STK | |||||
| Melampsora larici-populina | F1 (300 individuals): P. deltoides (I-69, R) × P. simonii (XYY, S) | 201709ab | Chr02-5594026 | LG 2, Chr.2 | 5,594,026 | 70.49; 65.49–75.49 | SNP–1222 | [84] | |
| 201707ab | Chr04-12165533 | LG 4, Chr.4 | 12,165,533 | 76.57; 71.57–81.57 | |||||
| 201709p2 | LG 8, Chr.8 | 191.79; 78.00–210.00 | |||||||
| 201809p2 | Chr14-18570439 | LG 9, Chr.9 | 0.00; 0.00–2.00 | ||||||
| 201809p2 | LG 10, Chr.10 | 228.24; 226.58–252.00 | |||||||
| 201709p2 | LG 12, Chr.12 | 146.0; 142.00–150.00 | |||||||
| 201809p2 | LG 13, Chr.13 | 70.00; 65.00–75.00 | |||||||
| LRDR-QTL-17 (overlapped region of 201709p2 and 201809p2) | Chr17-11257300-Chr17-12346306 | LG 17, Chr.17 | 11,257,300–12,346,306 | 65.67; 64.00–68.00; 66.00; 62.72–72.00 | |||||
| Potri.017G104100 15 disease-resistance genes | |||||||||
| LRDR-QTL-19 (overlapped region of 201707p2 and 201809p2) | Chr19-1553413 | LG 19, Chr.19 | 1,553,413 | 23.19; 18.19–28.19; 21.19; 19.19–26.78 | 21 disease-resistance genes | ||||
| Melamspora xcolumbiana | Collection P. trichocarpa (456 individuals) | 23949327 | Sc.5 | 23,949,327 | SNP–34K | IQD32 | [85] | ||
| 1402770 | Sc.6 | 1,402,770 | FAR1 | ||||||
| 8261867 | Sc.8 | 8,261,867 | PIP5K | ||||||
| 19215715 | Sc.10 | 19,215,715 | PRR7 | ||||||
| 2955 | Sc.143 | 2955 | NRT2.4 | ||||||
| Insects | F2 331 (350 individuals): P. trichocarpa (93-968) × P. deltoides (ILL-129) | LG Vb; Chr. 5 | 4; 0–19 | SSR, AFLP-183 | [74] | ||||
| LG I; Chr.1 | 5,467,692(PMGC_634); 6,435,691(PMGC_2789) | 9; 0–24 | 4 PG genes | ||||||
| LG XIV; Chr. 14 | 0; 0–28 | 1 PG gene | |||||||
| LG III; Chr.3 | 9,528,665(ORPM_30); 10,796,665(ORPM_203) | 37; 29–46 | |||||||
| LG IV; Chr.4 | 13,371,978(PMGC_2826) | 59; 45–85 | 1 PG gene | ||||||
| LG Va; Chr. 5 | 76; 62–86 | 1 PG gene | |||||||
| LG XVII; Chr. 17 | 54; 35–69 | 2 PG genes | |||||||
| LG Va; Chr. 5 | 19; 0–41 | 1 PG gene | |||||||
| LG VIIIa; Chr. 8 | 27; 12–27 | ||||||||
| LG XVII; Chr. 17 | 50; 33–70 | 2 PG genes | |||||||
| LG I; Chr.1 | 9,764,020(PMGC_2852), 11,239,328(PMGC_93) | 74; 32–125 | 2 PG genes | ||||||
| LG VI; Chr. 6 | 144; 134–144 | ||||||||
| LG XII; Chr. 12 | 4,407,861(WPMS_3), 12,292,045(PMGC_2885) | 17; 0–24 | 1 PG gene | ||||||
| LG III; Chr.3 | 6,609,278(PMGC_2501) | 14; 0–31 | |||||||
| Phloeomyzus passerinii L. | F1 (131 individuals): P. deltoides (D0-092b, R) × P. nigra (N074, S) | wpa-5 | 5_2426240 | LG V, Chr.5 | 1,975,251–2,578,834 | 43.7 | SNP, SSR–5667 | NPK1-related protein kinase 1 | [7] |
| wpa-16 | 16_3345538, 16_3345877 | LG XVI, Chr.16 | 2,980,973–3,749,017 | 43.4 | CCCH-type zinc finger protein with ARM repeat domain | ||||
| wpa-19 | 78_83250, 78_83287, 78_83295 | LG XIX, Chr.19 | 2,071,803–3,238,172 | 44.8 | 14 TIR-NB-LRR disease-resistance genes Phospholipase A2 | ||||
| Schizoempodium mesophyllincola | F2 331 (376 individuals): P. trichocarpa (93–968, R) × P. deltoides (ILL-129, S) | ORPM_26 | LGIII; Chr.6 | 6,013,759–6,013,972 | 33.642–59.393 | AFLP, RAPD, RFLP, SSR-841 | [75] | ||
| PMGC_2889B | LGI; Chr.1 | 108.686–118.167 | |||||||
| WPMS_15-PMGC_2839 | LGV; Chr. 5 | 23,655,307–25,782,064 | 63.447–76.062 | ||||||
| Sphaerulina musiva | Collection P. trichocarpa (1081 individuals) | Chr.3 | 3,517,268 | SNP–8,253,066 | Potri.003G028200_RLP | [86] | |||
| Chr.5 | 942,545 | Potri.005G012100_RLP | |||||||
| Chr.5 | 1,440,266 | Potri.005G018000_G-type lecRLK | |||||||
| Chr.9 | 4,548,711 | Potri.009G036300_L-type lecRLK | |||||||
| Biotic stress, Sphaerulina musiva responsive | Collection P. trichocarpa (917 individuals) | PtHCT2 | Chr18:13249087 | Chr. 18 | 13,222,67–13,252,693 | SNP–8,253,066 | Potri.018G105500 | [86,87] | |
| Marsonnina brunnea | F1 (84 individuals): P. deltoides (Zhongshi-8, R) × P. deltoides (D-124, S) | qMLSD-VI-1 | P_2217-G_2034 | LG VI, Chr.6 | 16,592,305–17,904,816 | 118.2; 92.2–137.8 | SSR, AFLP–1398 | Potri.006G164600.1Potri.006G171300.1 Potri.006G166700.1Potri.006G166800.1 | [76] |
| qMLSD-XVI-2 | P_2143–P_204 | LG XVI, Chr.16 | 10,022,916–12,773,381 | 138.3; 128.3–144.6 | Potri.016G115800.1Potri.016G115900.1 Potri.016G116000.1Potri.016G116100.1 Potri.016G114400.1Potri.016G107200.1 Potri.016G109200.1Potri.016G122700.1 Potri.016G123300.1Potri.016G123500.1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Biselli, C.; Vietto, L.; Rosso, L.; Cattivelli, L.; Nervo, G.; Fricano, A. Advanced Breeding for Biotic Stress Resistance in Poplar. Plants 2022, 11, 2032. https://doi.org/10.3390/plants11152032
Biselli C, Vietto L, Rosso L, Cattivelli L, Nervo G, Fricano A. Advanced Breeding for Biotic Stress Resistance in Poplar. Plants. 2022; 11(15):2032. https://doi.org/10.3390/plants11152032
Chicago/Turabian StyleBiselli, Chiara, Lorenzo Vietto, Laura Rosso, Luigi Cattivelli, Giuseppe Nervo, and Agostino Fricano. 2022. "Advanced Breeding for Biotic Stress Resistance in Poplar" Plants 11, no. 15: 2032. https://doi.org/10.3390/plants11152032
APA StyleBiselli, C., Vietto, L., Rosso, L., Cattivelli, L., Nervo, G., & Fricano, A. (2022). Advanced Breeding for Biotic Stress Resistance in Poplar. Plants, 11(15), 2032. https://doi.org/10.3390/plants11152032