
Visible Foliar Injury and Ecophysiological Responses to Ozone and Drought in Oak Seedlings

 ,
,  and
and
Abstract
:
1. Introduction
2. Materials and Methods
2.1. Plant Material and Experimental Setting
2.2. Evaluation of O3 and Drought Visible Foliar Injury
2.3. Measure of Growth Parameters
2.4. Assessment of Photosynthetic Parameters
2.5. Calculation of Accumulated Drought and Ozone Indexes
2.6. Statistical Analysis
3. Results
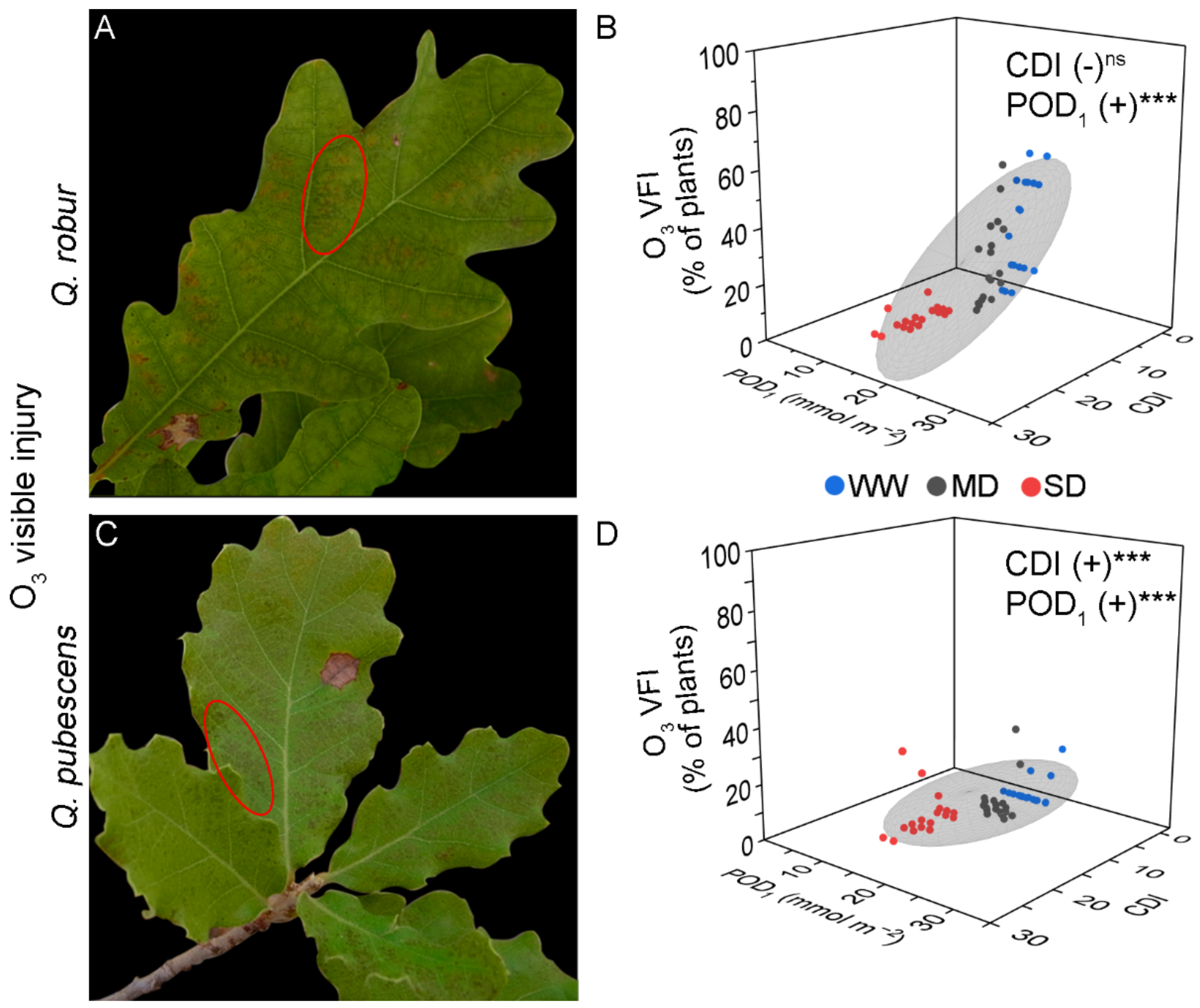
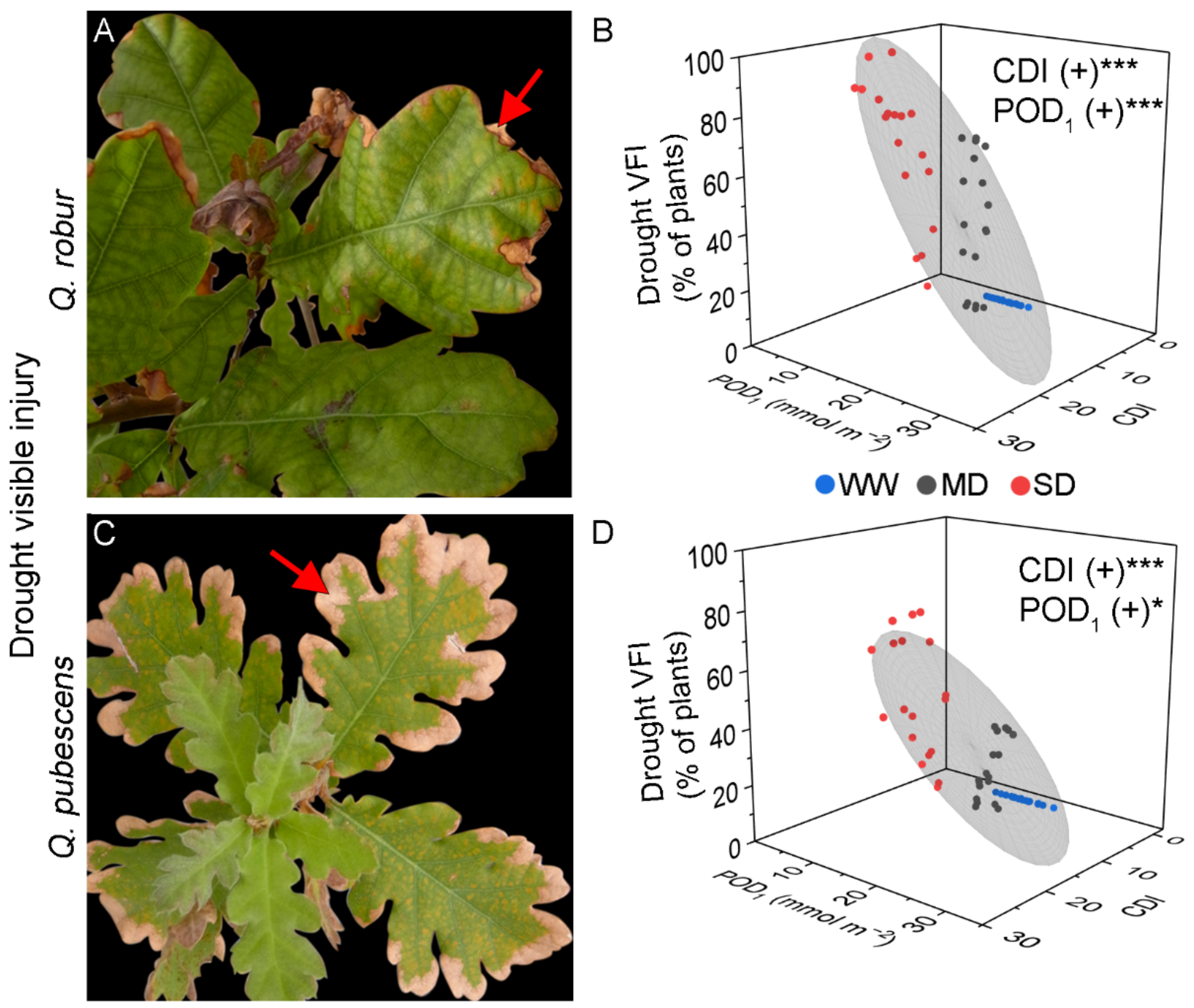
3.1. Visible Foliar Injury
3.2. Physiological Responses
3.3. Growth and Biomass
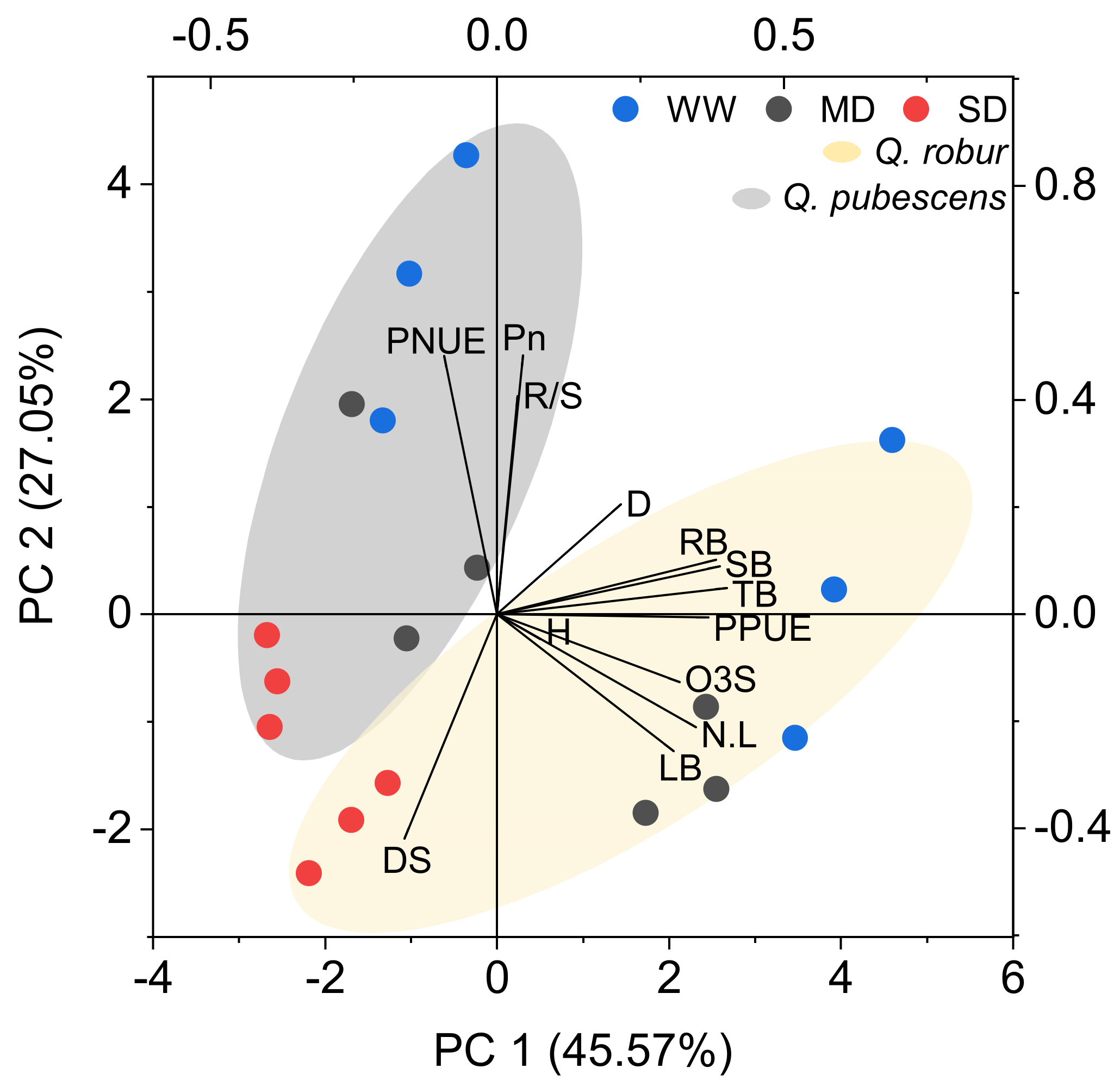
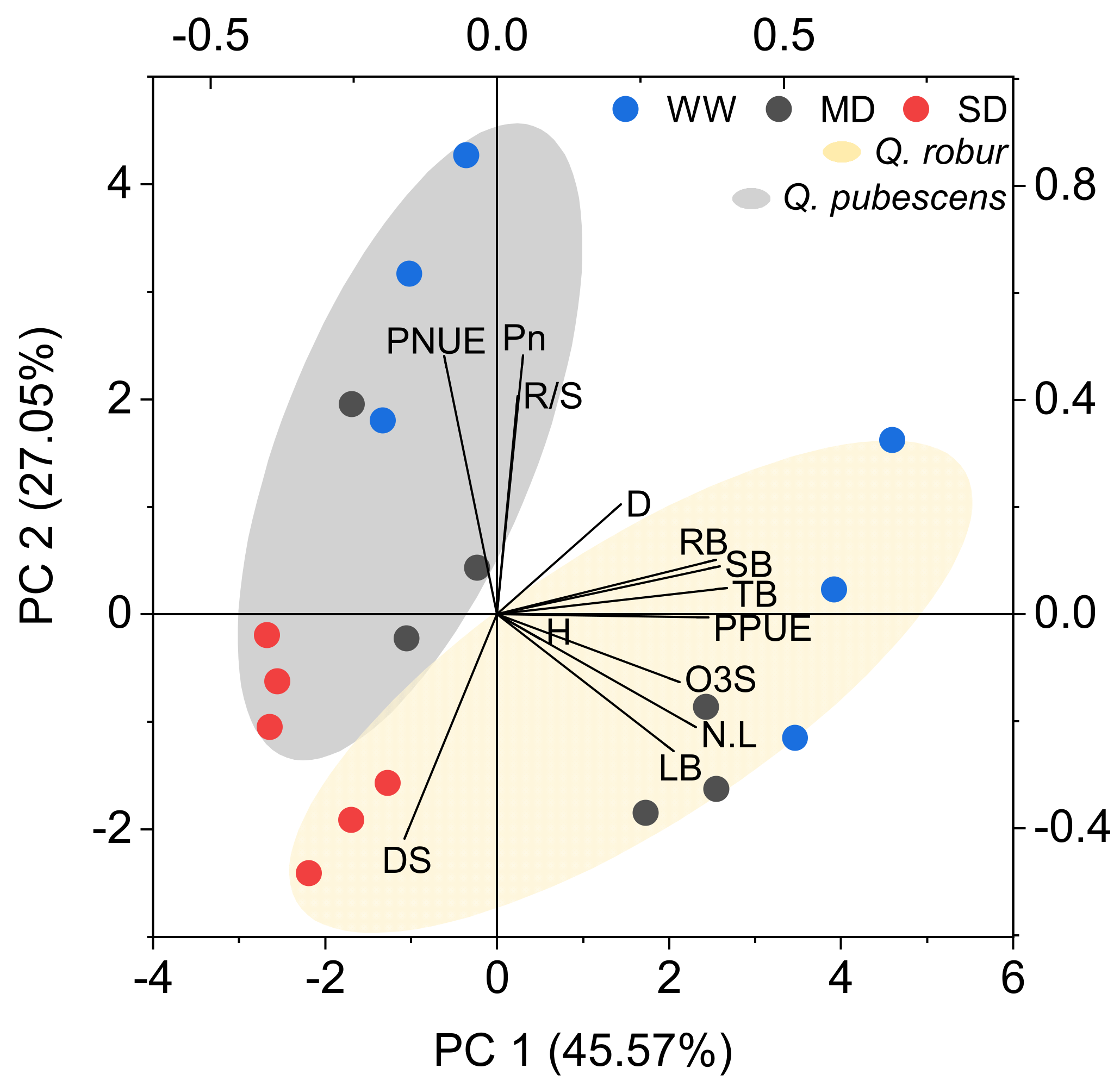
3.4. Principal Component Analysis
4. Discussion
4.1. Development of Visible Injury Due to Ozone and Drought Stress
4.2. Effects of Ozone and Drought Stress on Growth and Biomass Parameters
4.3. Effects of Ozone and Drought Stress on Physiological Parameters
4.4. Is the Ozone Visible Injury an Indicator to Explain Biomass Reduction or Physiological Damage in Mediterranean Oaks?
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Mills, G.; Pleijel, H.; Malley, C.S.; Sinha, B.; Cooper, O.R.; Schultz, M.G.; Neufeld, H.S.; Simpson, D.; Sharps, K.; Feng, Z.; et al. Tropospheric Ozone Assessment Report: Present-Day Tropospheric Ozone Distribution and Trends Relevant to Vegetation. Elementa 2018, 6, 47. [Google Scholar] [CrossRef]
- Paoletti, E. Ozone Impacts on Forests. CAB Rev. Perspect. Agric. Vet. Sci. Nutr. Nat. Resour. 2007, 2, 13. [Google Scholar] [CrossRef]
- Dusart, N.; Gandin, A.; Vaultier, M.N.; Joffe, R.; Cabané, M.; Dizengremel, P.; Jolivet, Y. Importance of Detoxification Processes in Ozone Risk Assessment: Need to Integrate the Cellular Compartmentation of Antioxidants? Front. For. Glob. Chang. 2019, 2, 45. [Google Scholar] [CrossRef] [Green Version]
- Grulke, N.E.; Heath, R.L. Ozone Effects on Plants in Natural Ecosystems. Plant Biol. 2020, 22, 12–37. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, M.; Hoshika, Y.; Inada, N.; Wang, X.; Mao, Q.; Koike, T. Photosynthetic Traits of Siebold’s Beech and Oak Saplings Grown under Free Air Ozone Exposure in Northern Japan. Environ. Pollut. 2013, 174, 50–56. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hoshika, Y.; Brilli, F.; Baraldi, R.; Fares, S.; Carrari, E.; Zhang, L.; Badea, O.; Paoletti, E. Ozone Impairs the Response of Isoprene Emission to Foliar Nitrogen and Phosphorus in Poplar. Environ. Pollut. 2020, 267, 115679. [Google Scholar] [CrossRef]
- Büker, P.; Feng, Z.; Uddling, J.; Briolat, A.; Alonso, R.; Braun, S.; Elvira, S.; Gerosa, G.; Karlsson, P.E.; Le Thiec, D.; et al. New Fl Ux Based Dose e Response Relationships for Ozone for European Forest Tree Species. Environ. Pollut. 2015, 206, 163–174. [Google Scholar] [CrossRef]
- Hoshika, Y.; Paoletti, E.; Agathokleous, E.; Sugai, T.; Koike, T. Developing Ozone Risk Assessment for Larch Species. Front. For. Glob. Chang. 2020, 3, 45. [Google Scholar] [CrossRef]
- CLRTAP. Mapping Critical Levels for Vegetation, Chapter III. Manual on Methodologies and Criteria for Modelling and Mapping Critical Loads and Levels and Air Pollution Effects, Risks and Trends. In UNECE Convention on Long-Range Transboundary Air Pollution; UNECE: Geneva, Switzerland, 2017. [Google Scholar]
- Sicard, P.; De Marco, A.; Carrari, E.; Dalstein-Richier, L.; Hoshika, Y.; Badea, O.; Pitar, D.; Fares, S.; Conte, A.; Popa, I.; et al. Epidemiological Derivation of Flux-Based Critical Levels for Visible Ozone Injury in European Forests. J. For. Res. 2020, 31, 1509–1519. [Google Scholar] [CrossRef]
- Bagard, M.; Jolivet, Y.; Hasenfratz-Sauder, M.-P.; Gérard, J.; Dizengremel, P.; Le Thiec, D. Ozone Exposure and Flux-Based Response Functions for Photosynthetic Traits in Wheat, Maize and Poplar. Environ. Pollut. 2015, 206, 411–420. [Google Scholar] [CrossRef]
- Schaub, M.; Calatayud, V.; Ferretti, M.; Brunialti, G.; Lövblad, G.; Krause, G.; Sanz, M.J. Part VIII: Monitoring of Ozone Injury. In Manual on Methods and Criteria for Harmonized Sampling, Assessment, Monitoring and Analysis of the Effects of Air Pollution on Forests; UNECE ICP Forests Programme Coordinating Centre, Ed.; Thünen Institute of Forest Ecosystems: Eberswalde, Germany, 2016; p. 14. [Google Scholar]
- Sicard, P.; De Marco, A.; Dalstein-Richier, L.; Tagliaferro, F.; Renou, C.; Paoletti, E. An Epidemiological Assessment of Stomatal Ozone Flux-Based Critical Levels for Visible Ozone Injury in Southern European Forests. Sci. Total Environ. 2016, 541, 729–741. [Google Scholar] [CrossRef]
- Sicard, P.; Hoshika, Y.; Carrari, E.; De Marco, A.; Paoletti, E. Testing Visible Ozone Injury within a Light Exposed Sampling Site as a Proxy for Ozone Risk Assessment for European Forests. J. For. Res. 2021, 32, 1351–1359. [Google Scholar] [CrossRef]
- Hanjra, M.A.; Qureshi, M.E. Global Water Crisis and Future Food Security in an Era of Climate Change. Food Policy 2010, 35, 365–377. [Google Scholar] [CrossRef]
- Xu, C.; McDowell, N.G.; Fisher, R.A.; Wei, L.; Sevanto, S.; Christoffersen, B.O.; Weng, E.; Middleton, R.S. Increasing Impacts of Extreme Droughts on Vegetation Productivity under Climate Change. Nat. Clim. Chang. 2019, 9, 948–953. [Google Scholar] [CrossRef] [Green Version]
- Peters, M.P.; Iverson, L.R.; Matthews, S.N. Spatio-Temporal Drought Trends by Forest Type in the Conterminous United States, 1960–2013; US Department of Agriculture, Forest Service, Northern Research Station: Madison, WI, USA, 2014.
- Vollenweider, P.; Ottiger, M.; Günthardt-Goerg, M.S. Validation of Leaf Ozone Symptoms in Natural Vegetation Using Microscopical Methods. Environ. Pollut. 2003, 124, 101–118. [Google Scholar] [CrossRef]
- Moura, B.B.; Alves, E.S.; Marabesi, M.A.; de Souza, S.R.; Schaub, M.; Vollenweider, P. Ozone Affects Leaf Physiology and Causes Injury to Foliage of Native Tree Species from the Tropical Atlantic Forest of Southern Brazil. Sci. Total Environ. 2018, 610–611, 912–925. [Google Scholar] [CrossRef]
- Vollenweider, P.; Menard, T.; Arend, M.; Kuster, T.M.; Günthardt-Goerg, M.S. Structural Changes Associated with Drought Stress Symptoms in Foliage of Central European Oaks. Trees—Struct. Funct. 2016, 30, 883–900. [Google Scholar] [CrossRef]
- Alonso, R.; Elvira, S.; González-Fernández, I.; Calvete, H.; García-Gómez, H.; Bermejo, V. Drought Stress Does Not Protect Quercus Ilex L. from Ozone Effects: Results from a Comparative Study of Two Subspecies Differing in Ozone Sensitivity. Plant Biol. 2014, 16, 375–384. [Google Scholar] [CrossRef]
- Hoshika, Y.; Moura, B.; Paoletti, E. Ozone Risk Assessment in Three Oak Species as Affected by Soil Water Availability. Environ. Sci. Pollut. Res. 2018, 25, 8125–8136. [Google Scholar] [CrossRef]
- Cocozza, C.; Paoletti, E.; Mrak, T.; Zavadlav, S.; Levanič, T.; Kraigher, H.; Giovannelli, A.; Hoshika, Y. Isotopic and Water Relation Responses to Ozone and Water Stress in Seedlings of Three Oak Species with Different Adaptation Strategies. Forests 2020, 11, 864. [Google Scholar] [CrossRef]
- Hoshika, Y.; Fares, S.; Pellegrini, E.; Conte, A.; Paoletti, E. Water Use Strategy Affects Avoidance of Ozone Stress by Stomatal Closure in Mediterranean Trees—A Modelling Analysis. Plant Cell Environ. 2020, 43, 611–623. [Google Scholar] [CrossRef]
- Pellegrini, E.; Hoshika, Y.; Dusart, N.; Cotrozzi, L.; Gérard, J.; Nali, C.; Vaultier, M.N.; Jolivet, Y.; Lorenzini, G.; Paoletti, E. Antioxidative Responses of Three Oak Species under Ozone and Water Stress Conditions. Sci. Total Environ. 2019, 647, 390–399. [Google Scholar] [CrossRef] [PubMed]
- Paoletti, E.; Carriero, G. A New-Generation 3D Ozone FACE (Free Air Controlled Exposure). Sci. Total Environ. 2017, 575, 1407–1414. [Google Scholar] [CrossRef] [PubMed]
- Moura, B.B.; Hoshika, Y.; Ribeiro, R.V.; Paoletti, E. Exposure- and Flux-Based Assessment of Ozone Risk to Sugarcane Plants. Atmos. Environ. 2018, 176, 252–260. [Google Scholar] [CrossRef]
- Innes, J.L.; Skelly, J.M.; Schaub, M. Ozone and Broadleaved Species: A Guide to the Identification of Ozone-Induced Foliar Injury/ Ozon, Laubholz-Und Krautpflanzen: Ein Fuhrer Zum Bestimmen von Ozonsymptomen; Haupt: Bern, Switzerland, 2001. [Google Scholar]
- Vollenweider, P.; Gunthardtgoerg, M. Erratum to “Diagnosis of Abiotic and Biotic Stress Factors Using the Visible Symptoms in Foliage”. Environ. Pollut. 2006, 140, 562–571. [Google Scholar] [CrossRef]
- Günthardt-Goerg, M.S.; Kuster, T.M.; Arend, M.; Vollenweider, P. Foliage Response of Young Central European Oaks to Air Warming, Drought and Soil Type. Plant Biol. 2013, 15, 185–197. [Google Scholar] [CrossRef]
- Chappelka, A.; Renfro, J.; Somers, G.; Nash, B. Evaluation of Ozone Injury on Foliage of Black Cherry (Prunus Serotina) and Tall Milkweed (Asclepias Exaltata) in Great Smoky Mountains National Park. Environ. Pollut. 1997, 95, 13–18. [Google Scholar] [CrossRef]
- Team R Development Core. A Language and Environment for Statistical Computing. 2018. Available online: https://www.R-project.org/ (accessed on 21 May 2022).
- Fernandes, F.F.; Moura, B.B. Foliage Visible Injury in the Tropical Tree Species, Astronium Graveolens Is Strictly Related to Phytotoxic Ozone Dose (PODy). Environ. Sci. Pollut. Res. 2021, 28, 41726–41735. [Google Scholar] [CrossRef]
- Gao, F.; Catalayud, V.; Paoletti, E.; Hoshika, Y.; Feng, Z. Water Stress Mitigates the Negative Effects of Ozone on Photosynthesis and Biomass in Poplar Plants. Environ. Pollut. 2017, 230, 268–279. [Google Scholar] [CrossRef]
- Grulke, N.E. The Physiological Basis of Ozone Injury Assessment Attributes in Sierran Conifers. Dev. Environ. Sci. 2003, 2, 55–81. [Google Scholar] [CrossRef]
- Yoshimura, K. Programmed Proteome Response for Drought Avoidance / Tolerance in the Root of a C 3 Xerophyte (Wild Watermelon) Under Water Deficits. Plant Cell Physiol. 2008, 49, 226–241. [Google Scholar] [CrossRef] [Green Version]
- Le Gall, H.; Philippe, F.; Domon, J.M.; Gillet, F.; Pelloux, J.; Rayon, C. Cell Wall Metabolism in Response to Abiotic Stress. Plants 2015, 4, 112. [Google Scholar] [CrossRef]
- Pinheiro, C.; Chaves, M.M. Photosynthesis and Drought: Can We Make Metabolic Connections from Available Data? J. Exp. Bot. 2011, 62, 869–882. [Google Scholar] [CrossRef] [Green Version]
- Pell, E.J.; Sinn, J.P.; Johansen, C.V. Nitrogen Supply as a Limiting Factor Determining the Sensitivity of Populus Tremuloides Michx. to Ozone Stress. New Phytol. 1995, 130, 437–446. [Google Scholar] [CrossRef]
- Matyssek, R.; Sandermann, H. Impact of Ozone on Trees: An Ecophysiological Perspective. In Progress in Botany; Esser, K., Lüttge, U., Beyschlag, W., Hellwig, F., Eds.; Springer: Berlin/Heidelberg, Germany, 2003; ISBN 978-3-642-55819-1. [Google Scholar]
- Jaleel, C.A.; Manivannan, P.; Wahid, A.; Farooq, M.; Al-Juburi, H.J.; Somasundaram, R.; Panneerselvam, R. Drought Stress in Plants: A Review on Morphological Characteristics and Pigments Composition. Int. J. Agric. Biol. 2009, 11, 100–105. [Google Scholar]
- Chaturvedi, R.K.; Tripathi, A.; Raghubanshi, A.S.; Singh, J.S. Functional Traits Indicate a Continuum of Tree Drought Strategies across a Soil Water Availability Gradient in a Tropical Dry Forest. For. Ecol. Manag. 2021, 482, 118740. [Google Scholar] [CrossRef]
- Moura, B.B.; Alves, E.S. Climatic Factors Influence Leaf Structure and Thereby Affect the Ozone Sensitivity of Ipomoea Nil “Scarlet O’Hara”. Environ. Pollut. 2014, 194, 11–16. [Google Scholar] [CrossRef]
- Serrano, L.; Peñuelas, J. Contribution of Physiological and Morphological Adjustments to Drought Resistance in Two Mediterranean Tree Species. Biol. Plant. 2005, 49, 551–559. [Google Scholar] [CrossRef]
- Watanabe, M.; Agathokleous, E.; Anav, A.; Araminiene, V.; Carrari, E.; De Marco, A.; Hoshika, Y.; Proietti, C.; Sicard, P.; Paoletti, E. Impacts of Ozone on the Ecophysiology of Forest Tree Species. In Tropospheric Ozone—A Hazard for Vegetation and Human Health; Agrawal, S.B., Agrawal, M., Singh, A., Eds.; Cambridge Scholars: Newcastle, UK, 2021; pp. 277–306. [Google Scholar]
- Shang, B.; Xu, Y.; Dai, L.; Yuan, X.; Feng, Z. Elevated Ozone Reduced Leaf Nitrogen Allocation to Photosynthesis in Poplar. Sci. Total Environ. 2019, 657, 169–178. [Google Scholar] [CrossRef]
- Weih, M.; Bonosi, L.; Ghelardini, L.; Rönnberg-Wästljung, A.C. Optimizing Nitrogen Economy under Drought: Increased Leaf Nitrogen Is an Acclimation to Water Stress in Willow (Salix Spp.). Ann. Bot. 2011, 108, 1347–1353. [Google Scholar] [CrossRef] [Green Version]
- Mrak, T.; Štraus, I.; Grebenc, T.; Gričar, J.; Hoshika, Y.; Carriero, G.; Paoletti, E.; Kraigher, H. Different Belowground Responses to Elevated Ozone and Soil Water Deficit in Three European Oak Species (Quercus Ilex, Q. Pubescens and Q. Robur). Sci. Total Environ. 2019, 651, 1310–1320. [Google Scholar] [CrossRef]
- Somers, G.L.; Chappelka, A.H.; Rosseau, P.; Renfro, J.R. Empirical Evidence of Growth Decline Related to Visible Ozone Injury. For. Ecol. Manag. 1998, 104, 129–137. [Google Scholar] [CrossRef]
- Marzuoli, R.; Gerosa, G.; Bussotti, F.; Pollastrini, M. Assessing the Impact of Ozone on Forest Trees in an Integrative Perspective: Are Foliar Visible Symptoms Suitable Predictors for Growth Reduction? A Critical Review. Forests 2019, 10, 1144. [Google Scholar] [CrossRef] [Green Version]
- Chirino, E.; Vilagrosa, A.; Hernández, E.I.; Matos, A.; Vallejo, V.R. Effects of a Deep Container on Morpho-Functional Characteristics and Root Colonization in Quercus Suber L. Seedlings for Reforestation in Mediterranean Climate. For. Ecol. Manag. 2008, 256, 779–785. [Google Scholar] [CrossRef]
- Giovannelli, A.; Traversi, M.L.; Anichini, M.; Hoshika, Y.; Fares, S.; Paoletti, E. Effect of Long-Term vs. Short-Term Ambient Ozone Exposure on Radial Stem Growth, Sap Flux and Xylem Morphology of O3-Sensitive Poplar Trees. Forests 2019, 10, 396. [Google Scholar] [CrossRef] [Green Version]
- Agathokleous, E.; Saitanis, C.J.; Wang, X.; Watanabe, M.; Koike, T. A Review Study on Past 40 Years of Research on Effects of Tropospheric O3 on Belowground Structure, Functioning, and Processes of Trees: A Linkage with Potential Ecological Implications. Water. Air. Soil Pollut. 2016, 227, 33. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Water Regime | O3 Treat. | Onset O3 Injury | Onset Drought Injury | ||||
|---|---|---|---|---|---|---|---|
| POD1 | AOT40 | CDI | |||||
| Q. robur | Q. pubescens | Q. robur | Q. pubescens | Q. robur | Q. pubescens | ||
| WW-treated | AA | 12.07 | Asymp. | 17.78 | Asymp. | Asymp. | Asymp. |
| ×1.2 | 12.40 | 15.69 | 16.41 | 23.77 | Asymp. | Asymp. | |
| ×1.4 | 11.62 | 20.46 | 15.15 | 33.38 | Asymp. | Asymp. | |
| MD-treated | AA | 13.02 | Asymp. | 21.74 | Asymp. | 6.10 | 6.52 |
| ×1.2 | 12.13 | 18.41 | 16.41 | 30.43 | 6.10 | 4.04 | |
| ×1.4 | 13.07 | 20.04 | 21.59 | 33.38 | 4.96 | 6.10 | |
| SD-treated | AA | Asymp. | Asymp. | Asymp. | Asymp. | 10.20 | 10.20 |
| ×1.2 | Asymp. | 13.13 | Asymp. | 30.43 | 10.20 | 10.20 | |
| ×1.4 | 10.72 | 12.81 | 26.23 | 26.23 | 10.20 | 10.20 | |
| Water Regime | O3 Treat./DOY | Q. robur | Q. pubescens | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 209 | 218 | 232 | 240 | 247 | 266 | 209 | 218 | 232 | 240 | 247 | 266 | |||
| O3 VFI incidence | WW-treat. | AA | - | - | 11 | 11 | 11 | 50 | - | - | - | - | - | - |
| ×1.2 | - | 11 | 44 | 44 | 44 | 44 | - | - | - | - | 11 | 11 | ||
| ×1.4 | 22 | 33 | 33 | 44 | 56 | 56 | - | - | - | - | - | 22 | ||
| MD-treat. | AA | - | - | - | 11 | 22 | 44 | - | - | - | - | - | - | |
| ×1.2 | - | 22 | 22 | 22 | 44 | 56 | - | - | - | - | - | 33 | ||
| ×1.4 | - | - | 22 | 33 | 33 | 33 | - | - | - | - | - | 22 | ||
| SD-treat. | AA | - | - | - | - | - | - | - | - | - | - | - | - | |
| ×1.2 | - | - | - | - | - | - | - | - | - | - | - | 22 | ||
| ×1.4 | - | - | - | - | 11 | 11 | - | - | - | - | 11 | 33 | ||
| Drought VFI incidence | WW-treat. | AA | - | - | - | - | - | - | - | - | - | - | - | - |
| ×1.2 | - | - | - | - | - | - | - | - | - | - | - | - | ||
| ×1.4 | - | - | - | - | - | - | - | - | - | - | - | - | ||
| MD-treat. | AA | - | - | 33 | 33 | 50 | 67 | - | - | - | 11 | 11 | 11 | |
| ×1.2 | - | - | 22 | 58 | 67 | 67 | 11 | 11 | 11 | 22 | 33 | 33 | ||
| ×1.4 | - | 44 | 33 | 44 | 50 | 63 | - | - | 22 | 33 | 33 | 33 | ||
| SD-treat. | AA | 22 | 22 | 67 | 78 | 83 | 89 | 22 | 22 | 39 | 67 | 67 | 67 | |
| ×1.2 | 11 | 56 | 61 | 78 | 78 | 89 | 11 | 11 | 22 | 33 | 44 | 44 | ||
| ×1.4 | 33 | 56 | 78 | 78 | 100 | 100 | 44 | 44 | 67 | 78 | 78 | 78 | ||
| Parameters | Model 1 (POD1, CDI) | Model 2 (AOT40, CDI) | ||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| POD1 | p | CDI | p | Intercept | p | R2 | AOT40 | p | CDI | p | Intercept | p | R2 | |||
| Q. robur | Injury | O3 | 6.736 | *** | −0.043 | n.s | −64.967 | *** | 0.717 | 0.002 | *** | −2−027 | *** | −20.600 | *** | 0.738 |
| Drought | 4.802 | *** | 5.882 | *** | −68.059 | *** | 0.887 | 0.001 | *** | 4.524 | *** | −31.350 | *** | 0.861 | ||
| Growth | Height | 0.316 | ** | 0.035 | n.s | −1.197 | n.s | 0.308 | 0.159 | *** | −0.041 | n.s | −0.213 | n.s | 0.501 | |
| Diameter | 0.032 | n.s | −0.051 | ** | 5.182 | *** | 0.323 | 0.019 | n.s | −0.059 | *** | 5.214 | *** | 0.338 | ||
| N. Leaves | 0.068 | n.s | −2.338 | *** | 190.955 | *** | 0.521 | 0.003 | n.s | −2.351 | *** | 191.913 | *** | 0.521 | ||
| Biomass | Leaf | 0.240 | *** | 0.013 | n.s | 1.741 | n.s | 0.404 | 0.068 | * | −0.040 | n.s | 3.743 | *** | 0.191 | |
| Shoot | −0.043 | n.s | −0.176 | *** | 12.517 | *** | 0.404 | −0.017 | n.s | −0.166 | *** | 12.268 | *** | 0.404 | ||
| Root | −0.784 | ** | −0.568 | *** | 40.098 | *** | 0.508 | −0.287 | * | −0.391 | *** | 35.097 | *** | 0.476 | ||
| Total | −0.588 | n.s | −0.732 | *** | 54.357 | *** | 0.428 | −0.236 | n.s | −0.597 | *** | 51.108 | *** | 0.427 | ||
| Ratio R/S | −0.063 | *** | −0.019 | *** | 2.600 | *** | 0.692 | −0.022 | *** | −0.005 | n.s | 2.163 | *** | 0.491 | ||
| Physiology | Pn | −0.629 | *** | −0.283 | *** | 14.579 | *** | 0.824 | 0.000 | *** | −0.152 | ** | 11.930 | *** | 0.738 | |
| PNUE | −0.569 | n.s | −0.307 | * | 14.724 | * | 0.558 | −0.152 | n.s | −0.175 | * | 10.032 | ** | 0.558 | ||
| PPUE | −3.751 | * | −1.560 | ** | 82.525 | ** | 0.916 | −0.668 | n.s | −0.671 | * | 41.067 | * | 0.767 | ||
| Q. pubescens | Injury | O3 | 1.827 | *** | 0.645 | *** | −26.982 | *** | 0.329 | 0.876 | *** | 0.090 | n.s | −16.377 | *** | 0.387 |
| Drought | 1.279 | * | 3.658 | *** | −23.673 | * | 0.787 | 0.660 | * | 3.259 | *** | −17.183 | ** | 0.792 | ||
| Growth | Height | 0.241 | *** | −0.001 | n.s | −0.472 | n.s | 0.373 | 0.115 | *** | −0.060 | * | 1.039 | n.s | 0.371 | |
| Diameter | −0.151 | *** | −0.052 | *** | 8.020 | *** | 0.485 | −0.052 | ** | −0.017 | n.s | 6.595 | *** | 0.244 | ||
| N. Leaves | −4.901 | *** | −1.801 | ** | 175.047 | *** | 0.390 | −2.011 | ** | −0.644 | n.s | 136.376 | *** | 0.283 | ||
| Biomass | Leaf | −0.019 | n.s | −0.012 | n.s | 4.183 | *** | −0.070 | −0.001 | n.s | −0.008 | n.s | 3.865 | *** | −0.075 | |
| Shoot | −0.205 | ** | −0.130 | *** | 13.137 | *** | 0.511 | −0.091 | ** | −0.081 | ** | 11.687 | *** | 0.474 | ||
| Root | −0.054 | n.s | −0.223 | *** | 22.085 | *** | 0.366 | −0.039 | n.s | −0.208 | *** | 22.072 | *** | 0.372 | ||
| Total | −0.278 | n.s | −0.364 | *** | 39.404 | *** | 0.400 | −0.132 | n.s | −0.297 | *** | 37.625 | *** | 0.398 | ||
| Ratio R/S | −0.022 | * | −0.017 | *** | 2.132 | *** | 0.316 | −0.009 | n.s | −0.012 | ** | 1.955 | *** | 0.277 | ||
| Physiology | Pn | −0.370 | *** | −0.255 | *** | 13.500 | *** | 0.795 | 0.000 | *** | −0.179 | *** | 12.290 | *** | 0.771 | |
| PNUE | −0.485 | * | −0.286 | ** | 14.671 | ** | 0.741 | −0.173 | * | −0.172 | ** | 10.622 | *** | 0.757 | ||
| PPUE | −2.385 | * | −1.389 | ** | 70.714 | ** | 0.954 | −0.575 | n.s | −0.788 | ** | 41.724 | ** | 0.901 | ||
| Q. ilex | Injury | O3 | ||||||||||||||
| Drought | ||||||||||||||||
| Growth | Height | 0.127 | n.s | 0.004 | n.s | 15.423 | *** | −0.050 | 0.051 | n.s | −0.026 | n.s | 15.623 | *** | −0.357 | |
| Diameter | 0.153 | ** | 0.031 | * | 0.799 | n.s | 0.305 | 0.042 | ** | −0.003 | n.s | 1.483 | *** | 0.229 | ||
| N. Leaves | 3.238 | ** | 1.194 | ** | 17.246 | n.s | 0.329 | 1.218 | *** | 0.445 | n.s | 23.955 | ** | 0.468 | ||
| Biomass | Leaf | −0.151 | n.s | −0.035 | n.s | 5.825 | *** | 0.022 | −0.029 | n.s | −0.002 | n.s | 4.846 | *** | −0.037 | |
| Shoot | 0.011 | n.s | −0.053 | n.s | 7.619 | *** | 0.158 | −0.000 | n.s | −0.055 | * | 7.750 | *** | 0.157 | ||
| Root | 0.453 | ** | 0.064 | n.s | 7.027 | *** | 0.230 | 0.072 | n.s | −0.031 | n.s | 10.315 | *** | 0.013 | ||
| Total | 0.313 | n.s | 0.024 | n.s | 20.471 | *** | 0.038 | 0.043 | n.s | −0.089 | n.s | 22.911 | *** | −0.005 | ||
| Ratio R/S | 0.095 | * | 0.012 | n.s | 0.158 | n.s | 0.294 | 0.022 | n.s | −0.008 | n.s | 0.7669 | n.s | 0.068 | ||
| Physiology | Pn | 0.094 | n.s | −0.173 | *** | 9.090 | *** | 0.630 | 0.000 | n.s | −0.196 | *** | 9.380 | *** | 0.624 | |
| PNUE | −0.014 | n.s | −0.122 | ** | 5.768 | ** | 0.840 | −0.014 | n.s | −0.118 | *** | 5.939 | *** | 0.846 | ||
| PPUE | −3.036 | n.s | −1.870 | * | 96.073 | * | 0.814 | −0.478 | n.s | −1.124 | * | 71.158 | * | 0.772 | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Moura, B.B.; Paoletti, E.; Badea, O.; Ferrini, F.; Hoshika, Y. Visible Foliar Injury and Ecophysiological Responses to Ozone and Drought in Oak Seedlings. Plants 2022, 11, 1836. https://doi.org/10.3390/plants11141836
Moura BB, Paoletti E, Badea O, Ferrini F, Hoshika Y. Visible Foliar Injury and Ecophysiological Responses to Ozone and Drought in Oak Seedlings. Plants. 2022; 11(14):1836. https://doi.org/10.3390/plants11141836
Chicago/Turabian StyleMoura, Barbara Baesso, Elena Paoletti, Ovidiu Badea, Francesco Ferrini, and Yasutomo Hoshika. 2022. "Visible Foliar Injury and Ecophysiological Responses to Ozone and Drought in Oak Seedlings" Plants 11, no. 14: 1836. https://doi.org/10.3390/plants11141836
APA StyleMoura, B. B., Paoletti, E., Badea, O., Ferrini, F., & Hoshika, Y. (2022). Visible Foliar Injury and Ecophysiological Responses to Ozone and Drought in Oak Seedlings. Plants, 11(14), 1836. https://doi.org/10.3390/plants11141836